The Chemical and Evolutionary Ecology of Tetrodotoxin (TTX) Toxicity in Terrestrial Vertebrates

Abstract
:1. Introduction
2. Background
3. TTX and TTX Analogs in Amphibians
4. Taxonomic Distribution of TTX and TTX Analogs in Amphibians
5. Ecology and Evolution of TTX Toxicity in Amphibians
6. Conclusions in Regards to the Origin/Biosynthesis of TTX in Amphibians
7. Future Directions
Acknowledgments
- Samples Availability: Available from the authors.
References and Notes
- Daly, JW. Marine toxins and nonmarine toxins: Convergence or symbiotic organisms. J Nat Prod 2004, 67(8), 1211–1215. [Google Scholar]
- Miyazawa, K; Noguchi, T. Distribution and origin of tetrodotoxin. J Toxicol: Toxin Rev 2001, 20(1), 11–33. [Google Scholar]
- Noguchi, T; Arakawa, O. Tetrodotoxin--distribution and accumulation in aquatic organisms, and cases of human intoxication. Mar Drugs 2008, 6(2), 220–242. [Google Scholar]
- Do, HK; Kogure, K; Simidu, U. Identification of deep-sea-sediment bacteria which produce tetrodotoxin. Appl Environ Microbiol 1990, 56(4), 1162–1163. [Google Scholar]
- Noguchi, T; Jeon, J.-K; Arakawa, O; Sugita, H; Deguchi, Y; Shida, Y; Hashimoto, K. Occurrence of tetrodotoxin and anhydrotetrodotoxin in Vibrio sp. isolated from the intestines of a xanthid crab Atergatis floridus. J Biochem 1986, 99, 311–314. [Google Scholar]
- Yasumoto, T; Yasumura, D; Yotsu, M; Michishita, T; Endo, A; Kotaki, Y. Bacterial production of tetrodotoxin and anhydrotetrodotoxin. Agric Biol Chem 1986, 50(3), 793–795. [Google Scholar]
- Daly, JW; Myers, CW; Whittaker, N. Further classification of skin alkaloids from neotropical poison frogs (Dendrobatidae), with a general survey of toxic/noxious substances in the amphibia. Toxicon 1987, 25(10), 1023–1095. [Google Scholar]
- Cardall, BL; Brodie, ED, Jr; Brodie, ED, III; Hanifin, CT. Secretion and regeneration of tetrodotoxin in the rough-skin newt (Taricha granulosa). Toxicon 2004, 44(8), 933–938. [Google Scholar]
- Hanifin, CT; Brodie, ED, III; Brodie, ED, Jr. Tetrodotoxin levels of the rough-skin newt, Taricha granulosa, increase in long-term captivity. Toxicon 2002, 40(8), 1149–1153. [Google Scholar]
- Lehman, EM; Brodie, ED, Jr; Brodie, ED, III. No evidence for an endosymbiotic bacterial origin of tetrodotoxin in the newt Taricha granulosa. Toxicon 2004, 44(3), 243–249. [Google Scholar]
- Kim, YH; Kim, YB; Yotsu-Yamashita, M. Potent neurotoxins: tetrodotoxin, chiriquitoxin, and zetekitoxin from Atelopus frogs in Central America. Toxin Rev 2003, 22(4), 521–532. [Google Scholar]
- Yasumoto, T; Yotsu, M; Murata, M. New tetrodotoxin analogues from the newt Cynops ensicauda. J Am Chem Soc 1988, 110, 2344. [Google Scholar]
- Yotsu, M; Iorizzi, M; Yasumoto, T. Distribution of tetrodotoxin, 6-epitetrodotoxin, and 11-deoxytetrodotoxin in newts. Toxicon 1990, 28(2), 238–241. [Google Scholar]
