
Carbamate Pesticide-Induced Apoptosis in Human T Lymphocytes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cells
2.3. Carbamate Pesticide-Induced Apoptosis in Jurkat T Cells Determined by FITC-Annexin V/PI Staining
2.4. Determination of Intracellular Levels of Active Caspase-3 in Jurkat Cells by Flow Cytometry
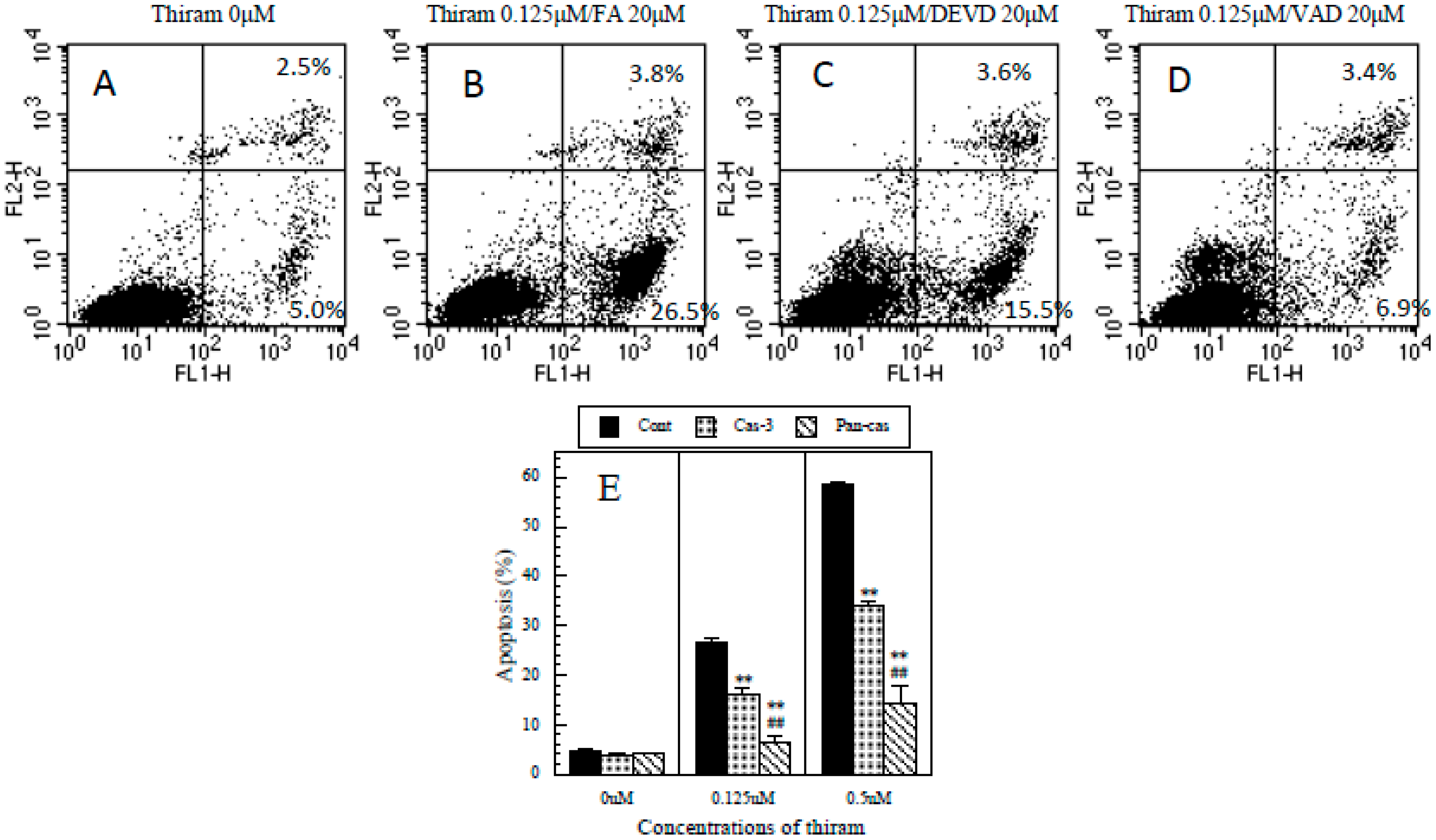
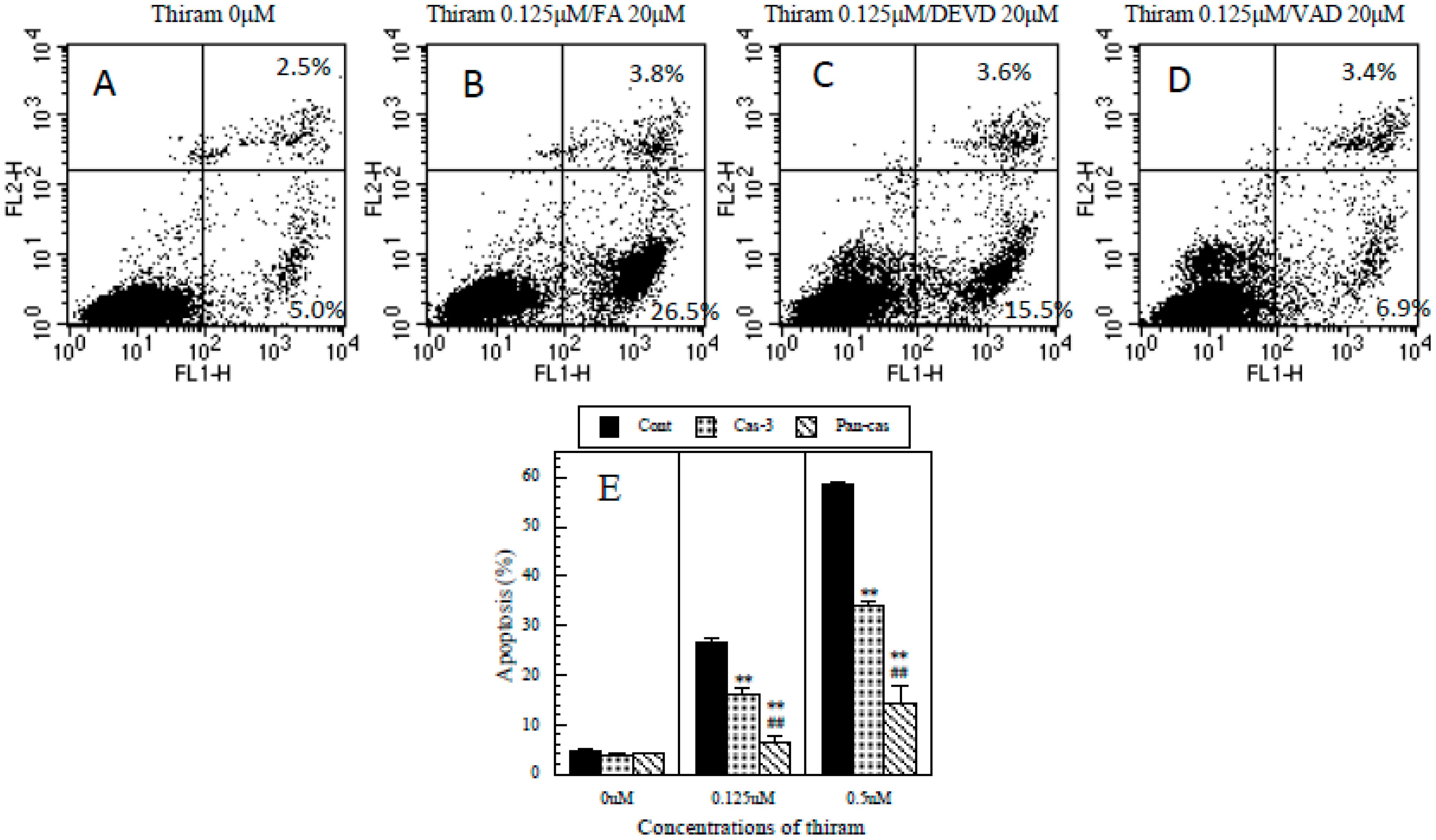
2.5. Protective Effects of Caspase-3 and General Caspase Inhibitors against Thiram-Induced Apoptosis in Jurkat T Cells
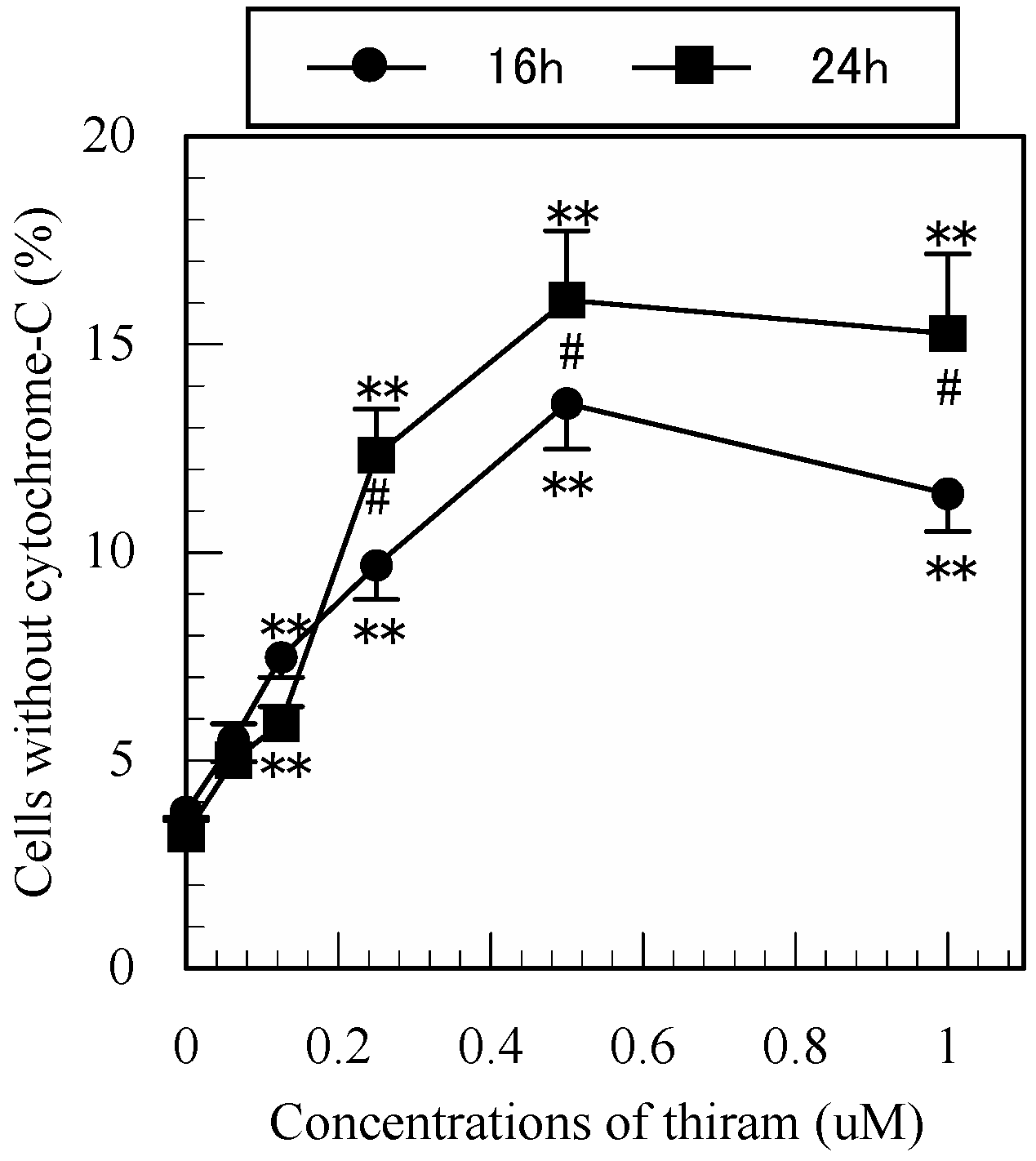
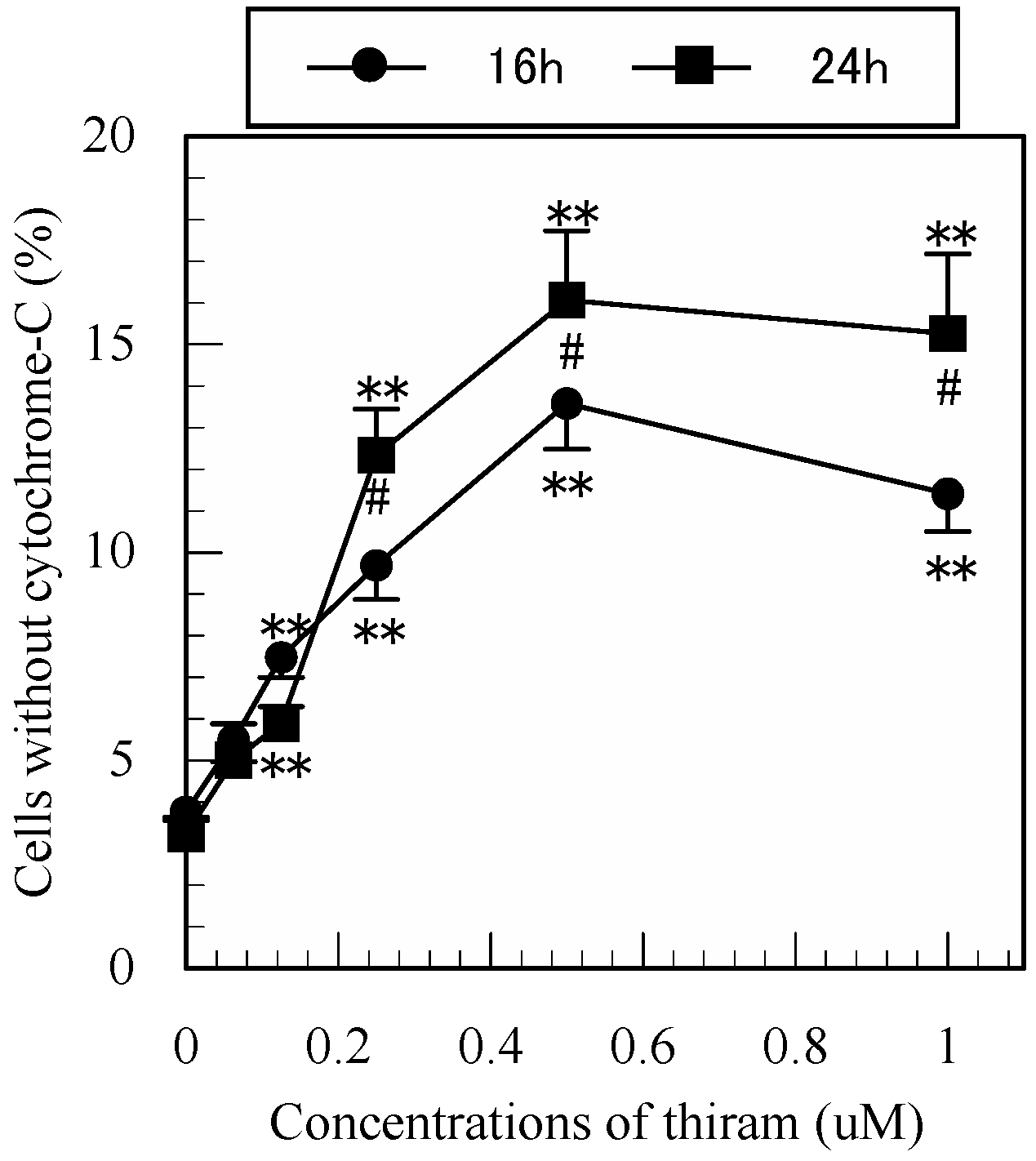
2.6. Analysis of Cytochrome-c Release
2.7. Statistical Analyses
3. Results
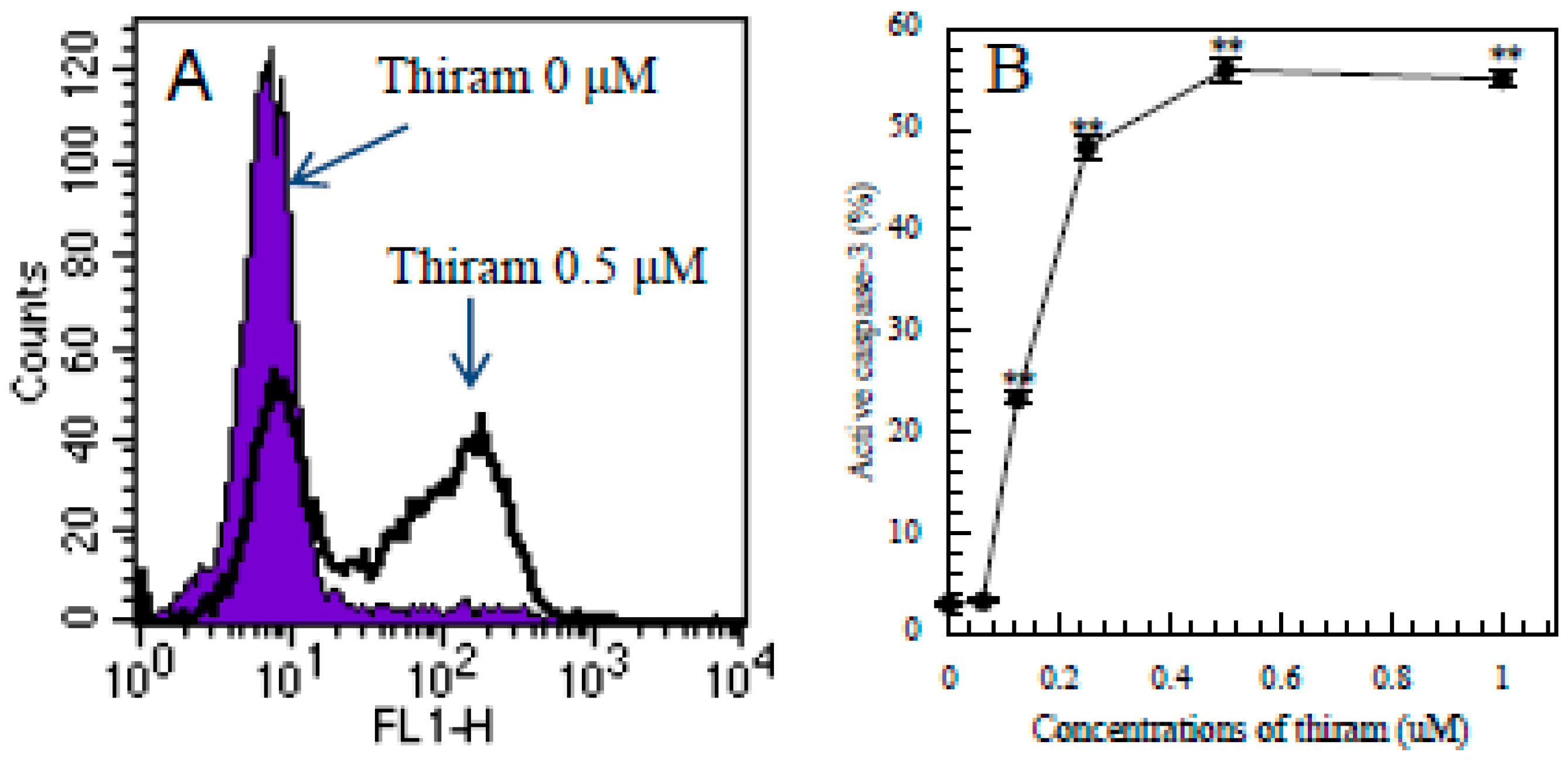
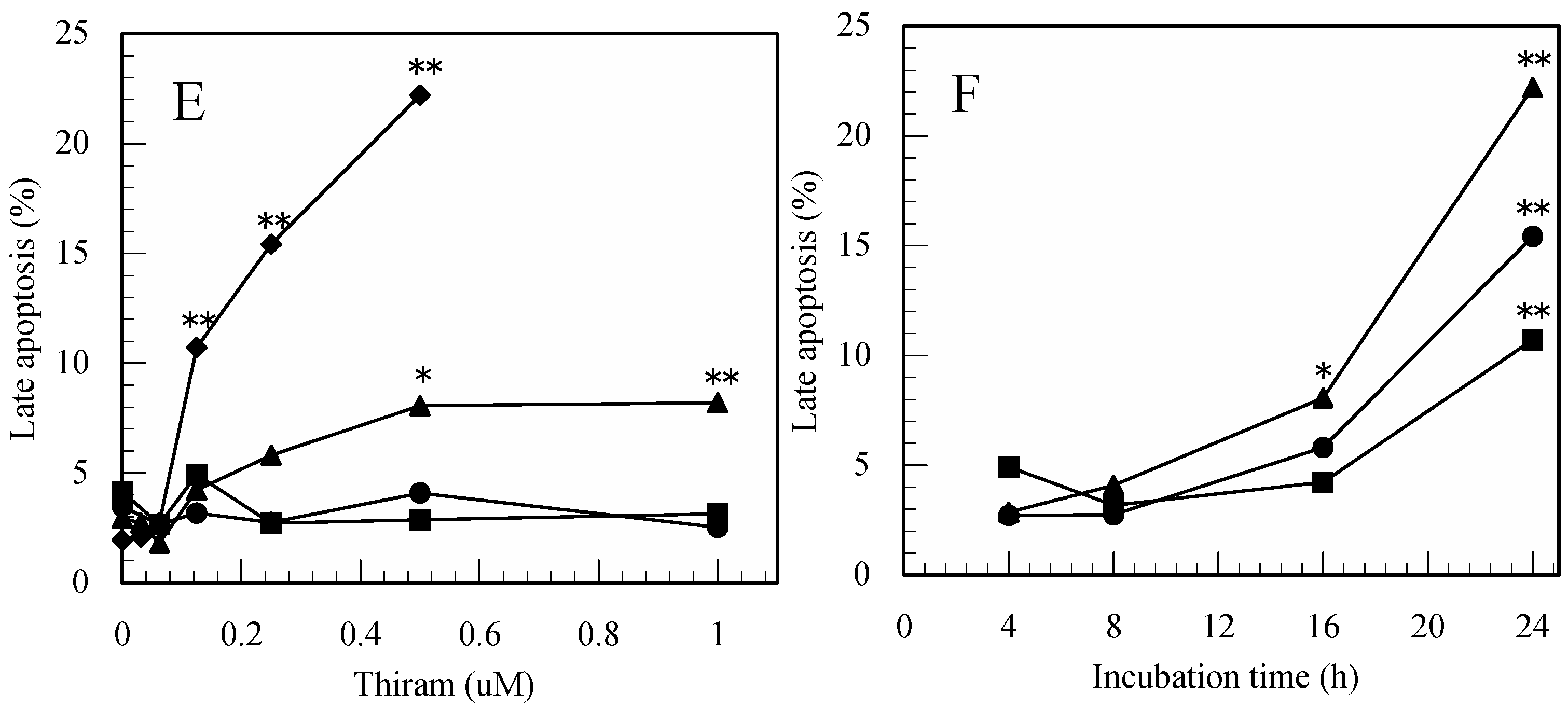
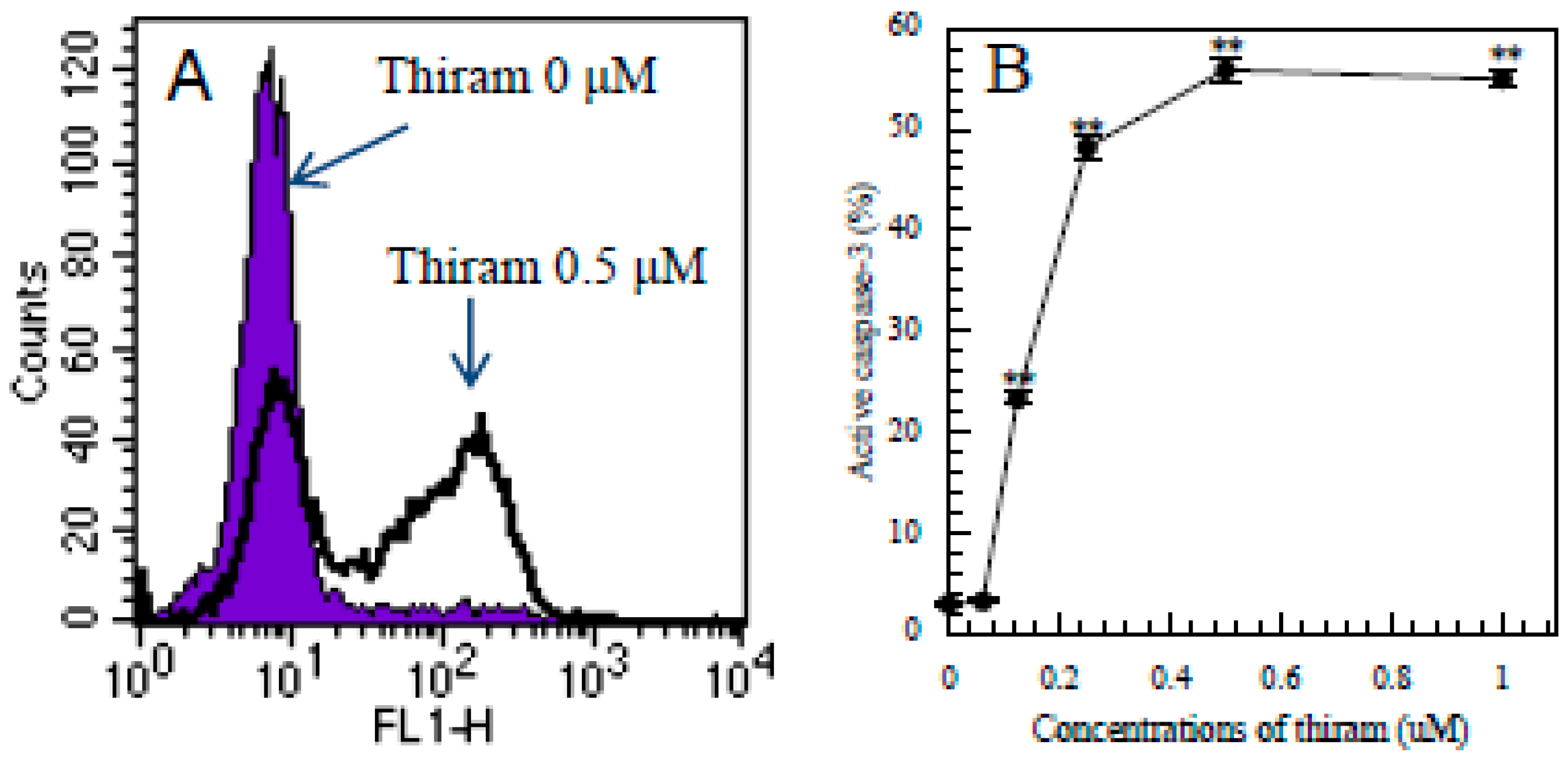
3.1. Thiram-Induced Apoptosis in Jurkat Cells Determined by FITC-Annexin-V/PI Staining
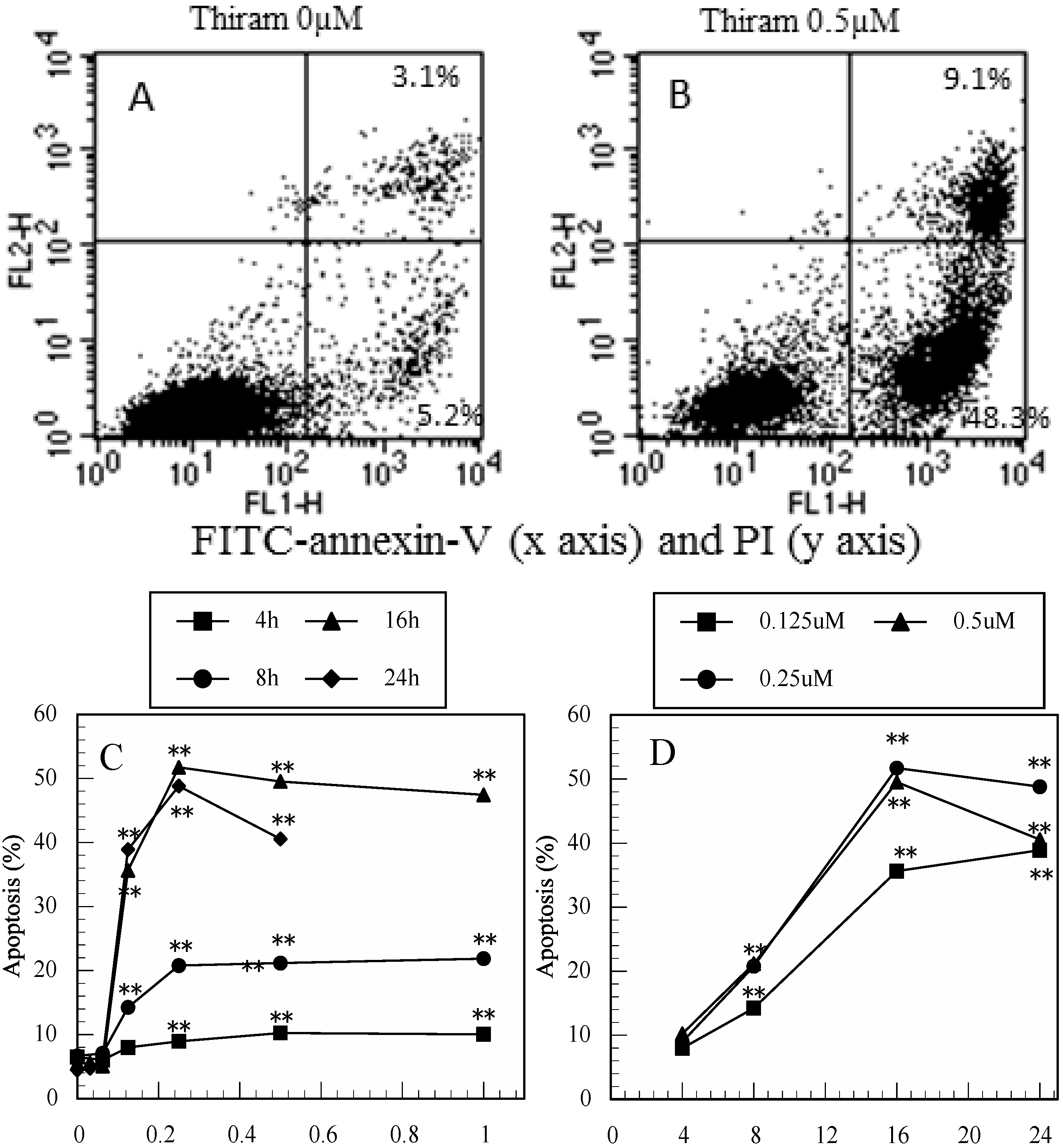
