Fucoidan from Marine Brown Algae Inhibits Lipid Accumulation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
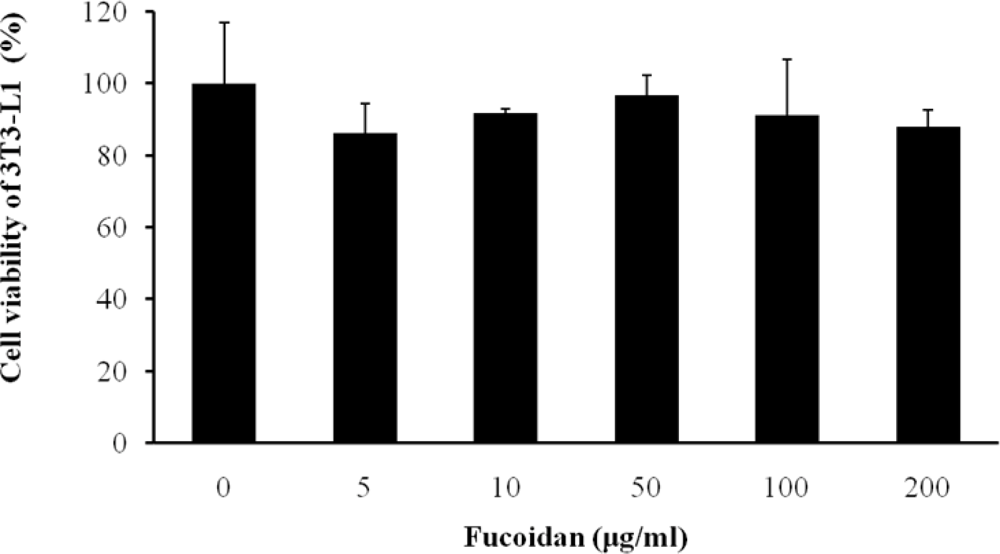
