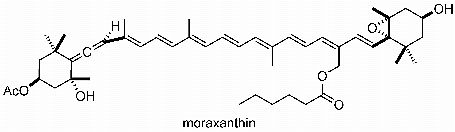
The New Carotenoid Pigment Moraxanthin Is Associated with Toxic Microalgae




 ,
,
Abstract
:
1. Introduction
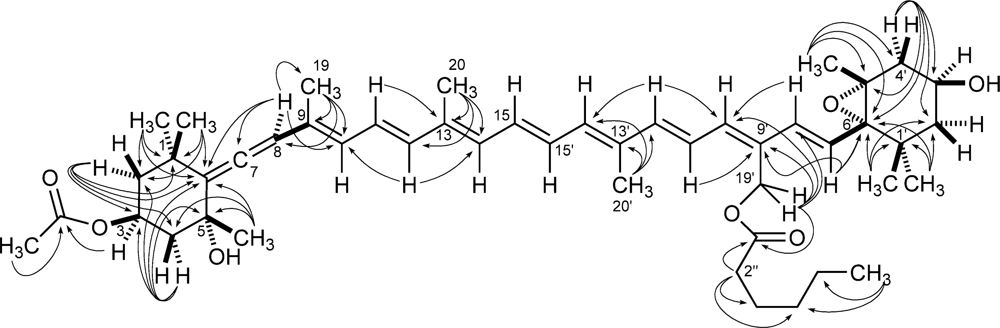
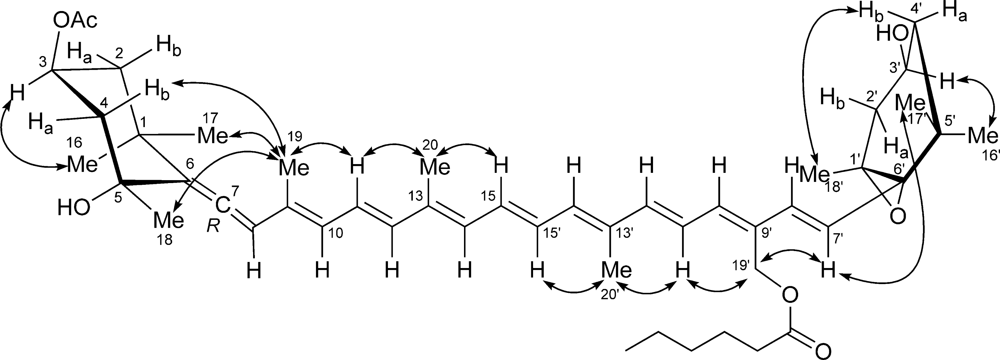
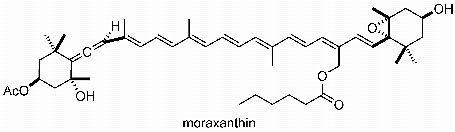
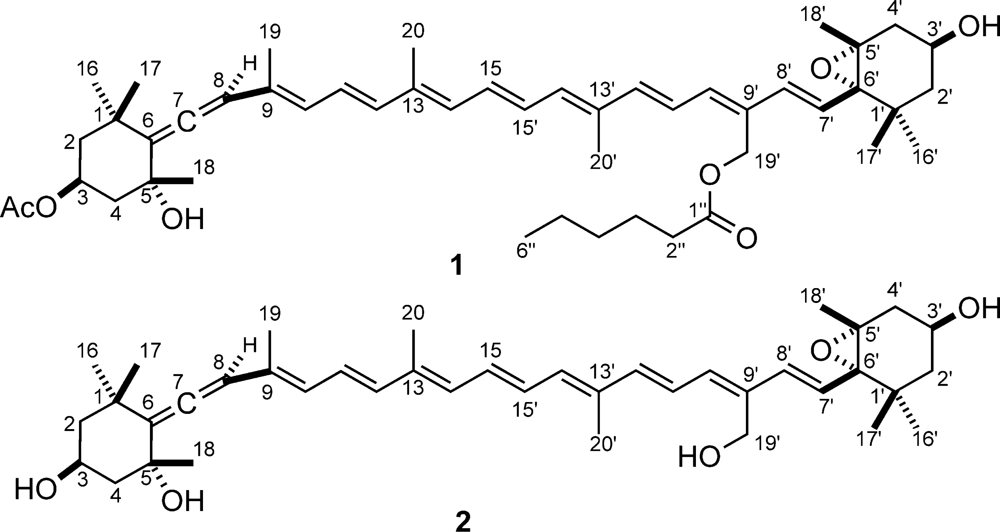
2. Results and Discussion
3. Experimental Section
3.1. General experimental procedures
