
Role for Functional SOD2 Polymorphism in Pulmonary Arterial Hypertension in a Chinese Population


Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Patients and Controls
2.3. DNA Extraction and SNP Genotyping
2.4. Construction of SOD2 Expression Plasmid
2.5. Cell Culture and Transient Transfection
2.6. Expression Levels of SOD2 mRNA and Protein
2.7. Statistical Analysis
3. Results
3.1. Characteristics of Study Subjects
3.2. Genotype Frequencies of SOD2 Polymorphism in PAH Cases and Controls
3.3. Correlation with Clinical Features and SOD2 rs4880
3.4. Effect of SOD2 Polymorphism on SOD2 Transcription
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- U.S. National Library of Medicine. Genetics Home. Available online: http://ghr.nlm.nih.gov/condition/pulmonary-arterial-hypertension (accessed on 15 August 2013).
- Thenappan, T.; Shah, S.J.; Rich, S.; Tian, L.; Archer, S.L.; Gomberg-Maitland, M. Survival in pulmonary arterial hypertension: A reappraisal of the NIH risk stratification equation. Eur. Respir. J. 2010, 35, 1079–1087. [Google Scholar] [CrossRef] [PubMed]
- Galie, N.; Humbert, M.; Vachiery, J.L.; Gibbs, S.; Lang, I.; Torbicki, A.; Simonneau, G.; Peacock, A.; Vonk Noordegraaf, A.; Beghetti, M. 2015 ESC/ERS Guidelines for the Diagnosis and Treatment of Pulmonary Hypertension. Eur. Heart J. 2016, 37, 67–119. [Google Scholar] [CrossRef] [PubMed]
- Stacher, E.; Graham, B.B.; Hunt, J.M.; Gandjeva, A.; Groshong, S.D.; McLaughlin, V.V.; Jessup, M.; Grizzle, W.E.; Aldred, M.A.; Cool, C.D.; et al. Modern age pathology of pulmonary arterial hypertension. Am. J. Respir. Crit. Care Med. 2012, 186, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Haase, E.; Bigam, D.L.; Nakonechny, Q.B.; Rayner, D.; Korbutt, G.; Cheung, P.Y. Cardiac function, myocardial glutathione, and matrix metalloproteinase-2 levels in hypoxic newborn pigs reoxygenated by 21%, 50%, or 100% oxygen. Shock 2005, 23, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yang, T.; Xu, X.; Wang, M.; Zhong, L.; Yang, Y.; Zhai, Z.; Xiao, F.; Wang, C. Oxidative stress and nitric oxide signaling related biomarkers in patients with pulmonary hypertension: A case control study. BMC Pulm. Med. 2015, 15, 50. [Google Scholar] [CrossRef] [PubMed]