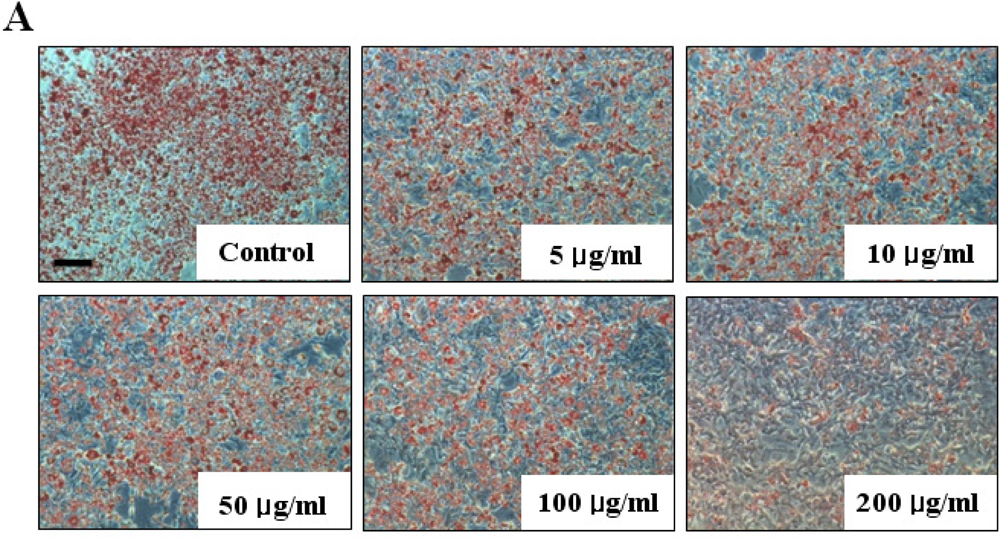
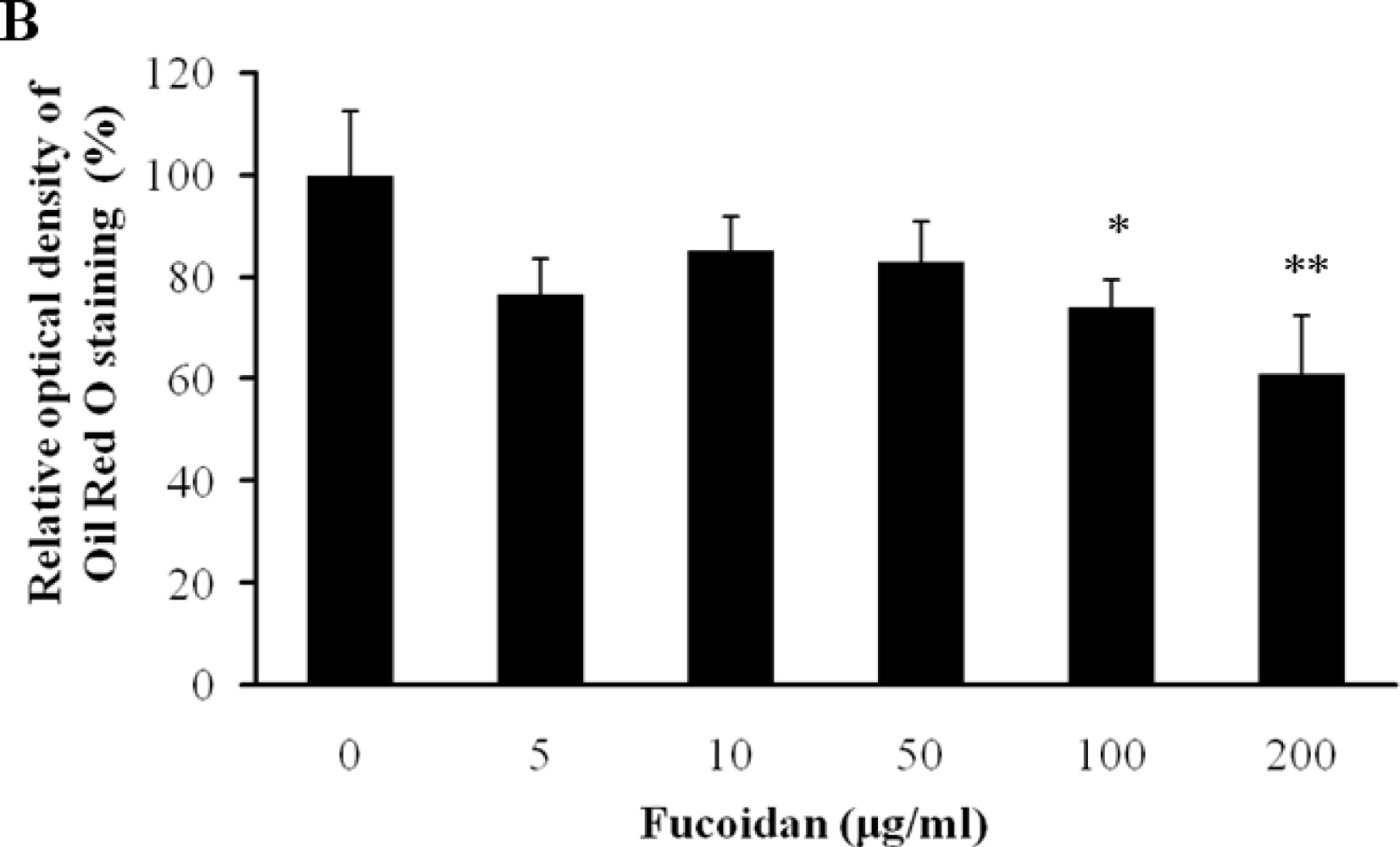
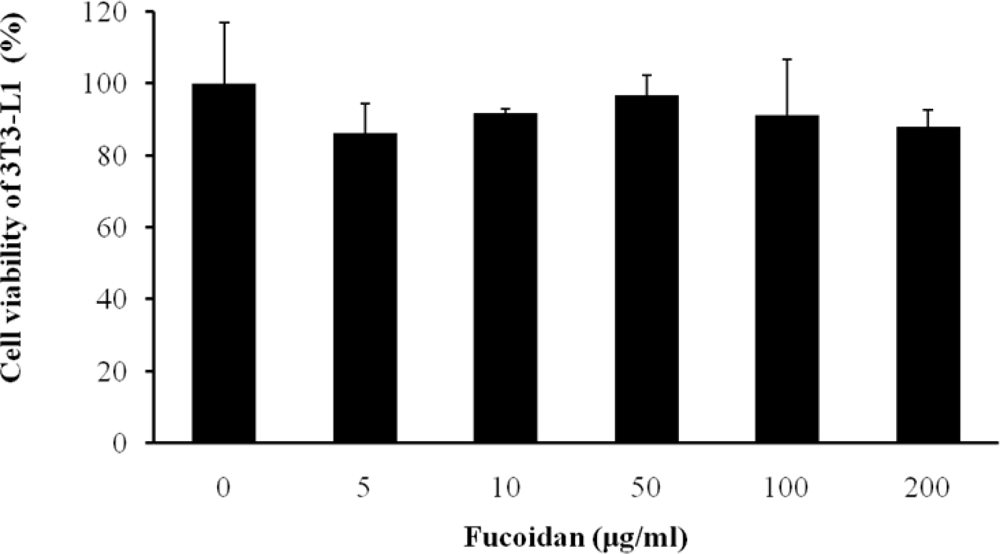
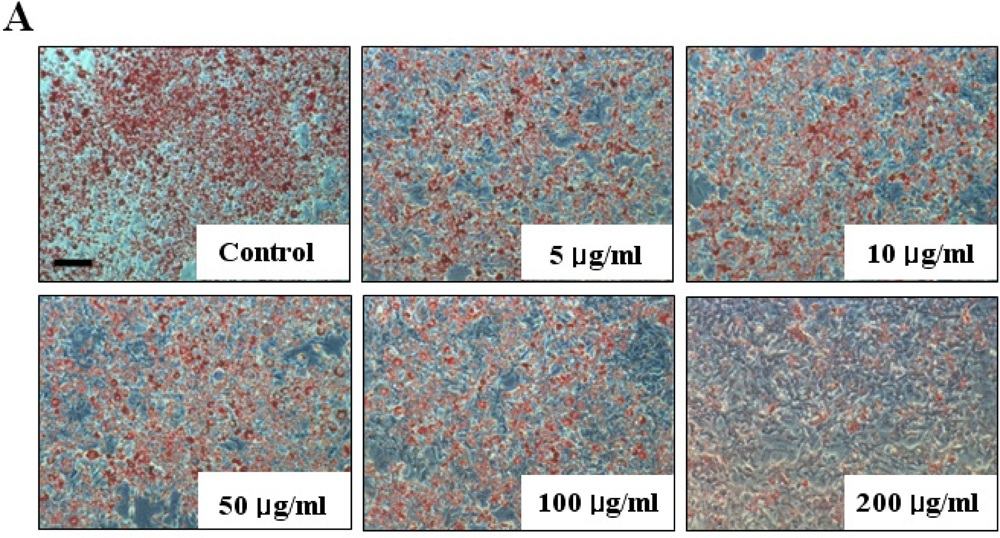
