Distribution of Ixodes scapularis in Northwestern Ontario: Results from Active and Passive Surveillance Activities in the Northwestern Health Unit Catchment Area

Abstract
1. Introduction
2. Materials and Methods
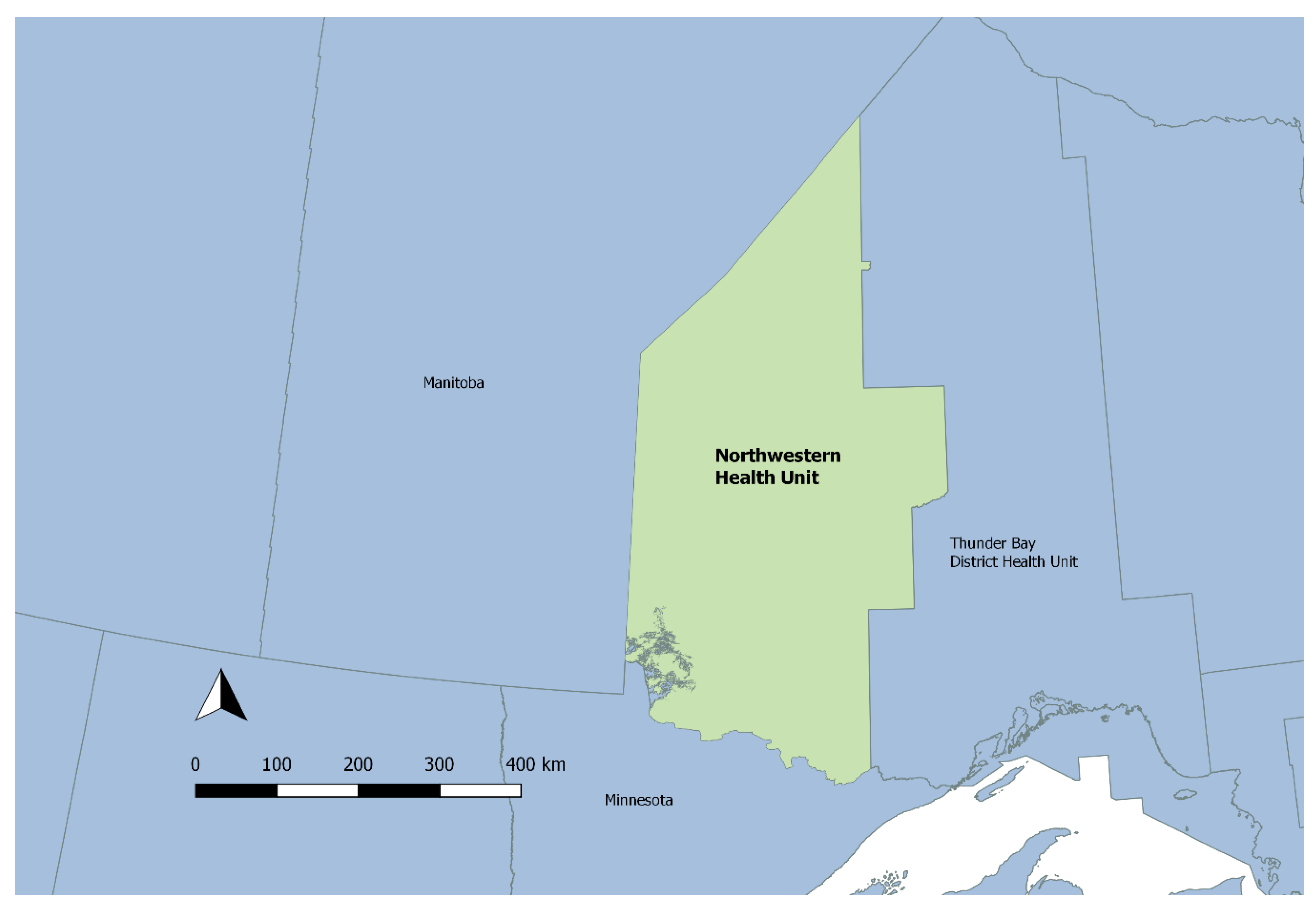
2.1. Study Setting
2.2. Tick Surveillance
2.2.1. Passive Tick Surveillance
2.2.2. Active Tick Surveillance
2.2.3. Pathogen Detection in Blacklegged Ticks
2.2.4. Data Variables and Sources
2.3. Statistical Analysis
3. Results
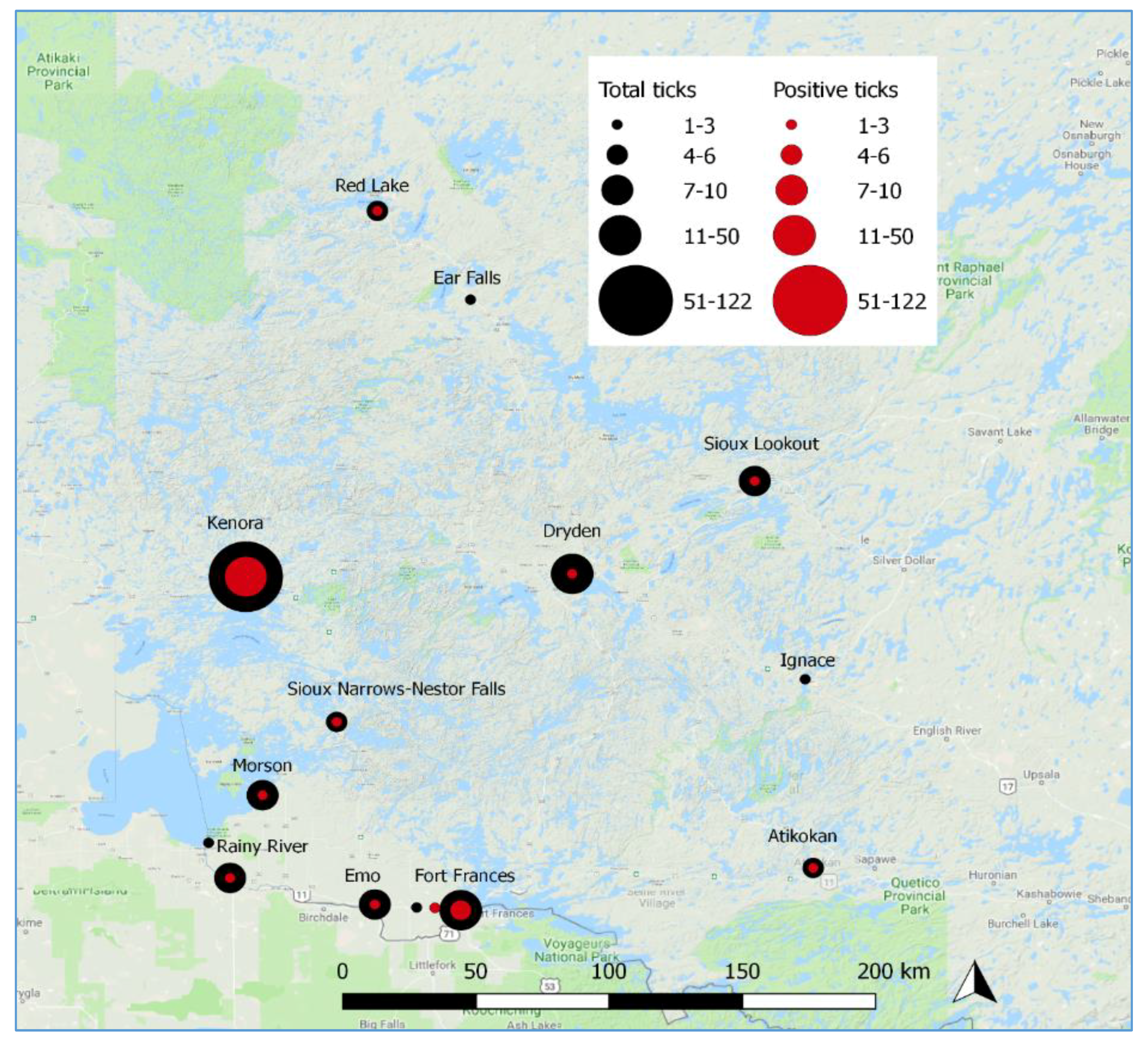
3.1. Passive Tick Surveillance
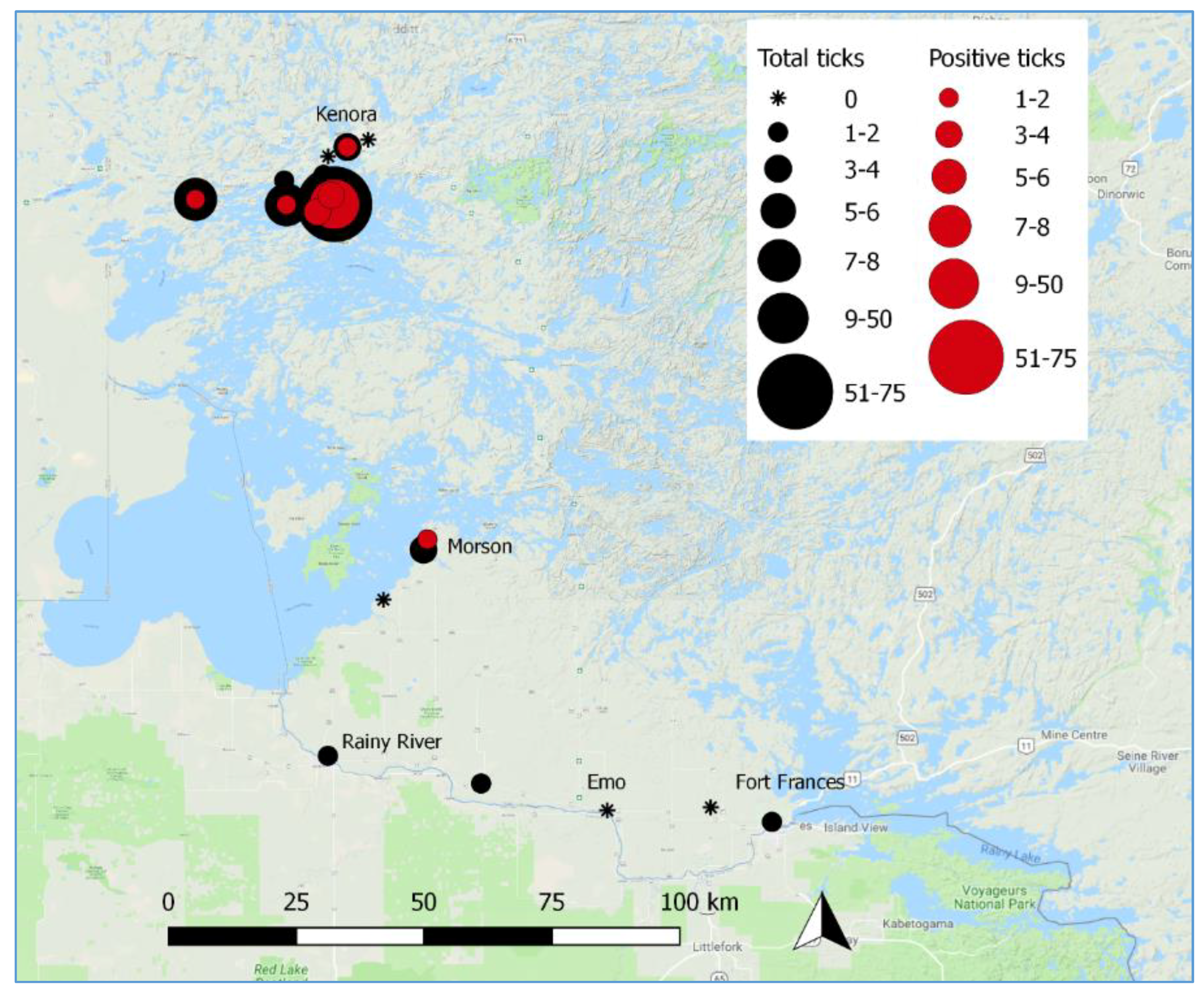
3.2. Active Tick Surveillance
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Average % of Forest Composition | ||
|---|---|---|
| Tree Species | Sites Where I. scapularis Were Detected | Sites Where No I. scapularis Were Detected |
| Trembling Aspen | 37.3% | 43.0% |
| White Birch | 8.8% | 8.0% |
| Bur Oak | 3.3% | 0.0% |
| Black Ash | 11.9% | 10.0% |
| Jack Pine | 2.3% | 4.5% |
| Black Spruce | 5.0% | 15.0% |
| White Pine | 4.6% | 0.0% |
| Balsam Fir | 8.8% | 5.5% |
| Balsam Poplar | 6.3% | 8.0% |
References
- Gasmi, S.; Ogden, N.H.; Lindsay, L.R.; Burns, S.; Fleming, S.; Badcock, J.; Hanan, S.; Gaulin, C.; Leblanc, M.A.; Russell, C.; et al. Surveillance for Lyme disease in Canada: 2009–2015. Can. Commun. Dis. Rep. 2017, 43, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, A.M.; Hinckley, A.F.; Mead, P.S.; Hook, S.A.; Kugeler, K.J. Surveillance for Lyme disease—United States, 2008–2015. MMWR. Surveill. Summ. 2017, 66, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, E.D. Borrelia burgdorferi (Lyme disease). Pediatr. Rev. 2014, 35, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Stanek, G.; Wormser, G.P.; Gray, J.; Strle, F. Lyme borreliosis. Lancet 2012, 379, 461–473. [Google Scholar] [CrossRef]
- Ogden, N.H.; Lindsay, L.R.; Morshed, M.; Sockett, P.N.; Artsob, H. The rising challenge of Lyme borreliosis in Canada. Can. Commun. Dis. Rep. 2008, 34, 1–19. [Google Scholar] [PubMed]
- Ogden, N.H.; Lindsay, L.R.; Morshed, M.; Sockett, P.N.; Artsob, H. The emergence of Lyme disease in Canada. CMAJ 2009, 180, 1221–1224. [Google Scholar] [CrossRef] [PubMed]
- Barker, I.K.; Surgeoner, G.A.; Artsob, H.; McEwen, S.A.; Elliott, L.A.; Campbell, G.D.; Robinson, J.T. Distribution of the Lyme disease vector, Ixodes dammini (Acari: Ixodidae) and isolation of Borrelia burgdorferi in Ontario, Canada. J. Med. Entomol. 1992, 29, 1011–1022. [Google Scholar] [CrossRef] [PubMed]
- Ogden, N.H.; Lindsay, L.R.; Hanincová, K.; Barker, I.K.; Bigras-Poulin, M.; Charron, D.F.; Heagy, A.; Francis, C.M.; O’Callaghan, C.J.; Schwartz, I.; et al. Role of migratory birds in introduction and range expansion of Ixodes scapularis ticks and of Borrelia burgdorferi and Anaplasma phagocytophilum in Canada. Appl. Environ. Microbiol. 