- Geraci, M.W.; Moore, M.; Gesell, T.; Yeager, M.E.; Alger, L.; Golpon, H.; Gao, B.; Loyd, J.E.; Tuder, R.M.; Voelkel, N.F. Gene expression patterns in the lungs of patients with primary pulmonary hypertension: A gene microarray analysis. Circ. Res. 2001, 88, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Heiss, E.H.; Schachner, D.; Werner, E.R.; Dirsch, V.M. Active NF-E2-related factor (Nrf2) contributes to keep endothelial NO synthase (eNOS) in the coupled state: Role of reactive oxygen species (ROS), eNOS, and heme oxygenase (HO-1) levels. J. Biol. Chem. 2009, 284, 31579–31586. [Google Scholar] [CrossRef] [PubMed]
- Roth, T.L.; Nayak, D.; Atanasijevic, T.; Koretsky, A.P.; Latour, L.L.; McGavern, D.B. Transcranial amelioration of inflammation and cell death after brain injury. Nature 2014, 505, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Birukov, K.G. Cyclic stretch, reactive oxygen species, and vascular remodeling. Antioxid. Redox Signal. 2009, 11, 1651–1667. [Google Scholar] [CrossRef] [PubMed]
- McCord, J.M.; Fridovich, I. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [PubMed]
- Salin, M.L.; McCord, J.M. Free radicals and inflammation. Protection of phagocytosine leukocytes by superoxide dismutase. J. Clin. Investig. 1975, 56, 1319–1323. [Google Scholar] [CrossRef] [PubMed]
- Shuvaev, V.V.; Han, J.; Yu, K.J.; Huang, S.; Hawkins, B.J.; Madesh, M.; Nakada, M.; Muzykantov, V.R. PECAM-targeted delivery of SOD inhibits endothelial inflammatory response. FASEB J. 2011, 25, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, M.A.; Ollosson, R.; Stewart, V.C.; Clark, J.B. Oxidation of nitric oxide by oxo manganese-salen complexes: A new mechanism for cellular protection by superoxide dismutase/catalase mimetics. Biochem. J. 2002, 366, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Robbins, D.; Zhao, Y. Manganese superoxide dismutase in cancer prevention. Antioxid. Redox Signal. 2014, 20, 1628–1645. [Google Scholar] [CrossRef] [PubMed]