3.2. Plant material
3.3. Pigment analysis
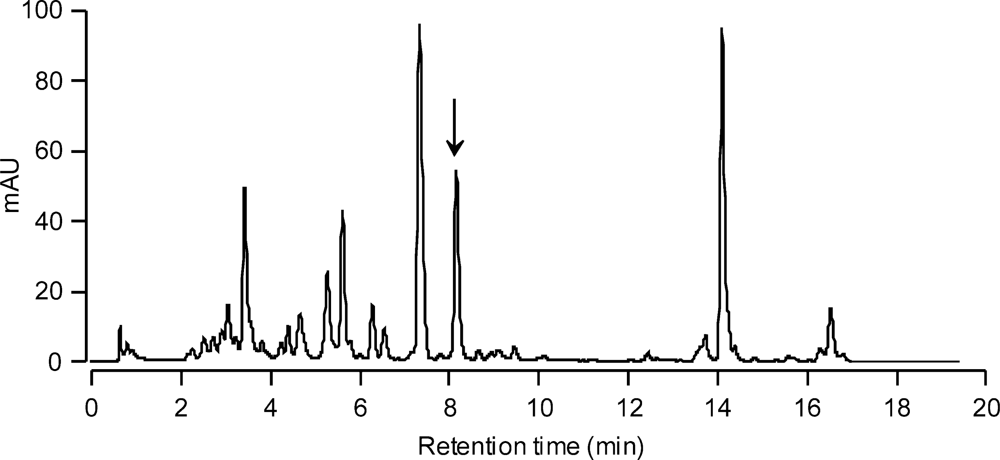
3.4. Analysis of algal bloom
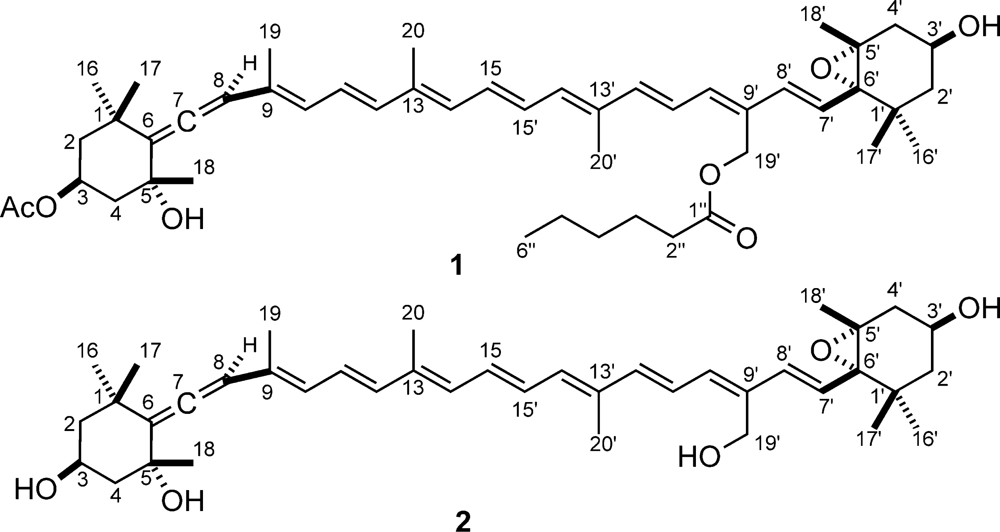
3.5. Extraction and isolation of moraxanthin (1)
3.6. Moraxanthin (1)
4. Conclusions
ELECTRONIC SUPPLEMENTARY INFORMATION
marinedrugs-09-00242-s001.pdfAcknowledgments
- Samples Availability: Available from the authors.