- Yotsu, M; Yasumoto, T; Kim, YH; Naoki, H; Kao, CY. The structure of chiriquitoxin from the Costa Rican frog Atelopus chiriquiensis. Tetrahedron Lett 1990, 31(22), 3187–3190. [Google Scholar]
- Yotsu-Yamashita, M. Chemistry of puffer fish toxin. J Toxicol: Toxin Rev 2001, 20(1), 51–66. [Google Scholar]
- Yotsu-Yamashita, M; Kim, YH; Dudley, SC, Jr; Choudhary, G; Pfahnl, A; Oshima, Y; Daly, JW. The structure of zetekitoxin AB, a saxitoxin analog from the Panamanian golden frog Atelopus zeteki: a potent sodium-channel blocker. Proc Natl Acad Sci USA 2004, 101(13), 4346–4351. [Google Scholar]
- Yotsu-Yamashita, M; Mebs, D. The levels of tetrodotoxin and its analogue 6-epitetrodotoxin in the red-spotted newt Notophthalmus viridescens. Toxicon 2001, 39(8), 1261–1263. [Google Scholar]
- Yotsu-Yamashita, M; Mebs, D; Kwet, A; Schneider, M. Tetrodotoxin and its analogue 6-epitetrodotoxin in newts (Triturus spp.; Urodela, Salamandridae) from southern Germany. Toxicon 2007, 50(2), 306–309. [Google Scholar]
- Yotsu-Yamashita, M; Tateki, E. First report on toxins in the Panamanian toads Atelopus limosus, A. glyphus and A certus. Toxicon 2010, 55(1), 153–156. [Google Scholar]
- Pires, OR, Jr; Sebben, A; Schwartz, EF; Largura, SW; Bloch, C, Jr; Morales, RA; Schwartz, CA. Occurrence of tetrodotoxin and its analogues in the Brazilian frog Brachycephalus ephippium (Anura: Brachycephalidae). Toxicon 2002, 40(6), 761–766. [Google Scholar]
- Fuhrman, FA. Tetrodotoxin, tarichatoxin, and chiriquitoxin: historical perspectives. Ann NY Acad Sci 1986, 479, 1–14. [Google Scholar]
- Kao, CY. Tetrodotoxin, saxitoxin and their significance in the study of excitation phenomena. Pharmacol Rev 1966, 18(2), 997–1049. [Google Scholar]
- Hashimoto, Y. Marine Toxins and Other Bioactive Marine Metabolites; Japan Scientific Societies Press: Tokyo, Japan, 1979; p. 369. [Google Scholar]
- Brown, MS; Mosher, H. Tarichatoxin: Isolation and purification. Science 1963, 140, 295–296. [Google Scholar]
- Buchwald, HD; Durham, L; Fischer, HG; Harada, R; Mosher, HS; Kao, CY; Fuhrman, FA. Identity of tarichatoxin and tetrodotoxin. Science 1964, 143, 474–475. [Google Scholar]
- Mosher, HS; Fuhrman, FA; Buchwald, HD; Fischer, HG. Tarichatoxin-tetrodotoxin: A potent neurotoxin. Science 1964, 144, 1100–1110. [Google Scholar]
- Narahashi, T. Tetrodotoxin: a brief history. Proc Jpn Acad, Ser B Phys Biol Sci 2008, 84(5), 147–154. [Google Scholar]
- Hille, B. Ionic Channels of Excitable Membranes; Sinauer Associates, Macmillan: Sunderland, MA, USA, 2001. [Google Scholar]
- Geffeney, S; Ruben, C. The Structural Basis and Functional Consequences of Interactions Between Tetrodotoxin and Voltage-Gated Sodium Channels. Mar Drugs 2006, 4(3), 143–156. [Google Scholar]
- Lipkind, GM; Fozzard, HA. A structural model of the tetrodotoxin and saxitoxin binding site of the Na+ channel. Biophys J 1994, 66(1), 1–13. [Google Scholar]
