
Induction of the Estrogenic Marker Calbindn-D9k by Octamethylcyclotetrasiloxane



Abstract
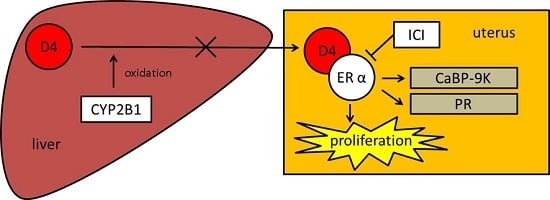
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Experimental Section
2.1. Chemicals
2.2. GH3 Cells
2.3. Animals
2.4. UT Assay and Liver/Body Weight Ratio Measurement
2.5. Quantitative Real-Time qPCR
2.6. Western Blotting
2.7. Statistical Analyses
3. Results
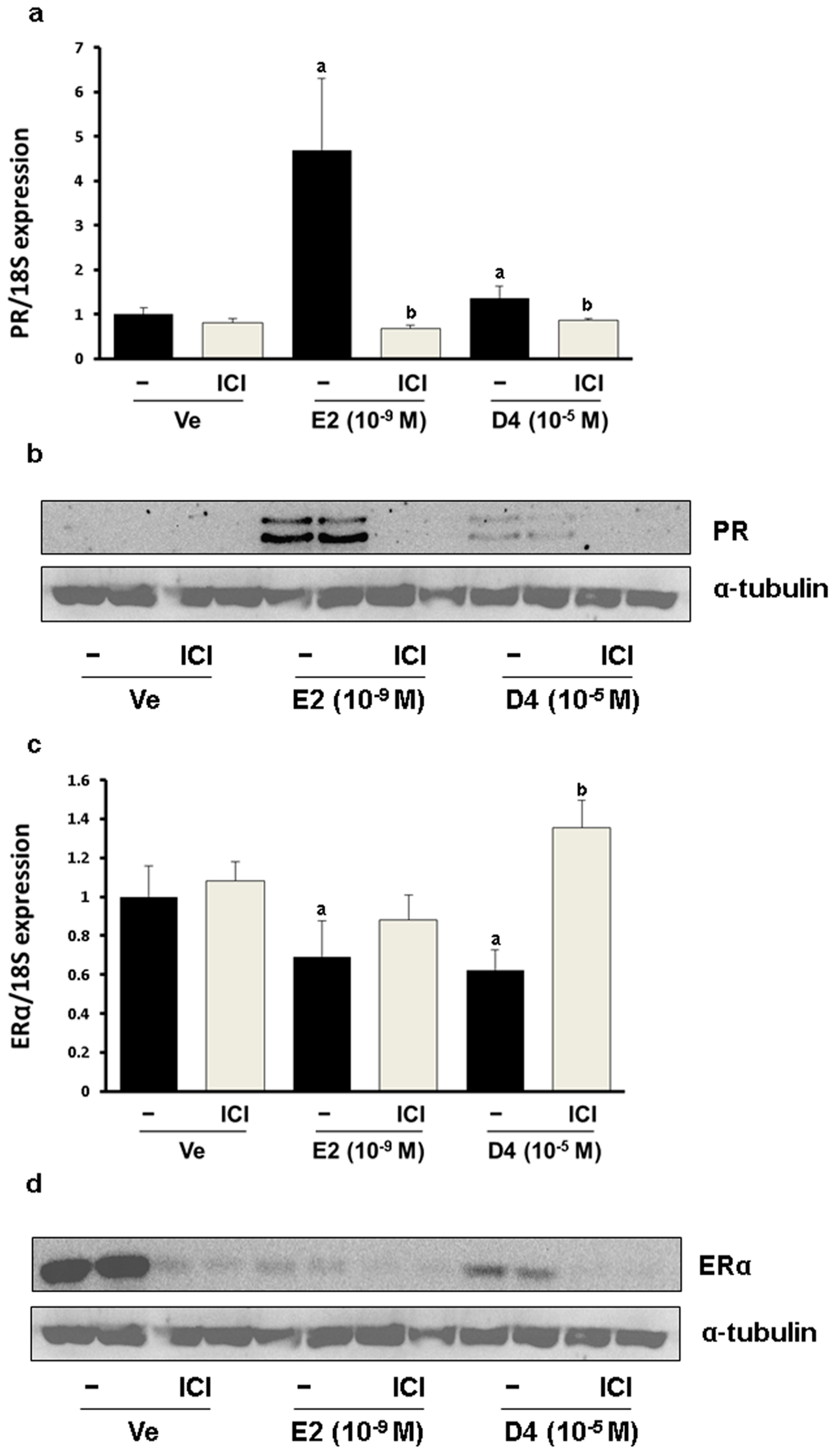
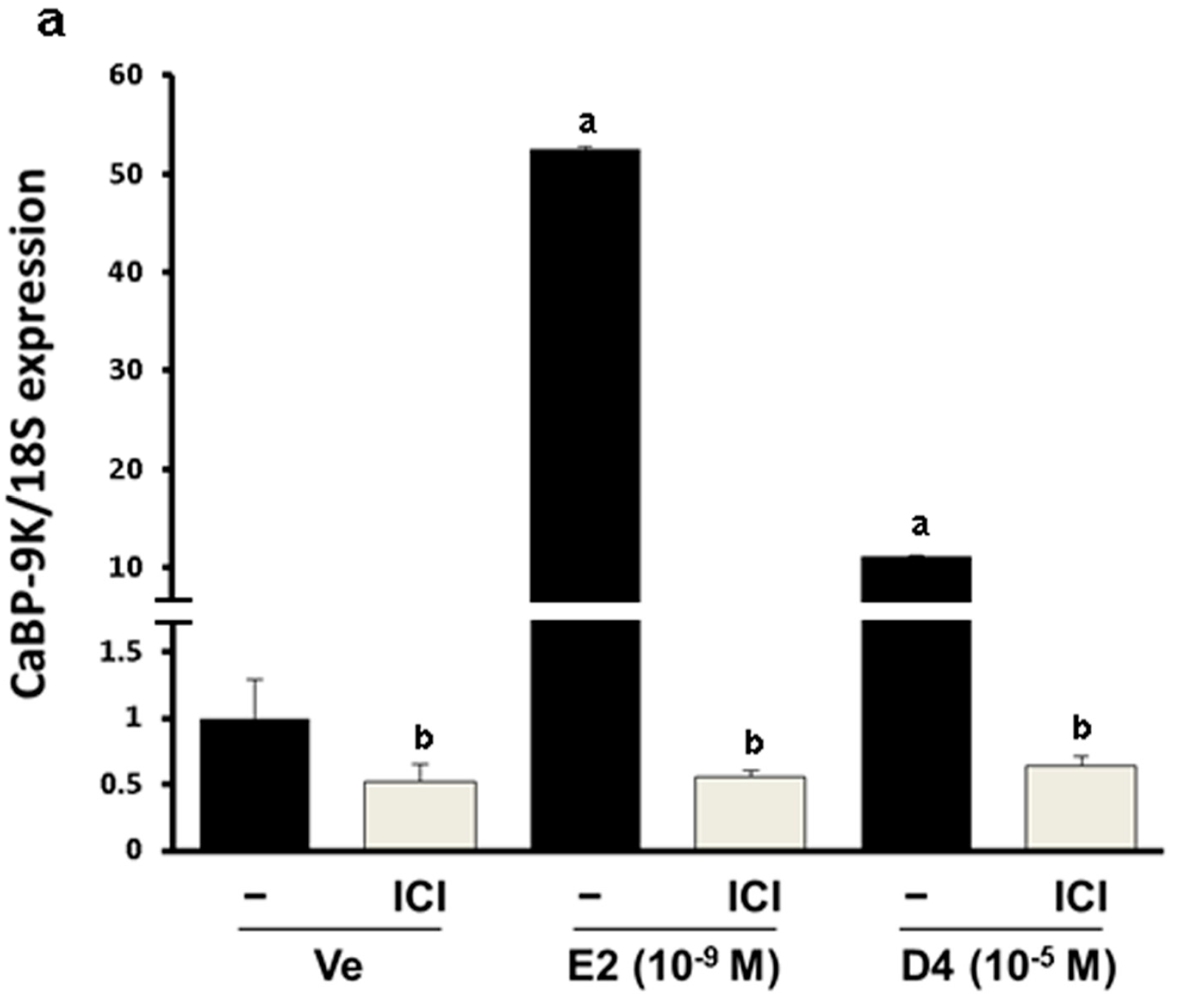
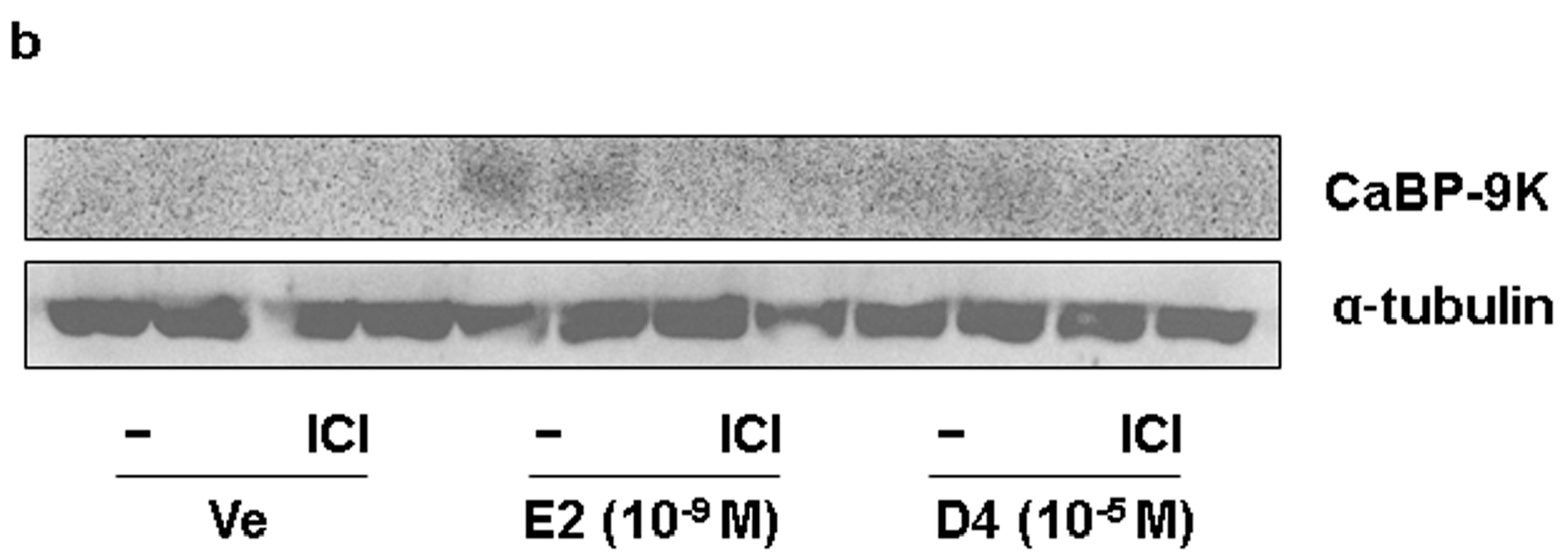
3.1. CaBP-9K, ER α, and PR Expression in GH3 Cells

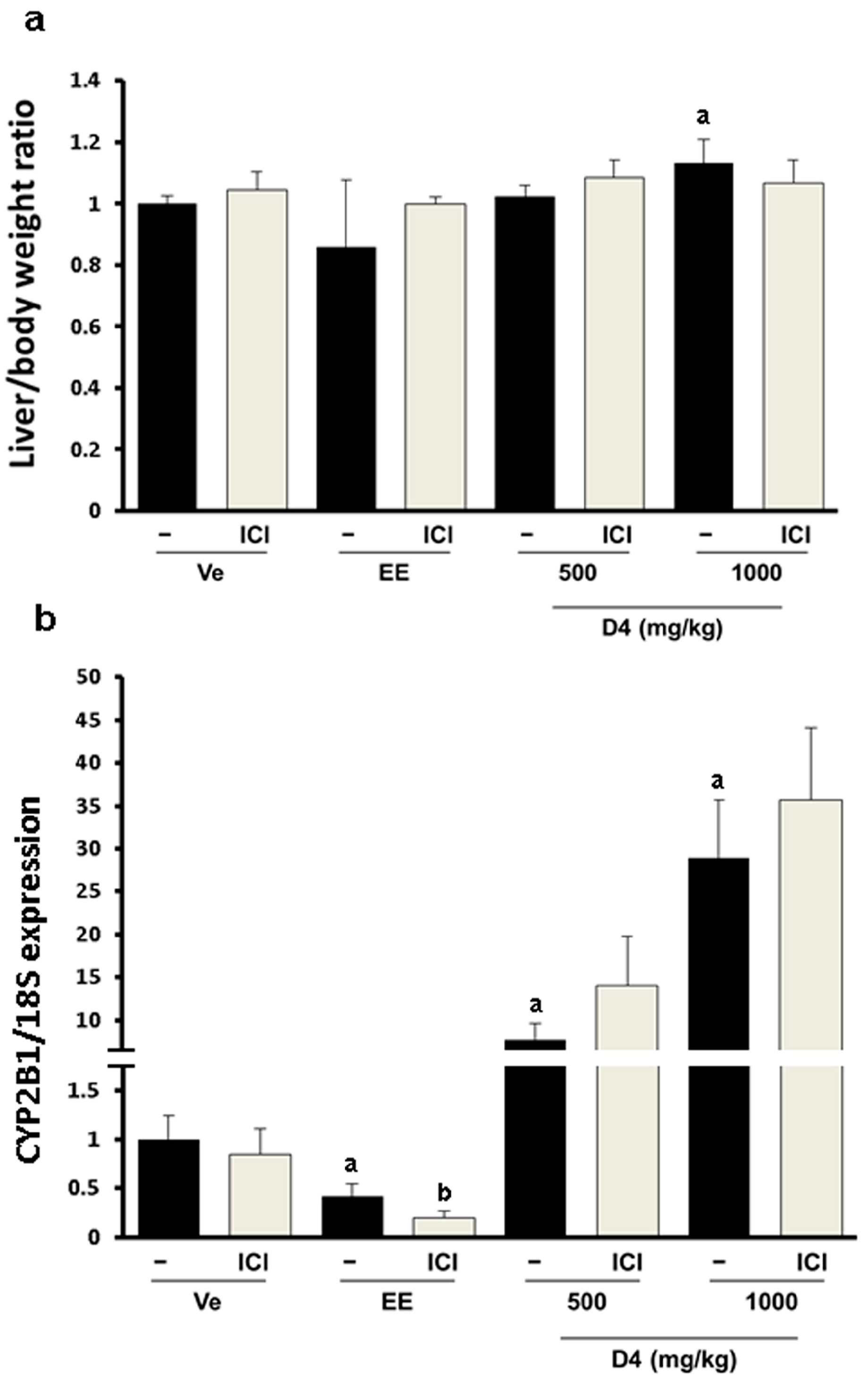
3.2. Relative Weight Ratio and CYP2B1/2 Expression in Immature Rat Liver
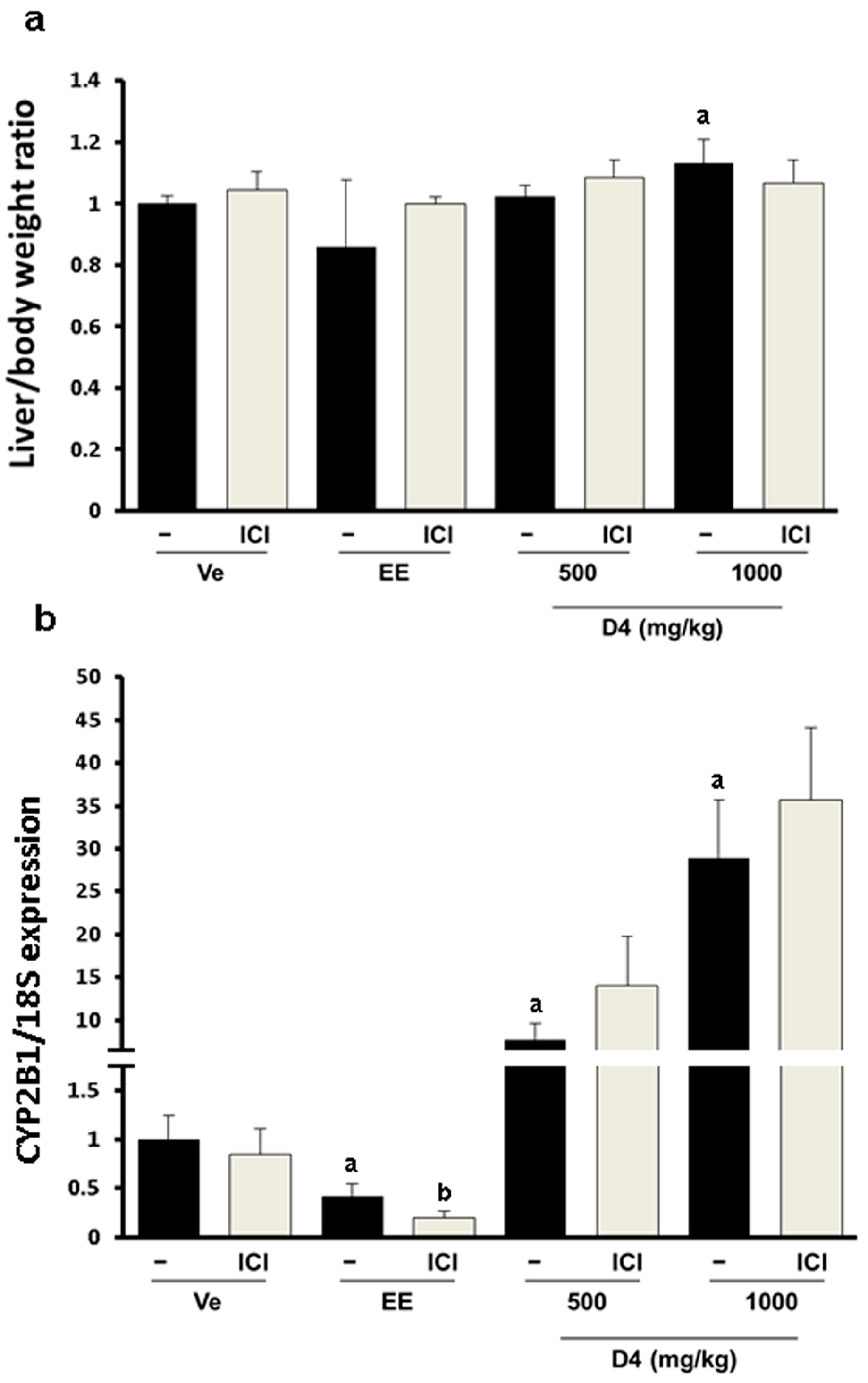
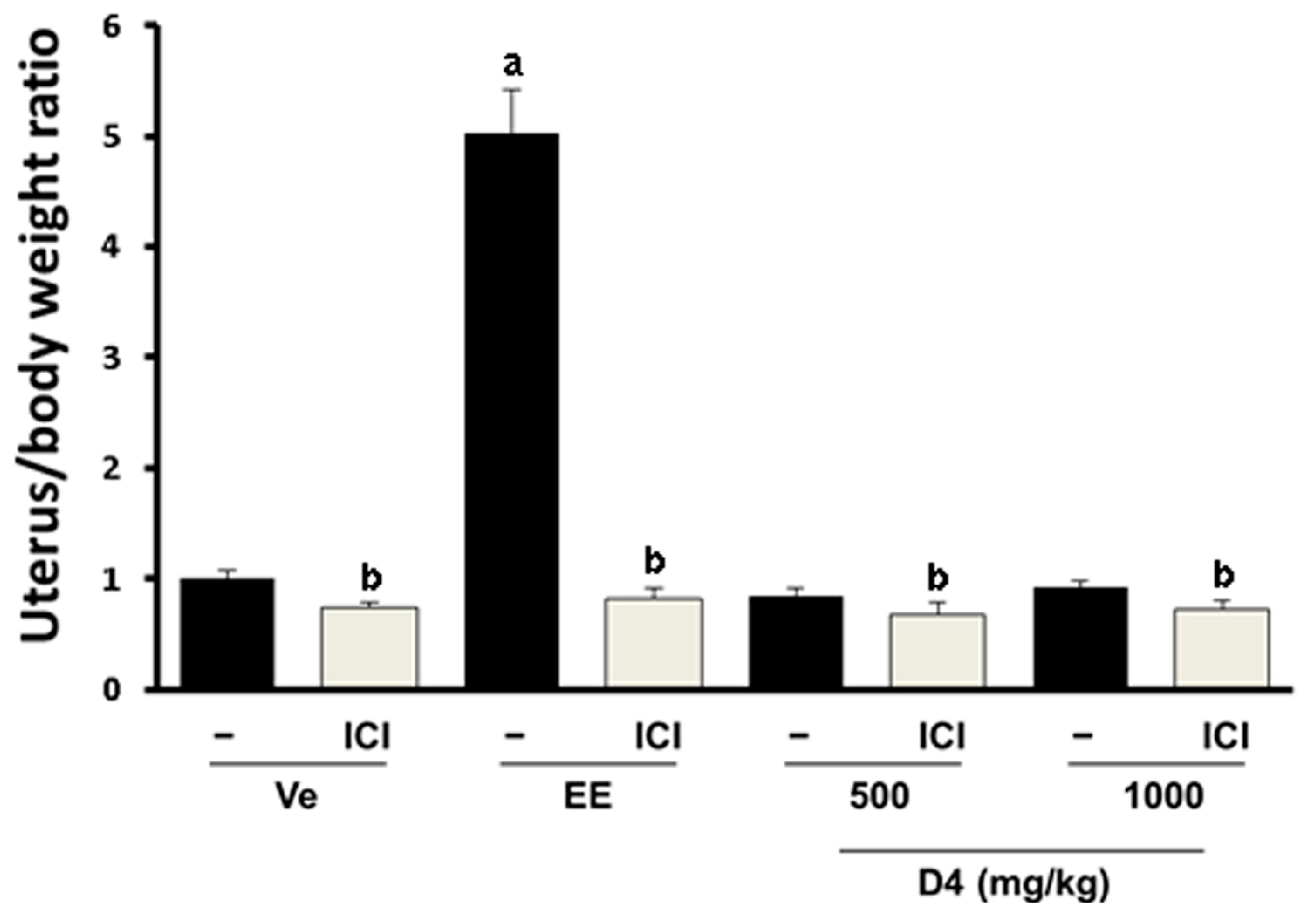
3.3. UT Assay Results in Immature Rats

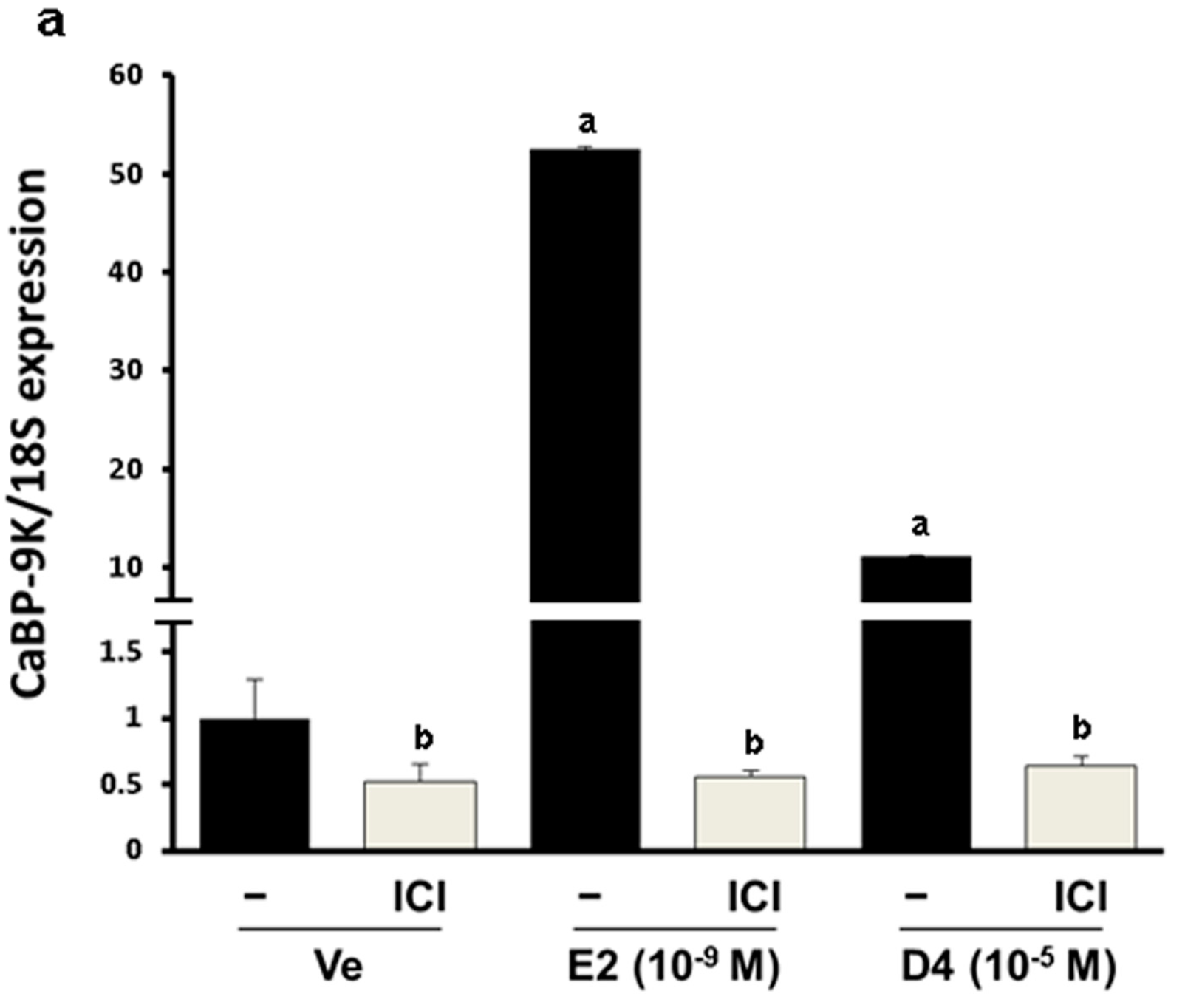
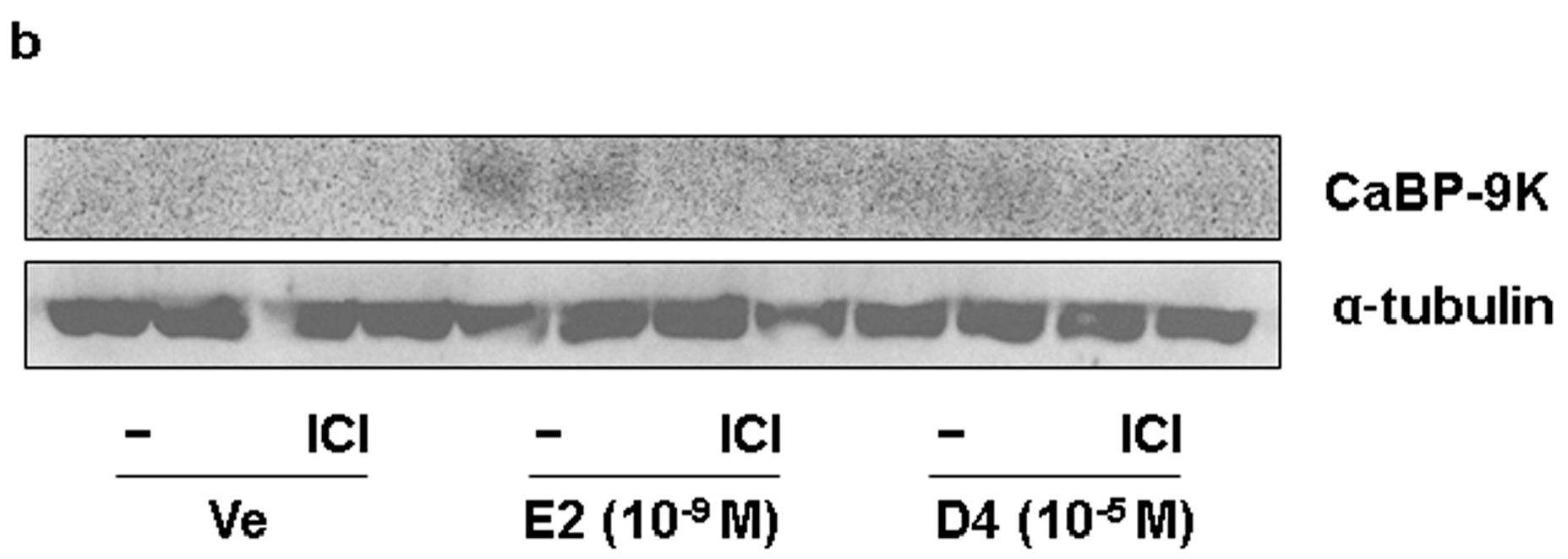
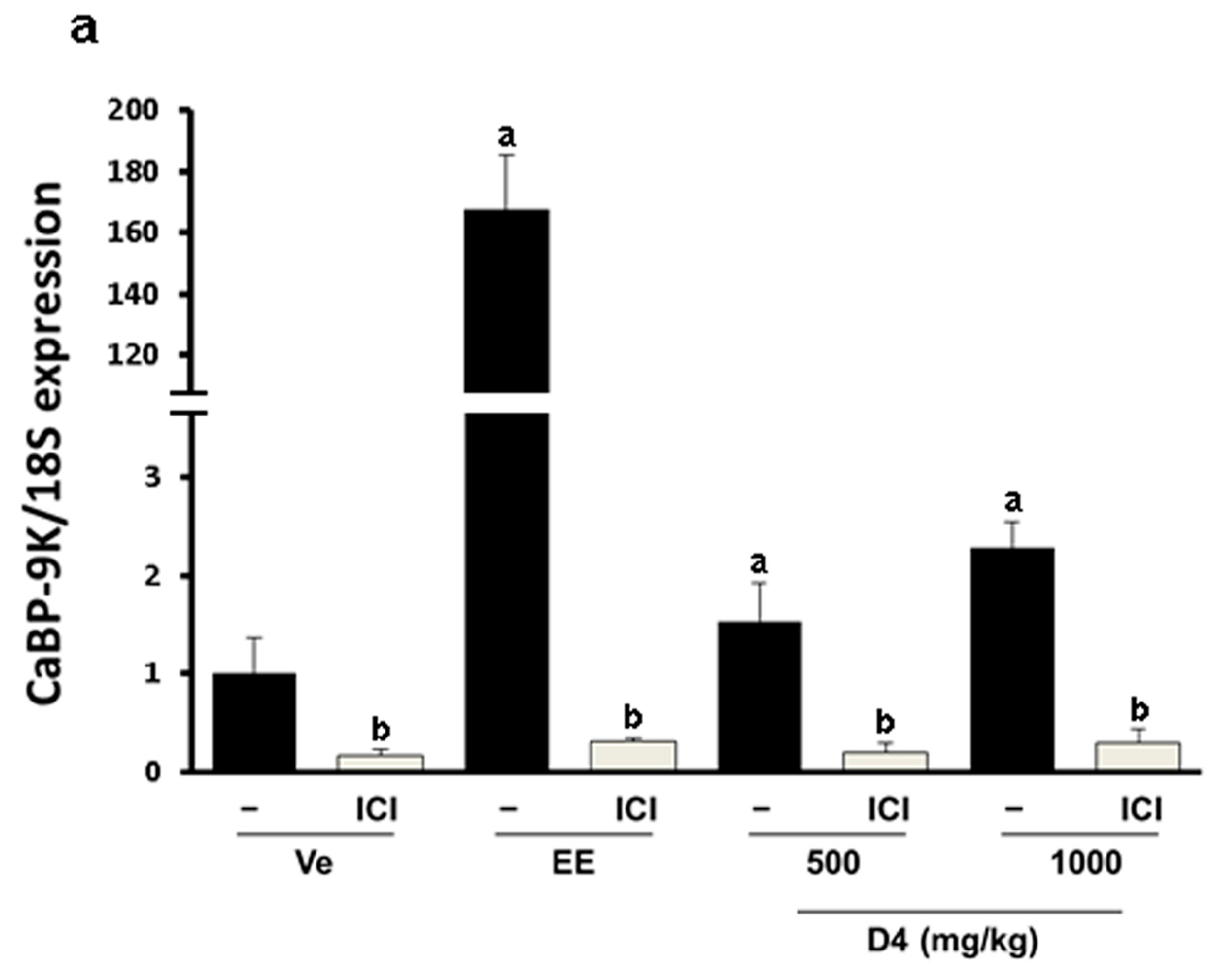
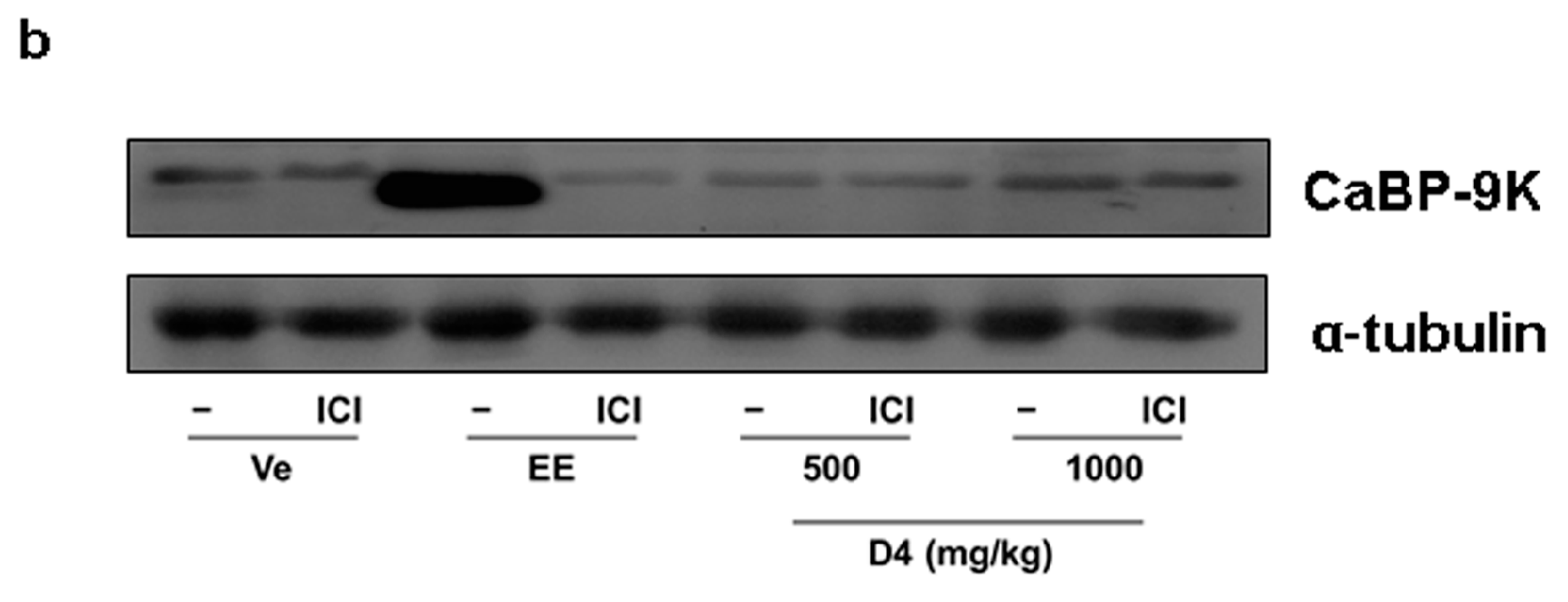
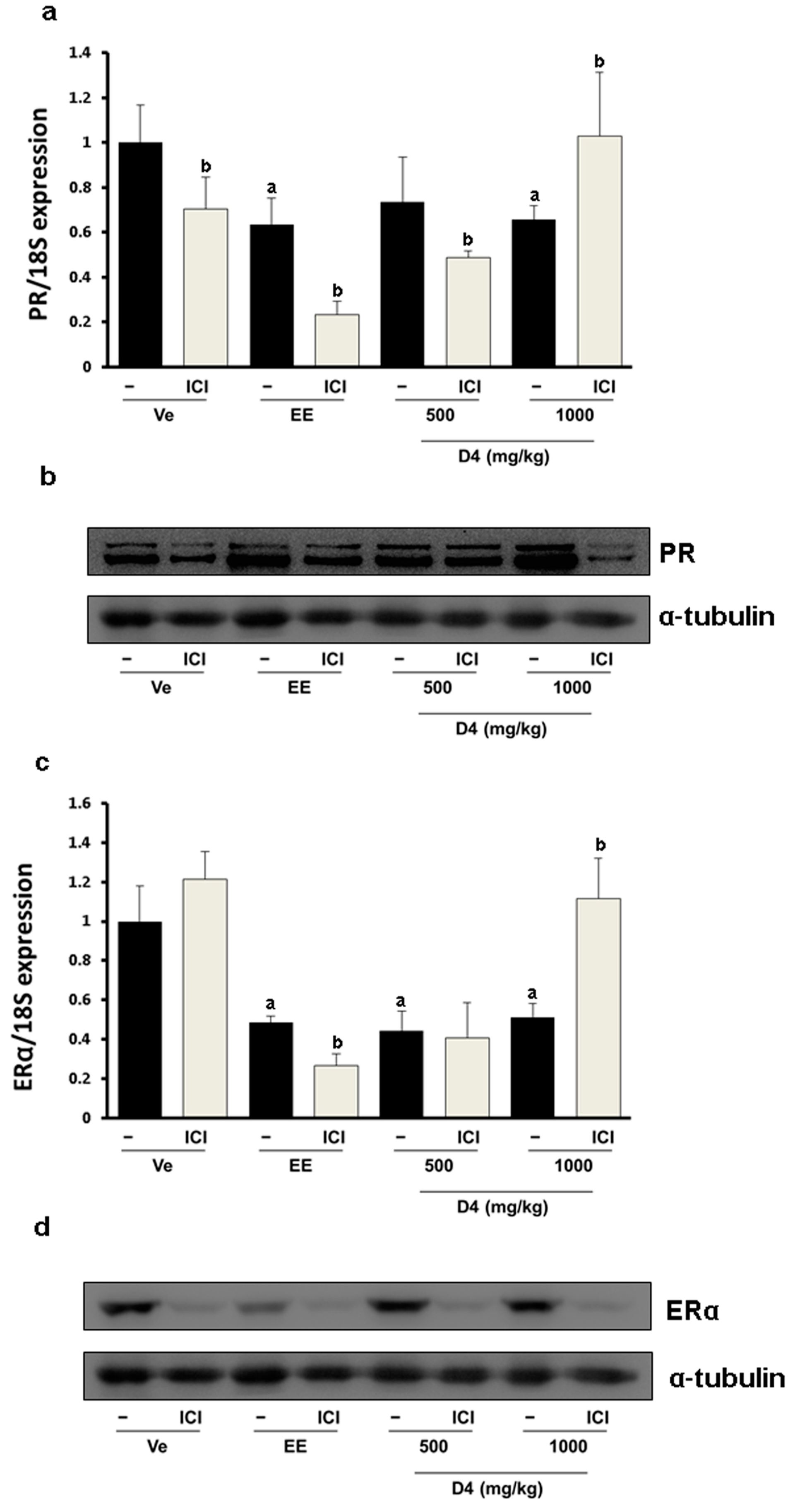
3.4. CaBP-9K, ER α, and PR Expression in Immature Rats Uterus
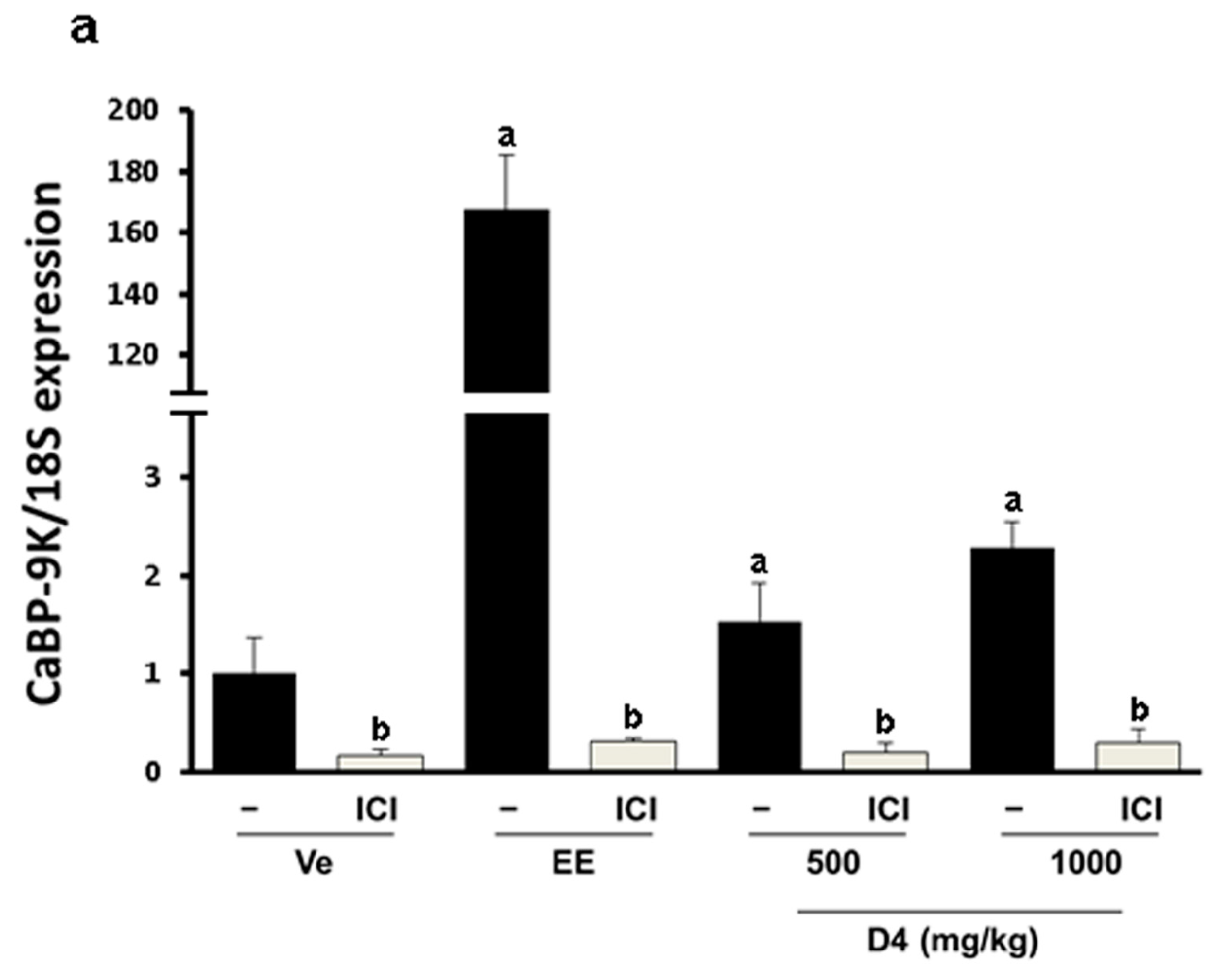
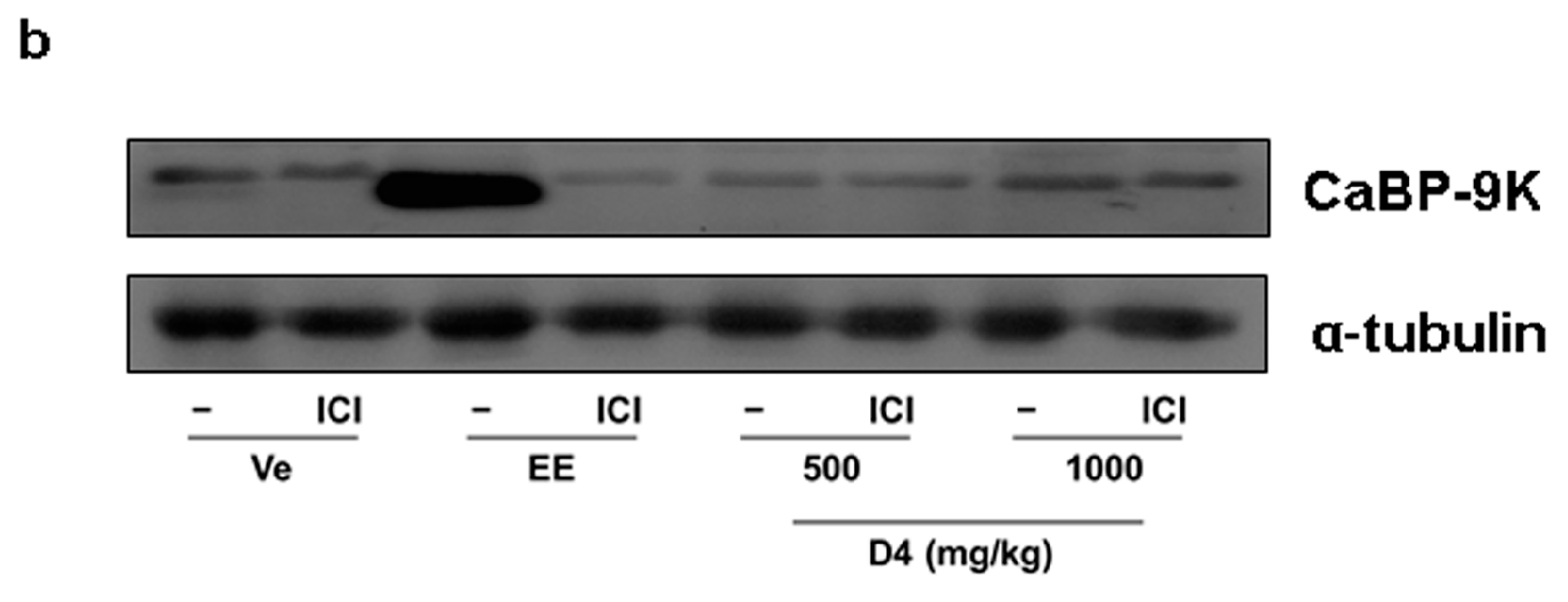
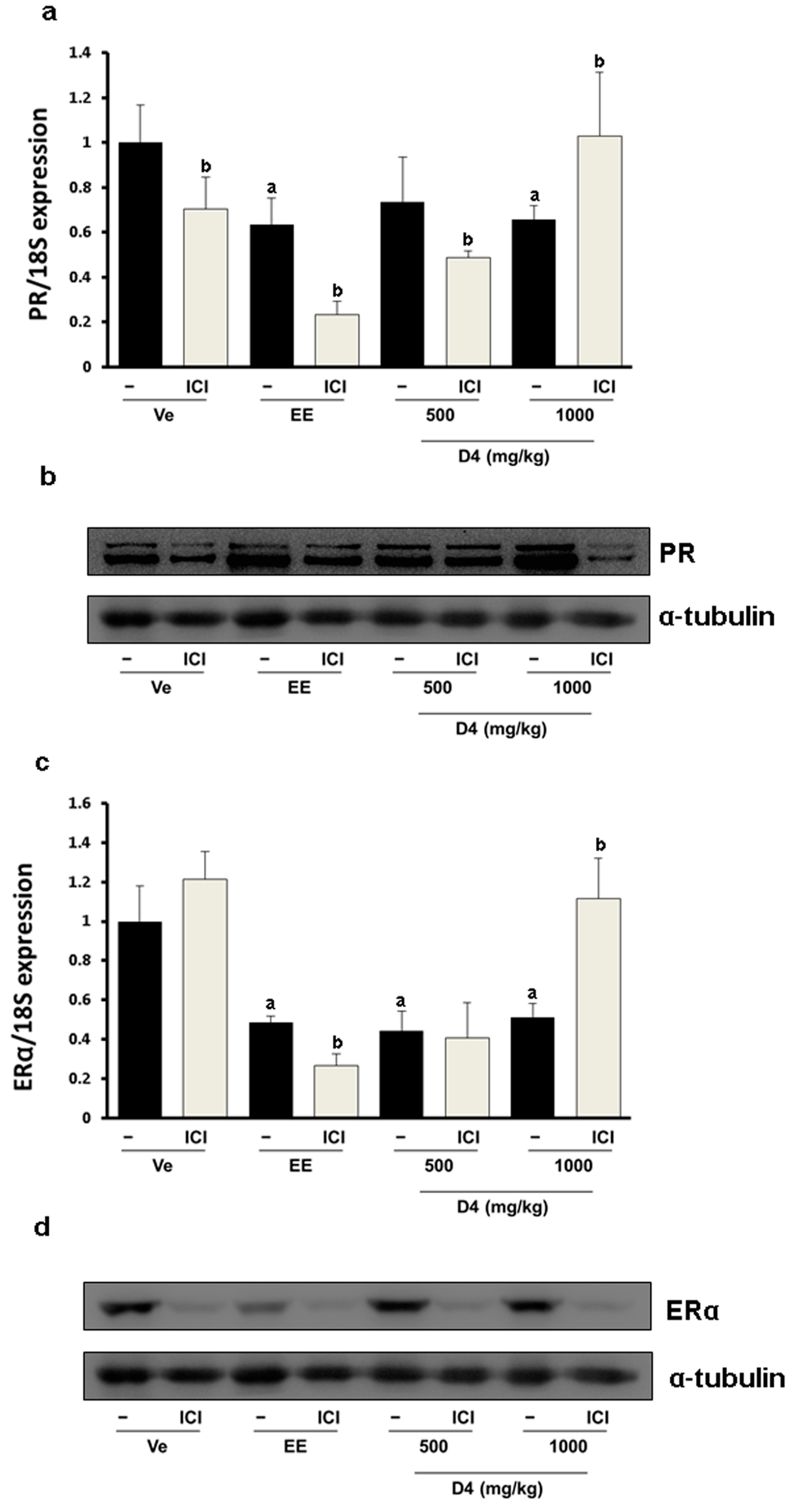
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mustonen, E.; Taponen, S.; Andersson, M.; Sukura, A.; Katila, T.; Taponen, J. Fertility and growth of nulliparous ewes after feeding red clover silage with high phyto-oestrogen concentrations. Animal 2014, 8, 1699–1705. [Google Scholar] [CrossRef] [PubMed]
- Diamanti-Kandarakis, E.; Bourguignon, J.P.; Giudice, L.C.; Hauser, R.; Prins, G.S.; Soto, A.M.; Zoeller, R.T.; Gore, A.C. Endocrine-disrupting chemicals: An endocrine society scientific statement. Endocr. Rev. 2009, 30, 293–342. [Google Scholar] [CrossRef] [PubMed]
- Hilakivi-Clarke, L. Maternal exposure to diethylstilbestrol during pregnancy and increased breast cancer risk in daughters. Breast Cancer Res. 2014, 16. [Google Scholar] [CrossRef]