References
- Falkowski, PG; Raven, JA. Aquatic Photosynthesis; Blackwell Science: Oxford, UK, 1997; p. 375. [Google Scholar]
- Rowan, KS. Photosynthetic Pigments of Algae; Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]
- Porra, RJ; Pfündel, EE; Engel, N. Metabolism and function of photosynthetic pigments. In Phytoplankton Pigments in Oceanography; Jeffrey, SW, Mantoura, RFC, Wright, SW, Eds.; UNESCO Publishing: Paris, France, 1997; pp. 85–126. [Google Scholar]
- Scheer, H. The pigments. In Light-Harvesting Antennas in Photosynthesis Advances in Photosynthesis and Respiration; Green, BR, Parsons, WE, Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; Volume 13, p. 29. [Google Scholar]
- Rodríguez, F; Chauton, M; Johnsen, G; Andresen, K; Olsen, LM; Zapata, M. Photoacclimation in phytoplankton: implications for biomass estimates, pigment functionality and chemotaxonomy. Mar Biol 2006, 148, 963–971. [Google Scholar]
- Mangoni, O; Carrada, GC; Modigh, M; Catalano, G; Saggiomo, V. Photoacclimation in Antarctic bottom ice algae: an experimental approach. Polar Biol 2009, 32, 325–335. [Google Scholar]
- Mackey, MD; Mackey, DJ; Higgins, HW; Wright, SW. CHEMTAX- a program for estimating class abundances from chemical markers: application to HPLC measurements of phytoplankton. Mar Ecol Progr Ser 1996, 144, 265–283. [Google Scholar]
- Jeffrey, SW; Vesk, M. Introduction to marine phytoplankton and their pigment signatures. In Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods; Jeffrey, SW, Mantoura, RFC, Wright, SW, Eds.; UNESCO Publishing: Paris, France, 1997; pp. 37–84. [Google Scholar]
- Zapata, M; Jeffrey, SW; Wright, SW; Rodríguez, F; Garrido, JL; Clementson, L. Photosynthetic pigments in 37 species (65 strains) of Haptophyta: implications for oceanographyand chemotaxonomy. Mar Ecol Prog Ser 2004, 270, 83–102. [Google Scholar]
- Yoshii, Y; Takaichi, S; Maoka, T; Suda, S; Sekiguchi, H; Nakayama, T; Inouye, I. Variation of siphonaxanthin series among the genus Nephroselmis (Prasinophyceae, Chlorophyta), including a novel methoxyl carotenoid. J Phycol 2005, 41, 827–834. [Google Scholar]
- Wright, SW; Jeffrey, SW. Pigment markers for phytoplankton production. In Marine Organic Matter: Biomarkers, Isotopes and DNA; Volkman, JK, Ed.; Springer-Verlag: Berlin, Germany, 2006; pp. 71–104. [Google Scholar]
- Latasa, M. Improving estimations of phytoplankton class abundances using CHEMTAX. Mar Ecol Prog Ser 2007, 329, 13–21. [Google Scholar]
- Jeffrey, SW. Application of pigment methods to oceanography. In Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods; Jeffrey, SW, Mantoura, RFC, Wright, SW, Eds.; UNESCO Publishing: Paris, France, 1997; p. 127. [Google Scholar]
- Laza-Martinez, A; Seoane, S; Zapata, M; Orive, E. Phytoplankton pigment patterns in a temperate estuary: from unialgal cultures to natural assemblages. J Plankton Res 2007, 29, 913–929. [Google Scholar]
- Mangoni, O; Modigh, M; Mozetic, P; Bergamasco, A; Rivaro, P; Saggiomo, V. Structure and photosynthetic properties of phytoplankton assemblages in a highly dynamic system, the Northern Adriatic Sea. Estuar Coast Shelf Sci 2008, 77, 633–644. [Google Scholar]
- Ras, J; Claustre, H; Uitz, J. Spatial variability of phytoplankton pigment distributions in the Subtropical South Pacific Ocean: comparison between in situ and predicted data. Biogeosciences 2008, 5, 353–369. [Google Scholar]
- Smith, WO, Jr; Tozzi, S; DiTullio, GR; Dinnimand, M; Mangoni, O; Modigh, M; Saggiomo, V. Phytoplankton photosynthetic pigments in the Ross Sea: patterns and relationships among functional groups. J Mar Systems 2010, 82, 177–185. [Google Scholar]
- Anderson, DM. Toxic algal blooms and red tides: a global perspective. In Red Tides: Biology, Environmental Science and Tosicology; Okaichi, T, Anderson, DM, Nemoto, T, Eds.; Elsevier: New York, NY, USA, 1989; pp. 11–16. [Google Scholar]
- Zingone, A; Enevoldsen, HO. The diversity of harmful algal blooms: a challenge for science and management. Ocean Coastal Manage 2000, 43, 725–748. [Google Scholar]
- Masò, M; Garcés, E. Harmful microalgae blooms (HAB); problematic and conditions that induce them. Mar Pollut Bull 2006, 53, 620–630. [Google Scholar]
- Zingone, A; Siano, R; D’Alelio, A; Sarno, D. Potentially toxic and harmful microalgae from coastal waters of the Campania region (Tyrrhenian Sea, Mediterranean Sea). Harmful Algae 2006, 5, 321–337. [Google Scholar]
- Smayda, TJ. Harmful algal blooms: Their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnol Oceanogr 1997, 1137–1153. [Google Scholar]
- Smayda, TJ; Reynolds, CS. Community Assembly in Marine Phytoplankton: Application of Recent Models to Harmful Dinoflagellate Blooms. J Plankton Res 2001, 23, 447–461. [Google Scholar]
- Hallegraeff, GM; Anderson, DM; Cembella, AD (Eds.) Manual on Harmful Marine Microalgae, Monographs on oceanographic methodology 11; UNESCO: Paris, France, 2004.