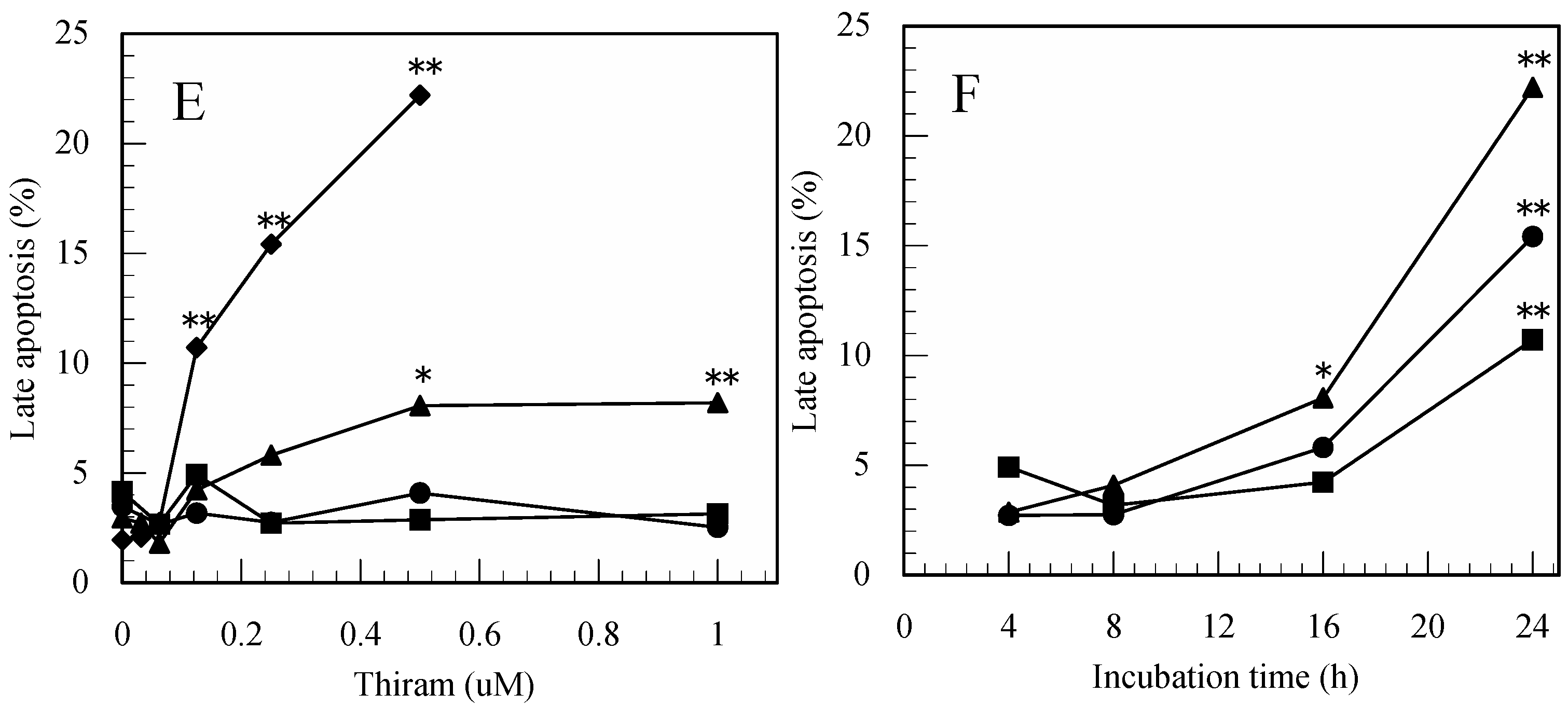
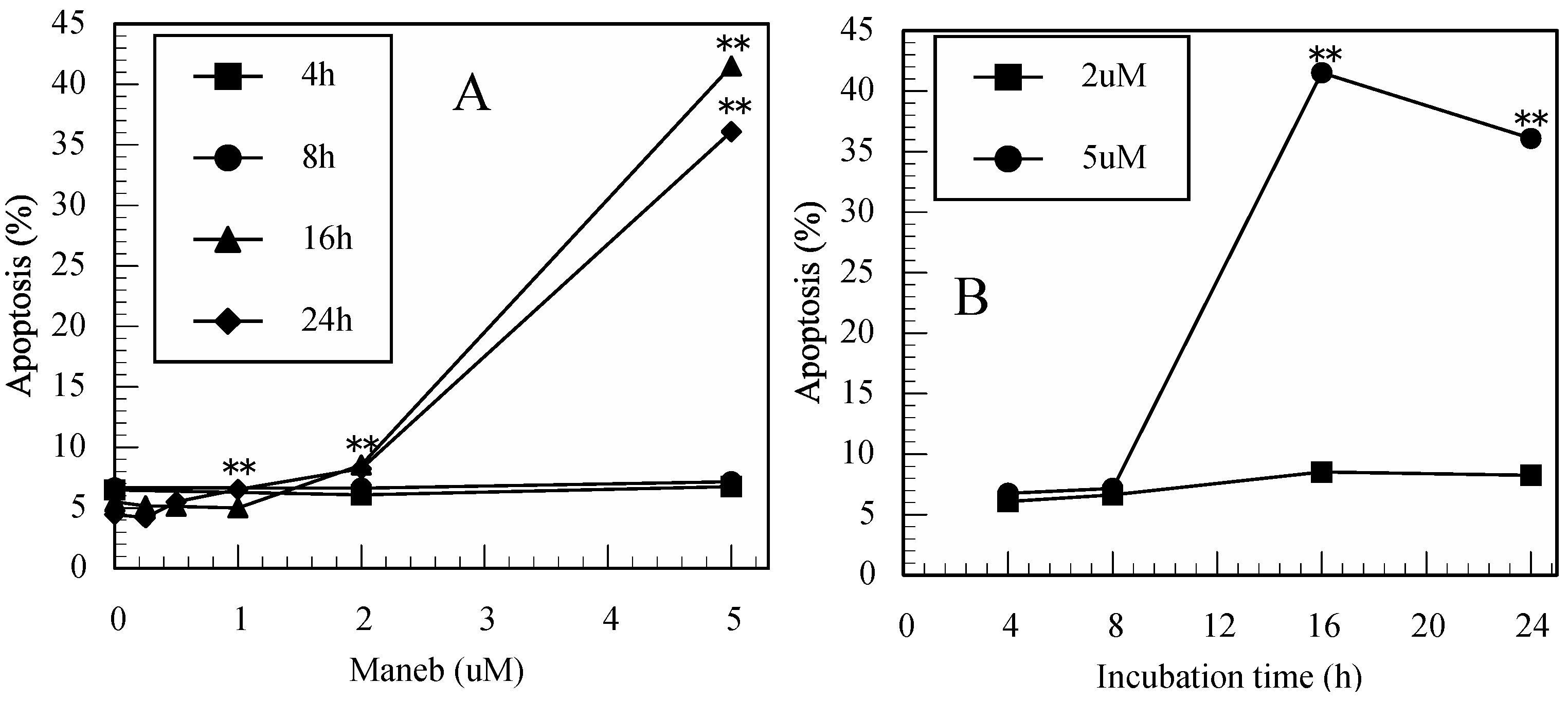
3.2. Maneb-Induced Apoptosis in Jurkat T Cells Determined by FITC-Annexin-V/PI Staining
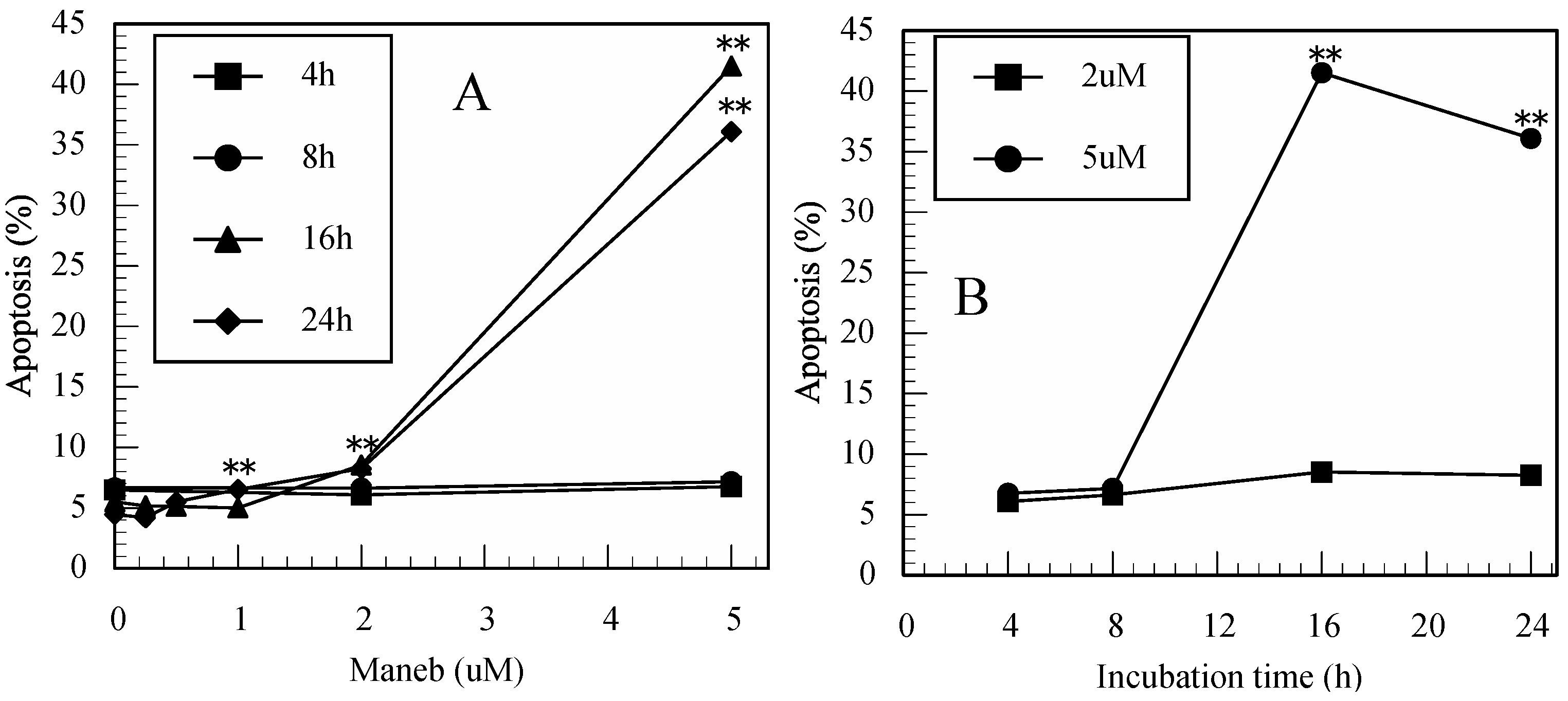
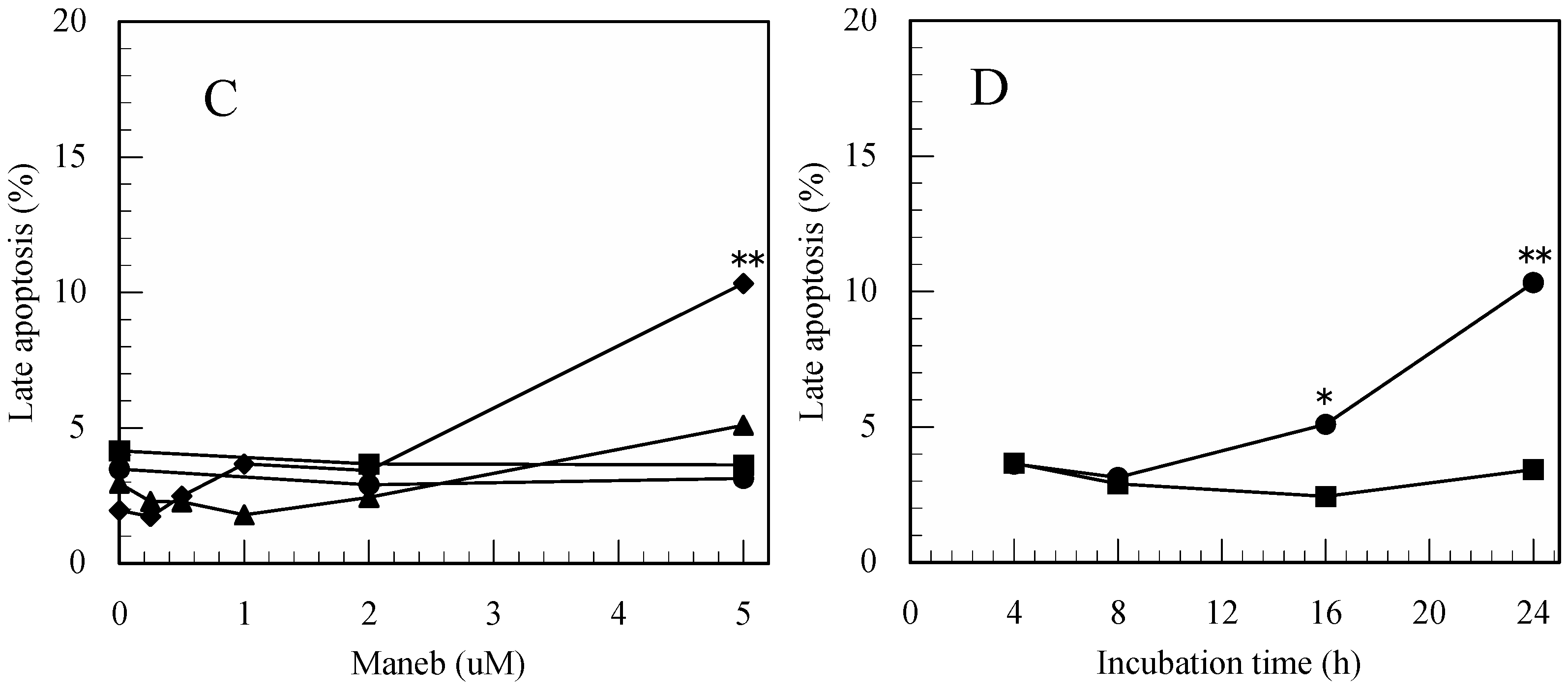
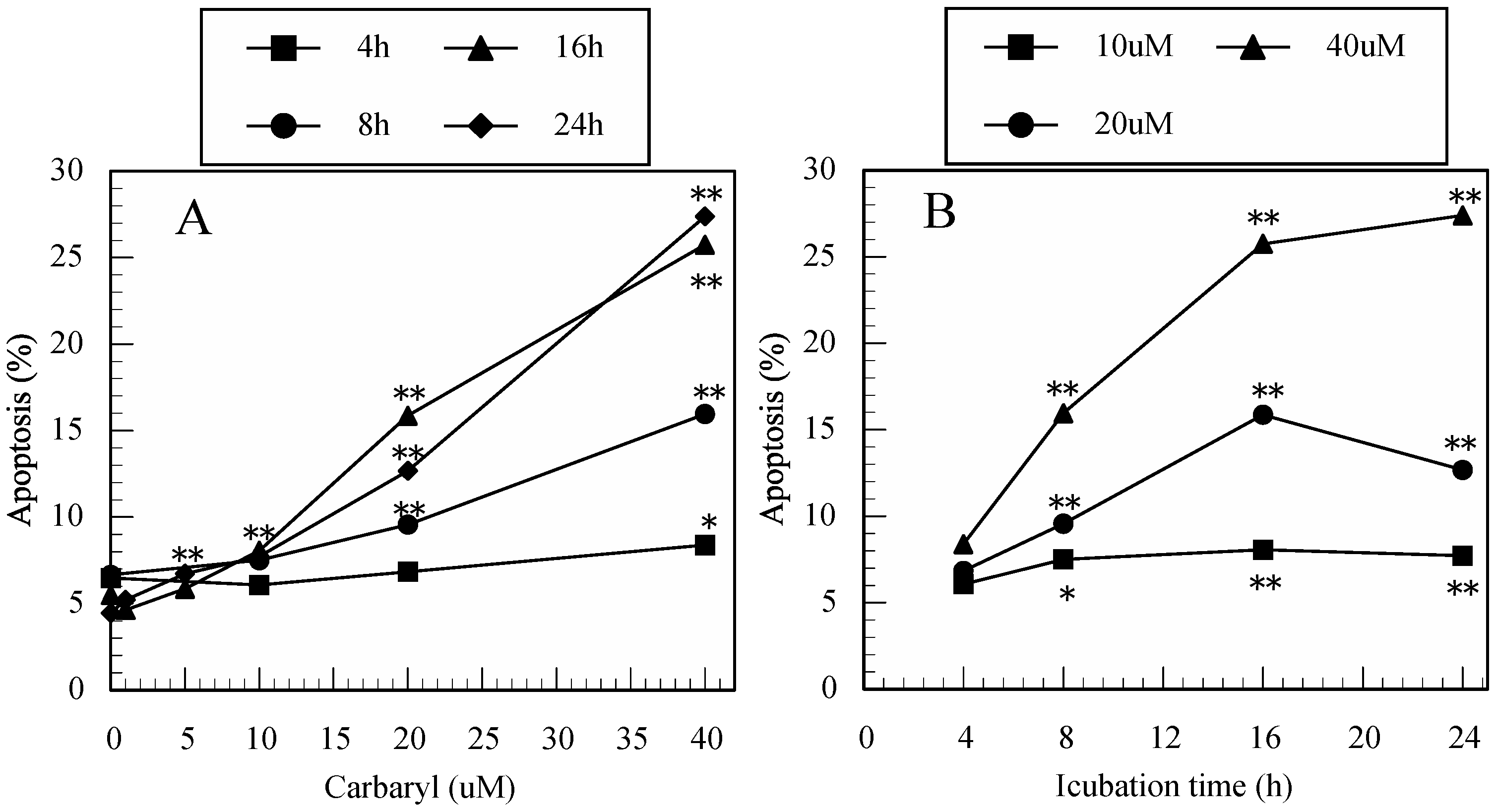
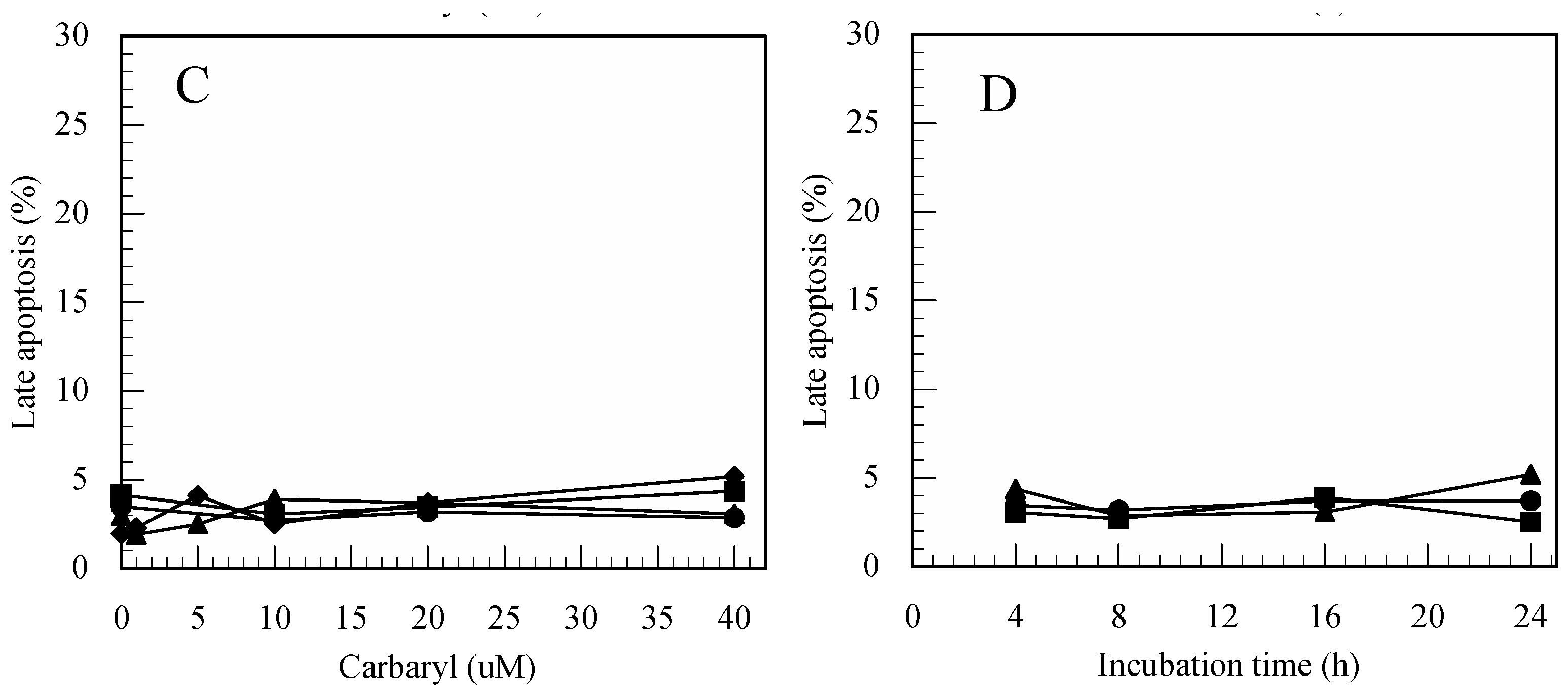
3.3. Carbaryl-Induced Apoptosis in Jurkat T Cells Determined by FITC-Annexin-V/PI Staining
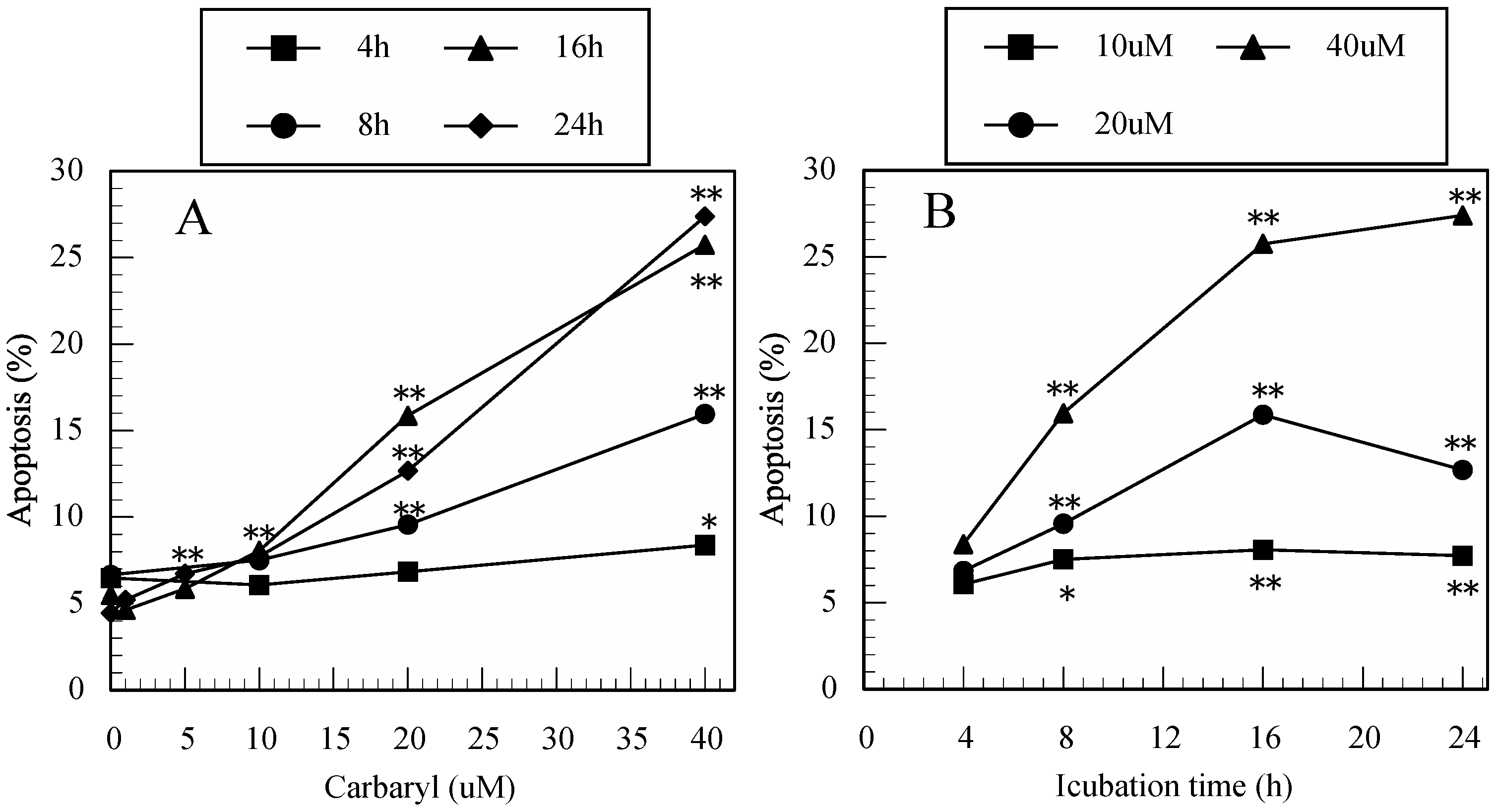
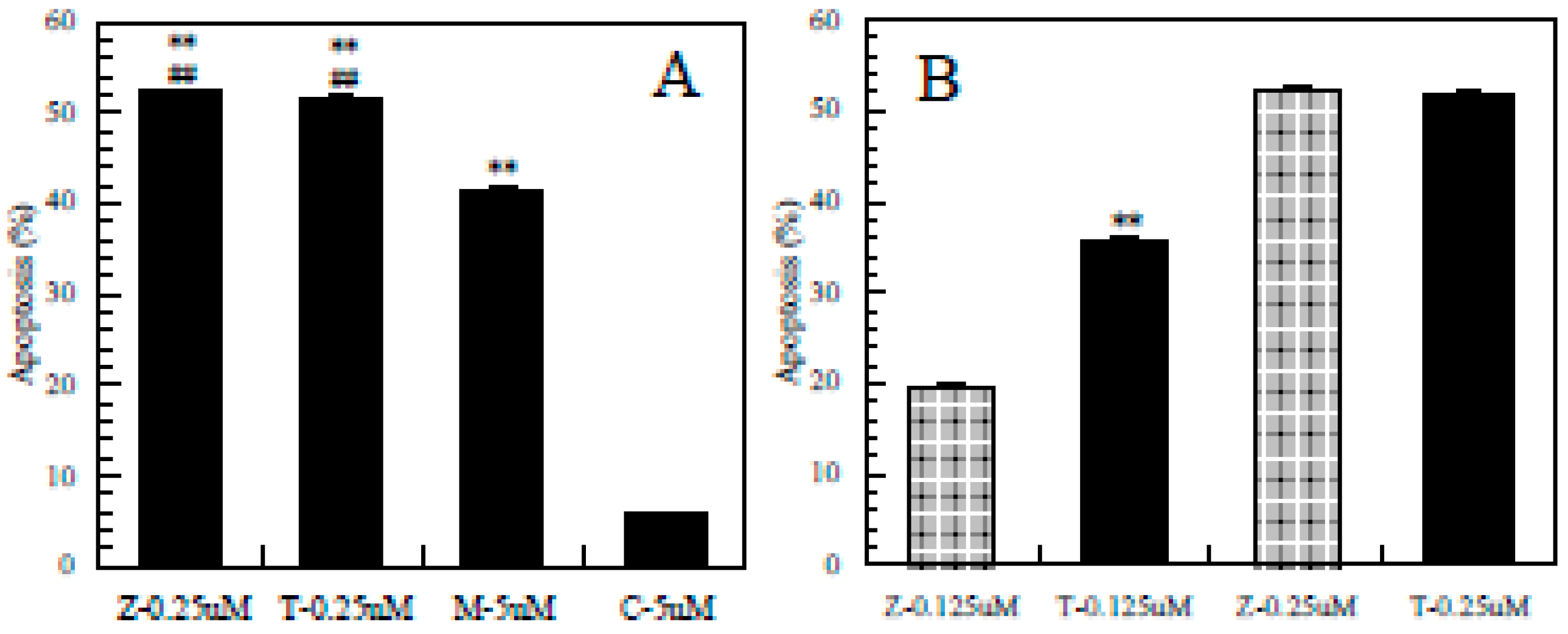
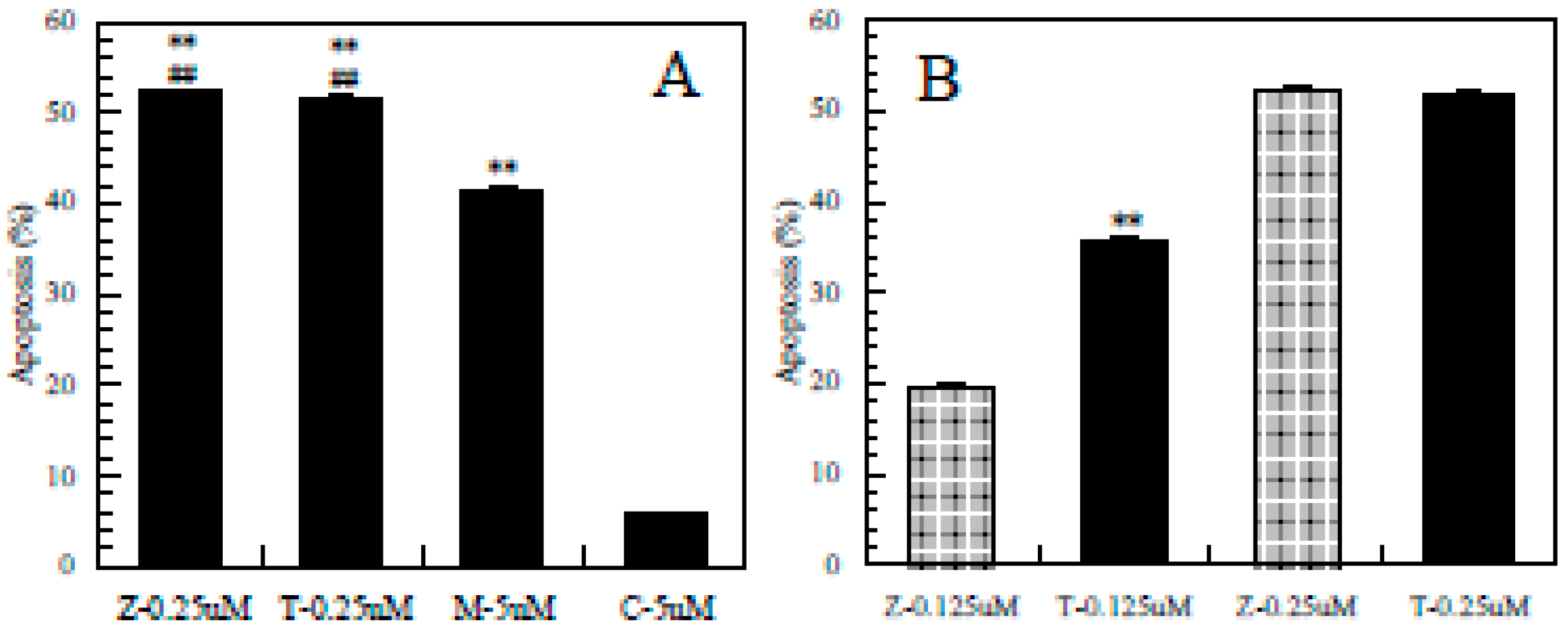
3.4. Difference in the Ability to Induce Apoptosis among the Pesticides
3.5. Detection of Intracellular Levels of Active Caspase-3 in Apoptotic Jurkat T Cells by Flow Cytometry
3.6. Protective Effects of Caspase-3 and General Caspase Inhibitors on Thiram-Induced Apoptosis in Jurkat T Cells
3.7. Detection of Mitochondrial Cytochrome-c Release in Apoptotic Jurkat T Cells by Flow Cytometry