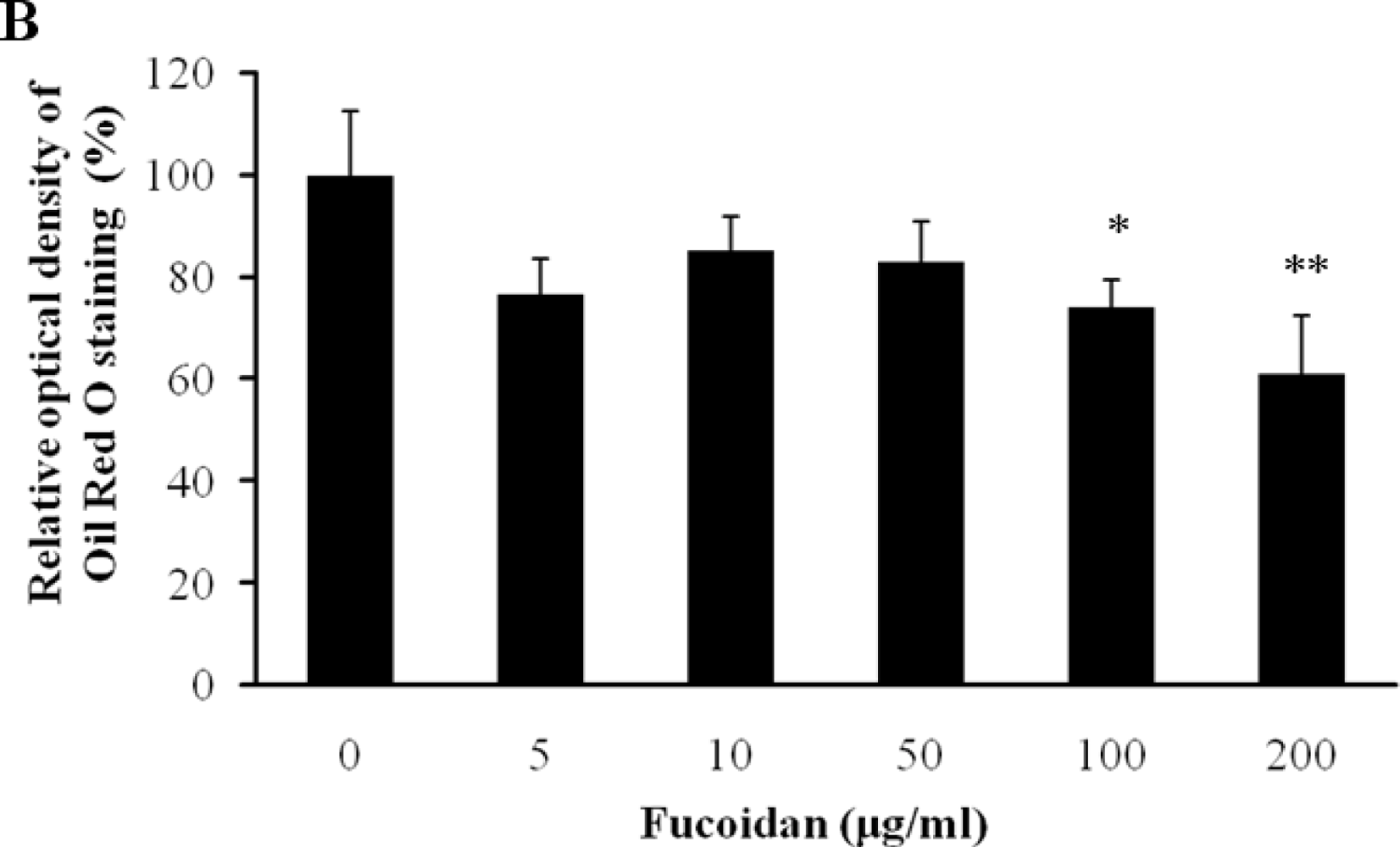
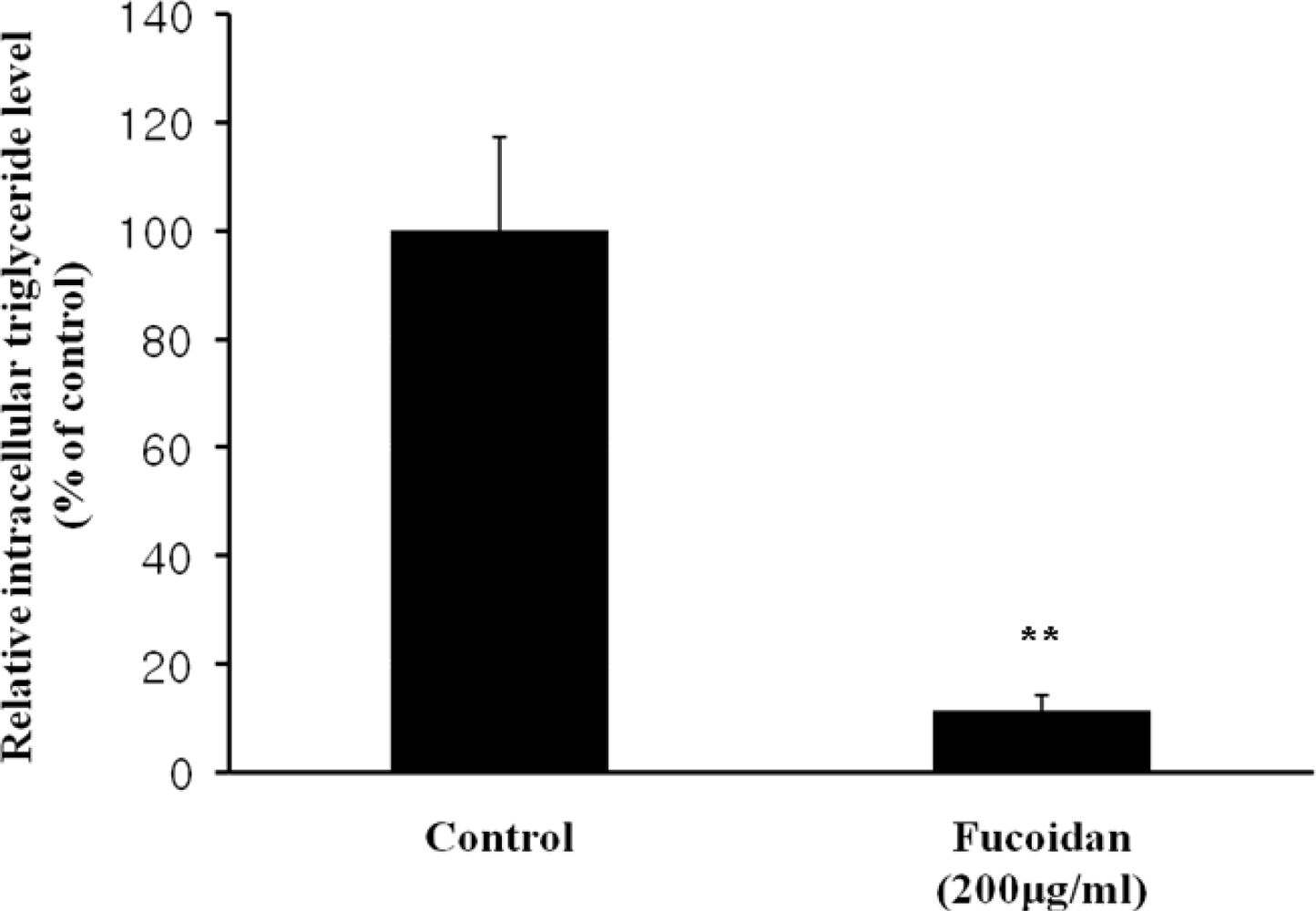
2.1. Cell Viability and Lipid Inhibitory Effect of Fucoidan on 3T3-L1 Adipocyte Differentiation