- Penzotti, JL; Fozzard, HA; Lipkind, GM; Dudley, SC, Jr. Differences in saxitoxin and tetrodotoxin binding revealed by mutagenesis of the Na+ channel outer vestibule. Biophys J 1998, 75(6), 2647–2657. [Google Scholar]
- Ishihara, F. Uber die physiologischen Wirkungen des Fugutoxins. Mitteilungen aus der Medizinischen Fakultat der Kaiserlichen Universitat zu Tokyo = Tokyo Teikoku Daigaku Igakubu kiyo 1918, 20, 375–426. [Google Scholar]
- Twitty, VC. Experiments on the phenomenon of paralysis produced by a toxin occurring in Triturus embryos. J Exp Zool A Comp Exp Biol 1937, 76(1), 67–104. [Google Scholar]
- Twitty, VC; Elliot, AE. The relative growth of the amphibian eye, studied by means of transplantation. J Exp Zool A Comp Exp Biol 1934, 68(2), 247–291. [Google Scholar]
- Twitty, VC; Johnson, HH. Motor inhibition in Amblystoma produced by Triturus transplants. Science 1934, 80(2064), 78–79. [Google Scholar]
- Moore, JW; Narahashi, T. Tetrodotoxin’s highly selective blockage of an ionic channel. Fed Proc 1967, 26(6), 1655–1663. [Google Scholar]
- Narahashi, T; Deguchi, T; Urakawa, M; Ohkubo, Y. Stabilization and rectification of muscle fiber membrane by tetrodotoxin. Am J Physiol Endocrinol Metabol 1960, 198, 934–938. [Google Scholar]
- Narahashi, T; Moore, JW; Scott, WR. Tetrodotoxin blockage of sodium conductance increase in lobster giant axons. J Gen Physiol 1964, 47, 965–974. [Google Scholar]
- Narahashi, T. Mechanism of action of tetrodotoxin and saxitoxin on excitable membranes. Fed Proc 1972, 31(3), 1124–1132. [Google Scholar]
- Kao, CY. Structure-activity relations of tetrodotoxin, saxitoxin, and analogues. Ann NY Acad Sci 1986, 479, 52–67. [Google Scholar]
- Kao, CY. Pharmacology of tetrodotoxin and saxitoxin. Fed Proc 1972, 31(3), 1117–1123. [Google Scholar]
- Lipkind, GM; Fozzard, HA. KcsA crystal structure as framework for a molecular model of the Na(+) channel pore. Biochemistry 2000, 39(28), 8161–8170. [Google Scholar]
- Hinman, A; Du Bois, J. A stereoselective synthesis of (−)-tetrodotoxin. J Am Chem Soc 2003, 125(38), 11510–11511. [Google Scholar]
- Yasumoto, T; Yotsu-Yamashita, M. Chemical and etiological studies on tetrodotoxin and its analogs. J Toxicol: Toxin Rev 1996, 15(2), 81–90. [Google Scholar]
- Kotaki, Y; Shimizu, Y. 1-Hydroxy-5,11-dideoxytetrodotoxin, the first N-hydroxy and ring-deoxy derivative of tetrodotoxin found in the newt Taricha granulosa. J Am Chem Soc 1993, 115(3), 827–830. [Google Scholar]
- Kim, YH; Brown, GB; Mosher, HS; Fuhrman, FA. Tetrodotoxin: Occurrence in atelopid frogs of Costa Rica. Science 1975, 189, 151–152. [Google Scholar]
- Fuhrman, FA; Fuhrman, GJ; Kim, YH; Mosher, HS. Pharmacology and chemistry of chiriquitoxin a new tetrodotoxin-like substance from the Costa Rican frog Atelopus chiriquiensis. Proc West Pharmacol Soc 1976, 19, 381–384. [Google Scholar]
- Yotsu-Yamashita, M; Mebs, D; Yasumoto, T. Tetrodotoxin and its analogues in extracts from the toad Atelopus oxyrhynchus (family: Bufonidae). Toxicon 1992, 30(11), 1489–1492. [Google Scholar]