2008, 74, 1780–1790. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, L.R.; Barker, I.K.; Surgeoner, G.A.; McEwen, S.A.; Gillespie, T.J.; Robinson, J.T. Survival and development of Ixodes scapularis (Acari: Ixodidae) under various climatic conditions in Ontario, Canada. J. Med. Entomol. 1995, 32, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Madhav, N.K.; Brownstein, J.S.; Tsao, J.I.; Fish, D. A dispersal model for the range expansion of blacklegged tick (Acari: Ixodidae). J. Med. Entomol. 2004, 41, 842–852. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, M.R.; Wilson, M.L. Microclimate-dependent survival of unfed adult Ixodes scapularis (Acari: Ixodidae) in nature: Life cycle and study design implications. J. Med. Entomol. 1996, 33, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Rand, P.W.; Lubelczyk, C.; Lavigne, G.R.; Elias, S.; Holman, M.S.; Lacombe, E.H.; Smith, R.P. Deer density and the abundance of Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 2003, 40, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Ogden, N.H.; Lindsay, L.R.; Beauchamp, G.; Charron, D.; Maarouf, A.; O’Callaghan, C.J.; Waltner-Toews, D.; Barker, I.K. Investigation of relationships between temperature and developmental rates of tick Ixodes scapularis (Acari: Ixodidae) in the laboratory and field. J. Med. Entomol. 2004, 41, 622–633. [Google Scholar] [CrossRef] [PubMed]