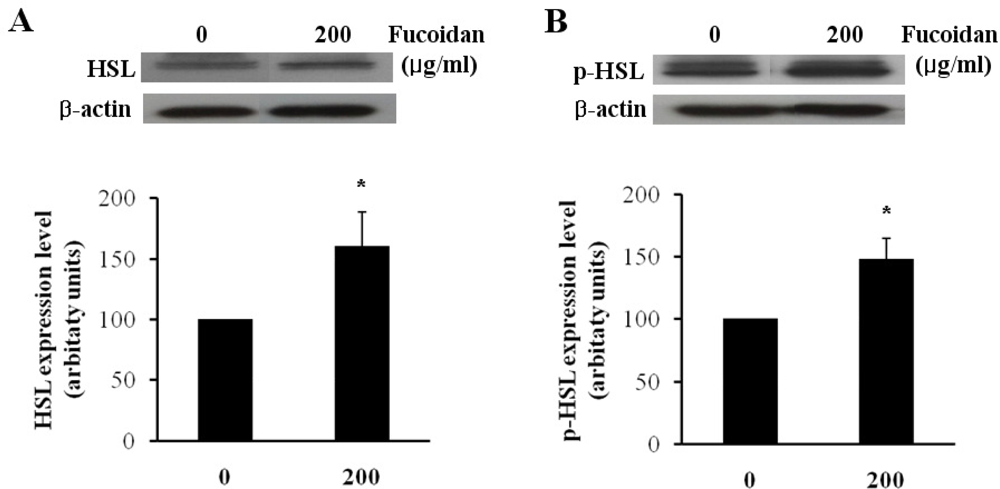
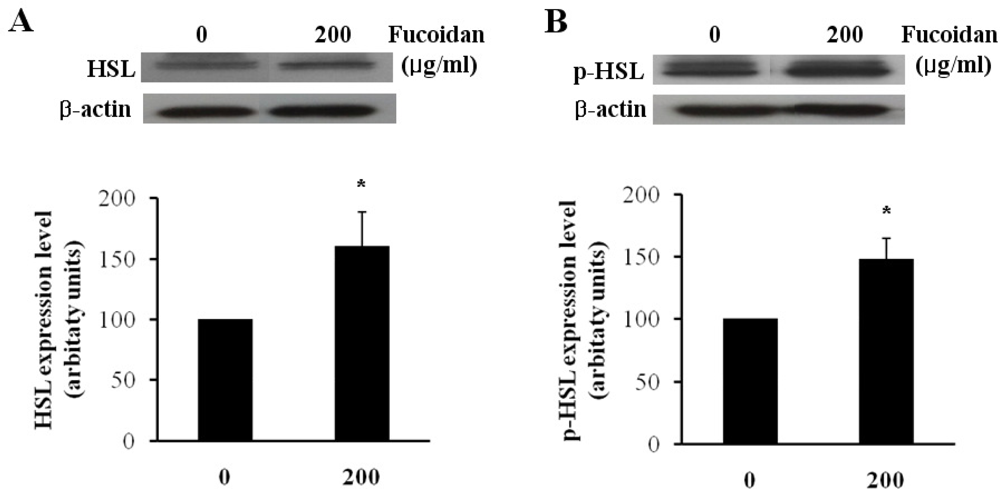
2.2. Expression Level of Lipolytic Gene
