
Possible Internalization of an Enterovirus in Hydroponically Grown Lettuce




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Virus and Cell Cultures
2.2. Hydroponic Cultures Preparation

2.3. Experimental Contamination
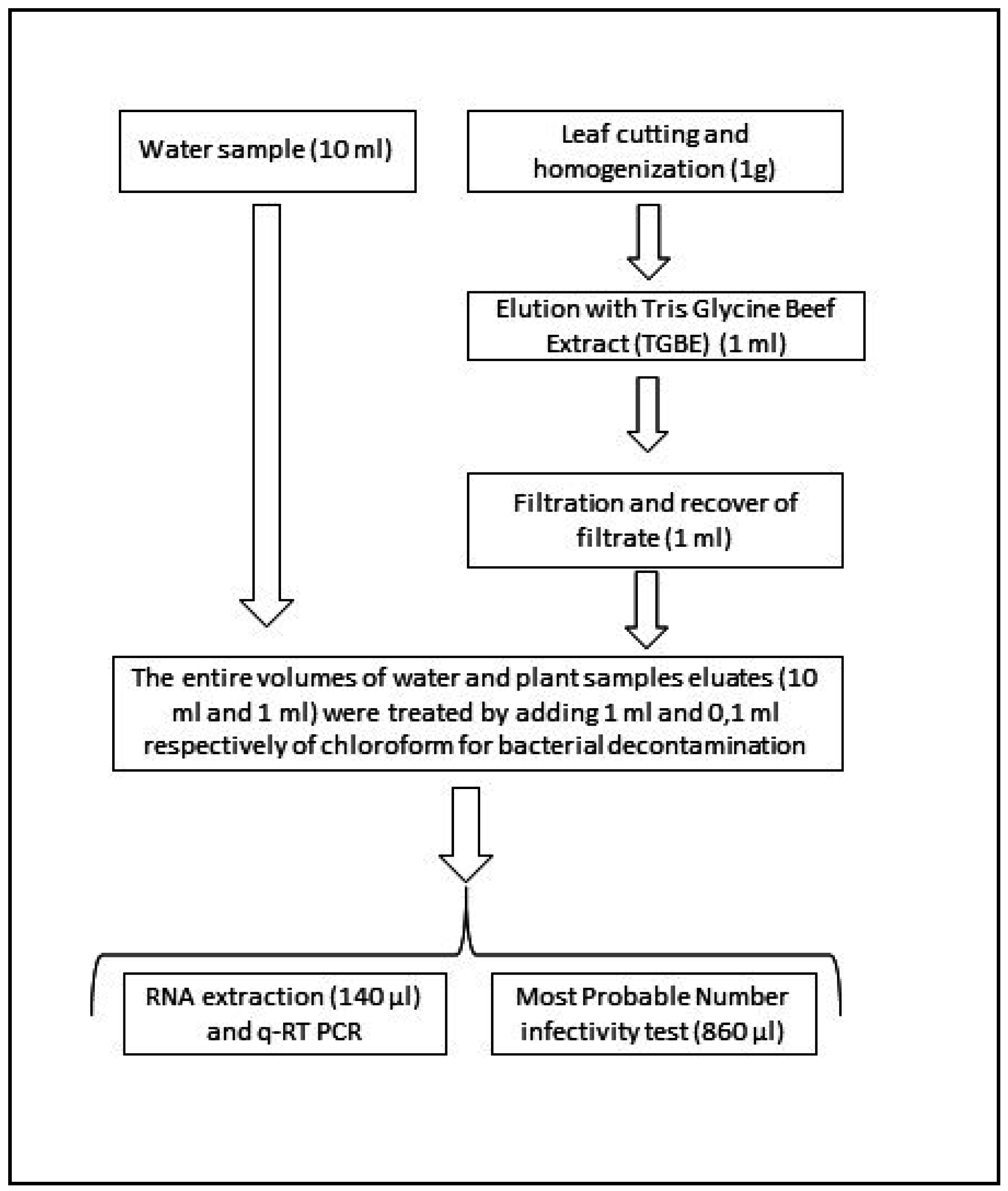
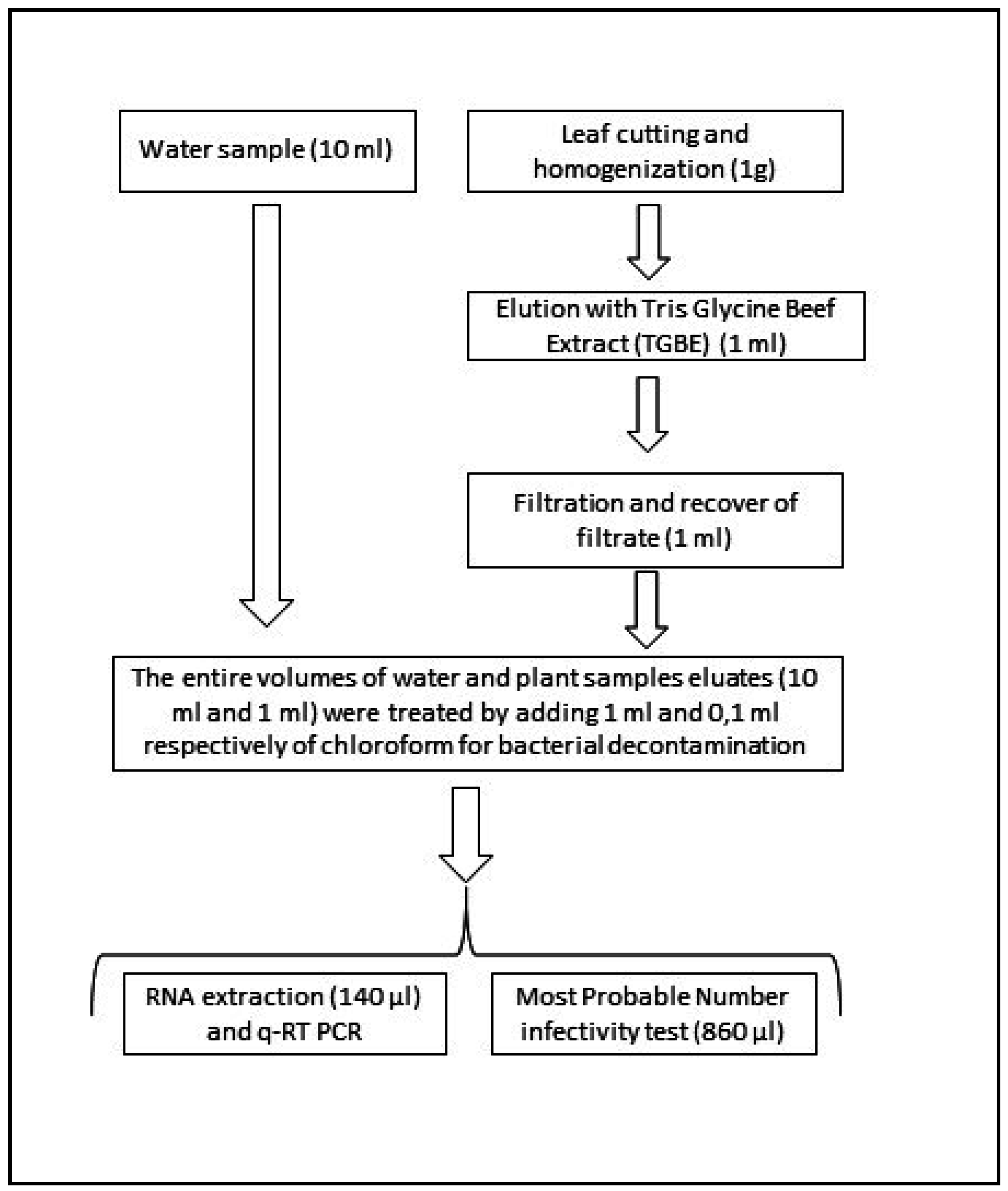
2.4. Water Sample Processing
2.5. Plant Sample Processing
2.6. Virus Quantification
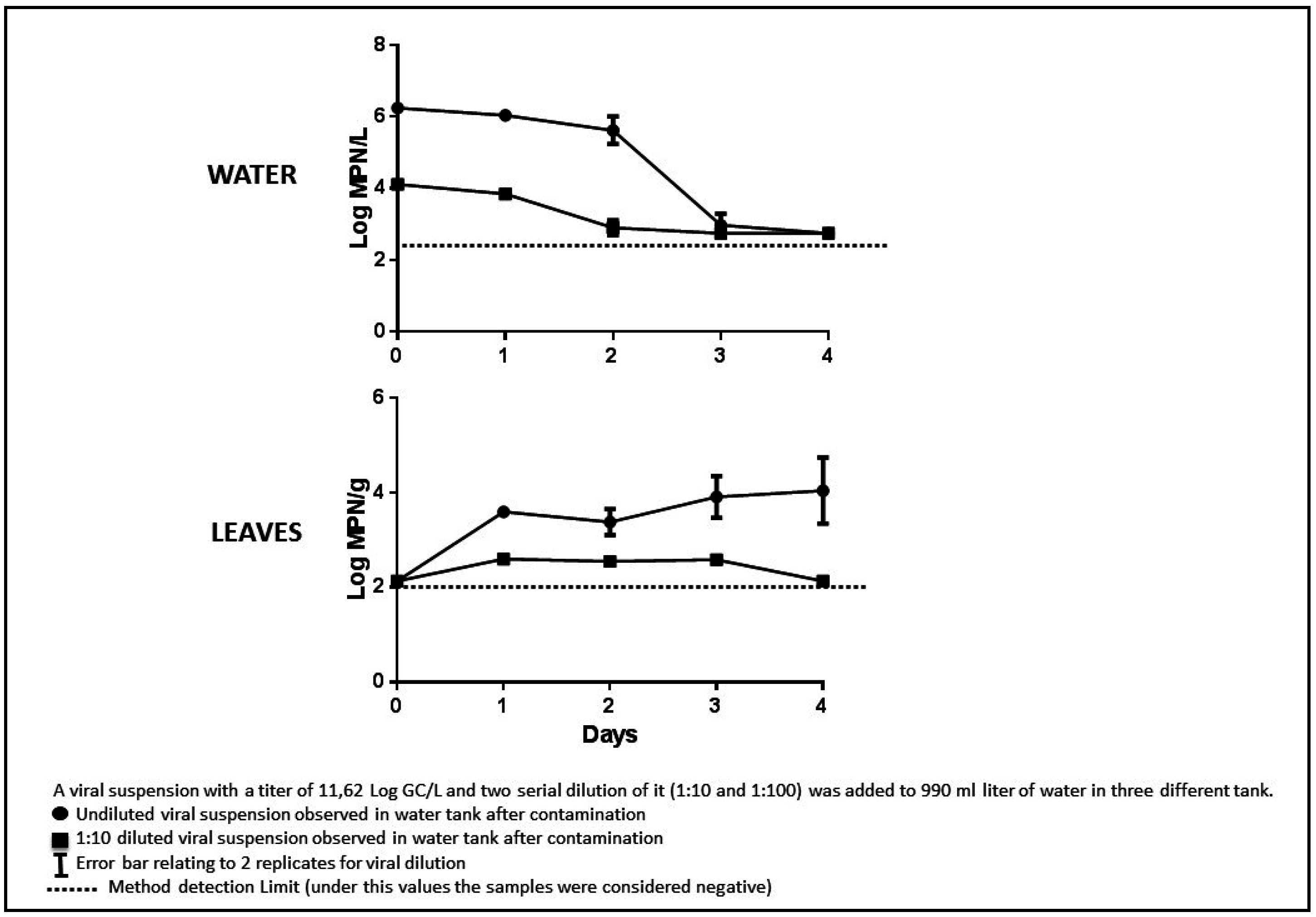
2.6.1. MPN Infectivity Tests
qRT-PCR
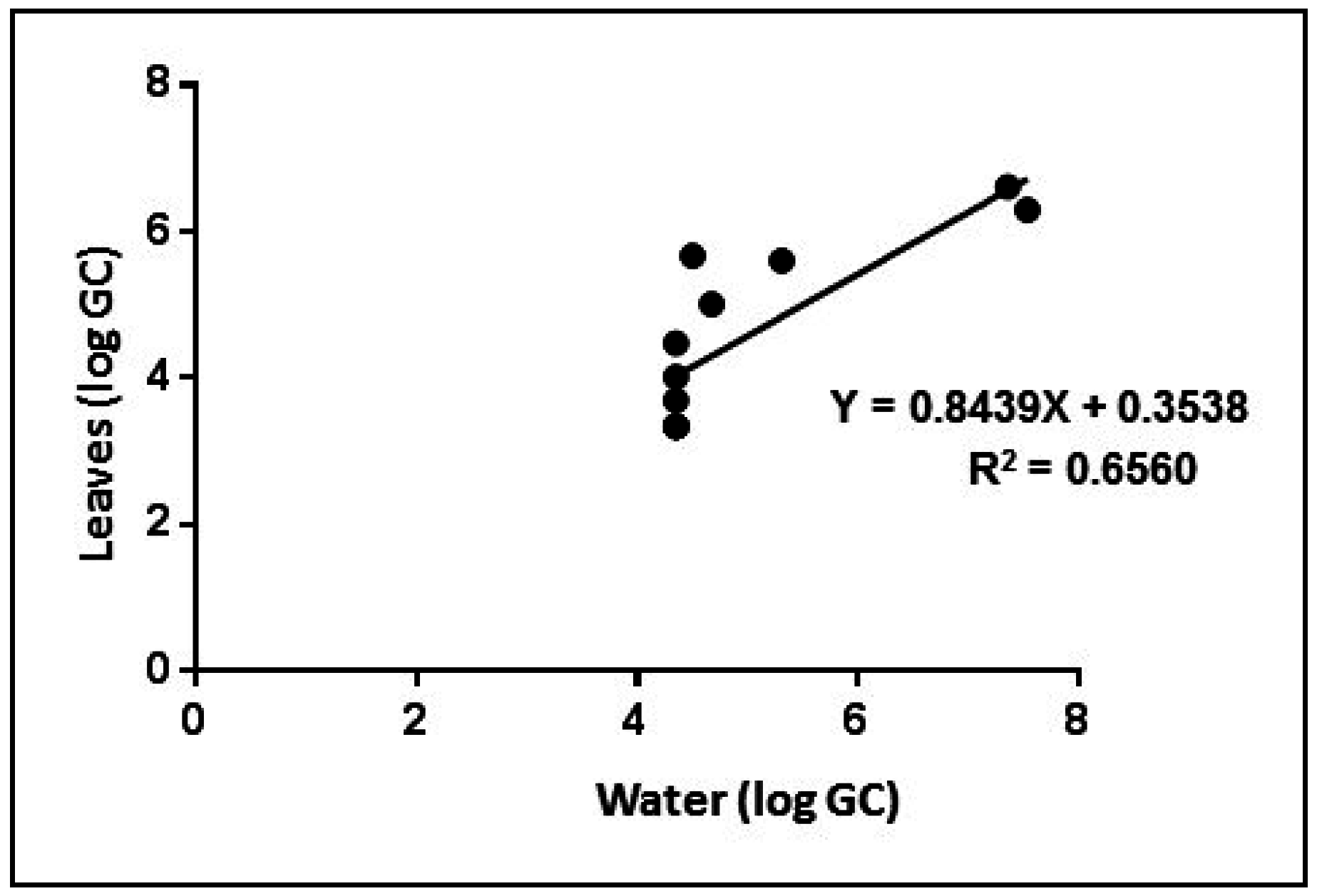
2.7. Data Analysis
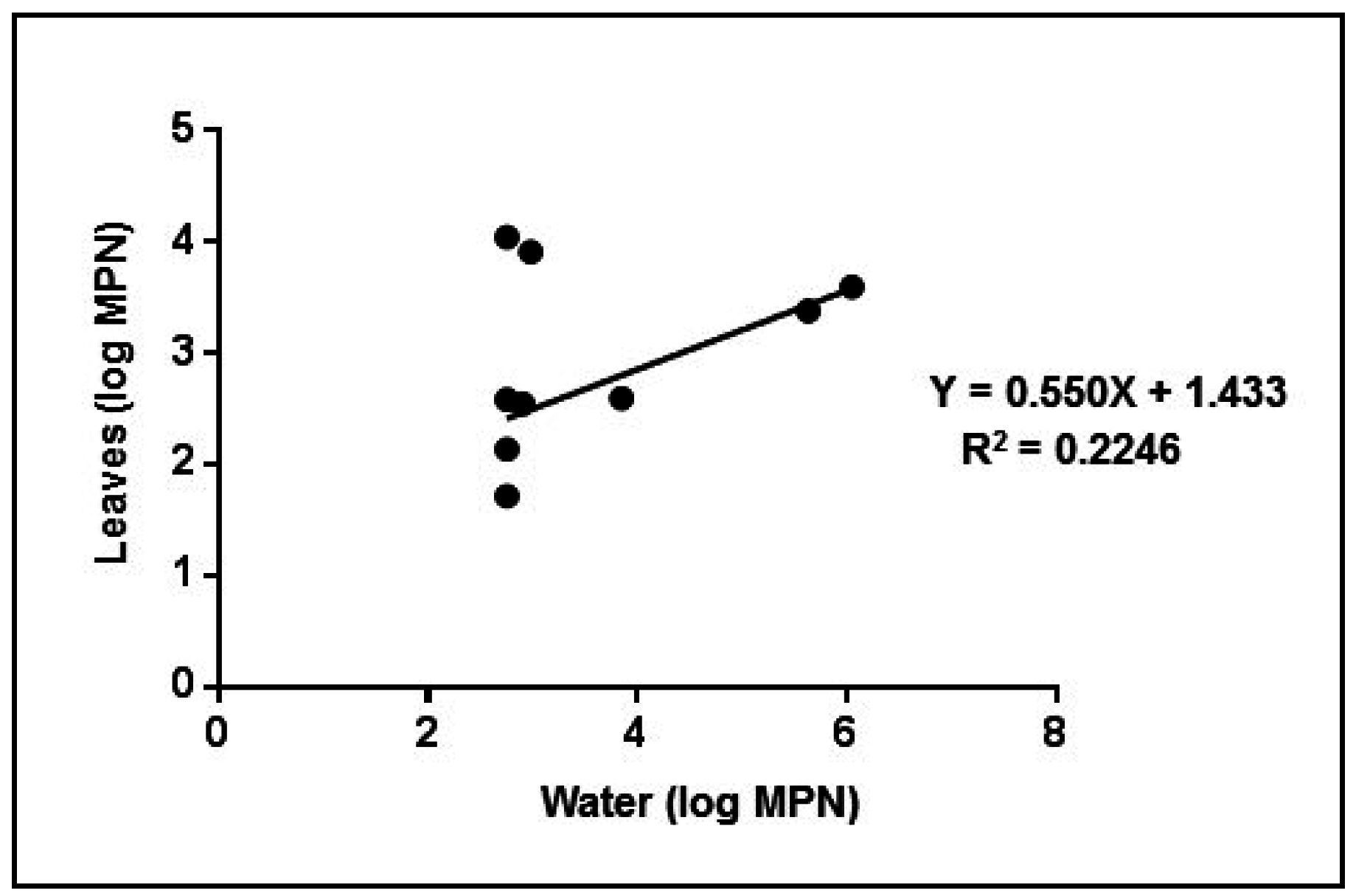
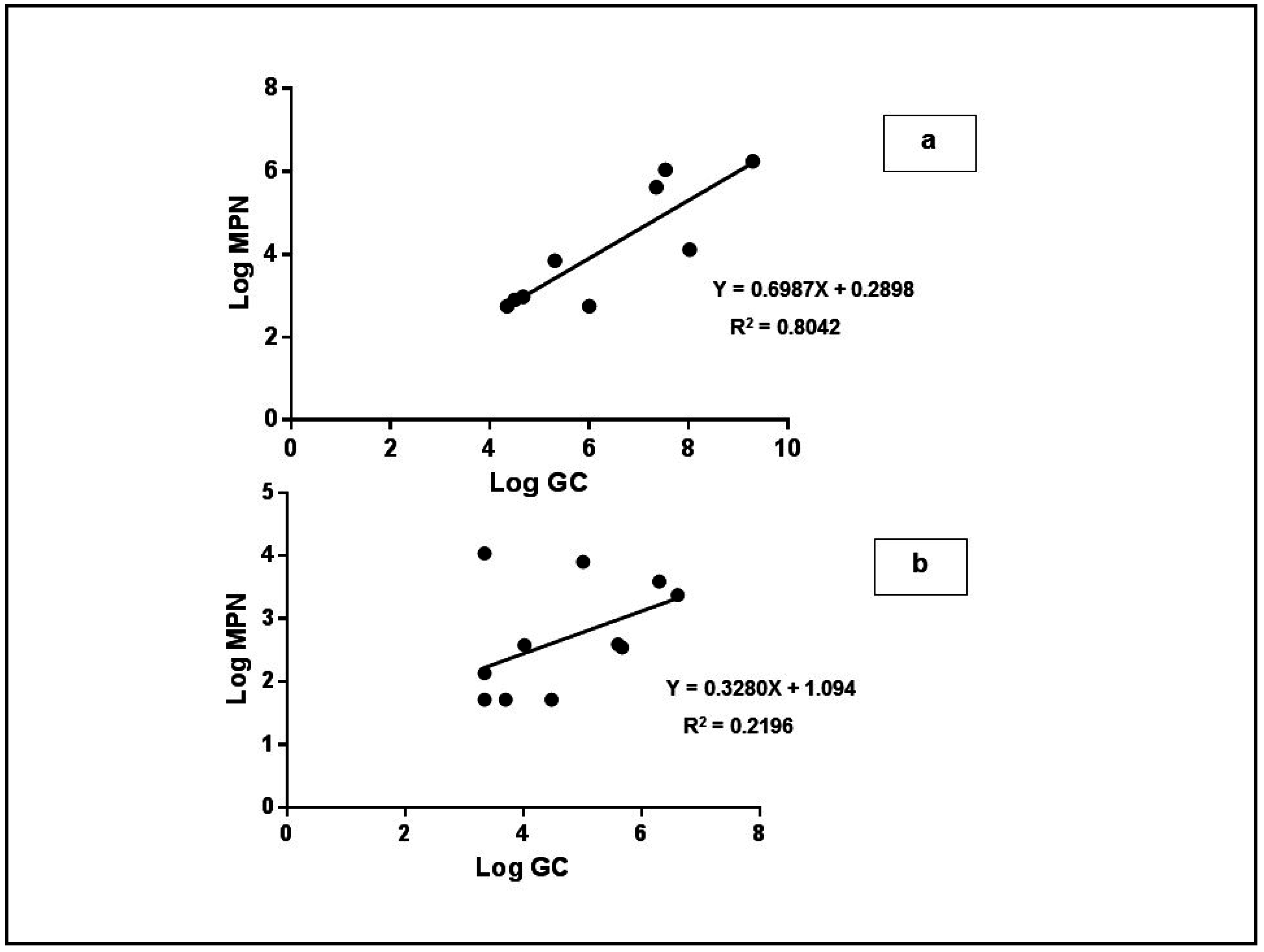
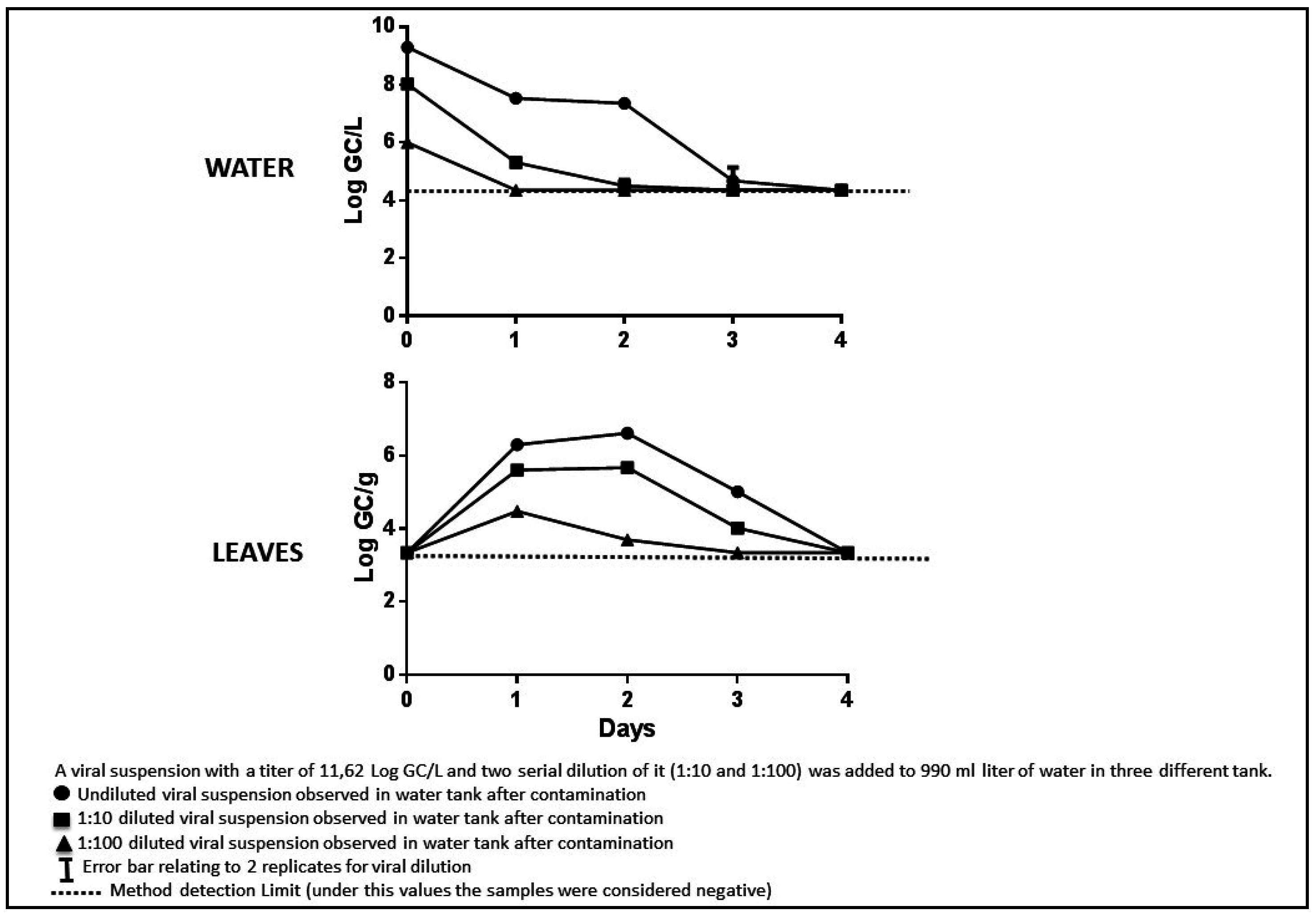
3. Results and Discussion
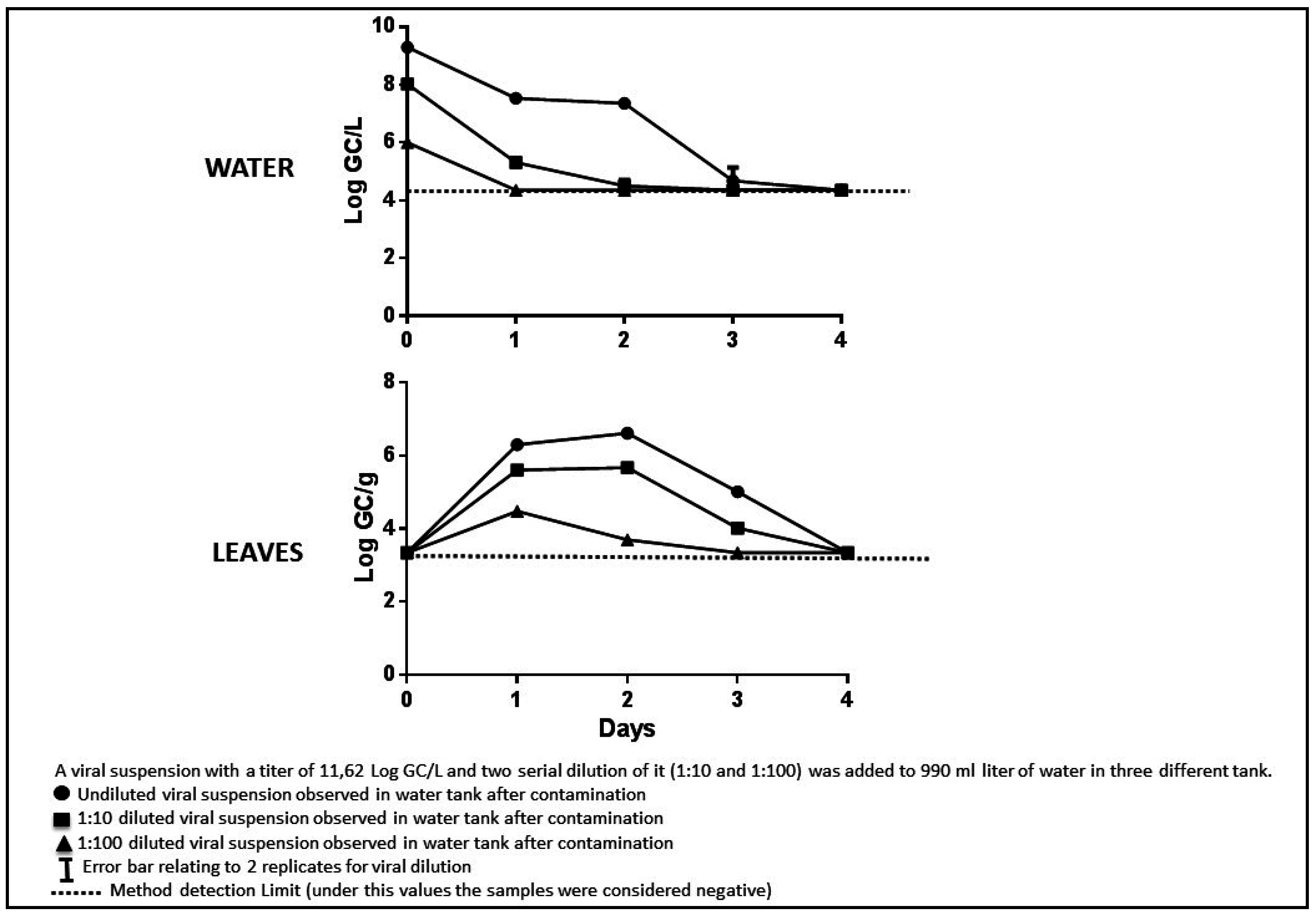
3.1. Starting Plant Contamination
3.2. Starting Water Titre
3.3. Methods’ Detection Limits
3.4. Experimental Tests
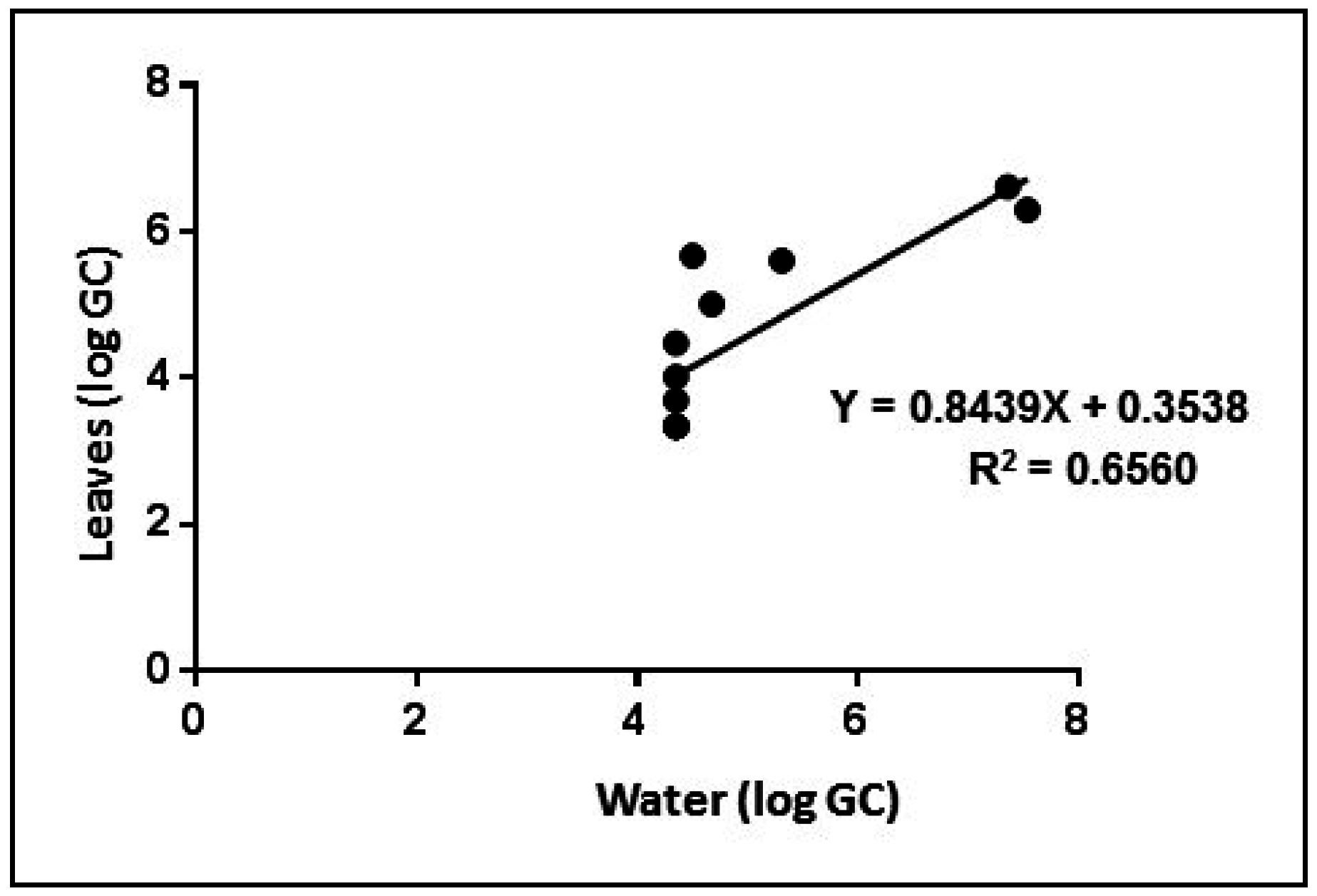
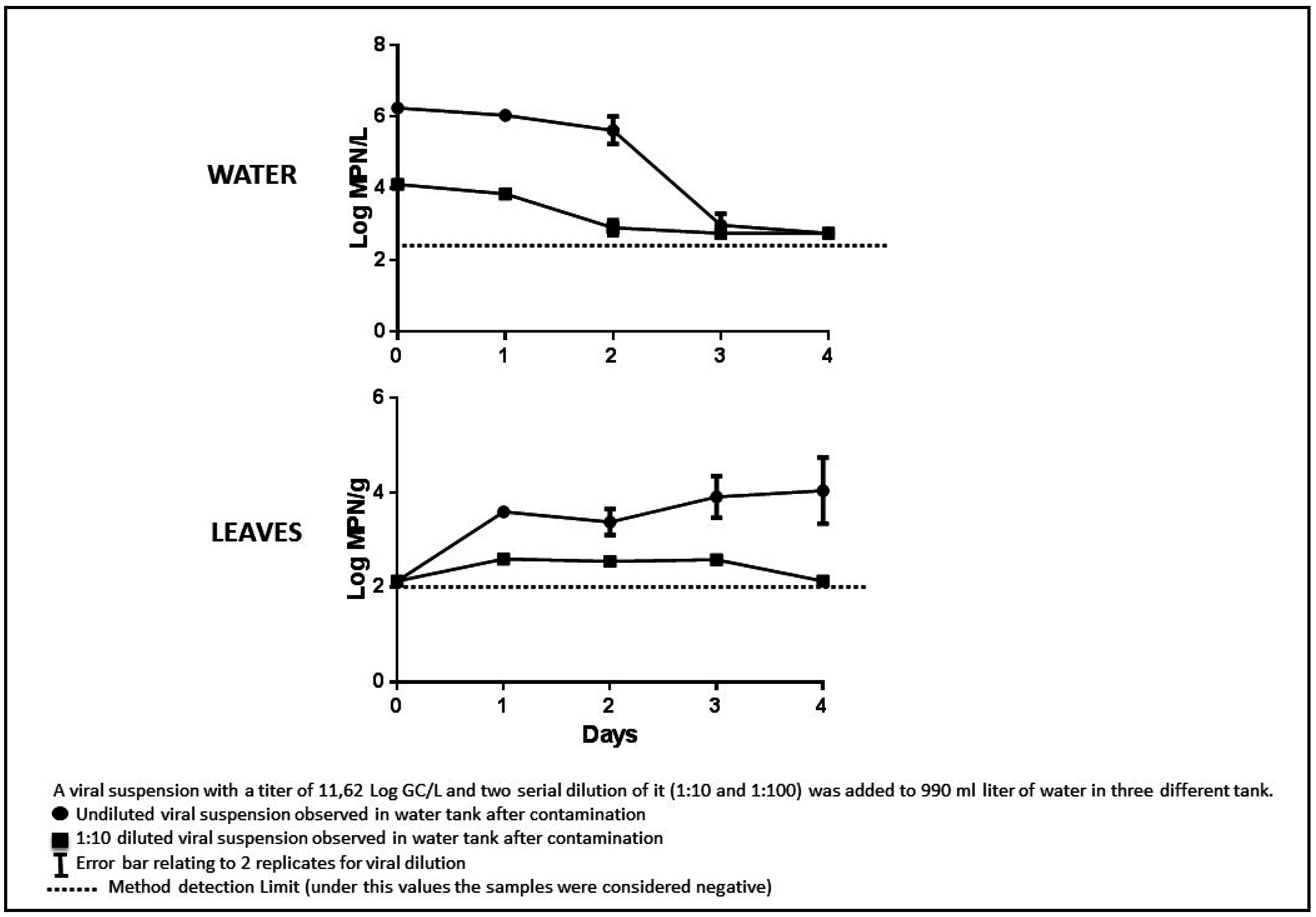
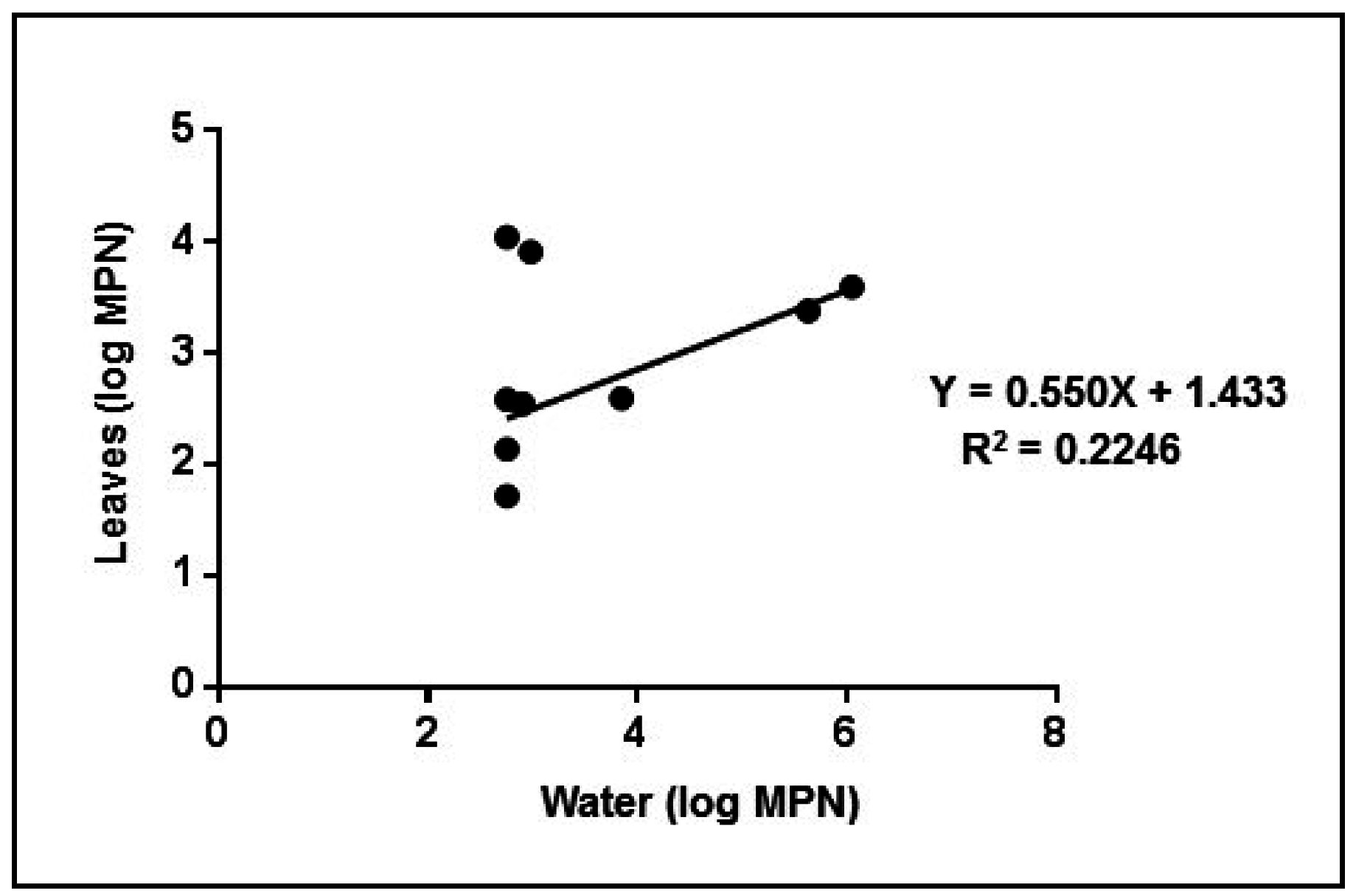

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Butot, S.; Putallaz, T.; Amoroso, R.; Sanchez, G. Inactivation of enteric viruses in minimally processed berries and herbs. Appl. Environ. Microbiol. 2009, 75, 4155–4161. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority and European Centre for Disease Prevention and Control. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2013. Scientific Report of EFSA and ECDC. EFSA J. 2015, 13. [Google Scholar] [CrossRef]
