
Meta-Prediction of the Effect of Methylenetetrahydrofolate Reductase Polymorphisms and Air Pollution on Alzheimer’s Disease Risk



Abstract
:1. Introduction
2. Materials and Methods
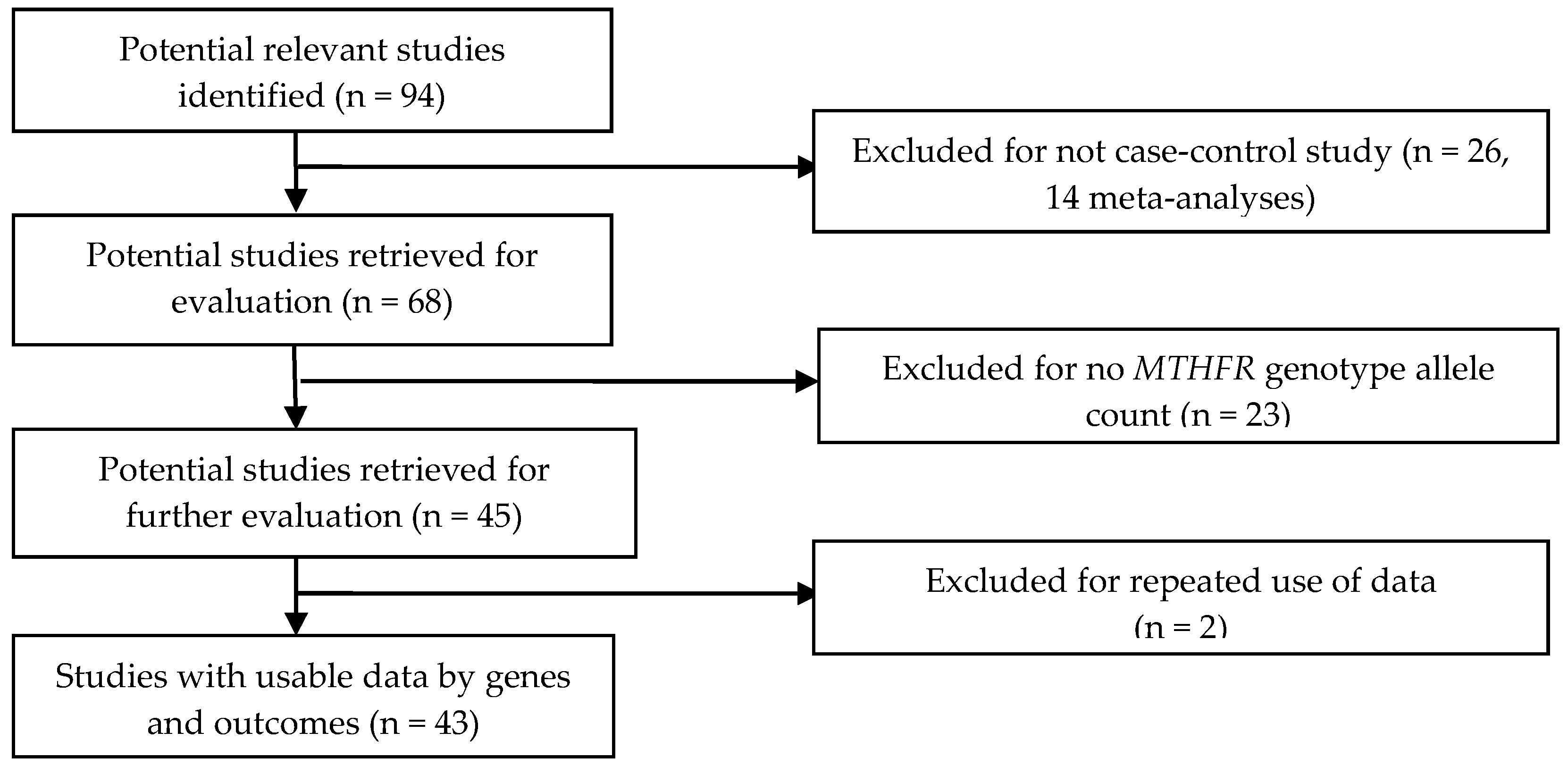
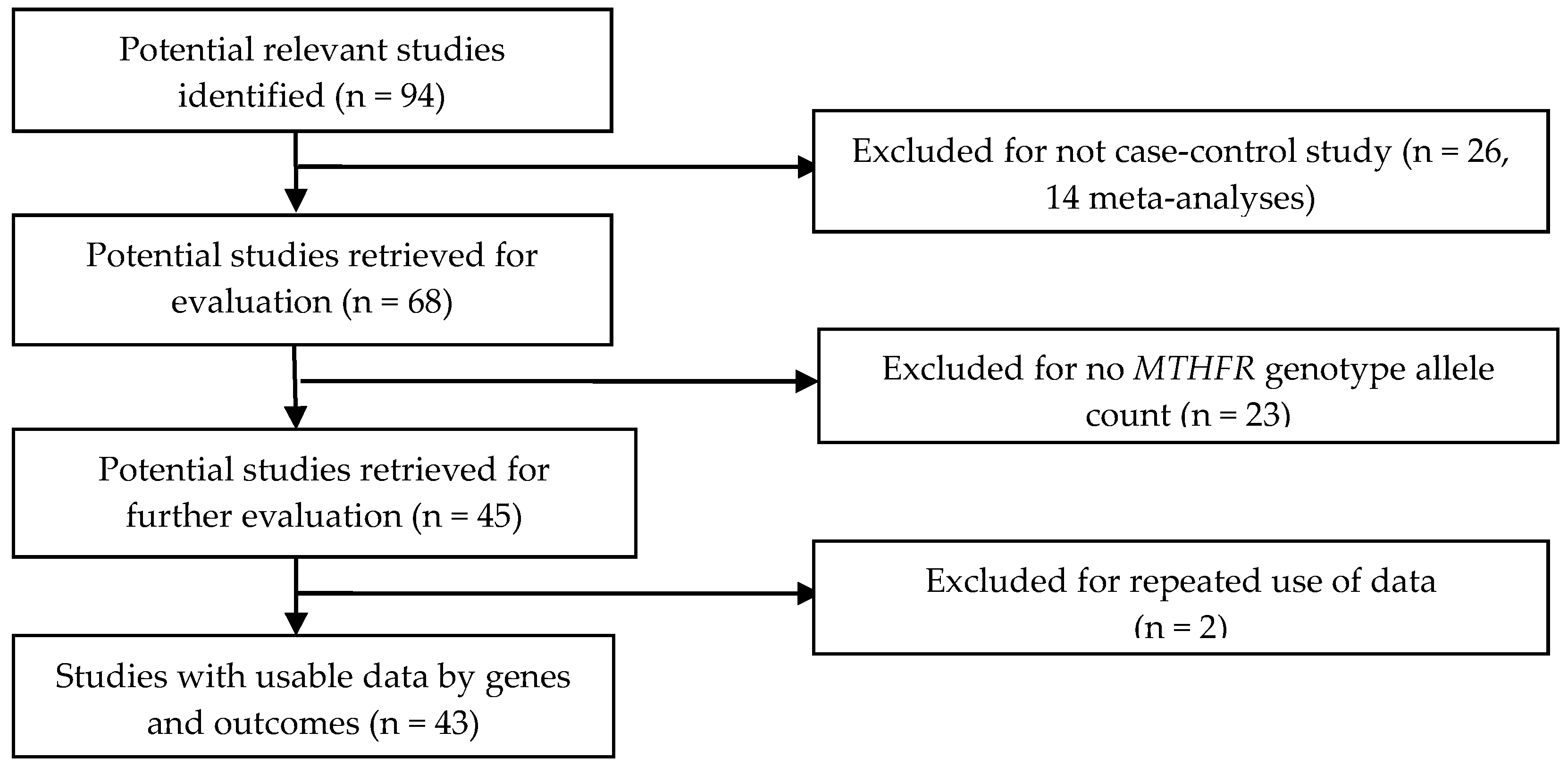
2.1. Selection Criteria and Identification of Studies
2.2. Data Extraction
2.3. Quality Assessment
2.4. Data Synthesis and Analysis
3. Results
3.1. Meta-Analysis
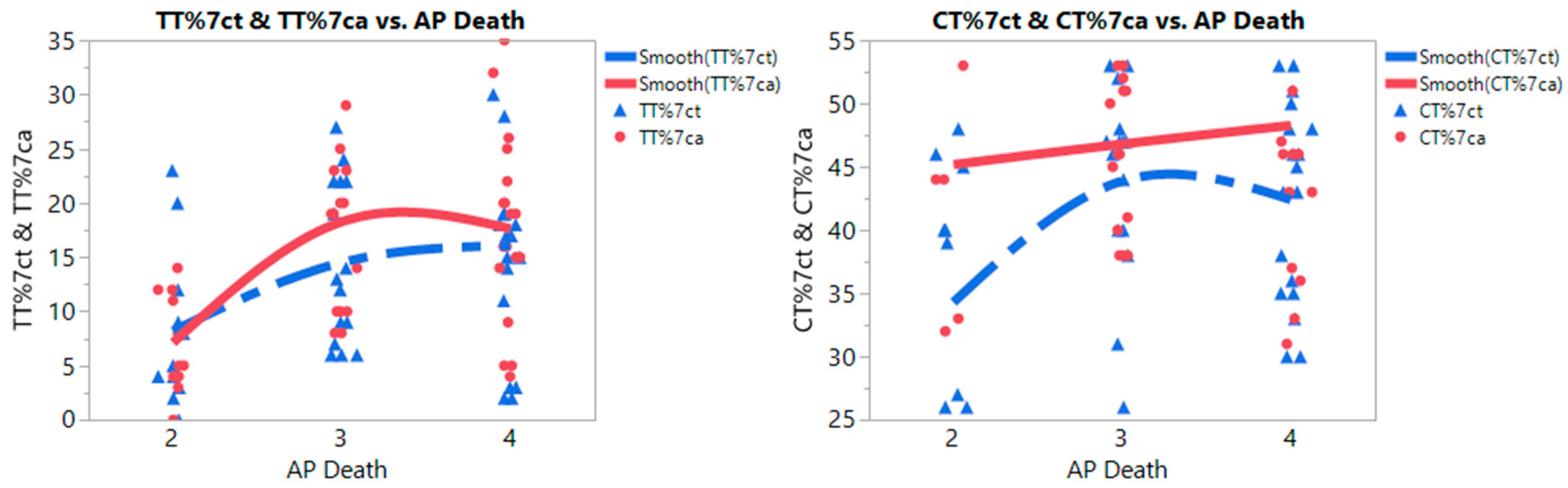
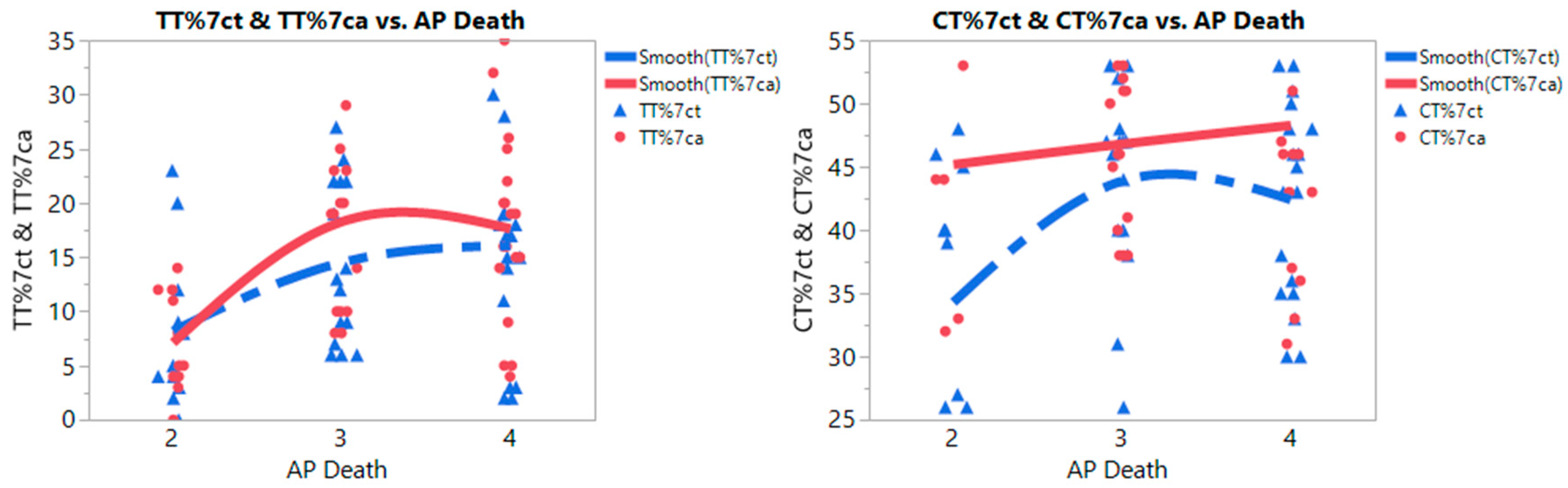
3.2. Meta-Prediction
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Alzheimer’s Association. 2016 Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2016, 12, 459–509. [Google Scholar]
- Alzheimer’s Association. 2015 Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2015, 11, 332–384. [Google Scholar]
- Hurd, M.D.; Martorell, P.; Delavande, A.; Mullen, K.J.; Langa, K.M. Monetary costs of dementia in the United States. N. Engl. J. Med. 2013, 368, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
- Friedman, E.M.; Shih, R.A.; Langa, K.M.; Hurd, M.D. US prevalence and predictors of informal caregiving for dementia. Health Aff. 2015, 34, 1637–1641. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Yu, J.T.; Tian, Y.; Tan, L. Epidemiology and etiology of Alzheimer’s disease: From genetic to non-genetic factors. Curr. Alzheimer Res. 2013, 10, 852–867. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, K.; Grundke-Iqbal, I. Alzheimer’s disease, a multifactorial disorder seeking multitherapies. Alzheimer’s Dement. 2010, 6, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Divyakolu, S.; Tejaswini, Y.; Thomas, W.; Thumoju, S.; Sreekanth, V.R.; Vasavi, M.; OmSai, V.R.; Nagaratna, V.; Hasan, Q.; Ahuja, Y.R. Evaluation of C677T polymorphism of the methylenetetrahydrofolate reductase (MTHFR) gene in various neurological disorders. J. Neurol. Disord. 2014, 2014, 142. [Google Scholar] [CrossRef]
- Kristin, E.D.; Bearden, D.; Watkins, D.; Hyland, K.; Rosenblatt, D.S.; Ficicioglu, C. Severe 5, 10-methylenetetrahydrofolate reductase deficiency and two MTHFR variants in an adolescent with progressive myoclonic epilepsy. Pediatr. Neurol. 2014, 51, 266–270. [Google Scholar]
- Bialecka, M.; Drozdzik, M.; Kurzawski, M.; Sitek, E.; Golab-Janowska, M.; Honczarenko, K.; Mak, M.; Robowski, P.; Roszman, A.; Slawek, J. COMT, MTHFR and SLC19A1 (RFC-1) Polymorphisms, Homocysteine Blood Levels and Cognitive Impairment in Parkinson’s Disease (Po4.175). Neurology 2013, 80, 104–175. [Google Scholar]
