Quantification of Dissolved and Particulate Polyunsaturated Aldehydes in the Adriatic Sea

Abstract
:1. Introduction
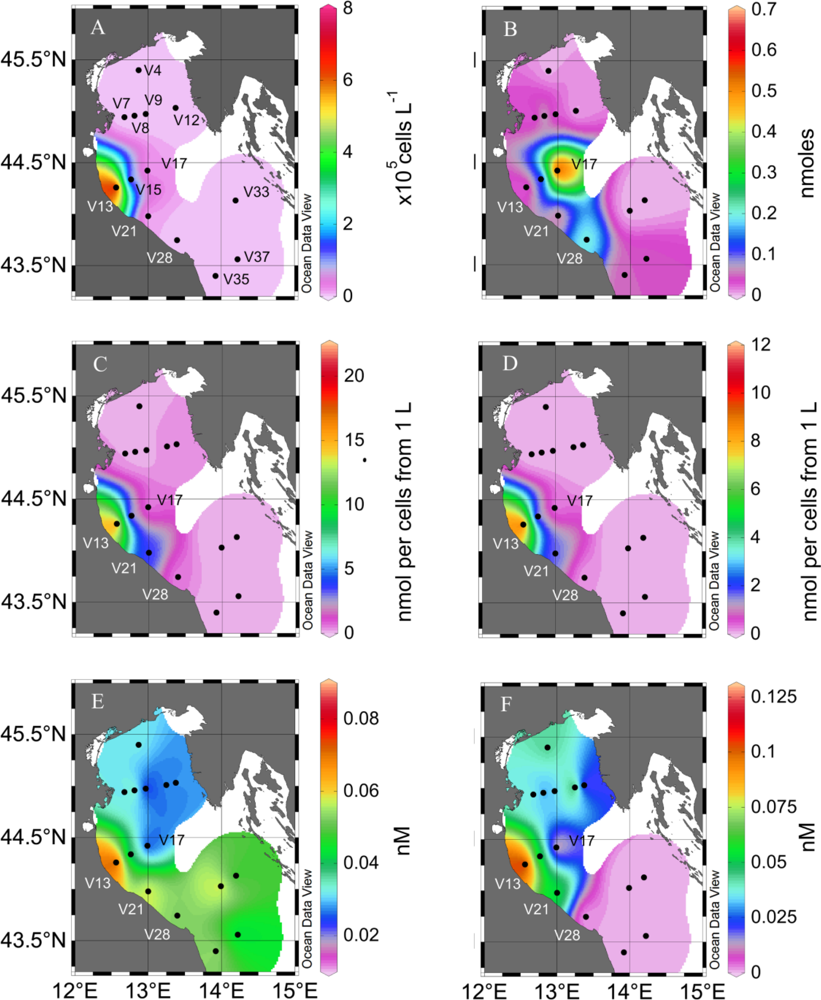
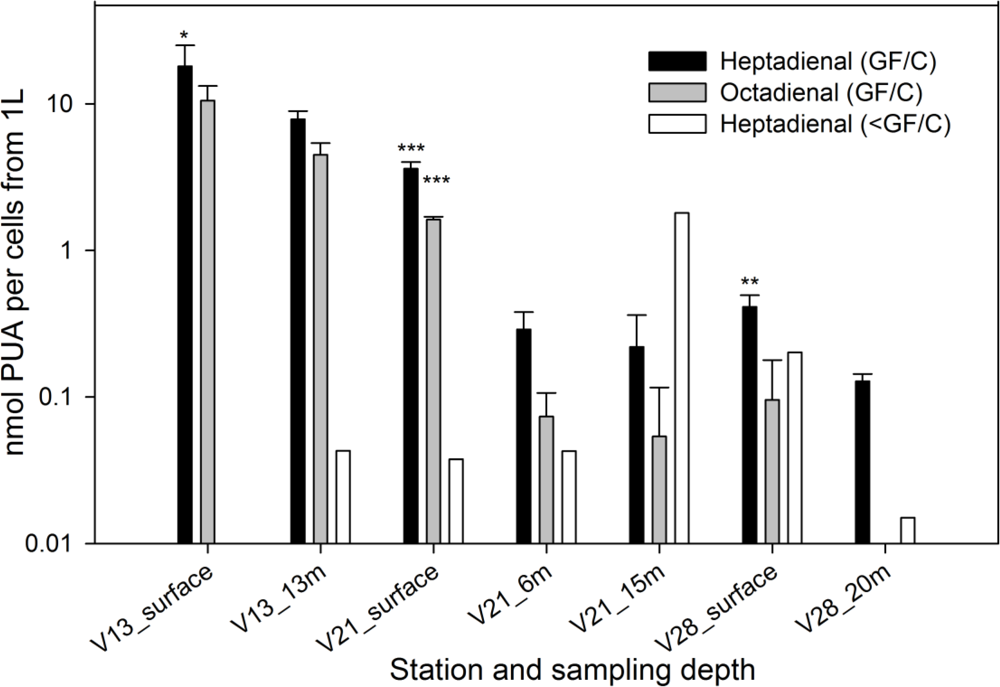
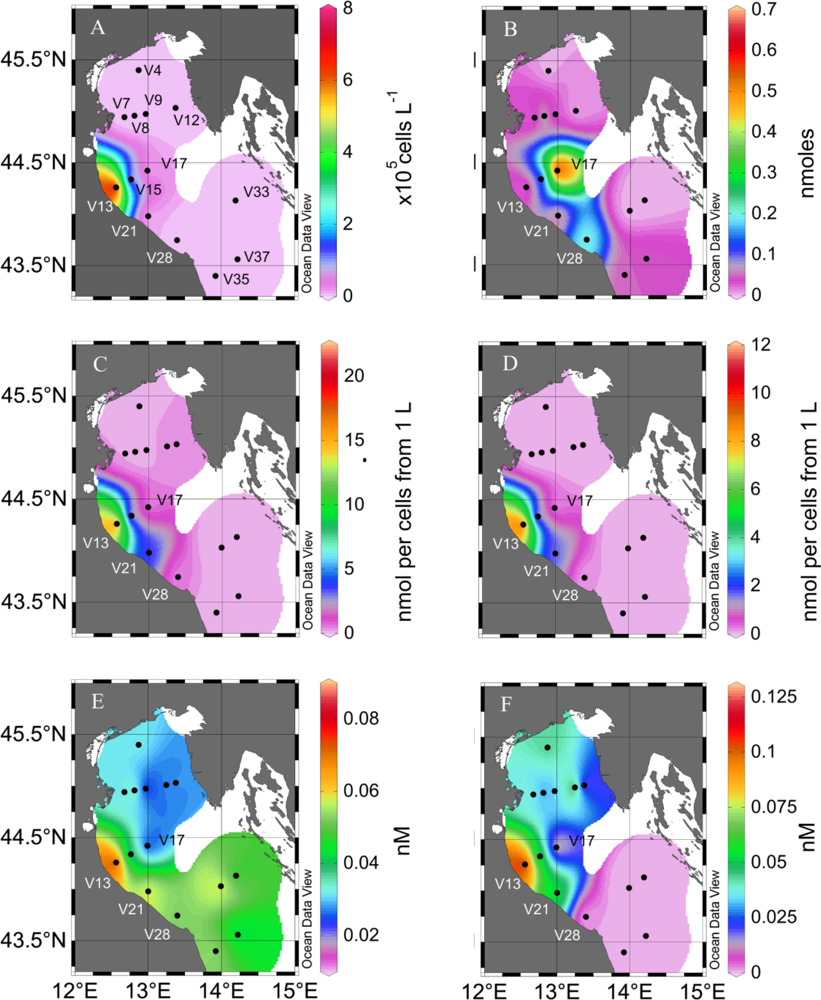
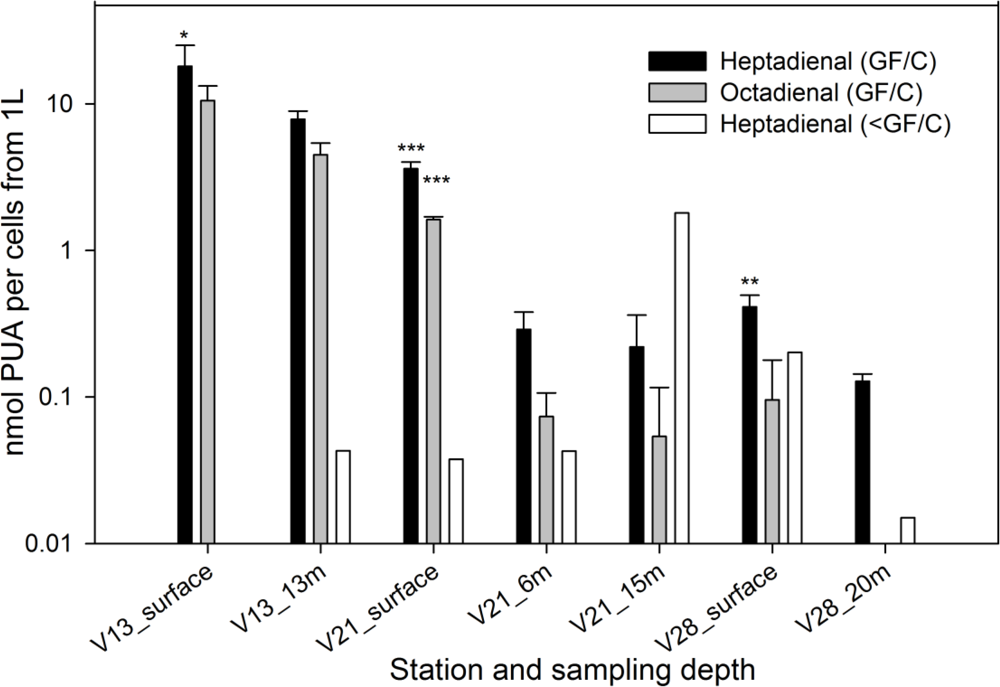
2. Results and Discussion
2.1. Particulate PUA
2.2. Dissolved PUA
3. Experimental Section
3.1. Field Sampling
3.2. Apparatus
3.3. Reagents
3.4. Data Analysis
3.5. Determination of the Particulate PUA
3.6. Determination of Dissolved PUA
3.7. Analysis and Quantification of PUA
4. Conclusions
Acknowledgments
References
- Wichard, T; Poulet, SA; Halsband-Lenk, C; Albaina, A; Harris, R; Liu, DY; Pohnert, G. Survey of the chemical defence potential of diatoms: Screening of fifty one species for alpha,beta,gamma,delta-unsaturated aldehydes. J Chem Ecol 2005, 31, 949–958. [Google Scholar]
- Ianora, A; Miralto, A. Toxigenic effects of diatoms on grazers, phytoplankton and other microbes: A review. Ecotoxicology 2010, 19, 493–511. [Google Scholar]
- Miralto, A; Barone, G; Romano, G; Poulet, SA; Ianora, A; Russo, GL; Buttino, I; Mazzarella, G; Laabir, M; Cabrini, M; Giacobbe, MG. The insidious effect of diatoms on copepod reproduction. Nature 1999, 402, 173–176. [Google Scholar]
- Adolph, S; Poulet, SA; Pohnert, G. Synthesis and biological activity of alpha,beta,gamma,delta-unsaturated aldehydes from diatoms. Tetrahedron 2003, 59, 3003–3008. [Google Scholar]
- Ianora, A; Miralto, A; Poulet, SA; Carotenuto, Y; Buttino, I; Romano, G; Casotti, R; Pohnert, G; Wichard, T; Colucci-D’Amato, L; Terrazzano, G; Smetacek, V. Aldehyde suppression of copepod recruitment in blooms of a ubiquitous planktonic diatom. Nature 2004, 429, 403–407. [Google Scholar]