- Ogden, N.H.; St.-Onge, L.; Barker, I.K.; Brazeau, S.; Bigras-Poulin, M.; Charron, D.F.; Francis, C.M.; Heagy, A.; Lindsay, L.R.; Maarouf, A.; et al. Risk maps for range expansion of the Lyme disease vector, Ixodes scapularis, in Canada now and with climate change. Int. J. Health Geogr. 2008, 7. [Google Scholar] [CrossRef] [PubMed]
- Ogden, N.H.; Maarouf, A.; Barker, I.K.; Bigras-Poulin, M.; Lindsay, L.R.; Morshed, M.G.; O’Callaghan, C.J.; Ramay, F.; Waltner-Toews, D.; Charron, D.F. Climate change and the potential for range expansion of the Lyme disease vector Ixodes scapularis in Canada. Int. J. Parasitol. 2006, 36, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Leighton, P.A.; Koffi, J.K.; Pelcat, Y.; Lindsay, L.R.; Ogden, N.H. Predicting the speed of tick invasion: An empirical model of range expansion for the Lyme disease vector Ixodes scapularis in Canada. J. Appl. Ecol. 2012, 49, 457–464. [Google Scholar] [CrossRef]
- Klich, M.; Lankester, M.W.; Wu, K.W. Spring migratory birds (Aves) extend the northern occurrence of blacklegged tick (Acari: Ixodidae). J. Med. Entomol. 1996, 33, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.N.; Christensen, C.I.; Scott, J.D. Isolation of Borrelia burgdorferi on mainland Ontario. Can. Commun. Dis. Rep. 1995, 21, 85–86. [Google Scholar] [PubMed]