- Vijayan, M.; Chinniah, R.; Ravi, P.M.; Sivanadham, R.; Joseph, A.K.; Vellaiappan, N.A.; Krishnan, J.I.; Karuppiah, B. MTHFR (C677T) CT genotype and CT-apoE3/3 genotypic combination predisposes the risk of ischemic stroke. Gene 2016, 591, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, J.; Zhao, L.; Liu, X.; Mi, W. Methylenetetrahydrofolate reductase polymorphisms and risk of acute lymphoblastic leukemia-evidence from an updated meta-analysis including 35 studies. BMC Med. Genet. 2012, 13, 77. [Google Scholar] [CrossRef] [PubMed]
- Gaughan, D.J.; Barbaux, S.; Kluijtmans, L.A.; Whitehead, A.S. The human and mouse methylenetetrahydrofolate reductase (MTHFR) genes: Genomic organization, mRNA structure and linkage to the CLCN6 gene. Gene 2000, 257, 279–289. [Google Scholar] [CrossRef]
- Peng, Q.; Lao, X.; Huang, X.; Qin, X.; Li, S.; Zeng, Z. The MTHFR C677T polymorphism contributes to increased risk of Alzheimer’s disease: Evidence based on 40 case-control studies. Neurosci. Lett. 2015, 586, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Crider, K.S.; Yang, T.P.; Berry, R.J.; Bailey, L.B. Folate and DNA methylation: A review of molecular mechanisms and the evidence for folate’s role. Adv. Nutr. 2012, 3, 21–38. [Google Scholar] [CrossRef] [PubMed]
- Crider, K.S.; Zhu, J.H.; Hao, L.; Yang, Q.H.; Yang, T.P.; Gindler, J.; Berry, R. J. MTHFR 677C →T genotype is associated with folate and homocysteine concentrations in a large, population-based, double-blind trial of folic acid supplementation. Am. J. Clin. Nutr. 2011, 93, 1365–1372. [Google Scholar] [CrossRef] [PubMed]
- Myers, V.; Broday, D.M.; Steinberg, D.M.; Drory, Y.; Gerber, Y. Exposure to particulate air pollution and long-term incidence of frailty after myocardial infarction. Ann. Epidemiol. 2013, 23, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Wellenius, G.A.; Burger, M.R.; Coull, B.A.; Schwartz, J.; Suh, H.H.; Koutrakis, P.; Mittleman, M.A. Ambient air pollution and the risk of acute ischemic stroke. Arch. Intern. Med. 2012, 172, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Shiao, S.; Yu, C. Meta-prediction of MTHFR gene polymorphism mutations and associated risk for colorectal cancer. Biol. Res. Nurs. 2016, 18, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Park, S.K.; Vokonas, P.S.; Sparrow, D.; Wilker, E.; Baccarelli, A.; Schwartz, J. Air pollution and homocysteine: More evidence that oxidative stress-related genes modify effects of particulate air pollution. Epidemiology 2010, 21, 198. [Google Scholar] [CrossRef] [PubMed]