- Pearson, L; Mihali, T; Moffitt, M; Kellmann, R; Neilan, B. On the Chemistry, Toxicology and Genetics of the Cyanobacterial Toxins, Microcystin, Nodularin, Saxitoxin and Cylindrospermopsin. Mar Drugs 2010, 8, 1650–1680. [Google Scholar]
- Ma, HY; Krock, B; Tillmann, U; Cembella, A. Preliminary Characterization of Extracellular Allelochemicals of the Toxic Marine Dinoflagellate Alexandrium tamarense Using a Rhodomonas salina Bioassay. Mar Drugs 2009, 7, 497–522. [Google Scholar] [Green Version]
- Lefebvre, KA; Bill, BD; Erickson, A; Baugh, KA; O’Rourke, L; Costa, PR; Nance, S; Trainer, VL. Characterization of intracellular and extracellular saxitoxin levels in both field and cultured Alexandrium spp. samples from Sequim Bay, Washington. Mar Drugs 2008, 6, 103–116. [Google Scholar]
- Wang, DZ. Neurotoxins from marine dinoflagellates: A brief review. Mar Drugs 2008, 6, 349–371. [Google Scholar]
- Landsberg, JH. The effects of harmful algal blooms on aquatic organisms. Rev Fish Sci 2002. [Google Scholar]
- Backer, LC; Kirkpatrick, B; Fleming, LE; Cheng, YS; Pierce, R; Bean, JA; Clark, R; Johnson, D; Wanner, A; Tamer, R; Zhou, Y; Baden, DG. Occupational exposure to aerosolized brevetoxins during Florida red tide events: effects on a healthy worker population. Environ Health Persp 2005, 113, 644–649. [Google Scholar]
- Pierce, RH; Henry, MS. Harmful algal toxins of the Florida red tide (Karenia brevis): natural chemical stressors in South Florida coastal ecosystems. Ecotoxicology 2008, 17, 623–631. [Google Scholar]
- Ramos, V; Vasconcelos, V. Palytoxin and Analogs: Biological and Ecological Effects. Mar Drugs 2010, 8, 2021–2037. [Google Scholar]