- Rosenblum, L.S.; Mirkin, I.R.; Allen, D.T.; Safford, S.; Hadler, S.C. A multifocal outbreak of hepatitis A traced to commercial distributed lettuce. Am. J. Public Health 1990, 80, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.H.; Kim, N.Y.; Koh, Y.J.; Lee, H.J. Epidemiology of foodborne norovirus outbreak in Incheon Korea. J. Korean Med. Sci. 2010, 25, 1128–1133. [Google Scholar] [CrossRef] [PubMed]
- Ethelberg, S.; Lisby, M.; Bottiger, B.; Schultz, A.C.; Villif, A.; Jensen, T.; Olsen, K.E.; Scheutz, F.; Kjelso, C.; Muller, L. Outbreaks of gastroenteritis linked to lettuce, Denmark, January 2010. Eurosurveillance 2010, 15, 6:1–6:3. [Google Scholar]
- DiCaprio, E.; Ma, Y.; Purgianto, A.; Hughes, J.; Li, J. Internalization and dissemination of human norovirus and animal calicivirus in hydroponically grown romaine lettuce. Appl. Environ. Microbiol. 2012, 78, 6143–6152. [Google Scholar] [CrossRef] [PubMed]
- Little, C.L.; Gillespie, I.A. Prepared salads and public health. J. Appl. Microbiol. 2008, 105, 1729–1743. [Google Scholar] [CrossRef] [PubMed]
- Widdowson, M.A.; Sulka, A.; Bulens, S.N.; Beard, R.S.; Chaves, S.S.; Hammond, R.; Salehi, E.D.; Swanson, E.; Totaro, J.; Woron, R. Norovirus and foodborne disease, United States, 1991–2000. Emer. Infect. Dis. 2005, 11, 95–102. [Google Scholar] [CrossRef] [PubMed]
- De Weal, C.S.; Bhuiya, F. Outbreaks by the numbers: Fruits and vegetables 1990–2005. In Proceedings of the 2007 Annual Meeting of the International Association for Food Protection, Lake Buena Vista, FL, USA, 8–11 July 2007.