- Kao, CY; Yeoh, PN; Goldfinger, MD; Fuhrman, FA; Mosher, HS. Chiriquitoxin, a new tool for mapping ionic channels. J Pharmacol Exp Ther 1981, 217(2), 416–429. [Google Scholar]
- Daly, JW; Kaneko, T; Wilham, J; Garraffo, HM; Spande, TF; Espinosa, A; Donnelly, MA. Bioactive alkaloids of frog skin: combinatorial bioprospecting reveals that pumiliotoxins have an arthropod source. Proc Natl Acad Sci USA 2002, 99(22), 13996–14001. [Google Scholar]
- Yotsu-Yamashita, M; Mebs, D. Occurrence of 11-oxotetrodotoxin in the red-spotted newt, Notophthalmus viridescens, and further studies on the levels of tetrodotoxin and its analogues in the newt’s efts. Toxicon 2003, 41(7), 893–897. [Google Scholar]
- Pires, OR; Sebben, A; Schwartz, EF; Bloch, C; Morales, RA; Schwartz, CA. The occurrence of 11-oxotetrodotoxin, a rare tetrodotoxin analogue, in the Brachycephalidae frog Brachycephalus ephippium. Toxicon 2003, 42(5), 563–566. [Google Scholar]
- Pires, OR, Jr; Sebben, A; Schwartz, EF; Morales, RA; Bloch, C, Jr; Schwartz, CA. Further report of the occurrence of tetrodotoxin and new analogues in the Anuran family Brachycephalidae. Toxicon 2005, 45(1), 73–79. [Google Scholar]
- Hanifin, CT; Yotsu-Yamashita, M; Yasumoto, T; Brodie, ED, III; Brodie, ED, Jr. Toxicity of dangerous prey: variation of tetrodotoxin levels within and among populations of the newt Taricha granulosa. J Chem Ecol 1999, 25(9), 2161–2175. [Google Scholar]
- Tsuruda, K; Arakawa, O; Noguchi, T. Toxicity and toxin profiles of the newt, Cynops pyrrhogaster from western Japan. J Nat Toxins 2001, 10(2), 79–89. [Google Scholar]
- Myers, GS. Notes on Pacific Coast Triturus. Copeia 2), 77–82.
- Twitty, VC. Of Scientists and Salamanders; WH Freeman & Co: San Francisco, CA, USA, 1966. [Google Scholar]
- Shindelman, J; Mosher, HS; Fuhrman, FA. Atelopidtoxin from the Panamanian frog Atelopus zeteki. Toxicon 1969, 7, 315–319. [Google Scholar]
- Wakely, JF; Fuhrman, GJ; Fuhrman, FA; Fischer, HG; Mosher, HS. The occurrence of tetrodotoxin (tarichatoxin) in amphibia and the distribution of the toxin in the organs of newts (Taricha). Toxicon 1966, 3(3), 195–203. [Google Scholar]
- Brodie, ED, Jr; Hensel, JL; Johnson, JA. Toxicity of the Urodele amphibians Taricha, Notophthalmus, Cynops, and Paramesotriton (Salamandridae). Copeia 2), 506–511.
- Tsuruda, K; Arakawa, O; Kawatsu, K; Hamano, Y; Takatani, T; Noguchi, T. Secretory glands of tetrodotoxin in the skin of the Japanese newt Cynops pyrrhogaster. Toxicon 2002, 40(2), 131–136. [Google Scholar]
- Levenson, CH; Woodhull, AM. The occurrence of a tetrodotoxin-like substance in the red-spotted newt Notophthalmus viridescens. Toxicon 1979, 17, 184–187. [Google Scholar]
- Brodie, ED, Jr. Investigations on the skin toxin of the adult rough-skinned newt Taricha granulosa. Copeia 2), 307–313.