- Richlen, ML; Morton, SL; Jamali, EA; Rajan, A; Anderson, DM. The catastrophic 2008–2009 red tide in the Arabian gulf region, with observations on the identification and phylogeny of the fish-killing dinoflagellate. Cochlodinium polykrikoides Harmful Algae 2010, 9, 163–172. [Google Scholar]
- Wiese, M; D’Agostino; Paul, M; Mihali, TK; Moffitt, MC; Neilan, BA. Neurotoxic Alkaloids: Saxitoxin and Its Analogs. Mar Drugs 2010, 8, 2185–2211. [Google Scholar]
- Hallegraeff, GM. Harmful algal blooms: a global overview. In Manual on Harmful Marine Microalgae, Monographs on oceanographic methodology 11; Hallegraeff, GM, Anderson, DM, Cembella, AD, Eds.; UNESCO: Paris, France, 2004; pp. 25–50. [Google Scholar]
- Bourdelais, A; Tomas, CR; Narr, J; Kubanek, J; Baden, D. A new fish-killing alga in coastal Delaware produces neurotoxins. Environ. Health Perspect 2002, 110, 465–470. [Google Scholar]
- Hallegraeff, GM. A review of harmful algal blooms and their apparent global increase. Phycologia 1993, 32, 79–99. [Google Scholar]
- Tomas, CR; Smayda, TJ. Red tide blooms of Cochlodinium polykrikoides in a coastal cove. Harmful Algae 2008, 7, 308–317. [Google Scholar]
- Anderson, DM. Harmful Algal Blooms and Ocean Observing Systems: Needs, Present Status and Future Potential. In Fisheries for Global Welfare and Environment; Memorial book of the 5th World Fisheries Congress, TERRAPUB: Tokyo, Japan, 2008; Tsukamoto, K, Kawamura, T, Takeuchi, T, Beard, TD, Jr, Kaiser, MJ, Eds.; pp. 317–334. [Google Scholar]
- Richardson, TL; Pinckney, JL. Monitoring of the toxic dinoflagellate Karenia brevis using gyroxanthin-based detection methods. J Appl Phycol 2004, 16, 315–328. [Google Scholar]
- Gárate-Lizárraga, I; Bustillos-Guzmán, JJ; Morquecho, LM; Lechuga-Deveze, CH. First Outbreak of Cochlodinium polykrikoides in the Gulf of California. Harmful Algae News 2000, 21, 7. [Google Scholar]