- Baert, L.; Mattison, K.; Loisy-Hamon, F.; Harlow, J.; Martyres, A.; Lebeau, B.; Stals, A.; Van Coillie, E.; Herman, L.; Uyttendaele, M. Norovirus prevalence in Belgian, Canadian and French fresh produce: A threat to human health? Int. J. Food Microbiol. 2011, 151, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Montaño-Remacha, C.; Ricotta, L.; Alfonsi, V.; Bella, A.; Tosti, M.E.; Ciccaglione, A.R.; Bruni, R.; Taffon, S.; Equestre, M.; Losio, M.N.; et al. Hepatitis A outbreak in Italy, 2013: A matched case-control study. Eurosurveillance 2014, 19, 37:1–37:7. [Google Scholar] [CrossRef]
- Kurdziel, A.S.; Wiljìkinson, N.; Langton, S.; Cook, N. Survival of poliovirus on soft fruits and salad vegetables. J. Food Prot. 2001, 64, 706–709. [Google Scholar] [PubMed]
- Cheong, S.; Lee, C.; Song, S.W.; Choi, W.C.; Lee, C.H.; Kim, S.J. Enteric viruses in raw vegetables and groundwater used for irrigation in South Korea. Appl. Environ. Microbiol. 2009, 75, 7745–7751. [Google Scholar] [CrossRef] [PubMed]
- Gilgen, M.; Wegmuller, B.; Burkhalter, P.; Buhler, H.P.; Muller, U.; Luthy, J.; Candrian, U. Reverse transcription PCR to detect enteroviruses in surface water. Appl. Environ. Microbiol. 1995, 61, 1226–1231. [Google Scholar] [PubMed]
- Rajtar, B.; Majek, M.; Polański, L.; Polz-Dacewicz, M. Enteroviruses in water environment—A potential threat to public health. Ann. Agr. Environ. Med. 2008, 15, 199–203. [Google Scholar]
- Kokkinos, P.; Kozyra, I.; Lazic, S.; Bouwknegt, M.; Rutjes, S.; Willems, K. Harmonised investigation of the occurrence of human enteric viruses in the leafy green vegetable supply chain in three European countries. Food Environ. Virol. 