- Killin, L.O.; Starr, J.M.; Shiue, I.J.; Russ, T.C. Environmental risk factors for dementia: A systematic review. BMC Geriatr. 2016, 16, 175. [Google Scholar] [CrossRef] [PubMed]
- Moulton, P.V.; Yang, W. Air pollution, oxidative stress, and Alzheimer’s disease. J. Environ. Public Health 2012, 2012, 472751. [Google Scholar] [CrossRef] [PubMed]
- Weuve, J.; Puett, R.C.; Schwartz, J.; Yanosky, J.D.; Laden, F.; Grodstein, F. Exposure to particulate air pollution and cognitive decline in older women. Arch. Intern. Med. 2012, 172, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Rai, V. Methylenetetrahydrofolate reductase (MTHFR) C677T polymorphism and Alzheimer disease risk: A meta-analysis. Mol. Neurobiol. 2016, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Zhao, H.; Kong, Y.; Ye, M. Association between the MTHFR gene and Alzheimer’s disease: A meta-analysis. Int. J. Neurosci. 2011, 121, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.Y.; Miao, L.; Li, Y.S.; Hu, G.Y. Meta-analysis of the methylenetetrahydrofolate reductase C677T polymorphism and susceptibility to Alzheimer’s disease. Neurosci. Res. 2010, 68, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yang, M.; Li, G.M.; Qiu, Y.; Zheng, J.; Du, X.; Liu, R.W. The MTHFR C677T polymorphism contributes to an increased risk for vascular dementia: A meta-analysis. J. Neurol. Sci. 2010, 294, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Riley, R.D.; Higgins, J.P.; Deeks, J.J. Interpretation of random effects meta-analyses. BMJ 2011, 342, d549. [Google Scholar] [CrossRef] [PubMed]
- Faraway, J.J. Extending the Linear Model with R: Generalized Linear, Mixed Effects and Nonparametric Regression Models; CRC Press: Boca Raton, FL, USA, 2016; Volume 124. [Google Scholar]
- Stroup, D.F.; Berlin, J.A.; Morton, S.C.; Olkin, I.; Williamson, G.D.; Rennie, D.; Thacker, S.B. Meta-analysis of observational studies in epidemiology: A proposal for reporting. JAMA 2000, 283, 2008–2012. [Google Scholar] [CrossRef] [PubMed]
