Development of a Recombinase Polymerase Amplification Assay for the Detection of Pathogenic Leptospira

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Leptospira Strains and Others Microorganisms

{kind=link}
| No. | Species | Serovar | Strain | Status | Reference | Result |
|---|---|---|---|---|---|---|
| 1 | L. alexanderi | Manzhuang | A23 | Pathogenic | [12] | + |
2.3. Clinical Samples
2.4. DNA Extractions
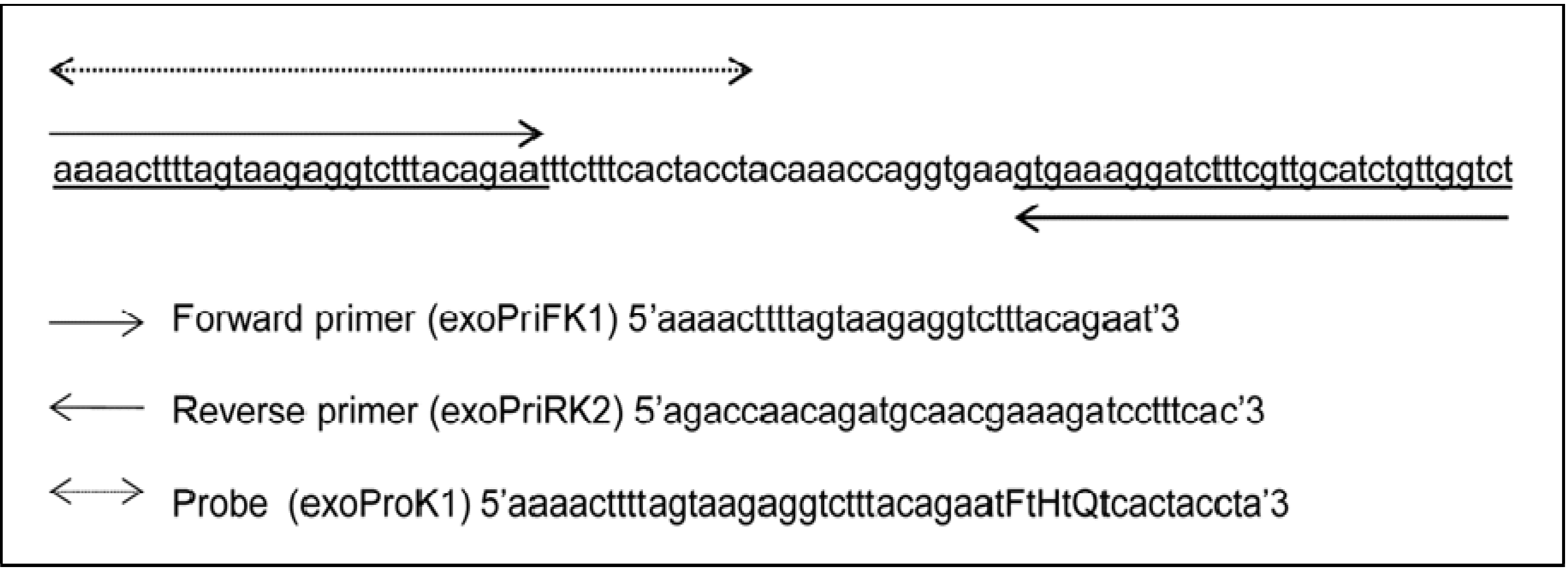
2.5. Primers and Probes Design
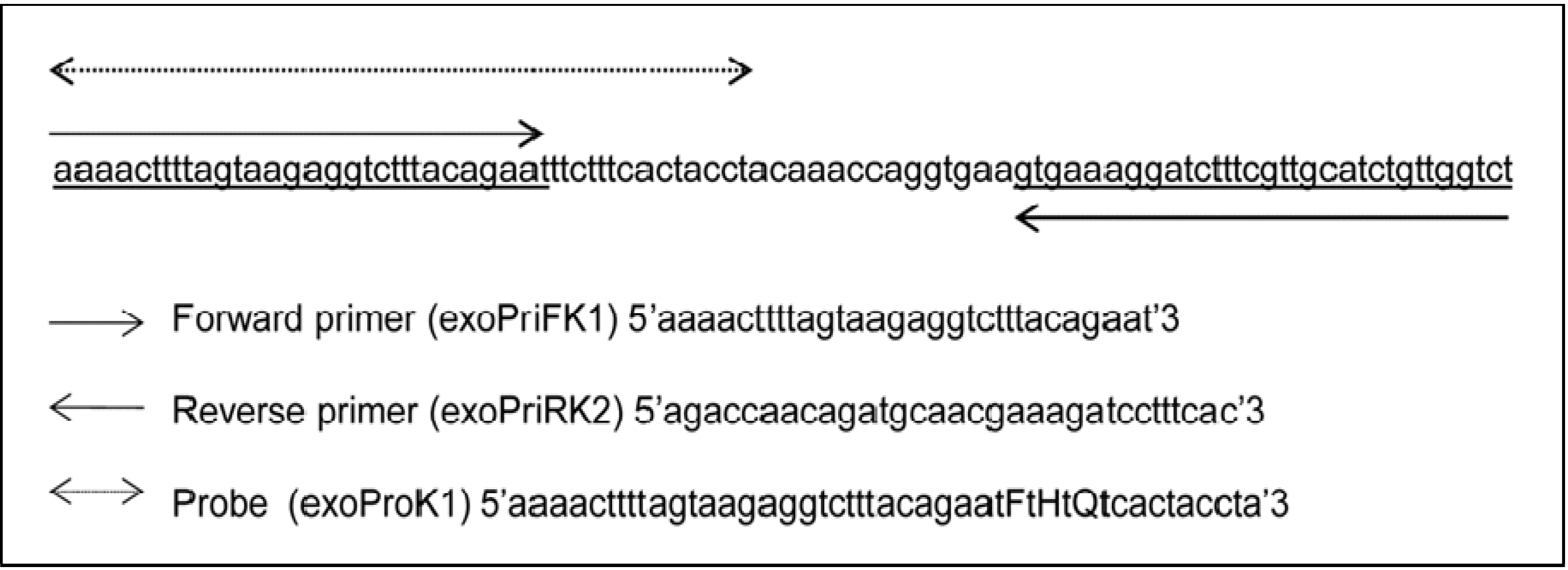
2.6. Recombinase Polymerase Amplification: Reagents and Conditions
2.7. Determination of Analytical and Diagnostic Sensitivity and Specificity
3. Results
3.1. Optimal Reaction Conditions and RPA Analytical Sensitivity and Specificity
3.2. The Diagnostic Sensitivity and Specificity
4. Discussion
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Picardeau, M.; Bertherat, E.; Jancloes, M.; Skouloudis, A.N.; Durski, K.; Hartskeerl, R.A. Rapid tests for diagnosis of leptospirosis: Current tools and emerging technologies. Diagn. Microbiol. Infect. Dis. 2014, 78, 1–8. [Google Scholar] [CrossRef]
- Ahmed, A.; Engelberts, M.F.; Boer, K.R.; Ahmed, N.; Hartskeerl, R.A. Development and validation of a real-time PCR for detection of pathogenic Leptospira species in clinical materials. PLoS One 2009, 4. [Google Scholar] [CrossRef]
- Levett, P.N. Leptospirosis. Clin. Microbiol. Rev. 2001, 14, 296–326. [Google Scholar] [CrossRef]
- Colebrander, A.; Terpstra, W.J.; Hartskeerl, R.A. Detectie van pathogene leptospiren met behulp van NASBA. Ned Tijdschr Geneeskd. 1994, 138, 436. [Google Scholar]
- Koizumi, N.; Nakajima, C.; Harunari, T.; Tanikawa, T.; Tokiwa, T.; Uchimura, E.; Furuya, T.; Mingala, C.N.; Villanueva, M.A.; Ohnishi, M.; Suzuki, Y. A new loop-mediated isothermal amplification method for rapid, simple, and sensitive detection of Leptospira spp. in urine. J. Clin. Microbiol. 2012, 50, 2072–2074. [Google Scholar] [CrossRef]
- Lin, X.; Chen, Y.; Lu, Y.; Yan, J.; Yan, J. Application of a loop-mediated isothermal amplification method for the detection of pathogenic Leptospira. Diagn. Microbiol. Infect. Dis. 2009, 63, 237–242. [Google Scholar]