2012, 4, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Allwood, P.B.; Malik, J.S.; Hedberg, C.W.; Goyal, S.M. Effect of temperature and sanitizers on the survival of feline calicivirus, Escherichia coli, and F-specific coliphage MS2 on leafy salad vegetables. J. Food Prot. 2004, 67, 1468–1472. [Google Scholar]
- Butot, S.; Putallaz, T.; Sánchez, G. Effects of sanitation, freezing and frozen storage on enteric viruses in berries and herbs. Int. J. Food Microbiol. 2008, 126, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Baert, L.; Debevere, J.; Uyttendaele, M. The efficacy of preservation methods to inactivate foodborne viruses. Int. J. Food Microbiol. 2009, 131, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Solomon, E.B.; Yaron, S.; Mattews, K.R. Transmission of Escherichia coli O157:H7 from contaminated manure and irrigation water to lettuce plant tissue and its subsequent internalization. Appl. Environ. Microbiol. 2002, 68, 397–400. [Google Scholar] [CrossRef] [PubMed]
- Erickson, M.C.; Webb, C.C.; Diaz-Perez, J.C.; Phatak, S.C.; Silvoy, J.J.; Davey, L.; Payton, A.S.; Liao, J.; Ma, L.; Doyle, M.P. Surface and internalized Escherichia coli O157:H7 on field-grown spinach and lettuce treated with spray-contaminated irrigation water. J. Food Prot. 2010, 73, 1023–1029. [Google Scholar] [PubMed]
- Hirneisen, K.A.; Sharma, M.; Kniel, K.E. Human enteric pathogen internalization by root uptake into food crops. Foodborne Pathog. Dis. 2009, 9, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Deboosere, N.; Pinon, A.; Caudrelier, Y.; Delobel, A.; Merle, G.; Perelle, S.; Temmam, S.; Loutreul, J.; Morin, T.; Estienney, M.; et al. Adhesion of human pathogenic enteric viruses and surrogate viruses to inert and vegetal food surfaces. Food Microbiol. 