- Kumar, E.; Holt, W.V. Impacts of endocrine disrupting chemicals on reproduction in wildlife. Adv. Exp. Med. Biol. 2014, 753, 55–70. [Google Scholar] [PubMed]
- Wan, H.T.; Mruk, D.D.; Wong, C.K.; Cheng, C.Y. Targeting testis-specific proteins to inhibit spermatogenesis: Lesson from endocrine disrupting chemicals. Expert Opin. Ther. Targets 2013, 17, 839–855. [Google Scholar] [CrossRef] [PubMed]
- Gross-Sorokin, M.Y.; Roast, S.D.; Brighty, G.C. Assessment of feminization of male fish in english rivers by the environment agency of england and wales. Environ. Health Perspect. 2006, 114, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Caserta, D.; di Segni, N.; Mallozzi, M.; Giovanale, V.; Mantovani, A.; Marci, R.; Moscarini, M. Bisphenol a and the female reproductive tract: An overview of recent laboratory evidence and epidemiological studies. Reprod. Biol. Endocrinol. 2014, 12. [Google Scholar] [CrossRef] [PubMed]
- Rutkowska, A.; Rachoń, D. Bisphenol a (BPA) and its potential role in the pathogenesis of the polycystic ovary syndrome (PCOS). Gynecol. Endocrinol. 2014, 30, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, K.; Tagami, T.; Akamizu, T.; Usui, T.; Saijo, M.; Kanamoto, N.; Hataya, Y.; Shimatsu, A.; Kuzuya, H.; Nakao, K. Thyroid hormone action is disrupted by bisphenol a as an antagonist. J. Clin. Endocrinol. Metab. 2002, 87, 5185–5190. [Google Scholar] [CrossRef] [PubMed]
- Hultman, M.T.; Rundberget, J.T.; Tollefsen, K.E. Evaluation of the sensitivity, responsiveness and reproducibility of primary rainbow trout hepatocyte vitellogenin expression as a screening assay for estrogen mimics. Aquat. Toxicol. 2015, 159, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Wanda, G.J.; Starcke, S.; Zierau, O.; Njamen, D.; Richter, T.; Vollmer, G. Estrogenic activity of griffonianone C, an isoflavone from the root bark of millettia griffoniana: Regulation of the expression of estrogen responsive genes in uterus and liver of ovariectomized rats. Planta Med. 2007, 73, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.M.; An, B.S.; Choi, K.C.; Jeung, E.B. Potential