- Land, W.; Schneeberger, H.; Schleibner, S.; Illner, W.D.; Abendroth, D.; Rutili, G.; Arfors, K.E.; Messmer, K. The beneficial effect of human recombinant superoxide dismutase on acute and chronic rejection events in recipients of cadaveric renal transplants. Transplantation 1994, 57, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Sutton, A.; Khoury, H.; Prip-Buus, C.; Cepanec, C.; Pessayre, D.; Degoul, F. The Ala16Val genetic dimorphism modulates the import of human manganese superoxide dismutase into rat liver mitochondria. Pharmacogenetics 2003, 13, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Galie, N.; Hoeper, M.M.; Humbert, M.; Torbicki, A.; Vachiery, J.L.; Barbera, J.A.; Beghetti, M.; Corris, P.; Gaine, S.; Gibbs, J.S.; et al. Guidelines for the diagnosis and treatment of pulmonary hypertension. Eur. Respir. J. 2009, 34, 1219–1263. [Google Scholar] [CrossRef] [PubMed]
- Costa, F.; Dornelles, E.; Manica-Cattani, M.F.; Algarve, T.D.; Filho, O.C.S.; Sagrillo, M.R.; Garcia, L.F.; Cruz, I.B. Influence of Val16Ala SOD2 polymorphism on the in vitro effect of clomiphene citrate in oxidative metabolism. Reprod. Biomed. Online 2012, 24, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.M.; Xu, X.; von Lohneysen, K.; Gilmartin, T.J.; Friedman, J.S. SOD2 deficient erythroid cells up-regulate transferrin receptor and down-regulate mitochondrial biogenesis and metabolism. PLoS ONE 2011, 6, e16894. [Google Scholar] [CrossRef] [PubMed]
- Miar, A.; Hevia, D.; Munoz-Cimadevilla, H.; Astudillo, A.; Velasco, J.; Sainz, R.M.; Mayo, J.C. Manganese superoxide dismutase (SOD2/MnSOD)/catalase and SOD2/GPx1 ratios as biomarkers for tumor progression and metastasis in prostate, colon, and lung cancer. Free Radic. Biol. Med. 2015, 85, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Zhou, Y.; Qiao, C.; Ni, T.; Li, Z.; You, Q.; Guo, Q.; Lu, N. Oroxylin A inhibits glycolysis-dependent proliferation of human breast cancer via promoting SIRT3-mediated SOD2 transcription and HIF1alpha destabilization. Cell Death Dis. 2015, 6, e1714. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Narasimhan, P.; Lee, Y.S.; Song, Y.S.; Endo, H.; Yu, F.; Chan, P.H. Mild hypoxia promotes survival and proliferation of SOD2-deficient astrocytes via c-Myc activation. J. Neurosci. 2006, 26, 4329–4337. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; He, Q.; Ding, X.; Zhao, T.; Zhao, L.; Wang, A. SOD2 is a C-myc target gene that promotes the migration and invasion of tongue squamous cell carcinoma involving cancer stem-like cells. Int. J. Biochem. Cell Biol. 2015, 60, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Drane, P.; Bravard, A.; Bouvard, V.; May, E. Reciprocal down-regulation of p53 and SOD2 gene expression-implication in p53 mediated apoptosis. Oncogene 2001, 20, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Kokoszka, J.E.; Coskun, P.; Esposito, L.A.; Wallace, D.C. Increased mitochondrial oxidative stress in the SOD2 (+/−) mouse results in the age-related decline of mitochondrial function culminating in increased apoptosis. Proc. Natl. Acad. Sci. USA 2001, 98, 2278–2283. [Google Scholar] [CrossRef] [PubMed]
- Voelkel, N.F.; Gomez-Arroyo, J. The role of vascular endothelial growth factor in pulmonary arterial hypertension. The angiogenesis paradox. Am. J. Respir. Cell Mol. Biol. 2014, 51, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Bowers, R.; Cool, C.; Murphy, R.C.; Tuder, R.M.; Hopken, M.W.; Flores, S.C.; Voelkel, N.F. Oxidative stress in severe pulmonary hypertension. Am. J. Respir. Crit. Care Med. 2004, 169, 764–769. [Google Scholar] [CrossRef]
- Cyr, A.R.; Hitchler, M.J.; Domann, F.E. Regulation of SOD2 in cancer by histone modifications and CpG methylation: Closing the loop between redox biology and epigenetics. Antioxid. Redox Signal. 2013, 18, 1946–1955. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, H.; Watanabe, T.; Hayashi, H.; Suzuki, Y.; Nakamura, T.; Ito, S.; Ono, M.; Hoshikawa, Y.; Okada, Y.; Kondo, T.; et al. Angiogenesis inhibitor vasohibin-1 enhances stress resistance of endothelial cells via induction of SOD2 and SIRT1. PLoS ONE 2012, 7, e46459. [Google Scholar] [CrossRef] [PubMed]
- Orlova, V.V.; Liu, Z.; Goumans, M.J.; ten Dijke, P. Controlling angiogenesis by two unique TGF-beta type I receptors signaling pathways. Histol. Histopathol. 2011, 26, 1219–1230. [Google Scholar]
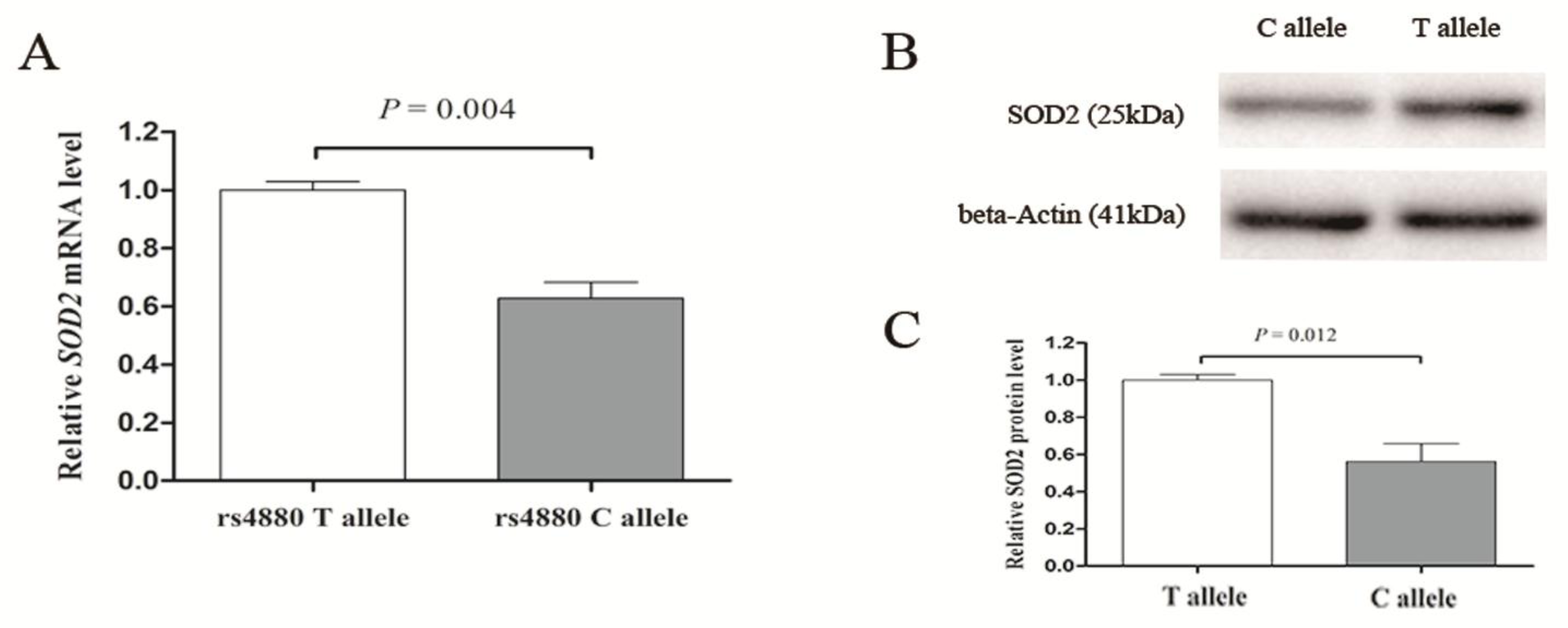

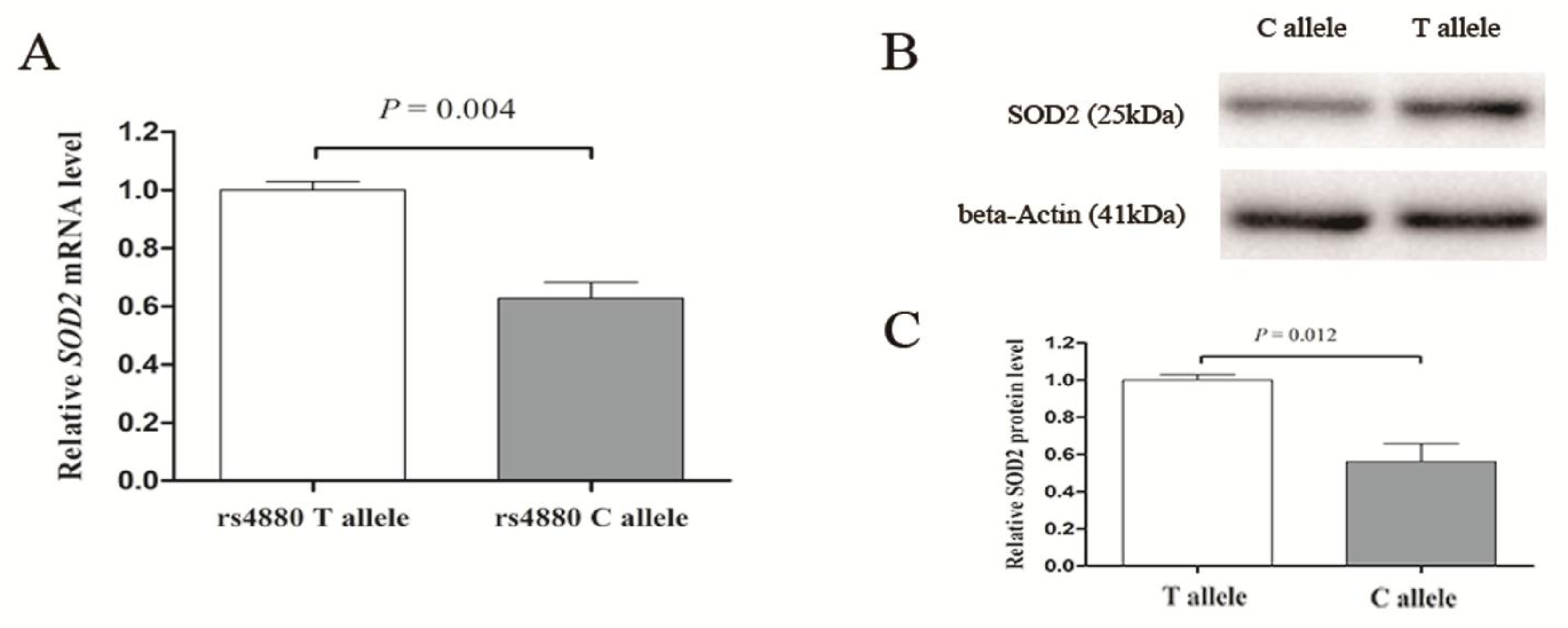{kind=link}
| Variables | Case (n = 460) | Control (n = 530) | p |
|---|---|---|---|
| age (years) | 44.6 ± 13.9 | 45.4 ± 13.6 | 0.861 |
| sex | 0.853 | ||
| male | 135 (29.3%) | 163 (30.8%) | |
| female | 325 (70.7%) | 367 (69.2%) | |
| BMI (kg/m2) | 25.6 ± 4.5 | 25.1 ± 5.3 | 0.399 |
| 6MWD (m) | 247.9 (146.2–363.3) | ||