- Casotti, R; Mazza, S; Brunet, C; Vantrepotte, V; Ianora, A; Miralto, A. Growth inhibition and toxicity of the diatom aldehyde 2-trans, 4-trans-decadienal on thalassiosira weissflogii (bacillariophyceae). J Phycol 2005, 41, 7–20. [Google Scholar]
- Ribalet, F; Berges, JA; Ianora, A; Casotti, R. Growth inhibition of cultured marine phytoplankton by toxic algal-derived polyunsaturated aldehydes. Aquat Toxicol 2007, 85, 219–227. [Google Scholar]
- Ribalet, F; Intertaglia, L; Lebaron, P; Casotti, R. Differential effect of three polyunsaturated aldehydes on marine bacterial isolates. Aquat Toxicol 2008, 86, 249–255. [Google Scholar]
- Adolph, S; Bach, S; Blondel, M; Cueff, A; Moreau, M; Pohnert, G; Poulet, SA; Wichard, T; Zuccaro, A. Cytotoxicity of diatom-derived oxylipins in organisms belonging to different phyla. J Exp Biol 2004, 207, 2935–2946. [Google Scholar]
- Vardi, A; Formiggini, F; Casotti, R; De Martino, A; Ribalet, F; Miralto, A; Bowler, C. A stress surveillance system based on calcium and nitric oxide in marine diatoms. PLoS Biol 2006, 4, 411–419. [Google Scholar]
- Vidoudez, C; Pohnert, G. Growth phase specific release of polyunsaturated aldehydes by the diatom skeletonema marinoi. J Plankton Res 2008, 30, 1305–1313. [Google Scholar]
- Pohnert, G. Wound-activated chemical defense in unicellular planktonic algae. Angew Chem Int Edit 2000, 39, 4352–4354. [Google Scholar]
- Pohnert, G. Phospholipase a(2) activity triggers the wound-activated chemical defense in the diatom thalassiosira rotula. Plant Physiol 2002, 129, 103–111. [Google Scholar]