- Morshed, M.G.; Scott, J.D.; Fernando, K.; Geddes, G.; McNabb, A.; Mak, S.; Durden, L. A distribution and characterization of Borrelia burgdorferi isolates from Ixodes scapularis and presence in mammalian hosts in Ontario, Canada. J. Med. Entomol. 2003, 43, 762–773. [Google Scholar] [CrossRef]
- Scott, J.D.; Foley, J.E.; Clark, K.L.; Anderson, J.F.; Durden, L.A.; Manord, J.M.; Smith, M.L. Established population of blacklegged ticks with high infection prevalence for the Lyme disease bacterium, Borrelia burgdorferi sensu lato, on Corkscrew Island, Kenora District, Ontario. Int. J. Med. Sci. 2016, 13, 881–891. [Google Scholar] [CrossRef] [PubMed]
- Ontario Agency for Health Protection and Promotion (Public Health Ontario). Ontario Lyme Disease Map 2018: Estimated Risk Areas; Queen’s Printer for Ontario: Toronto, ON, Canada, 2018. [Google Scholar]
- Beard, C. Lyme disease prevention and control—The way forward. Can. Commun. Dis. Rep. 2014, 40, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Statistics Canada Northwestern Health Unit, Ontario, Census Profile. Available online: https://www12.statcan.gc.ca/census-recensement/2016/dp-pd/prof/index.cfm?Lang=E (accessed on 8 August 2018).