2012, 32, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Verhaelen, K.; Bouwknegt, M.; Carratalà, A.; Lodder-Verschoor, F.; Diez-Valcarce, M.; Rodríguez-Lázaro, D.; de Roda Husman, A.M.; Rutjes, S.A. Virus transfer proportions between gloved fingertips, soft berries, and lettuce, and associated health risks. Int. J. Food Microbiol. 2013, 166, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.J.; Stagnitti, F.; Premier, R.; Boland, A.M.; Hale, G. Quantitative microbial risk assessment models for consumption of raw vegetables irrigated with reclaimed water. Appl. Environ. Microbiol. 2006, 72, 3284–3290. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Jin, Y.; Sims, T.; Kniel, K.E. Internalization of murine norovirus 1 by Lactuca sativa during irrigation. Appl. Environ. Microbiol. 2011, 77, 2508–2512. [Google Scholar] [CrossRef] [PubMed]
- Gerba, C.P.; Choi, C.Y. Role of irrigation water in crop contamination by viruses. In Viruses in Foods, 1st ed.; Goyal, S.M., Ed.; Springer: New York, NY, USA, 2010; pp. 257–267. [Google Scholar]
- Urbanucci, A.; Myrmel, M.; Berg, I.; von Bornsdorff, C.H.; Maunula, L. Potential internalisation of calicivirus in Lettuce. Int. J. Food Microbiol. 2009, 135, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Esseili, M.A.; Wang, Q.; Zhang, Z.; Saif, L.J. Internalization of sapovirus, a surrogate for norovirus, in romaine lettuce and the effect of lettuce latex on virus infectivity. Appl. Environ. Microbiol. 2012, 78, 6271–6279. [Google Scholar] [CrossRef] [PubMed]