- d’Ippolito, G; Tucci, S; Cutignano, A; Romano, G; Cimino, G; Miralto, A; Fontana, A. The role of complex lipids in the synthesis of bioactive aldehydes of the marine diatom skeletonema costatum. Biochim Biophys Acta 2004, 1686, 100–107. [Google Scholar]
- Barofsky, A; Pohnert, G. Biosynthesis of polyunsaturated short chain aldehydes in the diatom thalassiosira rotula. Org Lett 2007, 9, 1017–1020. [Google Scholar]
- Wichard, T; Pohnert, G. Formation of halogenated medium chain hydrocarbons by a lipoxygenase/hydroperoxide halolyase-mediated transformation in planktonic microalgae. J Am Chem Soc 2006, 128, 7114–7115. [Google Scholar]
- Poulet, SA; Escribano, R; Hidalgo, P; Cueff, A; Wichard, T; Aguilera, V; Vargas, CA; Pohnert, G. Collapse of calanus chilensis reproduction in a marine environment with high diatom concentration. J Exp Mar Biol Ecol 2007, 352, 187–199. [Google Scholar]
- Poulet, SA; Wichard, T; Ledoux, JB; Lebreton, B; Marchetti, J; Dancie, C; Bonnet, D; Cueff, A; Morin, P; Pohnert, G. Influence of diatoms on copepod reproduction. I. Field and laboratory observations related to calanus helgolandicus egg production. Mar Ecol Prog Ser 2006, 308, 129–142. [Google Scholar]
- Wichard, T; Poulet, SA; Boulesteix, AL; Ledoux, JB; Lebreton, B; Marchetti, J; Pohnert, G. Influence of diatoms on copepod reproduction. Ii. Uncorrelated effects of diatom-derived alpha,beta,gamma,delta-unsaturated aldehydes and polyunsaturated fatty acids on calanus helgolandicus in the field. Progr Oceanogr 2008, 77, 30–44. [Google Scholar]