estrogenic activity of triclosan in the uterus of immature rats and rat pituitary GH3 cells. Toxicol. Lett. 2012, 208, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, S.I.; Maznah, I.; Mahmud, R.B.; Saeed, M.I.; Imam, M.U.; Ishaka, A. Estrogen receptor modulatory effects of germinated brown rice bioactives in the uterus of rats through the regulation of estrogen-induced genes. Drug Des. Devel. Ther. 2013, 7, 1409–1420. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.S.; Jung, E.M.; Choi, K.C.; Oh, G.T.; Jeung, E.B. Compensatory induction of the TRPV6 channel in a calbindin-D9k knockout mouse: Its regulation by 1,25-hydroxyvitamin D3. J. Cell. Biochem. 2009, 108, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Dang, V.H.; Nguyen, T.H.; Choi, K.C.; Jeung, E.B. A calcium-binding protein, calbindin-D9k, is regulated through an estrogen-receptor mediated mechanism following xenoestrogen exposure in the GH3 cell line. Toxicol. Sci. 2007, 98, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Horii, Y.; Kannan, K. Survey of organosilicone compounds, including cyclic and linear siloxanes, in personal-care and household products. Arch. Environ. Contam. Toxicol. 2008, 55, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Dudzina, T.; von Goetz, N.; Bogdal, C.; Biesterbos, J.W.; Hungerbühler, K. Concentrations of cyclic volatile methylsiloxanes in european cosmetics and personal care products: Prerequisite for human and environmental exposure assessment. Environ. Int. 2014, 62, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Quinn, A.L.; Regan, J.M.; Tobin, J.M.; Marinik, B.J.; McMahon, J.M.; McNett, D.A.; Sushynski, C.M.; Crofoot, S.D.; Jean, P.A.; Plotzke, K.P. In vitro and in vivo evaluation of the estrogenic, androgenic, and progestagenic potential of two cyclic siloxanes. Toxicol. Sci. 2007, 96, 145–153. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Rhodes-Brower, S.; Miller, M.R.; Munson, A.E.; Germolec, D.R.; Walker, V.R.; Korach, K.S.; Meade, B.J. Octamethylcyclotetrasiloxane exhibits estrogenic activity in mice via ER α. Toxicol. Appl. Pharmacol. 2003, 192, 254–261. [Google Scholar] [CrossRef]