- Skrobot, O.A.; McKnight, A.J.; Passmore, P.A.; Seripa, D.; Mecocci, P.; Panza, F.; Karhunen, P. A validation study of vascular cognitive impairment genetics meta-analysis findings in an independent collaborative cohort. J. Alzheimer’s Dis. 2016, 53, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Teng, W.; Li, J.; Hao, F.; Wang, N. Homocysteine and Alzheimer’s disease: Evidence for a causal link from mendelian randomization. J. Alzheimer’s Dis. 2016, 52, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.G.; Ai, Q.L.; Wang, W.M.; Xiao, Z.C. Meta-analysis supports association of a functional SNP (rs1801133) in the MTHFR gene with Parkinson’s disease. Gene 2013, 531, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Grarup, N.; Sulem, P.; Sandholt, C.H.; Thorleifsson, G.; Ahluwalia, T.S.; Steinthorsdottir, V.; Albrechtsen, A. Genetic Architecture of Vitamin B12 and Folate Levels Uncovered Applying Deeply Sequenced Large Datasets. PLoS Genet. 2013, 9, e1003530. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, R.; Skrobot, O.A.; Dwyer, J.; Munafo, M.; Kehoe, P.G. Using Alzgene-like approaches to investigate susceptibility genes for vascular cognitive impairment. J. Alzheimer’s Dis. 2013, 34, 145–154. [Google Scholar]
- Nakamizo, T.; Nagayama, M. Stroke and the genetics of hyperhomocysteinemia. Brain Nerve Shinkei Kenkyu No Shinpo 2008, 60, 1295–1306. [Google Scholar] [PubMed]
- Bertram, L.; McQueen, M.B.; Mullin, K.; Blacker, D.; Tanzi, R.E. Systematic meta-analyses of Alzheimer disease genetic association studies: The AlzGene database. Nat. Genet. 2007, 39, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Zoccolella, S.; Martino, D.; Defazio, G.; Lamberti, P.; Livrea, P. Hyperhomocysteinemia in movement disorders: Current evidence and hypotheses. Curr. Vasc. Pharmacol. 2006, 4, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, M.; Kato, Y.; Hashimoto, M.; Yukawa, S.; Omori, K. Apolipoprotein E, Methylenetetrahydrofolate reductase (MTHFR) mutation and the risk of senile dementia: An epidemiological study using the polymerase chain reaction (PCR) method. J. Epidemiol. 2000, 10, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Bi, S.; Pang, S.H.; Zhang, Y.; Wu, J. Systematic meta-analyses of Alzheimer disease genetic association studies: The AlzGene database. Chin. J. Rehabil. Med. 2004, 19, 40–42. [Google Scholar]