- Ribalet, F; Vidoudez, C; Cassin, D; Pohnert, G; Ianora, A; Miralto, A; Casotti, R. High plasticity in the production of diatom-derived polyunsaturated aldehydes under nutrient limitation: Physiological and ecological implications. Protist 2009, 160, 444–451. [Google Scholar]
- Wichard, T; Gerecht, A; Boersma, M; Poulet, SA; Wiltshire, K; Pohnert, G. Lipid and fatty acid composition of diatoms revisited: Rapid wound-activated change of food quality parameters influences herbivorous copepod reproductive success. ChemBioChem 2007, 8, 1146–1153. [Google Scholar]
- Hansen, E; Ernstsen, A; Eilertsen, HC. Isolation and characterisation of a cytotoxic polyunsaturated aldehyde from the marine phytoplankter phaeocystis pouchetii (hariot) lagerheim. Toxicology 2004, 199, 207–217. [Google Scholar]
- Casotti, R. Stazione Zoologica A. Dohrn, Napoli, Italy. Unpublished work. 2011. [Google Scholar]
- Wichard, T; Poulet, SA; Pohnert, G. Determination and quantification of alpha,beta,gamma,delta-unsaturated aldehydes as pentafluorobenzyl-oxime derivates in diatom cultures and natural phytoplankton populations: Application in marine field studies. J Chromatogr B Analyt Technol Biomed Life Sci 2005, 814, 155–161. [Google Scholar]

{kind=link}
{kind=link}
| Station | V4 | V7 | V8 | V9 | V12 | V13 | V13 | V15 | V17 | V21 | V21 | V21 | V28 | V28 | V33 | V35 | V37 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Depth (m) | 0 | 0 | 0 | 0 | 0 | 0 | 13 | 0 | 0 | 0 | 6 | 15 | 0 | 17 | 0 | 0 | 0 |
| Species and cell densities (×103 cell L−1) | |||||||||||||||||
| Bacillariophyceae1 | 2.7 | 1.4 | 1.1 | 4.3 | 6.5 | 814 | 159.1 | 2.2 | 2.2 | 7.5 | 5.4 | 4.3 | 4.6 | 9.3 | 7 | 2.5 | 4.9 |
| Cerataulina pelagica | 2.2 | 4.3 | 37.7 | 58.6 | 2.2 | 1.1 | 0.8 | 3.6 | |||||||||