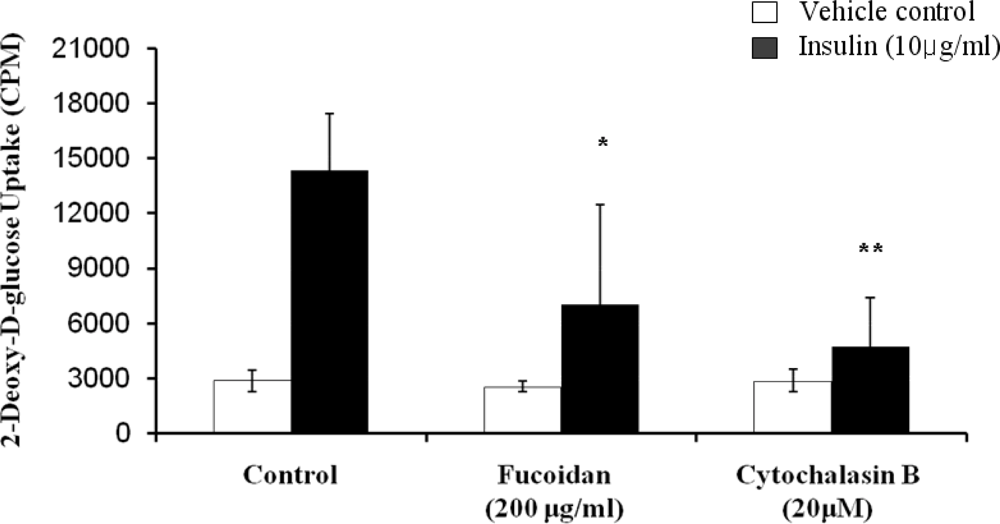
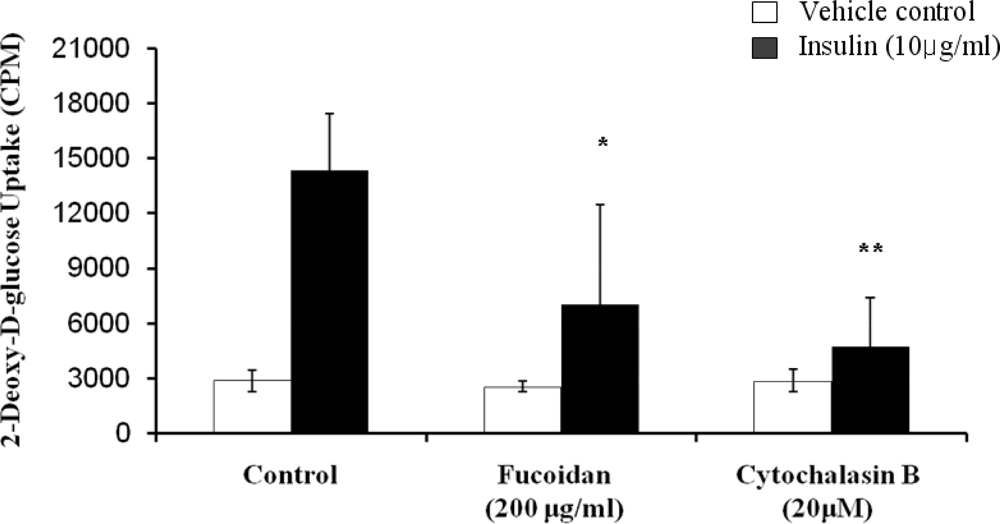
2.3. Inhibitory Effect of Fucoidan on Insulin-Induced Glucose Uptake
3. Experimental Section