- Korzekwa, K. Enzyme kinetics of oxidative metabolism: Cytochromes p450. Methods Mol. Biol. 2014, 1113, 149–166. [Google Scholar] [PubMed]
- Zhong, Y.; Dong, G.; Luo, H.; Cao, J.; Wang, C.; Wu, J.; Feng, Y.Q.; Yue, J. Induction of brain CYP2E1 by chronic ethanol treatment and related oxidative stress in hippocampus, cerebellum, and brainstem. Toxicology 2012, 302, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Ohno, M.; Motojima, K.; Okano, T.; Taniguchi, A. Induction of drug-metabolizing enzymes by phenobarbital in layered co-culture of a human liver cell line and endothelial cells. Biol. Pharm. Bull. 2009, 32, 813–817. [Google Scholar] [CrossRef] [PubMed]
- Meucci, V.; Arukwe, A. The xenoestrogen 4-nonylphenol modulates hepatic gene expression of pregnane x receptor, aryl hydrocarbon receptor, CYP3A and CYP1A1 in juvenile atlantic salmon (salmo salar). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 142, 142–150. [Google Scholar] [CrossRef] [PubMed]
- McKim, J.M.; Wilga, P.C.; Kolesar, G.B.; Choudhuri, S.; Madan, A.; Dochterman, L.W.; Breen, J.G.; Parkinson, A.; Mast, R.W.; Meeks, R.G. Evaluation of octamethylcyclotetrasiloxane (D4) as an inducer of rat hepatic microsomal cytochrome p450, UDP-glucuronosyltransferase, and epoxide hydrolase: A 28-day inhalation study. Toxicol. Sci. 1998, 41, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Rotroff, D.M.; Dix, D.J.; Houck, K.A.; Knudsen, T.B.; Martin, M.T.; McLaurin, K.W.; Reif, D.M.; Crofton, K.M.; Singh, A.V.; Xia, M.; et al. Using in vitro high throughput screening assays to identify potential endocrine-disrupting chemicals. Environ. Health Perspect. 2013, 121, 7–14. [Google Scholar] [PubMed]