| Chaetoceros spp. | 2.2 | 3.5 | |||||||||||||||
| Guinardia flaccida | 2.2 | 0.5 | |||||||||||||||
| Pseudosolenia calcar avis | 1.1 | 0.3 | 0.7 | 0.4 | |||||||||||||
| Skeletonema marinoi | 776.3 | 71.2 | 0.5 | 1.6 | 2.2 | ||||||||||||
| Thalassiosira rotula | 2.2 | 1.3 | 0.7 | ||||||||||||||
| Dinophyceae1 | 2.2 | 135.1 | 35.6 | 112.1 | 114.3 | 237.2 | 184.2 | 0.5 | 29.1 | 5.9 | 0.5 | 7.5 | 11.5 | 1.6 | 1.1 | 1.6 | |
| Gonyaulax polygramma | 8.6 | 0.3 | |||||||||||||||
| Gymnodinium spp. | 11.5 | 4.3 | 15.1 | 16.2 | 16.7 | 0.5 | 3.2 | 0.7 | 1.6 | 1.6 | |||||||
| Prorocentrum minimum | 1.6 | 122.2 | 20.5 | 15.1 | 81.9 | 221 | 142.3 | 27 | 5.9 | 0.5 | 3 | 9.3 | |||||
| Und. naked | 0.5 | 9.7 | 10.8 | 6.5 | 8.4 | 0.7 | |||||||||||
| Prymnesiophyceae1 | 15.1 | 10.1 | 21.6 | 56.1 | 30.2 | 5.4 | 4.2 | 18.6 | 33.4 | 1.6 | 16.7 | 17.3 | 12.7 | 27.3 | 6.5 | 16.2 | 11.9 |
| Emiliania huxleyi | 15.1 | 10.1 | 21.6 | 38.8 | 30.2 | 5.4 | 4.2 | 18.6 | 33.4 | 1.6 | 16.7 | 17.3 | 12.7 | 27.3 | 6.5 | 12.2 | 11.3 |
| Cryptophyceae1 | 11.3 | 46 | 18.3 | 97 | 28 | 248 | 12.6 | 5.7 | 9.2 | 39.9 | 18.3 | 5.4 | 13.5 | 18 | 19.4 | 4.3 | 3.2 |
| Nanoflagellates1 | 23.7 | 140.9 | 139.1 | 394.6 | 472.2 | 690 | 493.9 | 30.2 | 53.9 | 194.1 | 124.5 | 201.6 | 108.1 | 92 | 51.2 | 75.1 | 52 |
| Total1 | 55 | 333.5 | 215.6 | 664.1 | 651.2 | 1994.6 | 853.9 | 56.6 | 99.2 | 272.2 | 170.9 | 229.1 | 146.4 | 158.1 | 85.7 | 99.2 | 73.6 |
| Dissolved PUA (nM) | |||||||||||||||||
| Heptadienal | 0.032 | 0.03 | 0.039 | 0.019 | 0.028 | 0.079 | 0.128 | 0.033 | 0.022 | 0.059 | 0.062 | 0.051 | 0.053 | 0.049 | 0.046 | 0.054 | 0.041 |