- Seripa, D.; Dal Forno, G.; Matera, M.G.; Gravina, C.; Margaglione, M.; Palermo, M.T.; Masullo, C. Methylenetetrahydrofolate reductase and angiotensin converting enzyme gene polymorphisms in two genetically and diagnostically distinct cohort of Alzheimer patients. Neurobiol. Aging 2003, 24, 933–939. [Google Scholar] [CrossRef]
- Yoo, J.-H.; Choi, G.-D.; Kang, S.-S. Pathogenicity of thermolabile methylenetetrahydrofolate reductase for vascular dementia. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1921–1925. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Cook, D.J.; Eastwood, S.; Olkin, I.; Rennie, D.; Stroup, D.F. Improving the quality of reports of meta-analyses of randomised controlled trials: The QUOROM statement. Lancet 1999, 354, 1896–1900. [Google Scholar] [CrossRef]
- Downs, S.H.; Black, N. The feasibility of creating a checklist for the assessment of the methodological quality both of randomised and non-randomised studies of health care interventions. J. Epidemiol. Community Health 1998, 52, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Zintzaras, E.; Ioannidis, J. Heterogeneity testing in meta-analysis of genome searches. Genet. Epidemiol. 2005, 28, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.; Thompson, S.G.; Deeks, J.J.; Altman, D.G. Measuring inconsistency in meta-analyses. BMJ 2003, 327, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Pereira, T.V.; Rudnicki, M.; Cheung, B.M.; Baum, L.; Yamada, Y.; Oliveira, P.S.; Krieger, J.E. Three endothelial nitric oxide (NOS3) gene polymorphisms in hypertensive and normotensive individuals: Meta-analysis of 53 studies reveals evidence of publication bias. J. Hypertens. 2007, 25, 1763–1774. [Google Scholar] [CrossRef] [PubMed]
- Scott, I. Interpreting risks and ratios in therapy trials. Aust. Prescr. 2008, 31, 12–16. [Google Scholar] [CrossRef]
- Deeks, J.; Higgins, J.; Altman, D. Analysing Data and Undertaking Meta-Analyses: Cochrane Handbook for Systematic Reviews of Interventions Version 5.1. 0 [updated March 2011]; Cochrane Handbook for Systematic Reviews of Interventions Version; Cochrane: London, UK, 2011; Chapter 9; Volume 5. [Google Scholar]
- Albrecht, J. Key Concepts and Techniques in GIS; Sage: Thousand Oaks, CA, USA, 2007. [Google Scholar]
- World Health Organization. Global Health Risks. Available online: https://commons.wikimedia.org/wiki/File:Deaths_from_air_pollution.png 2012 (accessed on 12 October 2016).