- Vuorinen, A.; Odermatt, A.; Schuster, D. In silico methods in the discovery of endocrine disrupting chemicals. J. Steroid Biochem. Mol. Biol. 2013, 137, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Van Breemen, R.B.; Li, Y. Caco-2 cell permeability assays to measure drug absorption. Expert Opin. Drug Metab. Toxicol. 2005, 1, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Fasinu, P.; Bouic, P.J.; Rosenkranz, B. Liver-based in vitro technologies for drug biotransformation studies—A review. Curr. Drug Metab. 2012, 13, 215–224. [Google Scholar] [CrossRef] [PubMed]
- T’jollyn, H.; Snoeys, J.; Colin, P.; van Bocxlaer, J.; Annaert, P.; Cuyckens, F.; Vermeulen, A.; van Peer, A.; Allegaert, K.; Mannens, G.; et al. Physiology-based ivive predictions of tramadol from in vitro metabolism data. Pharm. Res. 2015, 32, 260–274. [Google Scholar] [CrossRef] [PubMed]
- Wetmore, B.A. Quantitative in vitro-to-in vivo extrapolation in a high-throughput environment. Toxicology 2014, 332, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.T.; An, B.S.; Yang, H.; Jung, E.M.; Hwang, I.; Jeung, E.B. calbindin-D9k as a sensitive molecular biomarker for evaluating the synergistic impact of estrogenic chemicals on GH3 rat pituitary cells. Int. J. Mol. Med. 2012, 30, 1233–1240. [Google Scholar] [PubMed]
- Vo, T.T.; Jeung, E.B. An evaluation of estrogenic activity of parabens using uterine calbindin-D9k gene in an immature rat model. Toxicol. Sci. 2009, 112, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Tinnanooru, P.; Dang, V.H.; Nguyen, T.H.; Lee, G.S.; Choi, K.C.; Jeung, E.B. Estrogen regulates the localization and expression of calbindin-D9k in the pituitary gland of immature male rats via the ER α-pathway. Mol. Cell. Endocrinol. 2008, 285, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Jung, E.M.; An, B.S.; Hwang, I.; Vo, T.T.; Kim, S.R.; Lee, S.M.; Choi, K.C.; Jeung, E.B. Additional effects of bisphenol a and paraben on the induction of calbindin-D9k and progesterone receptor via an estrogen receptor pathway in rat pituitary GH3 cells. J. Physiol. Pharmacol. 2012, 63, 445–455. [Google Scholar] [PubMed]