- Sonthayanon, P.; Chierakul, W.; Wuthiekanun, V.; Thaipadungpanit, J.; Kalambaheti, T.; Boonsilp, S.; Amornchai, P.; Smythe, L.D.; Limmathurotsakul, D.; Day, N.P.; Peacock, S.J. Accuracy of loop-mediated isothermal amplification for diagnosis of human leptospirosis in Thailand. Am. J. Trop. Med. Hyg. 2011, 84, 614–620. [Google Scholar] [CrossRef]
- Suwancharoen, D.; Kulchim, C.; Chirathaworn, C.; Yoshida, S. Development of a novel primer combination to detect pathogenic Leptospira by loop-mediated isothermal amplification. J. Microbiol. Methods 2012, 91, 171–173. [Google Scholar] [CrossRef]
- Ahmed, A.; Grobusch, M.P.; Klatser, P.R.; Hartskeerl, R.A. Molecular approaches in the detection and characterization of Leptospira. J. Bacteriol. Parasitol. 2012, 3. [Google Scholar] [CrossRef]
- Piepenburg, O.; Williams, C.H.; Stemple, D.L.; Armes, N.A. DNA detection using recombination proteins. PLoS Biol. 2006, 4. [Google Scholar] [CrossRef]
- Hartskeerl, R.A. Leptospirosen. In LCI-richtlijnen Infectieziektebestrijding; Van Steenbergen, J.E., Timen, A., Beaujean, D.J.M.A., Eds.; Landelijke Coordinatie Infectieziektebestrijding: Bilthoven, The Netherland, 2008; pp. 238–243. [Google Scholar]
- Brenner, D.J.; Kaufmann, A.F.; Sulzer, K.R.; Steigerwalt, A.G.; Rogers, F.C.; Weyant, R.S. Further determination of DNA relatedness between serogroups and serovars in the family Leptospiraceae with a proposal for Leptospira alexanderi sp. nov. and four new Leptospira genomospecies. Int. J. Syst. Bacteriol. 1999, 49, 839–858. [Google Scholar] [CrossRef]
- Levett, P.N.; Morey, R.E.; Galloway, R.L.; Steigerwalt, A.G. Leptospira broomii sp. nov., isolated from humans with leptospirosis. Int. J. Syst. Evol. Microbiol. 2006, 56, 671–673. [Google Scholar] [CrossRef]
- Matthias, M.A.; Ricaldi, J.N.; Cespedes, M.; Diaz, M.M.; Galloway, R.L.; Saito, M.; Steigerwalt, A.G.; Patra, K.P.; Ore, C.V.; Gotuzzo, E.; et al. Human leptospirosis caused by a new, antigenically unique Leptospira associated with a Rattus species reservoir in the Peruvian Amazon. PLoS Negl. Trop. Dis. 2008, 2. [Google Scholar] [CrossRef]