3.1. Materials
3.2. Cell Culture and Differentiation
3.3. Cell Viability
3.4. Western Blotting
3.5. Glucose Uptake Activity Assay
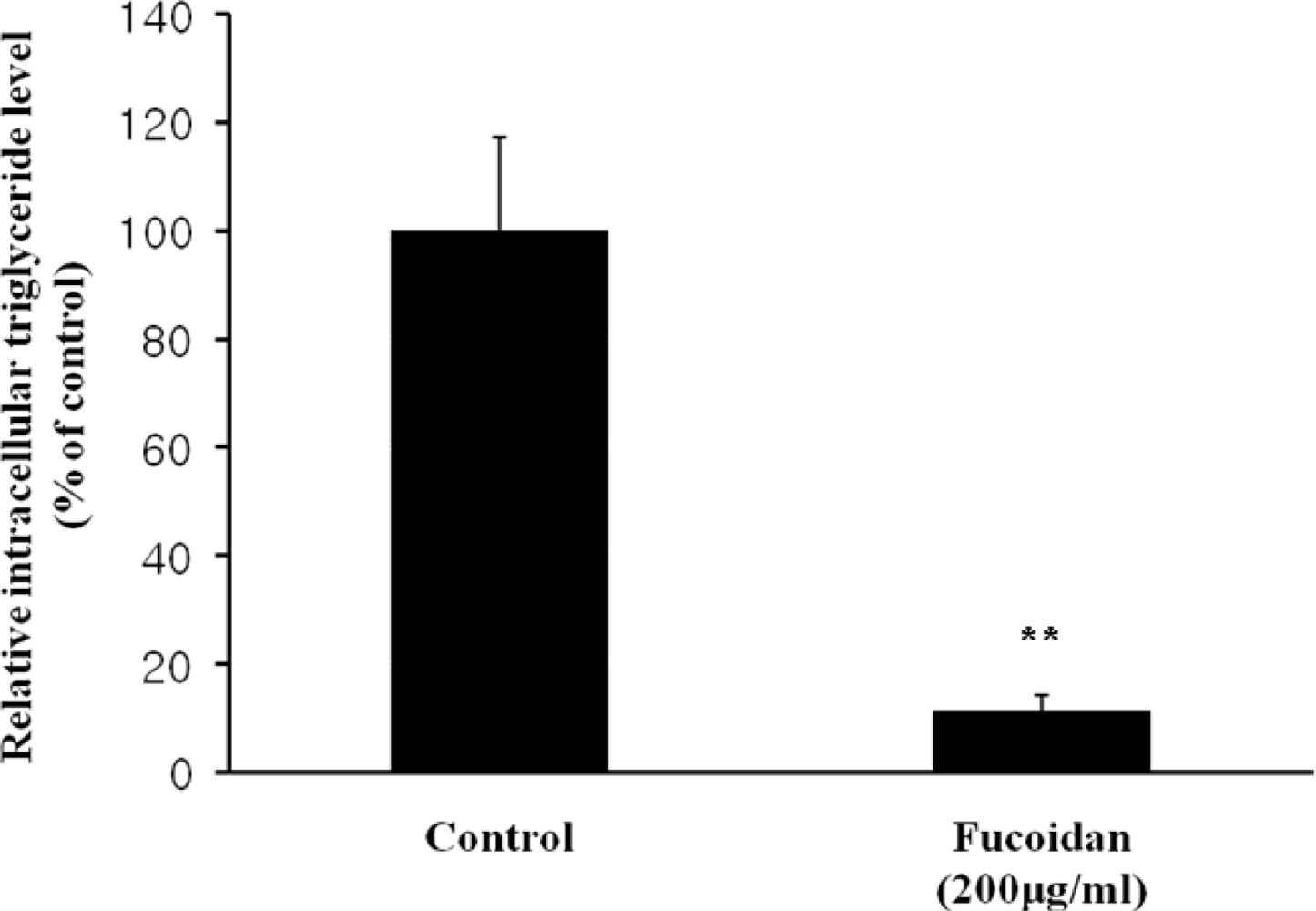
3.6. Triglyceride Accumulation Content Assay
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
- Samples Availability: Available from the authors.