- Hanifin, CT; Brodie, ED, III; Brodie, ED, Jr. A predictive model to estimate total skin tetrodotoxin in the newt Taricha granulosa. Toxicon 2004, 43(3), 243–249. [Google Scholar]
- Hanifin, CT; Brodie, ED, III; Brodie, ED, Jr. Tetrodotoxin levels in eggs of the rough-skin newt, Taricha granulosa, are correlated with female toxicity. J Chem Ecol 2003, 29(8), 1729–1739. [Google Scholar]
- Shimizu, Y; Kobayashi, M. Apparent lack of tetrodotoxin biosynthesis in captured Taricha torosa and Taricha granulosa. Chem Pharm Bull (Tokyo) 1983, 31, 3625–3631. [Google Scholar]
- Hanifin, CT; Brodie, ED, Jr; Brodie, ED, III. Phenotypic mismatches reveal escape from arms-race coevolution. PLoS Biol 2008, 6(3), e60. [Google Scholar]
- Twitty, VC. Taricha rivularis (Twitty) red-bellied newt. In Catalogue of American Amphibians and Reptiles; Riemer, WJ, Ed.; American Society of Ichthyologists and Herpetologists: Kensington, MD, USA, 1964; pp. 9.1–9.2. [Google Scholar]
- Sebben, A; Schwartz, CA; Valente, D; Mendes, EG. A tetrodotoxin-like substance found in the Brazilian frog Brachycephalus ephippium. Toxicon 1986, 24(6), 799–806. [Google Scholar]
- Daly, JW; Gusovsky, F; Myers, CW; Yotsu-Yamashita, M; Yasumoto, T. First occurrence of tetrodotoxin in a dendrobatid frog (Colostethus inguinalis), with further reports for the bufonid genus. Atelopus Toxicon 1994, 32(3), 279–285. [Google Scholar]
- Pavelka, LA; Kim, YH; Mosher, HS. Tetrodotoxin and tetrodotoxin-like compounds from the eggs of the Costa Rican frog Atelopus chiriquiensis. Toxicon 1977, 15, 135–139. [Google Scholar]
- Mebs, D; Schmidt, K. Occurrence of tetrodotoxin in the frog Atelopus oxyrhynchus. Toxicon 1989, 27(7), 819–822. [Google Scholar]
- Mebs, D; Yotsu-Yamashita, M; Yasumoto, T; Lotters, S; Schluter, A. Further report of the occurrence of tetrodotoxin in Atelopus species (family: Bufonidae). Toxicon 1995, 33(2), 246–249. [Google Scholar]
- Daly, JW; Padgett, WL; Saunders, RL; Cover, JF, Jr. Absence of tetrodotoxins in a captive-raised riparian frog Atelopus varius. Toxicon 1997, 35(5), 705–709. [Google Scholar]
- Brown, GB; Kim, YH; Kuntzel, H; Mosher, HS. Chemistry and pharmacology of skin toxins from the frog Atelopus zeteki (atelopidtoxin: zetekitoxin). Toxicon 1977, 15(2), 115–128. [Google Scholar]
- Tanu, MB; Mahmud, Y; Tsuruda, K; Arakawa, O; Noguchi, T. Occurrence of tetrodotoxin in the skin of a rhacophoridid frog Polypedates sp from Bangladesh. Toxicon 2001, 39(7), 937–941. [Google Scholar]
- Hirota, K; Kaneko, Y. Cloning and distribution of a putative tetrodotoxin-resistant Na+ channel in newt retina. Zool Sci 1999, 16, 587–594. [Google Scholar]
- Kaneko, Y; Matsumoto, G; Hanyu, Y. TTX resistivity of Na+ channel in newt retinal neuron. Biochem Biophys Res Commun 1997, 240, 651–656. [Google Scholar]
- Williams, BL. Behavioral and chemical ecology of marine organisms with respect to tetrodotoxin. Mar Drugs 2010, 8, 381–398. [Google Scholar]