- Nelder, M.P.; Russell, C.; Lindsay, L.R.; Dhar, B.; Patel, S.N.; Johnson, S.; Moore, S.; Kristjanson, E.; Li, Y.; Ralevski, F. Population-based passive tick surveillance and detection of expanding foci of blacklegged ticks Ixodes scapularis and the Lyme disease agent Borrelia burgdorferi in Ontario, Canada. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Ontario Agency for Health Protection and Promotion (Public Health Ontario). Tick Dragging: Standard Operating Procedure; Queen’s Printer for Ontario: Toronto, ON, Canada, 2015. [Google Scholar]
- Guerra, M.; Walker, E.; Jones, C.; Paskewitz, S.; Roberto Cortinas, M.; Ashley Stancil, L.B.; Bobo, M.; Kitron, U. Predicting the risk of Lyme disease: Habitat suitability for Ixodes scapularis in the north central United States. Emerg. Infect. Dis. 2002, 8, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Dibernardo, A.; Cote, T.; Ogden, N.H.; Lindsay, L.R. The prevalence of Borrelia miyamotoi infection, and co-infections with other Borrelia spp. in Ixodes scapularis ticks collected in Canada. Parasites Vectors 2014, 7, 183. [Google Scholar] [CrossRef] [PubMed]
- Pritt, B.S.; Mead, P.S.; Johnson, D.K.H.; Neitzel, D.F.; Respicio-Kingry, L.B.; Davis, J.P.; Schiffman, E.; Sloan, L.M.; Schriefer, M.E.; Replogle, A.J.; et al. Identification of a novel pathogenic Borrelia species causing Lyme borreliosis with unusually high spirochaetaemia: A descriptive study. Lancet Infect. Dis. 2016, 16, 556–564. [Google Scholar] [CrossRef]
- Courtney, J.W.; Kostelnik, L.M.; Zeidner, N.S.; Massung, R.F. Multiplex real-time PCR for detection of Anaplasma phagocytophilum and Borrelia burgdorferi. J. Clin. Microbiol. 2004, 42, 3164–3168. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, R.; Tsuji, M.; Oda, K.; Zamoto-Niikura, A.; Wei, Q.; Kaswabuchi-Kurata, T.; Nishida, A.; Ishihara, C. Babesia microti-group parasites compared phylogenetically by complete sequencing of the CCTeta gene in 36 isolates. J. Vet. Med. Sci. 2009, 71, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Dupuis, A.P.; Peters, R.J.; Prusinski, M.A.; Falco, R.C.; Ostfeld, R.S.; Kramer, L.D. Isolation of deer tick virus (Powassan virus, lineage II) from Ixodes scapularis and detection of antibody in vertebrate hosts sampled in the Hudson Valley, New York State. Parasites Vectors 2013, 6, 185. [Google Scholar] [CrossRef] [PubMed]
- Ogden, N.; Koffi, J.; Lindsay, L. Assessment of a screening test to identify Lyme disease risk. Can. Commun. Dis. Rep. 2014, 40, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Hamer, S.A.; Roy, P.L.; Hickling, G.J.; Walker, E.D.; Foster, E.S.; Barber, C.C.; Tsao, J.I. Zoonotic pathogens in Ixodes scapularis, Michigan. Emerg. Infect. Dis. 2007, 13, 1131–1133. [Google Scholar] [CrossRef] [PubMed]
- Northwesten Health Unit Tick-Borne Disease & Tick Surveillance. Available online: https://www.nwhu.on.ca/ourservices/healthstatistics/Pages/Tick-Surveillance.aspx (accessed on 15 August 2018).
- Ogden, N.H.; Trudel, L.; Artsob, H.; Barker, I.K.; Beauchamp, G.; Charron, D.F.; Drebot, M.A.; Galloway, T.D.; O’handley, R.; Thompson, R.A.; et al. Ixodes scapularis ticks collected by passive surveillance in Canada: Analysis of geographic distribution and infection with Lyme Borreliosis agent Borrelia burgdorferi. J. Med. Entomol 2006, 43, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, L.R.; Mathison, S.W.; Barker, I.K.; McEwen, S.A.; Surgeoner, G.A. Abundance of Ixodes scapularis (Acari: Ixodidae) larvae and nymphs in relation to host density and habitat on Long Point, Ontario. J. Med. Entomol. 1999, 36, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.J.; Neitzel, D.F.; Moen, R.A.; Craft, M.E.; Hamilton, K.E.; Johnson, L.B.; Mulla, D.J.; Munderloh, U.G.; Redig, P.T.; Smith, K.E.; et al. Disease risk in a dynamic environment: The spread of tick-borne pathogens in Minnesota, USA. Ecohealth 2015, 12, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Aenishaenslin, C.; Bouchard, C.; Koffi, J.K.; Pelcat, Y.; Ogden, N.H. Evidence of rapid changes in Lyme disease awareness in Canada. Ticks Tick. Borne. Dis. 2016, 7, 1067–1074. [Google Scholar] [CrossRef] [PubMed]