- Perolat, P.; Chappel, R.J.; Adler, B.; Baranton, G.; Bulach, D.M.; Billinghurst, M.L.; Letocart, M.; Merien, F.; Serrano, M.S. Leptospira fainei sp. nov. isolated from pigs in Australia. Int. J. Syst. Bacteriol. 1998, 48, 851–858. [Google Scholar] [CrossRef]
- Victoria, B.; Ahmed, A.; Zuerner, R.L.; Ahmed, N.; Bulach, D.M.; Quinteiro, J.; Hartskeerl, R.A. Conservation of the S10-spc-α locus within otherwise highly plastic genomes provides phylogenetic insight into the genus Leptospira. PLoS One 2008, 3. [Google Scholar] [CrossRef]
- Goris, M.G.A.; Leeflang, M.M.; Boer, K.R.; Goeijenbier, M.; van Gorp, E.C.M.; Wagenaar, J.F.; Hartskeerl, R.A. Establishment of valid laboratory case definition for human leptospirosis. J. Bacteriol. Parasitol. 2012, 3. [Google Scholar] [CrossRef]
- Ellinghausen, H.C., Jr.; McCullough, W.G. Nutrition of Leptospira Pomona and growth of 13 other serotypes: Fractionation of oleic albumin complex and a medium of bovine albumin and polysorbate 80. Am. J. Vet. Res. 1965, 26, 45–51. [Google Scholar]
- Johnson, R.C.; Harris, V.G. Differentiation of pathogenic and saprophytic letospires. I. Growth at low temperatures. J. Bacteriol. 1967, 94, 27–31. [Google Scholar]
- Ahmed, A.; Engelberts, M.F.; Boer, K.R.; Ahmed, N.; Hartskeerl, R.A. Development and validation of a real-time PCR for detection of pathogenic Leptospira species in clinical materials. PLoS One 2009, 4. [Google Scholar] [CrossRef]
- BLAST: Basic Local Alignment Search Tool. Available online: http://www.blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 28 March 2013).
- Stoddard, R.A.; Gee, J.E.; Wilkins, P.P.; McCaustland, K.; Hoffmaster, A.R. Detection of pathogenic Leptospira spp. through TaqMan polymerase chain reaction targeting the lipL32 gene. Diagn. Microbiol. Infect. Dis. 2009, 64, 247–255. [Google Scholar] [CrossRef]
- Newcombe, R.G. Two-sided confidence intervals for the single proportion: Comparison of seven methods. Stat. Med. 1998, 17, 857–872. [Google Scholar] [CrossRef]
- Wilson, E. Probable inference, the law of succession, and statistical inference. J. Am. Stat. Assoc. 1927, 22, 209–212. [Google Scholar] [CrossRef]
- VassarStats: Website for Statistical Computation. Available online: http://www.vassarstats.net (accessed on 12 December 2013).