- Brodie, ED, III; Brodie, ED, Jr. Costs of exploiting poisonous prey: evolutionary trade-offs in a predator-prey arms race. Evolution 1999, 53(2), 626–631. [Google Scholar]
- Lee, P; Ruben, PC. The Biophysical Costs Associated with Tetrodotoxin Resistance in the Garter Snake, Thamnophis sirtalis. Mar Drugs 2010, 8, submitted. [Google Scholar]
- Duellman, WE; Trueb, L. Biology of Amphibians, 1st ed; McGraw-Hill: New York, NY, USA, 1986. [Google Scholar]
- Daly, JW. The chemistry of poisons in amphibian skin. Proc Natl Acad Sci USA 1995, 92(1), 9–13. [Google Scholar]
- Brodie, ED, Jr. Antipredator adaptations of salamanders: evolution and convergence among terrestrial species. In Plant, animal, and microbial adaptations to terrestrial environment; Margaris, NS, Arianoutsu-Faraggitaki, M, Reiter, RJ, Eds.; Plenum: New York, NY, USA, 1983. [Google Scholar]
- Brodie, ED, Jr; Ducey, PK; Baness, EA. Antipredator Skin Secretions of Some Tropical Salamanders (Bolitoglossa) are Toxic to Snake Predators. Biotropica 1991, 23(1), 58–62. [Google Scholar]
- Tyler, MJ. Frog and cane toad secretions. In Toxic Plants and Animals: A Guide for Australia; Covacevich, J, Davies, P, Pearn, J, Eds.; Qld Museum: Brisbane, Australia, 1987. [Google Scholar]
- Erspammer, V. Bioactive secretions of amphibian integument. In Amphibian Biology; Heatwole, H, Barthalmus, GT, Eds.; Surrey Beatty & Sons Pty, Ltd: Chipping Norton, UK, 1994. [Google Scholar]
- Duda, TF, Jr; Vanhoye, D; Nicolas, P. Roles of diversifying selection and coordinated evolution in the evolution of amphibian antimicrobial peptides. Mol Biol Evol 2002, 19(6), 858–864. [Google Scholar]
- Erspamer, V; Erspamer, GF; Mazzanti, G; Endean, R. Active peptides in the skins of one hundred amphibian species from Australia and Papua New Guinea. Comp Biochem Physiol 1984, 77C(1), 99–108. [Google Scholar]
- Mangoni, ML; Maisetta, G; Di Luca, M; Gaddi, LM; Esin, S; Florio, W; Brancatisano, FL; Barra, D; Campa, M; Batoni, G. Comparative analysis of the bactericidal activities of amphibian peptide analogues against multidrug-resistant nosocomial bacterial strains. Antimicrob Agents Chemother 2008, 52(1), 85–91. [Google Scholar]
- Brodie, ED, Jr; Nussbaum, RA; DiGiovanni, M. Antipredator adaptations of Asian salamanders (Salamandridae). Herpetologica 1984, 40(1), 56–68. [Google Scholar]
- Johnson, JA; Brodie, ED, Jr. The selective advantage of the defensive posture of the newt Taricha granulosa. Am Midl Nat 1975, 93(1), 139–148. [Google Scholar]
- Neuwirth, M; Daly, JW; Myers, CW; Tice, LW. Morphology of the granular secretory glands in skin of poison-dart frogs (Dendrobatidae). Tissue Cell 1979, 11(4), 755–771. [Google Scholar]
- Chen, T; Farragher, S; Bjourson, AJ; Orr, DF; Rao, P; Shaw, C. Granular gland transcriptomes in stimulated amphibian skin secretions. Biochem J 2003, 371 Pt 1, 125–130. [Google Scholar]
- Kim, H.-H; Noh, Y.-T; Chung, Y.-W; Chi, Y.-D. The ultrastructure of the mucous secreting cells in the amphibians skin. Korean J Zool 1978, 21(1), 29–39. [Google Scholar]