- Kim, Y.R.; Jung, E.M.; Choi, K.C.; Jeung, E.B. Synergistic effects of octylphenol and isobutyl paraben on the expression of calbindin-D9k in GH3 rat pituitary cells. Int. J. Mol. Med. 2012, 29, 294–302. [Google Scholar] [PubMed]
- Imaoka, S.; Enomoto, K.; Oda, Y.; Asada, A.; Fujimori, M.; Shimada, T.; Fujita, S.; Guengerich, F.P.; Funae, Y. Lidocaine metabolism by human cytochrome p-450s purified from hepatic microsomes: Comparison of those with rat hepatic cytochrome p-450s. J. Pharmacol. Exp. Ther. 1990, 255, 1385–1391. [Google Scholar] [PubMed]
- Falany, C.N.; Li, G. Effects of age and pregnancy on cytochrome p450 induction by octamethyltetracyclosiloxane in female sprague-dawley rats. J. Biochem. Mol. Toxicol. 2005, 19, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Dorne, J.L. Human variability in hepatic and renal elimination: Implications for risk assessment. J. Appl. Toxicol. 2007, 27, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Genualdi, S.; Harner, T.; Cheng, Y.; Macleod, M.; Hansen, K.M.; van Egmond, R.; Shoeib, M.; Lee, S.C. Global distribution of linear and cyclic volatile methyl siloxanes in air. Environ. Sci. Technol. 2011, 45, 3349–3354. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Moody, R.P.; Koniecki, D.; Zhu, J. Low molecular weight cyclic volatile methylsiloxanes in cosmetic products sold in canada: Implication for dermal exposure. Environ. Int. 2009, 35, 900–904. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Wong, J.W.; Begley, T.H.; Hayward, D.G.; Limm, W. Determination of siloxanes in silicone products and potential migration to milk, formula and liquid simulants. Food Addit. Contam. Part A Chem. Anal. Control Expo Risk Assess. 2012, 29, 1311–1321. [Google Scholar] [CrossRef] [PubMed]
- Flassbeck, D.; Pfleiderer, B.; Klemens, P.; Heumann, K.G.; Eltze, E.; Hirner, A.V. Determination of siloxanes, silicon, and platinum in tissues of women with silicone gel-filled implants. Anal. Bioanal. Chem. 2003, 375, 356–362. [Google Scholar] [PubMed]