- Haake, D.A.; Chao, G.; Zuerner, R.L.; Barnett, J.K.; Barnett, D.; Mazel, M.; Matsunaga, J.; Levett, P.N.; Bolin, C.A. The leptospiral major outer membrane protein LipL32 is a lipoprotein expressed during mammalian infection. Infect. Immun. 2000, 68, 2276–2285. [Google Scholar] [CrossRef]
- Gravekamp, C.; van de Kemp, H.; Franzen, M.; Carrington, D.; Schoone, G.J.; Van Eys, G.J.; Everard, C.O.; Hartskeerl, R.A.; Terpstra, W.J. Detection of seven species of pathogenic leptospires by PCR using two sets of primers. J. Gen. Microbiol. 1993, 139, 1691–1700. [Google Scholar] [CrossRef]
- Hookey, J.V. Characterization of Leptospiraceae by 16S DNA restriction fragment length polymorphisms. J. Gen. Microbiol. 1993, 139, 1681–1689. [Google Scholar] [CrossRef]
- Hookey, J.V.; Bryden, J.; Gatehouse, L. The use of 16S rDNA sequence analysis to investigate the phylogeny of Leaptospirceae and related spirochaetes. J. Gen. Microbiol. 1993, 139, 2585–2590. [Google Scholar] [CrossRef]
- Letocart, M.; Baranton, G.; Perolat, P. Rapid identification of pathogenic Leptospira species (Leptospira interrogans, L. borgpetersenii, and L. kirschneri) with species-specific DNA probes produced by arbitrarily primed PCR. J. Clin. Microbiol. 1997, 35, 248–253. [Google Scholar]
- Murgia, R.; Riquelme, N.; Baranton, G.; Cinco, M. Oligonucleotides specific for pathogenic and saprophytic Leptospira occurring in water. FEMS Microbiol. Lett. 1997, 148, 27–34. [Google Scholar] [CrossRef]
- Segers, R.P.; van Gestel, J.A.; Van Eys, G.J.; van der Zeijst, B.A.; Gaastra, W. Presence of putative sphingomyelinase genes among members of the family Leptospiraceae. Infect. Immun. 1992, 60, 1707–1710. [Google Scholar]
- Ahmed, A.; Anthony, R.M.; Hartskeerl, R.A. A simple and rapid molecular method for Leptospira species identification. Infect. Genet. Evol. 2010, 10, 955–962. [Google Scholar] [CrossRef]
- Limmathurotsakul, D.; Turner, E.L.; Wuthiekanun, V.; Thaipadungpanit, J.; Suputtamongkol, Y.; Chierakul, W.; Smythe, L.D.; Day, N.P.; Cooper, B.; Peacock, S.J. Fool’s gold: Why imperfect reference tests are undermining the evaluation of novel diagnostics: A reevaluation of 5 diagnostic tests for leptospirosis. Clin. Infect. Dis. 2012, 55, 322–331. [Google Scholar] [CrossRef]
- Nervig, R.M.; Ellinghausen, H.C., Jr. Viability of Leptospira interrogans serotype Grippotyphosa in swine urine and blood. Cornell Vet. 1978, 68, 70–77. [Google Scholar]
- Word Health Organization. Human Leptospirosis: Guidance for Diagnosis, Surveillance and Control; WHO: Geneva, Switzerland, 2003. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ahmed, A.; Van der Linden, H.; Hartskeerl, R.A. Development of a Recombinase Polymerase Amplification Assay for the Detection of Pathogenic Leptospira. Int. J. Environ. Res. Public Health 2014, 11, 4953-4964. https://doi.org/10.3390/ijerph110504953
Ahmed A, Van der Linden H, Hartskeerl RA. Development of a Recombinase Polymerase Amplification Assay for the Detection of Pathogenic Leptospira. International Journal of Environmental Research and Public Health. 2014; 11(5):4953-4964. https://doi.org/10.3390/ijerph110504953
Chicago/Turabian StyleAhmed, Ahmed, Hans Van der Linden, and Rudy A. Hartskeerl. 2014. "Development of a Recombinase Polymerase Amplification Assay for the Detection of Pathogenic Leptospira" International Journal of Environmental Research and Public Health 11, no. 5: 4953-4964. https://doi.org/10.3390/ijerph110504953
APA StyleAhmed, A., Van der Linden, H., & Hartskeerl, R. A. (2014). Development of a Recombinase Polymerase Amplification Assay for the Detection of Pathogenic Leptospira. International Journal of Environmental Research and Public Health, 11(5), 4953-4964. https://doi.org/10.3390/ijerph110504953