- Bustillos-Guzmán, J; Gárate-Lizárraga, I; López-Cortés, D; Hernández-Sandoval, F. The use of pigment “fingerprints” in the study of harmful algal blooms. Rev Biol Trop 2004, 52(Suppl. 1), 17–26. [Google Scholar]
- Bowers, H; Tengs, T; Sayaka, G; Tomas, CR; Ono, C; Yoshimatsu, S; Oldach, D. Development of real-time PCR assays for the detection of Chattonella species in culture and environmental samples. In Harmful Algae; Steidinger, KA, Landsberg, JH, Tomas, CR, Vargo, GA, Eds.; Florida Fish and Wildlife Conservation Commission, Florida Institute of Oceanography and Intergovernmental Oceanographic Commission: St. Petersburg, FL, USA, 2004; pp. 231–233. [Google Scholar]
- Bowers, HA; Tomas, CR; Tengs, T; Kempton, JW; Lewitus, AJ; Oldach, DW. 35 Raphidophyceae (Chadefaud ex Silva) systematics and rapid identification: sequence analysis and 36 real-time PCR analysis. J Phycol 2006, 42, 1333–1348. [Google Scholar]
- Vidussi, F; Claustre, H; Bustillos-Guzman, J; Cailliau, C; Marty, J-C. Determination of chlorophylls and carotenoids of marine phytoplankton: separation of chlorophyll a from divinyl-chlorophyll a and zeaxanthin from lutein. J Plank Res 1996, 18, 2377–2382. [Google Scholar]
- Mantoura, RFC; Repeta, D. Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods; Jeffrey, SW, Mantoura, RFC, Wright, SW, Eds.; UNESCO Publishing: Paris, France, 1997; pp. 407–428. [Google Scholar]
- Britton, G; Liaaen-Jensen, S; Pfander, H. Carotenoids Handbook; Birkhäuser Verlag: Basel, Switzerland, 2004; p. 238. [Google Scholar]
- Andersen, R. Algal Culturing Techniques; Elsevier Academic Press: San Diego, CA, USA, 2005; p. 578. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
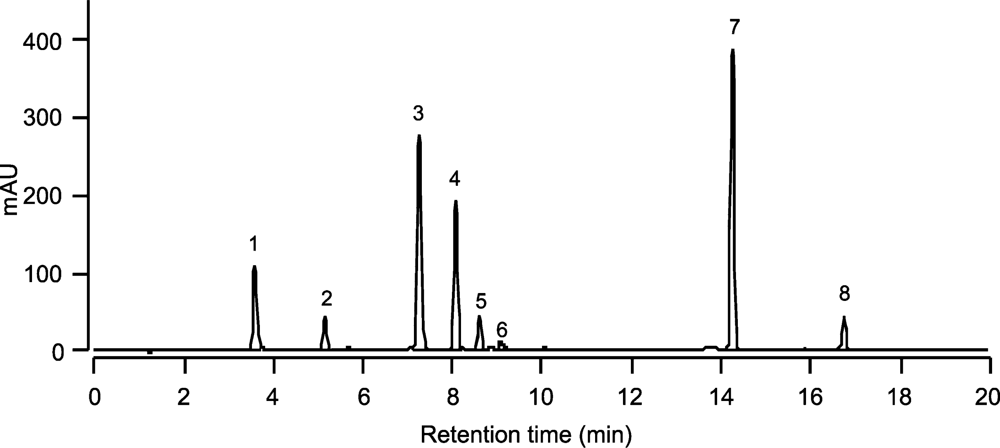
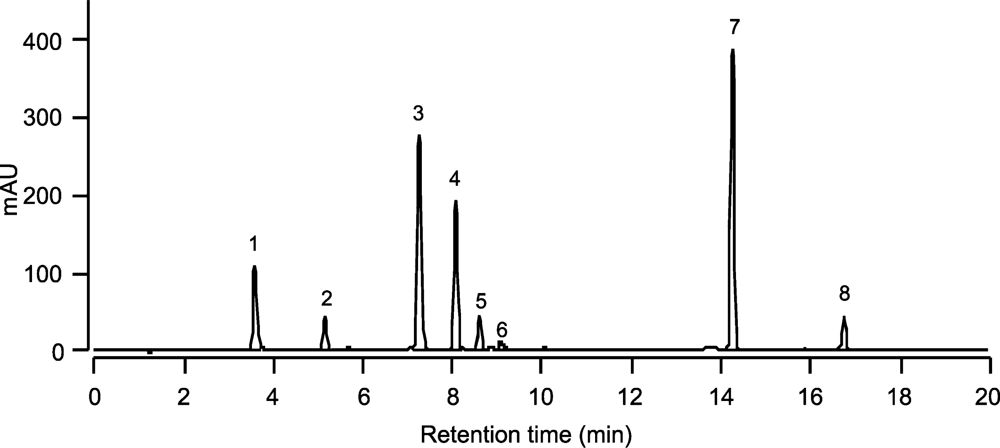
| Peak No. | Pigments | Retention time (min) | Absorption maxima (nm) | ||