- Alum, A. Control of Viral Contamination of Reclaimed Water Irrigated Vegetables by Drip Irrigation. Ph.D. Dissertation, University of Arizona, Tucson, AZ, USA, 2001. [Google Scholar]
- Gerba, C.P.; Kitajima, M.; Iker, B. Viral presence in waste water and sewage and control methods. In Viruses in Food and Water. Risks, Surveillance and Control, 1st ed.; Nigel, C., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2013; pp. 293–311. [Google Scholar]
- Steele, M.; Odumeru, J. Irrigation water as source of foodborne pathogens on fruit and vegetables. J. Food Prot. 2006, 67, 2839–2849. [Google Scholar]
- Bosch, A.; Sánchez, G.; Abbaszadegan, M.; Carducci, A.; Guix, S.; Le Guyader, F.S.; Netshikweta, R.; Pintó, R.M.; vander Poel, W.V.H.; Rutjes, S.; et al. Analytical methods for virus detection in water and food. Food Anal. Method. 2011, 4, 4–12. [Google Scholar] [CrossRef]
- Calgua, B.; Rodriguez-Manzano, J.; Hundesa, A.; Suñen, E.; Calvo, M.; Bofill-Mas, S.; Girones, R. New methods for the concentration of viruses from urban sewage using quantitative PCR. J. Virol. Method 2013, 187, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Michen, B.; Graule, T. Isoelectric points of viruses. J. Appl. Microbiol. 2010, 109, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Hurley, M.A.; Roscoe, M.E. Automated statistical analysis of microbial enumeration by dilution series. J. Appl. Bacteriol. 1983, 55, 159–164. [Google Scholar] [CrossRef]