- Elkan, E; Cooper, JE. Skin Biology of Reptiles and Amphibians. Proc R Soc Edinburgh 1980, 79b, 115–125. [Google Scholar]
- Hoffman, CW; Dent, JN. Effects of neurotransmitters upon the discharge of secretory product from the cutaneous glands of the red-spotted newt. J Exper Zool 1977, 202, 155–162. [Google Scholar]
- Toledo, RC; Jared, C. Cutaneous granular glands and amphibian venoms. Comp Biochem Physiol A: Physiol 1995, 111A(1), 1–29. [Google Scholar]
- Williams, BL; Brodie, ED, III; Brodie, ED, Jr. A resistant predator and its toxic prey: persistence of newt toxin leads to poisonous (not venomous) snakes. J Chem Ecol 2004, 30(10), 1901–1919. [Google Scholar]
- Fuhrman, FA; Fuhrman, GJ; Dull, DL; Mosher, HS. Toxins from eggs of fishes and amphibia. J Agric Food Chem 1969, 17, 417–424. [Google Scholar]
- Gregory, PT; Nelson, KJ. Predation on fish and intersite variation in the diet of common garter snakes, Thamnophis sirtalis, on Vancouver Island. Can J Zool/Rev Can Zool 1991, 69, 988–994. [Google Scholar]
- Macartney, JM; Gregory, PT. Differential susceptibility of sympatric garter snake species to amphibian skin secretions. Am Midl Nat 1981, 106(2), 271–281. [Google Scholar]
- Brandon, RA; Labanick, GM; Huheey, JE. Learned avoidence of brown efts, Notophthalmus viridescen louisianensis (Amphibia, Urodela, Salamandridae), by chickens. J Herpetol 1979, 13(2), 171–176. [Google Scholar]
- Brodie, ED, Jr. Investigations on the Skin Toxin of the Red-Spotted Newt, Notophthalmus viridescens viridescens. Am Midl Nat 1968, 80(1), 276–280. [Google Scholar]
- Brodie, ED, Jr; Formanowicz, DR, Jr. Larvae of the predaceous diving beetle Dytiscus verticalis acquire an avoidance response to skin secretions of the newt Notophthalmus viridescens. Herpetologica 1981, 37(3), 172–176. [Google Scholar]
- Hurlbert, SH. Predator Responses to the Vermilion-Spotted Newt (Notophthalmus viridescens). J Herpetol 1970, 4(1/2), 47–55. [Google Scholar]
- Pough, FH. Leech-repellent property of eastern red-spotted newts, Notophthalmus viridescens. Science 1971, 174, 1144–1146. [Google Scholar]
- Shure, DJ; Wilson, LA; Hochwender, C. Predation on aposematic efts of Notophthalmus viridescens. J Herpetol 1989, 23(6), 437–439. [Google Scholar]
- Webster, DA. Toxicity of the spotted newt, Notophthalmus viridescens, to trout. Copeia 1), 2.
- Brodie, ED, III; Brodie, ED, Jr. Tetrodotoxin resistance in garter snakes - an evolutionary response of predators to dangerous prey. Evolution 1990, 44(3), 651–659. [Google Scholar]
- Brodie, ED, Jr; Ridenhour, BJ; Brodie, ED, III. The evolutionary response of predators to dangerous prey: hotspots and coldspots in the geographic mosaic of coevolution between garter snakes and newts. Evolution 2002, 56(10), 2067–2082. [Google Scholar]
- Feldman, CR; Brodie, ED, Jr; Brodie, ED, 3rd; Pfrender, ME. The evolutionary origins of beneficial alleles during the repeated adaptation of garter snakes to deadly prey. Proc Natl Acad Sci USA 2009, 106(32), 13415–13420. [Google Scholar]