|---|---|---|---|---|---|
| 1 | Chlorophyll c1 + c2 (Chlc1 + c2) | 3.59 | 445 | 583 | 634 |
| 2 | Unknown (RT 5.15) | 5.15 | 423 | 446 | 476 |
| 3 | Diadinoxanthin (Dd) | 7.24 | 422 | 446 | 475 |
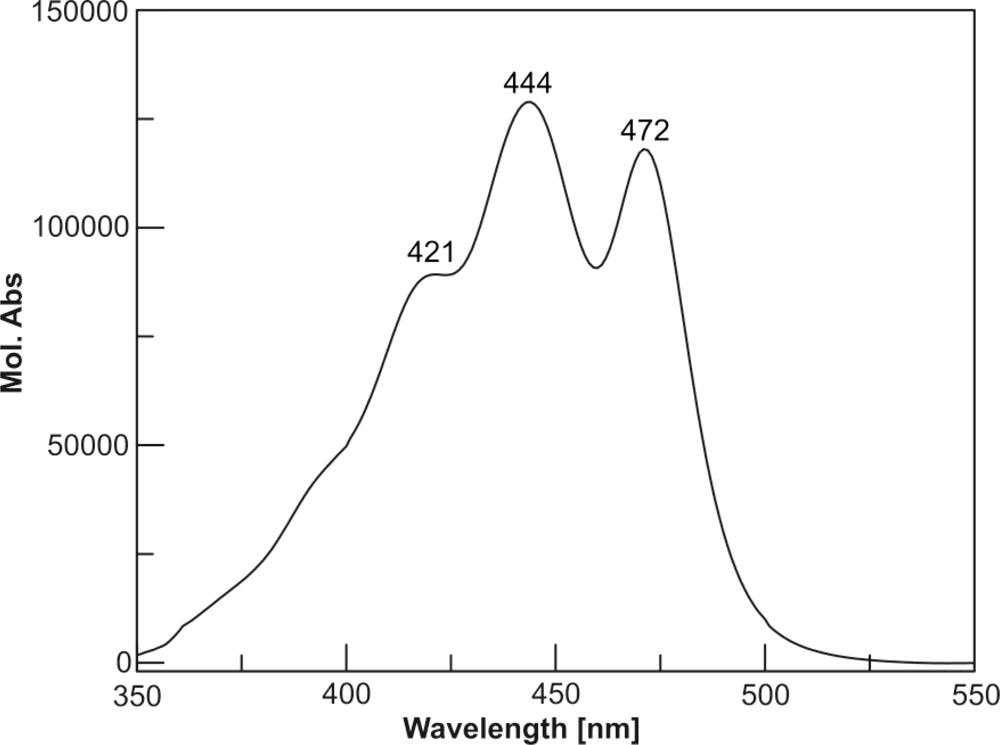
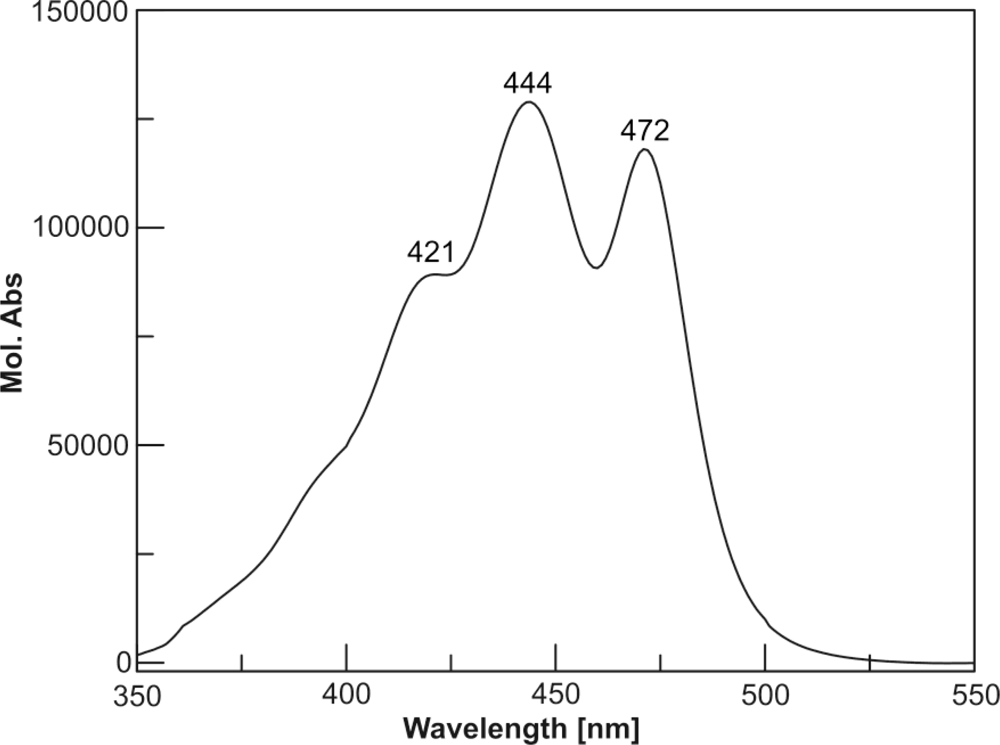
| 4 | Moraxanthin | 8.07 | 421 | 444 | 472 |
| 5 | Diatoxanthin (Dt) | 8.60 | 426 | 451 | 478 |
| 6 | Zeaxanthin (Zea) | 9.08 | 449 | 477 | |
| 7 | Chlorophyll a (Chla) | 14.25 | 432 | 617 | 665 |
| 8 | β-carotene (β-car) | 16.75 | 450 | 478 | |
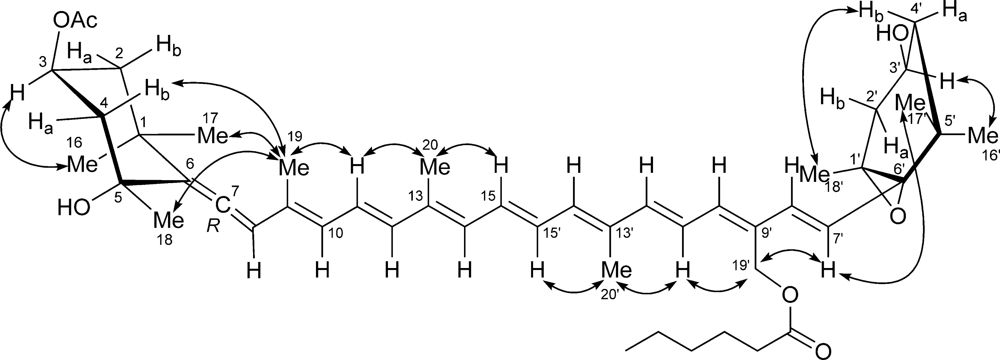
| Pos. | δH (J in Hz) | δC, mult | Pos. | δH (J in Hz) | δC, mult | ||
|---|---|---|---|---|---|---|---|
| 1 | - | 35.8, C | 1’ | - | 35.2, C | ||
| 2 | α | 2.05, ddd (12.4, 4.2, 2.2) | 45.8, CH2 | 2’ | α | 1.48, ddd (12.8, 3.3, 1.7) | 47.1, CH2 |
| β | 1.39, dd (12.4, 11.5) | β | 1.11, dd (12.8, 10.2) | ||||
| 3 | 5 .69, dddd (11.5, 11.5, 4.2, 4.2) | 67.7, CH | 3’ | 3.78, ddddd (10.2, 8.6, 5.2, 4.3, 3.3) | 63.7, CH | ||
| 3’-OH | 0.57, d (4.3) | - | |||||
| 4 | α | 2.30, ddd (12.8, 4.2, 2.2) | 45.5, CH2 | 4’ | α | 2.24, ddd (14.2, 5.2, 1.7) | 41.0, CH2 |
| β | 1.41, ddd (12.8, 11.5, 1.7) | β | 1.48, dd (14.2, 8.6) | ||||
| 5 | - | 72.0, C | 5’ | - | 66.6, C | ||
| 5-OH | 0.77, d (1.7) | - | - | - | |||
| 6 | - | 117.8, C | 6’ | - | 69.9, C | ||
| 7 | - | 202.1, C | 7’ | 6.22, d (15.7) | 125.9, CH | ||
| 8 | 6.03, s | 103.3, CH | 8’ | 6.63, d (15.7) | 134.9, CH | ||
| 9 | - | 131.9, C | 9’ | - | 132.4, C | ||
| 10 | 6.20, d (11.4) | 129.0, CH | 10’ | 6.27, d (11.6) | 136.3, CH | ||
| 11 | 6.70, dd (15.0, 11.4) | 125.3, CH | 11’ | 6.95, dd (14.9, 11.6) | 124.1, CH | ||
| 12 | 6.46, d (15.0) | 137.5, CH | 12’ | 6.39, d (14.9) | 141.0, CH | ||
| 13 | - | 136.8, C | 13’ | - | 136.4, C | ||