- Donaldson, K.A.; Griffin, D.W.; Paul, D.H. Detection quantitation and identification of enteroviruses from surface waters and sponge tissue from the Florida Keys using real-time RT-PCR. Water Res. 2002, 36, 2505–2514. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Snyder, W.C. Nutrition of strawberry plant under controlled conditions: (1) Effects of deficiencies of boron and certain other elements: (b) Susceptibility to injury from sodium salts. Proc. Am. Soc. Hort. Sci. 1933, 30, 288–294. [Google Scholar]
- Carducci, A.; Battistini, R.; Rovini, E.; Verani, M. Viral removal by wastewater treatment: Monitoring of indicators and pathogens. Food Environ. Virol. 2009, 1, 85–91. [Google Scholar] [CrossRef]
- Butot, S.; Putallaz, T.; Sànchez, G. Procedure for rapid concentration and detection of enteric viruse from berries and vegetables. Appl. Environ. Microbiol. 2007, 73, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Carducci, A.; Ciurli, A.; Verani, M. Deep viral contamination of lettuce through root absorbtion. In Proceedings of the 16th International Symposium on Health-Related Water Microbiology, Rotorua, New Zealand, 18–23 September 2011.
- Nordgren, J.; Matussek, A.; Mattsson, A.; Svensson, L.; Lindgren, P.E. Prevalence of norovirus and factors influencing virus concentrations during one year in a full-scale wastewater treatment plant. Water Res. 2009, 43, 1117–1125. [Google Scholar] [CrossRef] [PubMed]