| Octadienal | 0.043 | 0.039 | 0.027 | 0.032 | 0.005 | 0.118 | 0.109 | 0.048 | 0 | 0.055 | 0.037 | 0.05 | 0 | 0 | 0 | 0 | 0 |
| Particulate > 1.2 μ (nmol per cells from 1 L, ±Standard deviation, n = 3) | |||||||||||||||||
| Heptadienal | 0 | 0 | 0 | 0 | 0 | 18.1±7.1 | 7.9±1.1 | 0.04±0.01 | 0.05±0.02 | 3.61±0.4 | 0.29±0.091 | 0.22±0.14 | 0.41±0.08 | 0.13±0.02 | 0.02±0.02 | 0.03±0.02 | 0 |
| Octadienal | 0 | 0 | 0 | 0 | 0 | 10.5±2.7 | 4.5±0.9 | 0 | 0 | 1.62±0.07 | 0.074±0.033 | 0.05±0.06 | 0.1±0.08 | 0 | 0 | 0 | 0 |
| 0.2 < Particulate < 1.2 μ (nmol per cells from 1 L)2 | |||||||||||||||||
| Heptadienal | 0 | 0 | 0.106 | 0 | 0 | 0 | 0.043 | 0.064 | 0.635 | 0.038 | 0.043 | 1.804 | 0.202 | 0.015 | 0 | 0.046 | 0.041 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Vidoudez, C.; Casotti, R.; Bastianini, M.; Pohnert, G. Quantification of Dissolved and Particulate Polyunsaturated Aldehydes in the Adriatic Sea. Mar. Drugs 2011, 9, 500-513. https://doi.org/10.3390/md9040500
Vidoudez C, Casotti R, Bastianini M, Pohnert G. Quantification of Dissolved and Particulate Polyunsaturated Aldehydes in the Adriatic Sea. Marine Drugs. 2011; 9(4):500-513. https://doi.org/10.3390/md9040500
Chicago/Turabian StyleVidoudez, Charles, Raffaella Casotti, Mauro Bastianini, and Georg Pohnert. 2011. "Quantification of Dissolved and Particulate Polyunsaturated Aldehydes in the Adriatic Sea" Marine Drugs 9, no. 4: 500-513. https://doi.org/10.3390/md9040500
APA StyleVidoudez, C., Casotti, R., Bastianini, M., & Pohnert, G. (2011). Quantification of Dissolved and Particulate Polyunsaturated Aldehydes in the Adriatic Sea. Marine Drugs, 9(4), 500-513. https://doi.org/10.3390/md9040500