- Teng, Z.; Wang, L.; Cai, S.; Yu, P.; Wang, J.; Gong, J.; Liu, Y. The 677C> T (rs1801133) polymorphism in the MTHFR gene contributes to lorectal cancer risk: A meta-analysis based on 71 research studies. PLoS ONE 2013, 8, e55332. [Google Scholar] [CrossRef] [PubMed]
- Duval, S.; Tweedie, R. Trim and fill: A simple funnel-plot-based method of testing and adjusting for publication bias in meta-analysis. Biometrics 2000, 56, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Egger, M.; Smith, G.D.; Schneider, M.; Minder, C. Bias in meta-analysis detected by a simple, graphical test. BMJ 1997, 315, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Thornton, A.; Lee, P. Publication bias in meta-analysis: Its causes and consequences. J. Clin. Epidemiol. 2000, 53, 207–216. [Google Scholar] [CrossRef]
- Akaike, H. Akaike’s Information Criterion. In International Encyclopedia of Statistical Science; Springer: Berlin/Heidelberg, Germany, 2011; p. 25. [Google Scholar]
- Jaccard, J.; Becker, M.A.; Wood, G. Pairwise multiple comparison procedures: A review. Psychol. Bull. 1984, 96, 589. [Google Scholar] [CrossRef]
- Kang, S.S.; Zhou, J.; Wong, P.W.; Kowalisyn, J.; Strokosch, G. Intermediate homocysteinemia: A thermolabile variant of methylenetetrahydrofolate reductase. Am. J. Hum. Genet. 1988, 43, 414. [Google Scholar] [PubMed]
- Block, M.L.; Calderón-Garcidueñas, L. Air pollution: Mechanisms of neuroinflammation and CNS disease. Trends Neurosci. 2009, 32, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Levesque, S.; Surace, M.J.; McDonald, J.; Block, M.L. Air pollution & the brain: Subchronic diesel exhaust exposure causes neuroinflammation and elevates early markers of neurodegenerative disease. J. Neuroinflamm. 2011, 8, 105. [Google Scholar]
- Genc, S.; Zadeoglulari, Z.; Fuss, S.H.; Genc, K. The adverse effects of air pollution on the nervous system. J. Toxicol. 2012, 2012, 782462. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Genotype (Number of Studies) | AD Case N = 4732 n (%) | Control N = 5979 n (%) | Test of Heterogeneity | Statistical Model | Test of Association | |||
|---|---|---|---|---|---|---|---|---|
| Q | p | I2 (%) | Risk Ratio (95% CI) | p | ||||
| TT (44) | 822 (17.37) | 925 (15.47) | 58.28 | 0.0600 | 26.2 | Fixed | 1.13 (1.04, 1.23) | 0.0047 |
| Caucasian (13) | 268 (15.76) | 219 (14.53) | 10.00 | 0.6158 | 0 | Fixed | 1.04 (0.88, 1.22) | 0.6605 |
| East Asian (20) | 486 (21.39) | 633 (18.39) | 29.15 | 0.0636 | 34.8 | Fixed | 1.19 (1.07, 1.32) | 0.0015 |
| South Asian (4) | 15 (5.08) | 14 (3.18) | 4.24 | 0.2364 | 29.3 | Fixed | 1.36 (0.66, 2.81) | 0.4045 |
| Mixed (2) | 9 (7.20) | 15 (7.11) | 2.56 | 0.1098 | 60.9 | Fixed | 1.01 (0.43, 2.33) | 0.9906 |
| Middle Eastern (3) | 35 (13.57) | 33 (13.36) | 5.84 | 0.0539 | 65.8 | Fixed | 0.99 (0.64, 1.53) | 0.9593 |
| African (2) | 9 (11.11) | 11 (8.33) | 3.34 | 0.0678 | 70.0 | Fixed | 1.14 (0.41, 3.21) | 0.8016 |
| CT (44) | 2246 (47.46) | 2607 (43.60) | 67.85 | 0.0092 | 36.6 | Random | 1.12 (1.06, 1.19) | <0.0001 |
| Caucasian (13) | 803 (47.21) | 659 (43.73) | 6.33 | 0.8986 | 0 | Fixed | 1.07 (0.99, 1.15) | 0.0864 |
| East Asian (20) | 1089 (47.93) | 1604 (46.60) | 32.15 | 0.0301 | 40.9 | Random | 1.08 (1.00, 1.17) | 0.0638 |
| South Asian (4) | 118 (40.00) | 108 (24.55) | 5.74 | 0.1249 | 47.7 | Fixed | 1.56 (1.25, 1.95) | <0.0001 |