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Variations | Degrees of Freedom | Variances | Observed Variance Ratios | p Values |
|---|---|---|---|---|---|
| Period of treatment (h) | 10.12 | 1 | 10.12 | 10.11 | 0.004 |
| Concentration of thiram (µM) | 638.74 | 5 | 127.75 | 127.64 | 0.000 |
| Interactions | 36.28 | 5 | 7.26 | 7.25 | 0.000 |
| Errors | 24.02 | 24 | 1.00 | ||
| Total | 709.17 | 35 |
4. Discussion
5. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Richardson, M.L. C62: Carbaryl. In The Dictionary of Substances and Their Effects; The Royal Society of Chemistry: Cambridge, UK, 1993; Volume 2, pp. 99–102. [Google Scholar]
- Richardson, M.L. M19: Maneb. In The Dictionary of Substances and Their Effects; The Royal Society of Chemistry: Cambridge, UK, 1993; Volume 5, pp. 323–327. [Google Scholar]
- Richardson, M.L. T130: Thiram. In The Dictionary of Substances and Their Effects; The Royal Society of Chemistry: Cambridge, UK, 1993; Volume 7, pp. 437–442. [Google Scholar]
- Richardson, M.L. Z19: Ziram. In The Dictionary of Substances and Their Effects; The Royal Society of Chemistry: Cambridge, UK, 1993; Volume 7, pp. 908–911. [Google Scholar]
- McDuffie, H.H.; Pahwa, P.; McLaughlin, J.R.; Spinelli, J.J.; Fincham, S.; Dosman, J.A.; Robson, D.; Skinnider, L.F.; Choi, N.W. Non-Hodgkin’s lymphoma and specific pesticide exposures in men: Cross-Canada study of pesticides and health. Cancer Epidemiol. Biomarkers Prev. 2001, 10, 1155–1163. [Google Scholar] [PubMed]
- Li, Q.; Kobayashi, M.; Kawada, T. Effect of ziram on natural killer, lymphokine-activated killer, and cytotoxic T lymphocyte activity. Arch. Toxicol. 2012, 86, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Kobayashi, M.; Kawada, T. Effect of carbamate pesticides on perforin, granzymes A-B-3/K, and granulysin in human natural killer cells. Int. J. Immunopathol. Pharmacol. 2015. (In press) [Google Scholar]