References
- Larsson, B; Svardsudd, K; Welin, L; Wilhelmsen, L; Bjorntorp, P; Tibblin, G. Abdominal adipose tissue distribution, obesity and risk of cardiovascular disease and death: 13 year follow up of participants in the study of men born in 1913. Br. Med. J 1984, 288, 1401–1404. [Google Scholar]
- Spiegelman, BM; Flier, JS. Obesity and the regulation of energy balance. Cell 2001, 104, 531–543. [Google Scholar]
- Kopelman, PG. Obesity as a medical problem. Nature 2000, 404, 635–643. [Google Scholar]
- Kawada, T; Takahashi, N; Fushiki, T. Biochemical and physiological characteristics of fat cell. J. Nutr. Sci. Vitaminol (Tokyo) 2001, 47, 1–12. [Google Scholar]
- Shi, Y; Burn, P. Lipid metabolism enzymes: Emerging drug targets for the treatment of obesity. Nat. Rev. Drug. Discov 2004, 3, 695–710. [Google Scholar]
- Chapman, VJ; Chapman, DJ. Seaweeds and Their Uses, 3rd ed; Chapman and Hall: London, UK, 1980; pp. 279–312. [Google Scholar]
- Patankar, MS; Oehninger, S; Barnett, T; Williams, RL; Clark, GF. A revised structure for fucoidan may explain some of its biological activities. J. Biol. Chem 1993, 29, 21770–21776. [Google Scholar]
- Li, B; Lu, F; Wei, X; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar]
- Kuznetsova, TA; Besednova, NN; Mamaev, AN; Momot, AP; Shevchenko, NM; Zvyagintseva, TN. Anticoagulant activity of fucoidan from brown algae. Fucus evanescens of the Okhotsk Sea. Bull. Exp. Biol. Med 2003, 5, 471–473. [Google Scholar]
- Maruyama, H; Tamauchi, H; Iizuka, M; Nakano, T. The role of NK cells in antitumor activity of dietary fucoidan from Undaria pinnatifida sporophylls (Mekabu). Planta Med 2006, 15, 1415–1417. [Google Scholar]
- Hayashi, K; Nakano, T; Hashimoto, M; Kanekiyo, K; Hayashi, T. Defensive effects of a fucoidan from brown alga Undaria pinnatifida against herpes simplex virus infection. Int. Immunopharmacol 2008, 8, 109–116. [Google Scholar]
- Wang, J; Zhang, Q; Zhang, Z; Li, Z. Antioxidant activity of sulfated polysaccharide fractions extracted from Laminaria japonica. Int. J. Biol. Macromol 2008, 2, 127–132. [Google Scholar]
- Kim, KJ; Lee, OH; Lee, BY. Fucoidan, a sulfated polysaccharide, inhibits adipogenesis through the mitogen-activated protein kinase pathway in 3T3-L1 preadipocytes. Life Sci 2010, 22, 791–797. [Google Scholar]
- Kim, MJ; Chang, UJ; Lee, JS. Inhibitory effects of fucoidan in 3T3-L1 adipocyte differentiation. Mar. Biotechnol 2009, 5, 557–562. [Google Scholar]
- Rocha de Souza, MC; Marques, CT; Guerra Dore, CM; Oliveira Rocha, HA; Ferreira da Silva, FR; Leite, EL. Antioxidant activities of sulfated polysaccharides from brown and red seaweeds. J. Appl. Phycol 2007, 19, 153–160. [Google Scholar]
- Student, AK; Hsu, RY; Lane, MD. Induction of fatty acid synthetase synthesis in differentiating 3T3-L1 preadipocytes. J. Biol. Chem 1980, 255, 4745–4750. [Google Scholar]
- Langin, D; Holm, C; Lafontan, M. Adipocyte hormone sensitive lipase: A major regulator of lipid metabolism. Proc. Nutr. Soc 1996, 55, 93–109. [Google Scholar]
- Kim, SO; Lee, EJ; Choe, WK. The effects of Ginseng Saponin-Re, Rc and green tea catechine; ECGC (Epigallocatechin Gallate) on leptin, hormone sensitive lipase and resistin mRNA expressions in 3T3-L1 adipocytes. Korean J. Nutr 2006, 39, 748–755. [Google Scholar]
- Cha, SY; Jang, JY; Lee, YH; Lee, G; Lee, HJ; Hwang, KT; Kim, YJ; Jun, WJ; Lee, JM. Lipolytic effect of methanol extracts from Luffa cylindrica in mature 3T3-L1 adipocytes. J. Korean Soc. Food Sci. Nutr 2010, 6, 813–819. [Google Scholar]
- Newsholme, EA. Carbohydrate metabolism in vivo: Regulation of the blood glucose level. Clin. Endocrinol. Metab 1976, 3, 543–578. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Park, M.-K.; Jung, U.; Roh, C. Fucoidan from Marine Brown Algae Inhibits Lipid Accumulation. Mar. Drugs 2011, 9, 1359-1367. https://doi.org/10.3390/md9081359
Park M-K, Jung U, Roh C. Fucoidan from Marine Brown Algae Inhibits Lipid Accumulation. Marine Drugs. 2011; 9(8):1359-1367. https://doi.org/10.3390/md9081359
Chicago/Turabian StylePark, Min-Kyoung, Uhee Jung, and Changhyun Roh. 2011. "Fucoidan from Marine Brown Algae Inhibits Lipid Accumulation" Marine Drugs 9, no. 8: 1359-1367. https://doi.org/10.3390/md9081359
APA StylePark, M.-K., Jung, U., & Roh, C. (2011). Fucoidan from Marine Brown Algae Inhibits Lipid Accumulation. Marine Drugs, 9(8), 1359-1367. https://doi.org/10.3390/md9081359