| Mixed (2) | 71 (56.80) | 101 (47.87) | 1.46 | 0.2263 | 31.7 | Fixed | 1.21 (0.98, 1.48) | 0.0704 |
| Middle Eastern (3) | 124 (48.06) | 86 (34.82) | 1.32 | 0.5156 | 0 | Fixed | 1.35 (1.09, 1.66) | 0.0052 |
| African (2) | 41 (50.62) | 49 (37.12) | 0.47 | 0.4920 | 0 | Fixed | 1.50 (1.10, 2.04) | 0.0107 |
| CC (44) | 1664 (35.16) | 2447 (40.93) | 61.63 | 0.0325 | 30.2 | Random | 0.85 (0.80, 0.90) | <0.0001 |
| Caucasian (13) | 630 (37.04) | 629 (41.74) | 11.43 | 0.4926 | 0 | Fixed | 0.91 (0.84, 0.99) | 0.0335 |
| East Asian (20) | 697 (30.68) | 1205 (35.01) | 21.75 | 0.2969 | 12.6 | Fixed | 0.84 (0.78, 0.91) | <0.0001 |
| South Asian (4) | 162 (54.92) | 318 (72.27) | 19.47 | 0.0002 | 84.6 | Random | 0.78 (0.59, 1.04) | 0.0937 |
| Mixed (2) | 45 (36.00) | 95 (45.02) | 0.45 | 0.5036 | 0 | Fixed | 0.78 (0.59, 1.03) | 0.0828 |
| Middle Eastern (3) | 99 (38.37) | 128 (51.82) | 1.23 | 0.5395 | 0 | Fixed | 0.76 (0.63, 0.92) | 0.0054 |
| African (2) | 31 (38.27) | 72 (54.55) | 0.003 | 0.9564 | 0 | Fixed | 0.66 (0.48, 0.92) | 0.0150 |
| T (44) | 1945 (41.10) | 2228 (37.27) | 37.59 | 0.7044 | 0 | Fixed | 1.12 (1.06, 1.17) | <0.0001 |
| C (44) | 2787 (58.90) | 3751 (62.73) | 33.14 | 0.8608 | 0 | Fixed | 0.93 (0.90, 0.96) | <0.0001 |
| CC + CT (44) | 3910 (82.63) | 5054 (84.53) | 70.18 | 0.0055 | 38.7 | Random | 0.98 (0.96, 1.00) | 0.0624 |
| TT + CT (44) | 3068 (64.84) | 3532 (59.07) | 70.84 | 0.0048 | 39.3 | Random | 1.11 (1.07, 1.16) | <0.0001 |
| Subgroups | ||||||||
| TT Risk > 1 | 4062 | 5205 | ||||||
| TT (37) | 761 (18.73) | 833 (16.00) | 44.31 | 0.1610 | 18.8 | Fixed | 1.19 (1.09, 1.30) | 0.0002 |
| CT (37) | 1906 (46.92) | 2278 (43.77) | 57.03 | 0.0143 | 36.9 | Random | 1.10 (1.04, 1.17) | 0.0023 |
| CC (37) | 1395 (34.34) | 2094 (40.23) | 56.40 | 0.0164 | 36.2 | Random | 0.84 (0.79, 0.90) | <0.0001 |
| CC + CT (37) | 3301 (81.27) | 4372 (84.00) | 55.24 | 0.0211 | 34.8 | Random | 0.97 (0.95, 0.99) | 0.0041 |
| TT + CT (37) | 2667 (65.66) | 3111 (59.77) | 66.60 | 0.0014 | 45.9 | Random | 1.12 (1.07, 1.17) | <0.0001 |
| TT Risk < 1 | 545 | 563 | ||||||
| TT (5) | 52 (9.54) | 77 (13.68) | 1.06 | 0.9000 | 0 | Fixed | 0.66 (0.47, 0.92) | 0.0142 |
| T (5) | 269 (49.36) | 228 (40.50) | 4.72 | 0.3177 | 15.2 | Fixed | 1.23 (1.07, 1.40) | 0.0025 |
| CC (5) | 224 (41.10) | 258 (45.83) | 3.60 | 0.4630 | 0 | Fixed | 0.90 (0.79, 1.04) | 0.1552 |
| CC + CT (5) | 493 (90.46) | 486 (86.32) | 0.87 | 0.9293 | 0 | Fixed | 1.06 (1.01, 1.10) | 0.0133 |
| TT + CT (5) | 321 (58.90) | 305 (54.17) | 3.60 | 0.4634 | 0 | Fixed | 1.08 (0.97, 1.20) | 0.1492 |
| TT Risk varied | 125 | 211 | ||||||
| TT (2) | 9 (7.20) | 15 (7.11) | 2.56 | 0.1098 | 60.9 | Fixed | 1.01 (0.43, 2.33) | 0.9906 |
| CT (2) | 71 (56.80) | 101 (47.87) | 1.46 | 0.2263 | 31.7 | Fixed | 1.21 (0.98, 1.48) | 0.0704 |
| CC (2) | 45 (36.00) | 95 (45.02) | 0.45 | 0.5036 | 0 | Fixed | 0.79 (0.60, 1.04) | 0.0960 |
| CC + CT (2) | 116 (92.80) | 196 (92.89) | 2.76 | 0.0964 | 63.8 | Fixed | 1.00 (0.94, 1.06) | 0.9902 |
| TT + CT (2) | 80 (64.00) | 116 (54.98) | 0.13 | 0.718 | 0 | Fixed | 1.18 (0.99, 1.41) | 0.0656 |
| Partition Tree | Tukey’s Test | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Variable | AICc | AP Death Levels | Count | Mean | SD | Levels Compared | Difference | SE Difference | Lower CI | Upper CI | p |