- Carducci, A.; Federigi, I.; Verani, M. Virus occupational exposure in solid waste processing facilities. Ann. Occup. Hyg. 2013, 57, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.; Voice, T.C.; Xagoraraki, I. Effect of organic carbon on sorption of human adenovirus to soil particles and laboratory containers. Water Res. 2013, 47, 3339–3346. [Google Scholar] [CrossRef] [PubMed]
- Girones, R.; Ferrús, M.A.; Alonso, J.L.; Rodriguez-Manzano, J.; Calgua, B.; Corrêa Ade, A.; Hundesa, A.; Carratala, A.; Bofill-Mas, S. Molecular detection of pathogens in water—The pros and cons of molecular techniques. Water Res. 2010, 44, 4325–4339. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carducci, A.; Caponi, E.; Ciurli, A.; Verani, M. Possible Internalization of an Enterovirus in Hydroponically Grown Lettuce. Int. J. Environ. Res. Public Health 2015, 12, 8214-8227. https://doi.org/10.3390/ijerph120708214
Carducci A, Caponi E, Ciurli A, Verani M. Possible Internalization of an Enterovirus in Hydroponically Grown Lettuce. International Journal of Environmental Research and Public Health. 2015; 12(7):8214-8227. https://doi.org/10.3390/ijerph120708214
Chicago/Turabian StyleCarducci, Annalaura, Elisa Caponi, Adriana Ciurli, and Marco Verani. 2015. "Possible Internalization of an Enterovirus in Hydroponically Grown Lettuce" International Journal of Environmental Research and Public Health 12, no. 7: 8214-8227. https://doi.org/10.3390/ijerph120708214
APA StyleCarducci, A., Caponi, E., Ciurli, A., & Verani, M. (2015). Possible Internalization of an Enterovirus in Hydroponically Grown Lettuce. International Journal of Environmental Research and Public Health, 12(7), 8214-8227. https://doi.org/10.3390/ijerph120708214