| NT-proBNP (pg/mL) | 1139.9 (360.9–2471.8) | ||
| mPAP (mmHg) | 51 (40–59) | ||
| PVRI (dyn*sec*m2/cm5) | 1485.2 (1137.3–2742.7) | ||
| CO (L/min) | 3.86 ± 1.81 | ||
| CI (L/min/m2) | 2.23 ± 0.88 |
| rs4880 T > C | Cases, n (%) | Controls, n (%) | OR (95% CI) a | p a |
|---|---|---|---|---|
| TT | 319 (69.35%) | 426 (80.37%) | Reference | - |
| TC | 111 (24.13%) | 99 (18.68%) | 1.35 (0.79–2.81) | 0.195 |
| CC | 30 (6.52%) | 5 (0.94%) | 3.81 (1.22–5.67) | 0.013 |
| TC + CC | 141 (30.65%) | 104 (19.62%) | 2.75 (1.23–3.98) | 0.010 |
| p trend | 0.023 |
| Variables | Case (n = 460) | p Value a | |
|---|---|---|---|
| TT (n = 319) | TC + CC (n = 141) | ||
| 6MWD (m) | 258.7 ± 49.6 | 239.2 ± 41.3 | 0.001 |
| NT-proBNP (pg/mL) | 1075.7 ± 256.5 | 1177.8 ± 243.4 | <0.001 |
| mPAP (mmHg) | 50 ± 5 | 52 ± 7 | <0.001 |
| PVRI (dyn*sec*m2/cm5) | 1461.6 ± 252.9 | 1516.0 ± 274.2 | 0.063 b |
| CO (L/min) | 4.01 ± 1.84 | 3.58 ± 1.73 | 0.021 |
| CI (L/min/m2) | 2.36 ± 0.79 | 2.18 ± 0.91 | 0.035 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, M.; Xu, M.; Han, L.; Yuan, C.; Mei, Y.; Zhang, H.; Chen, S.; Sun, K.; Zhu, B. Role for Functional SOD2 Polymorphism in Pulmonary Arterial Hypertension in a Chinese Population. Int. J. Environ. Res. Public Health 2017, 14, 266. https://doi.org/10.3390/ijerph14030266
Xu M, Xu M, Han L, Yuan C, Mei Y, Zhang H, Chen S, Sun K, Zhu B. Role for Functional SOD2 Polymorphism in Pulmonary Arterial Hypertension in a Chinese Population. International Journal of Environmental Research and Public Health. 2017; 14(3):266. https://doi.org/10.3390/ijerph14030266
Chicago/Turabian StyleXu, Ming, Min Xu, Lei Han, Chao Yuan, Yong Mei, Hengdong Zhang, Shi Chen, Kai Sun, and Baoli Zhu. 2017. "Role for Functional SOD2 Polymorphism in Pulmonary Arterial Hypertension in a Chinese Population" International Journal of Environmental Research and Public Health 14, no. 3: 266. https://doi.org/10.3390/ijerph14030266
APA StyleXu, M., Xu, M., Han, L., Yuan, C., Mei, Y., Zhang, H., Chen, S., Sun, K., & Zhu, B. (2017). Role for Functional SOD2 Polymorphism in Pulmonary Arterial Hypertension in a Chinese Population. International Journal of Environmental Research and Public Health, 14(3), 266. https://doi.org/10.3390/ijerph14030266