| TT%ct | 318.732 | 2 | 11 | 8.182 | 7.427 | 4/2 | 7.874 | 3.319 | −0.198 | 15.945 | 0.0572 |
| 3 and 4 | 33 | 15.364 | 8.930 | 3/2 | 6.3515 | 3.443 | −2.021 | 14.724 | 0.1680 | ||
| 4/3 | 1.5222 | 3.032 | −5.851 | 8.896 | 0.8707 | ||||||
| TT%ca | 308.987 | 2 | 11 | 7.091 | 4.549 | 3/2 | 11.042 | 3.089 | 3.731 | 18.753 | 0.0021 |
| 4 and 3 | 33 | 17.939 | 8.441 | 4/2 | 10.520 | 2.978 | 3.279 | 17.762 | 0.0029 | ||
| 3/4 | 0.722 | 2.720 | −5.893 | 7.337 | 0.9619 | ||||||
| CT%ct | 317.809 | 2 | 11 | 34.273 | 10.762 | 3/2 | 9.661 | 3.407 | 1.377 | 17.944 | 0.0190 |
| 4 and 3 | 33 | 43.091 | 7.666 | 4/2 | 8.116 | 3.284 | 0.130 | 16.102 | 0.0457 | ||
| 3/4 | 1.544 | 3.000 | −5.751 | 8.840 | 0.8646 | ||||||
| CT%ca | 339.208 | 2 and 3 | 26 | 46.115 | 11.389 | 4/2 | 3.096 | 4.195 | −7.104 | 13.296 | 0.7425 |
| 4 | 18 | 48.278 | 9.999 | 3/2 | 1.618 | 4.351 | −8.962 | 12.198 | 0.9267 | ||
| 4/3 | 1.478 | 3.832 | −7.840 | 10.796 | 0.9214 | ||||||
| CC%ct | 366.743 | 3 and 4 | 33 | 41.576 | 14.331 | 2/3 | 16.055 | 5.959 | 1.564 | 30.545 | 0.0269 |
| 2 | 11 | 57.455 | 16.336 | 2/4 | 15.732 | 5.745 | 1.762 | 29.703 | 0.0242 | ||
| 4/3 | 0.322 | 5.248 | −12.440 | 13.084 | 0.9979 | ||||||
| CC%ca | 364.011 | 4 and 3 | 33 | 34.545 | 12.696 | 2/4 | 13.616 | 5.567 | 0.078 | 27.154 | 0.0484 |
| 2 | 11 | 47.727 | 18.778 | 2/3 | 12.661 | 5.775 | −1.382 | 26.703 | 0.0845 | ||
| 3/4 | 0.956 | 5.086 | −11.412 | 13.323 | 0.9807 | ||||||
| RRTT | 118.544 | 2 | 11 | 1.073 | 1.016 | 4/2 | 0.377 | 0.342 | −0.455 | 1.209 | 0.5180 |
| 3 and 4 | 33 | 1.424 | 0.838 | 3/2 | 0.321 | 0.355 | −0.542 | 1.184 | 0.6413 | ||
| 4/3 | 0.057 | 0.313 | −0.703 | 0.817 | 0.9821 | ||||||
| RRCT | 7.777 | 3 and 4 | 33 | 1.124 | 0.206 | 2/3 | 0.250 | 0.100 | 0.006 | 0.493 | 0.0433 |
| 2 | 11 | 1.336 | 0.359 | 2/4 | 0.181 | 0.097 | −0.054 | 0.415 | 0.1593 | ||
| 4/3 | 0.069 | 0.088 | −0.145 | 0.283 | 0.7164 | ||||||
| RRCC | −21.842 | 2 and 4 | 29 | 0.822 | 0.185 | 3/2 | 0.026 | 0.072 | −0.149 | 0.201 | 0.9299 |
| 3 | 15 | 0.848 | 0.167 | 3/4 | 0.026 | 0.063 | −0.128 | 0.180 | 0.9132 | ||
| 4/2 | 0.0004 | 0.069 | −0.168 | 0.169 | 1.0000 | ||||||
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.-M.; Chen, Z.-F.; Young, L.; Shiao, S.P.K. Meta-Prediction of the Effect of Methylenetetrahydrofolate Reductase Polymorphisms and Air Pollution on Alzheimer’s Disease Risk. Int. J. Environ. Res. Public Health 2017, 14, 63. https://doi.org/10.3390/ijerph14010063
Wu S-M, Chen Z-F, Young L, Shiao SPK. Meta-Prediction of the Effect of Methylenetetrahydrofolate Reductase Polymorphisms and Air Pollution on Alzheimer’s Disease Risk. International Journal of Environmental Research and Public Health. 2017; 14(1):63. https://doi.org/10.3390/ijerph14010063
Chicago/Turabian StyleWu, Suh-Mian, Zhao-Feng Chen, Lufei Young, and S. Pamela K. Shiao. 2017. "Meta-Prediction of the Effect of Methylenetetrahydrofolate Reductase Polymorphisms and Air Pollution on Alzheimer’s Disease Risk" International Journal of Environmental Research and Public Health 14, no. 1: 63. https://doi.org/10.3390/ijerph14010063
APA StyleWu, S.-M., Chen, Z.-F., Young, L., & Shiao, S. P. K. (2017). Meta-Prediction of the Effect of Methylenetetrahydrofolate Reductase Polymorphisms and Air Pollution on Alzheimer’s Disease Risk. International Journal of Environmental Research and Public Health, 14(1), 63. https://doi.org/10.3390/ijerph14010063