- Li, Q.; Kobayashi, M.; Kawada, T. Ziram induces apoptosis and necrosis in human immune cells. Arch. Toxicol. 2011, 85, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Kobayashi, M.; Kawada, T. Mechanism of ziram-induced apoptosis in human T lymphocytes. Arch. Toxicol. 2012, 86, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Kobayashi, M.; Kawada, T. Mechanism of ziram-induced apoptosis in human natural killer cells. Int. J. Immunopathol. Pharmacol. 2012, 25, 883–891. [Google Scholar] [PubMed]
- Li, Q.; Kobayashi, M.; Kawada, T. Carbamate pesticide-induced apoptosis and necrosis in human natural killer cells. J. Biol. Regul. Homeost. Agents 2014, 28, 23–32. [Google Scholar] [PubMed]
- Kerr, J.F.; Wyllie, A.H.; Currie, A.R. Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Majno, G.; Joris, I. Apoptosis, oncosis, and necrosis. An overview of cell death. Am. J. Pathol. 1995, 146, 3–15. [Google Scholar] [PubMed]
- Patel, T.; Gores, G.J.; Kaufmann, S.H. The role of proteases during apoptosis. FASEB J. 1996, 10, 587–597. [Google Scholar] [PubMed]
- Stennicke, H.R.; Jurgensmeier, J.M.; Shin, H.; Deveraux, Q.; Woff, B.B.; Yang, X.; Zhou, Q.; Ellerby, H.M.; Ellerby, L.M.; Bredesen, D.; et al. Pro-caspase-3 is a major physiologic target of caspase-8. J. Biol. Chem. 1998, 273, 27084–27090. [Google Scholar] [CrossRef] [PubMed]
- Okada, S.; Li, Q.; Whitin, J.C.; Clayberger, C.; Krensky, A.M. Intracellular mediators of granulysin-induced cell death. J. Immunol. 2003, 171, 2556–2562. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.C.; Waterhouse, N.J.; Juin, P.; Evan, G.I.; Green, D.R. The coordinate release of cytochrome C during apoptosis is rapid, complete and kinetically invariant. Nat. Cell. Biol. 2000, 2, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Stahnke, K.; Mohr, A.; Liu, J.; Meyer, L.H.; Karawajew, L.; Debatin, K.M. Identification of deficient mitochondrial signaling in apoptosis resistant leukemia cells by flow cytometric analysis of intracellular cytochrome c, caspase-3 and apoptosis. Apoptosis 2004, 9, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Taylor, T.R.; Whalen, M.M. Effects of ziram on tumor-cell-binding capacity, cell-surface marker expression, and ATP levels of human natural killer cells. Cell. Biol. Toxicol. 2009, 25, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Trace-Standard Of Agricultural Chemicals. Available online: http://m5.ws001.squarestart.ne.jp/zaidan/agrdtl.php?a_inq=28700 (accessed on 13 February 2015).
- Vandekar, M.; Plestina, R.; Wilhelm, K. Toxicity of carbamates for mammals. Bull. World Health Organ. 1971, 44, 241–249. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Kobayashi, M.; Kawada, T. Carbamate Pesticide-Induced Apoptosis in Human T Lymphocytes. Int. J. Environ. Res. Public Health 2015, 12, 3633-3645. https://doi.org/10.3390/ijerph120403633
Li Q, Kobayashi M, Kawada T. Carbamate Pesticide-Induced Apoptosis in Human T Lymphocytes. International Journal of Environmental Research and Public Health. 2015; 12(4):3633-3645. https://doi.org/10.3390/ijerph120403633
Chicago/Turabian StyleLi, Qing, Maiko Kobayashi, and Tomoyuki Kawada. 2015. "Carbamate Pesticide-Induced Apoptosis in Human T Lymphocytes" International Journal of Environmental Research and Public Health 12, no. 4: 3633-3645. https://doi.org/10.3390/ijerph120403633
APA StyleLi, Q., Kobayashi, M., & Kawada, T. (2015). Carbamate Pesticide-Induced Apoptosis in Human T Lymphocytes. International Journal of Environmental Research and Public Health, 12(4), 3633-3645. https://doi.org/10.3390/ijerph120403633