- Luu, H.M.; Hutter, J.C. Bioavailability of octamethylcyclotetrasiloxane (D(4)) after exposure to silicones by inhalation and implantation. Environ. Health Perspect. 2001, 109, 1095–1101. [Google Scholar] [CrossRef] [PubMed]
- Meeks, R.G. Bioavailability of D4 after inhalation and implantation exposure to silicones. Environ. Health Perspect. 2002, 110, A442–A443. [Google Scholar] [CrossRef] [PubMed]
- Stoica, G.E.; Franke, T.F.; Moroni, M.; Mueller, S.; Morgan, E.; Iann, M.C.; Winder, A.D.; Reiter, R.; Wellstein, A.; Martin, M.B.; et al. Effect of estradiol on estrogen receptor-α gene expression and activity can be modulated by the ERBB2/PI 3-K/AKT pathway. Oncogene 2003, 22, 7998–8011. [Google Scholar] [CrossRef] [PubMed]
- Okubo, T.; Yokoyama, Y.; Kano, K.; Kano, I. ER-dependent estrogenic activity of parabens assessed by proliferation of human breast cancer MCF-7 cells and expression of ER α and PR. Food Chem. Toxicol. 2001, 39, 1225–1232. [Google Scholar] [CrossRef]
- Long, X.; Nephew, K.P. Fulvestrant (ICI 182,780)-dependent interacting proteins mediate immobilization and degradation of estrogen receptor-α. J. Biol. Chem. 2006, 281, 9607–9615. [Google Scholar] [CrossRef] [PubMed]
- Wardell, S.E.; Marks, J.R.; McDonnell, D.P. The turnover of estrogen receptor α by the selective estrogen receptor degrader (SERD) fulvestrant is a saturable process that is not required for antagonist efficacy. Biochem. Pharmacol. 2011, 82, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Yeh, W.L.; Shioda, K.; Coser, K.R.; Rivizzigno, D.; McSweeney, K.R.; Shioda, T. Fulvestrant-induced cell death and proteasomal degradation of estrogen receptor α protein in MCF-7 cells require the CSK C-SRC tyrosine kinase. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Kocanova, S.; Mazaheri, M.; Caze-Subra, S.; Bystricky, K. Ligands specify estrogen receptor α nuclear localization and degradation. BMC Cell Biol. 