| 14 | 6.28, d (11.5) | 133.0, CH | 14’ | 6.27, d (11.5) | 134.5, CH2 | ||
| 15 | 6.66, dd (14.3, 11.5) | 131.1, CH | 15’ | 6.60, dd (14.3, 11.5) | 130.3, CH | ||
| 16 | 1.43, s | 29.0, CH3 | 16’ | 1.15, s | 25.2, CH3 | ||
| 17 | 1.08, s | 32.1, CH3 | 17’ | 1.14, s | 29.4, CH3 | ||
| 18 | 1.17, s | 30.9, CH3 | 18’ | 1.19, s | 20.0, CH3 | ||
| 19 | 1.78, s | 13.8, CH3 | 19’ | a | 5.07, d (12.3) | 58.1, CH2 | |
| b | 5.13, d (12.3) | ||||||
| 20 | 1.87, s | 12.7, CH3 | 20’ | 1.86, s | 12.6, CH3 | ||
| 1″ | - | 172.8, C | Ac | CH3 | 1.74, s | 20.8, CH3 | |
| 2″ | 2.14, t (7.5) | 34.2, CH2 | CO | - | 169.2, C | ||
| 3″ | 1.55, quintet (7.5) | 24.8, CH2 | |||||
| 4″ | 1.12, overlapped | 31.3, CH2 | |||||
| 5″ | 1.14, overlapped | 22.4, CH2 | |||||
| 6″ | 0.78, t (7.0) | 13.8, CH3 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mangoni, O.; Imperatore, C.; Tomas, C.R.; Costantino, V.; Saggiomo, V.; Mangoni, A. The New Carotenoid Pigment Moraxanthin Is Associated with Toxic Microalgae. Mar. Drugs 2011, 9, 242-255. https://doi.org/10.3390/md9020242
Mangoni O, Imperatore C, Tomas CR, Costantino V, Saggiomo V, Mangoni A. The New Carotenoid Pigment Moraxanthin Is Associated with Toxic Microalgae. Marine Drugs. 2011; 9(2):242-255. https://doi.org/10.3390/md9020242
Chicago/Turabian StyleMangoni, Olga, Concetta Imperatore, Carmelo R. Tomas, Valeria Costantino, Vincenzo Saggiomo, and Alfonso Mangoni. 2011. "The New Carotenoid Pigment Moraxanthin Is Associated with Toxic Microalgae" Marine Drugs 9, no. 2: 242-255. https://doi.org/10.3390/md9020242
APA StyleMangoni, O., Imperatore, C., Tomas, C. R., Costantino, V., Saggiomo, V., & Mangoni, A. (2011). The New Carotenoid Pigment Moraxanthin Is Associated with Toxic Microalgae. Marine Drugs, 9(2), 242-255. https://doi.org/10.3390/md9020242