- McDermid, J.; Fera, S.; Hogg, A. Climate Change Projections for Ontario: An Updated Synthesis for Policymakers and Planners; Queen’s Printer for Ontario: Peterborough, ON, Canada, 2015; ISBN 9781424921263. [Google Scholar]



| Species | 2013 | 2014 | 2015 | 2016 | 2017 | Total |
|---|---|---|---|---|---|---|
| Dermacentor | ||||||
| D. variabilis | 35 | 39 | 72 | 201 | 265 | 612 |
| D. albipictus | 0 | 0 | 0 | 0 | 1 | 1 |
| Ixodes | ||||||
| I. scapularis | 14 | 39 | 17 | 109 | 72 | 251 |
| I. cookei | 2 | 0 | 1 | 12 | 2 | 17 |
| I. muris | 0 | 1 | 0 | 0 | 1 | 2 |
| Unknown * | 2 | 3 | 0 | 0 | 0 | 5 |
| Amblyomma | ||||||
| A. cajennense | 1 | 0 | 0 | 0 | 0 | 1 |
| Unknown * | 0 | 0 | 4 | 9 | 6 | 19 |
| Year | Total I. scapularis Submissions n | Submission Rate Per 100,000 Population % (95% CI) | I. scapularis Positive for B. burgdorferi % (95% CI) |
|---|---|---|---|
| 2013 | 14 | 17.2 (9.4–28.9) | 21.4 (4.7–50.8) |
| 2014 | 39 | 47.9 (34.1–65.5) | 7.7 (1.6–20.9) |
| 2015 | 17 | 20.9 (12.2–33.5) | 5.9 (0.1–28.7) |
| 2016 | 109 | 133.7 (109.8–161.2) | 15.6 (9.4–23.8) |
| 2017 | 72 | 88.1 (68.9–11.1) | 13.9 (6.9–24.1) |
| Total | 251 | 61.6 (54.2–69.7) | 13.5 (9.6–18.4) |
| Pathogen | Male n = 54 | Female n = 47 |
|---|---|---|
| B. burgdorferi | 37 (68.5%) | 24 (51.1%) |
| B. miyamotoi | 0 (0.0%) | 0 (0.0%) |
| B. microti | 0 (0.0%) | 0 (0.0%) |
| A. phagocytophilum | 5 (9.3%) | 3 (6.4%) |
| Powassan virus | 0 (0.0%) | 1 (2.1%) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schillberg, E.; Lunny, D.; Lindsay, L.R.; Nelder, M.P.; Russell, C.; Mackie, M.; Coats, D.; Berry, A.; Young Hoon, K.N. Distribution of Ixodes scapularis in Northwestern Ontario: Results from Active and Passive Surveillance Activities in the Northwestern Health Unit Catchment Area. Int. J. Environ. Res. Public Health 2018, 15, 2225. https://doi.org/10.3390/ijerph15102225
Schillberg E, Lunny D, Lindsay LR, Nelder MP, Russell C, Mackie M, Coats D, Berry A, Young Hoon KN. Distribution of Ixodes scapularis in Northwestern Ontario: Results from Active and Passive Surveillance Activities in the Northwestern Health Unit Catchment Area. International Journal of Environmental Research and Public Health. 2018; 15(10):2225. https://doi.org/10.3390/ijerph15102225
Chicago/Turabian StyleSchillberg, Erin, Dorian Lunny, L. Robbin Lindsay, Mark P. Nelder, Curtis Russell, Mike Mackie, Dave Coats, Alex Berry, and Kit Ngan Young Hoon. 2018. "Distribution of Ixodes scapularis in Northwestern Ontario: Results from Active and Passive Surveillance Activities in the Northwestern Health Unit Catchment Area" International Journal of Environmental Research and Public Health 15, no. 10: 2225. https://doi.org/10.3390/ijerph15102225
APA StyleSchillberg, E., Lunny, D., Lindsay, L. R., Nelder, M. P., Russell, C., Mackie, M., Coats, D., Berry, A., & Young Hoon, K. N. (2018). Distribution of Ixodes scapularis in Northwestern Ontario: Results from Active and Passive Surveillance Activities in the Northwestern Health Unit Catchment Area. International Journal of Environmental Research and Public Health, 15(10), 2225. https://doi.org/10.3390/ijerph15102225