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- McKim, J.M.; Wilga, P.C.; Breslin, W.J.; Plotzke, K.P.; Gallavan, R.H.; Meeks, R.G. Potential estrogenic and antiestrogenic activity of the cyclic siloxane octamethylcyclotetrasiloxane (D4) and the linear siloxane hexamethyldisiloxane (HMDS) in immature rats using the uterotrophic assay. Toxicol. Sci. 2001, 63, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.E.; Zhu, X.; Li, J.; Lyle, C.; Dowdy, S.; Podratz, K.C.; Byck, D.; Chen, H.B.; Jiang, S.W. Microarray analysis on gene regulation by estrogen, progesterone and tamoxifen in human endometrial stromal cells. Int. J. Mol. Sci. 2015, 16, 5864–5885. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.; Bergfeld, W.F.; Belsito, D.V.; Hill, R.A.; Klaassen, C.D.; Liebler, D.C.; Marks, J.G.; Shank, R.C.; Slaga, T.J.; Snyder, P.W.; et al. Safety assessment of cyclomethicone, cyclotetrasiloxane, cyclopentasiloxane, cyclohexasiloxane, and cycloheptasiloxane. Int. J. Toxicol. 2011, 30, S149–S227. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.; Ahn, C.; An, B.-S.; Jeung, E.-B. Induction of the Estrogenic Marker Calbindn-D9k by Octamethylcyclotetrasiloxane. Int. J. Environ. Res. Public Health 2015, 12, 14610-14625. https://doi.org/10.3390/ijerph121114610
Lee D, Ahn C, An B-S, Jeung E-B. Induction of the Estrogenic Marker Calbindn-D9k by Octamethylcyclotetrasiloxane. International Journal of Environmental Research and Public Health. 2015; 12(11):14610-14625. https://doi.org/10.3390/ijerph121114610
Chicago/Turabian StyleLee, Dongoh, Changhwan Ahn, Beum-Soo An, and Eui-Bae Jeung. 2015. "Induction of the Estrogenic Marker Calbindn-D9k by Octamethylcyclotetrasiloxane" International Journal of Environmental Research and Public Health 12, no. 11: 14610-14625. https://doi.org/10.3390/ijerph121114610
APA StyleLee, D., Ahn, C., An, B.-S., & Jeung, E.-B. (2015). Induction of the Estrogenic Marker Calbindn-D9k by Octamethylcyclotetrasiloxane. International Journal of Environmental Research and Public Health, 12(11), 14610-14625. https://doi.org/10.3390/ijerph121114610