- Geffeney, SL; Fujimoto, E; Brodie, ED, III; Brodie, ED, Jr; Ruben, PC. Evolutionary diversification of TTX-resistant sodium channels in a predator-prey interaction. Nature 2005, 434(7034), 759–763. [Google Scholar]
- Brodie, ED, III; Brodie, ED, Jr. Evolutionary response of predators to dangerous prey - reduction of toxicity of newts and resistance of garter snakes in island populations. Evolution 1991, 45(1), 221–224. [Google Scholar]
- Brodie, ED, III; Feldman, CR; Hanifin, CT; Motychak, JE; Mulcahy, DG; Williams, BL; Brodie, ED, Jr. Parallel arms races between garter snakes and newts involving tetrodotoxin as the phenotypic interface of coevolution. J Chem Ecol 2005, 31(2), 343–356. [Google Scholar]
- Geffeney, SL; Brodie, ED, Jr; Ruben, PC; Brodie, ED, III. Mechanisms of adaptation in a predator-prey arms race: TTX-resistant sodium channels. Science 2002, 297(5585), 1336–1339. [Google Scholar]
- Gall, BG; Brodie, ED, III; Brodie, ED, Jr. Predator avoidance during oviposition: female newts avoid depositing eggs near invertebrate predators. Copeia 2010. submitted. [Google Scholar]
- Zimmer, RK; Ferrer, RP. Neuroecology, chemical defense, and the keystone species concept. Biol Bull Mar Biol Lab Woods Hole 2007, 213(3), 208–225. [Google Scholar]
- Kuchta, SR. Experimental support for aposematic coloration in the salamander Ensatina eschscholtzii xanthoptica: Implications for mimicry of Pacific Newts. Copeia 2005, (2), 265–271. [Google Scholar]
- Kuchta, SR; Krakauer, AH; Sinervo, B. Why does the yellow-eyed Ensatina have yellow eyes? Batesian mimicry of Pacific newts (genus Taricha) by the salamander Ensatina eschscholtzii xanthoptica. Evolution 2008, 62(4), 984–990. [Google Scholar]
- Howard, RR; Brodie, ED. Experimental study of mimicry in salamanders involving Notophthalmus viridescens viridescens and Pseudotriton ruber schencki. Nature 1971, 233(5317), 277. [Google Scholar]
- Huheey, JE; Brandon, RA. Studies in warning coloration and mimicry. VI. Comments on the warning coloration of red efts and their presumed mimicry by red salamanders. Herpetologica 1974, 30(2), 149–155. [Google Scholar]
- Ferrer, RP; Zimmer, RK. Chemical neuroecology and community dynamics. Ann NY Acad Sci 2009, 1170, 450–455. [Google Scholar]
- Zimmer, RK; Schar, DW; Ferrer, RP; Krug, PJ; Kats, LB; Michel, WC. The scent of danger: Tetrodotoxin (TTX) as an olfactory cue of predation risk. Ecol Monogr 2006, 76(4), 585–600. [Google Scholar]
- Chau, R; Kalaitzis, JA; Neilan, BA. Biosynthesis of tetrodotoxin and structurally related molecules. Mar Drugs 2010, 8, submitted. [Google Scholar]
- Matsumura, K. Reexamination of tetrodotoxin production by bacteria. Appl Environ Microbiol 1995, 61, 3468–3470. [Google Scholar]
- Janzen, FJ; Krenz, JG; Haselkorn, TS; Brodie, ED, Jr; Brodie, ED, III. Molecular phylogeography of common garter snakes (Thamnophis sirtalis) in western North America: implications for regional historical forces. Mol Ecol 2002, 11(9), 1739–1751. [Google Scholar]
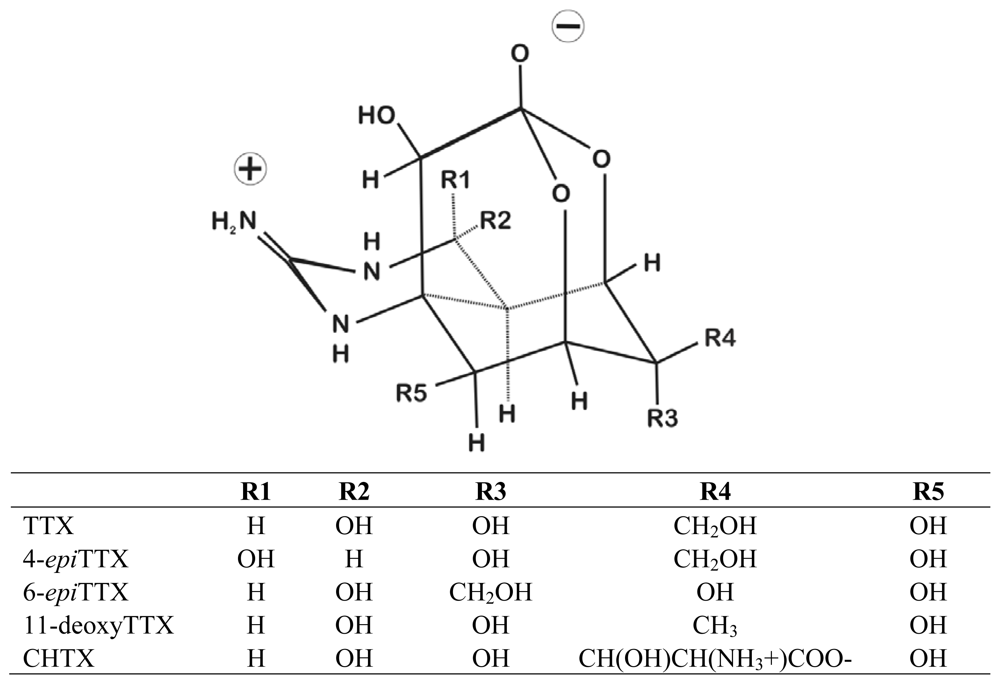

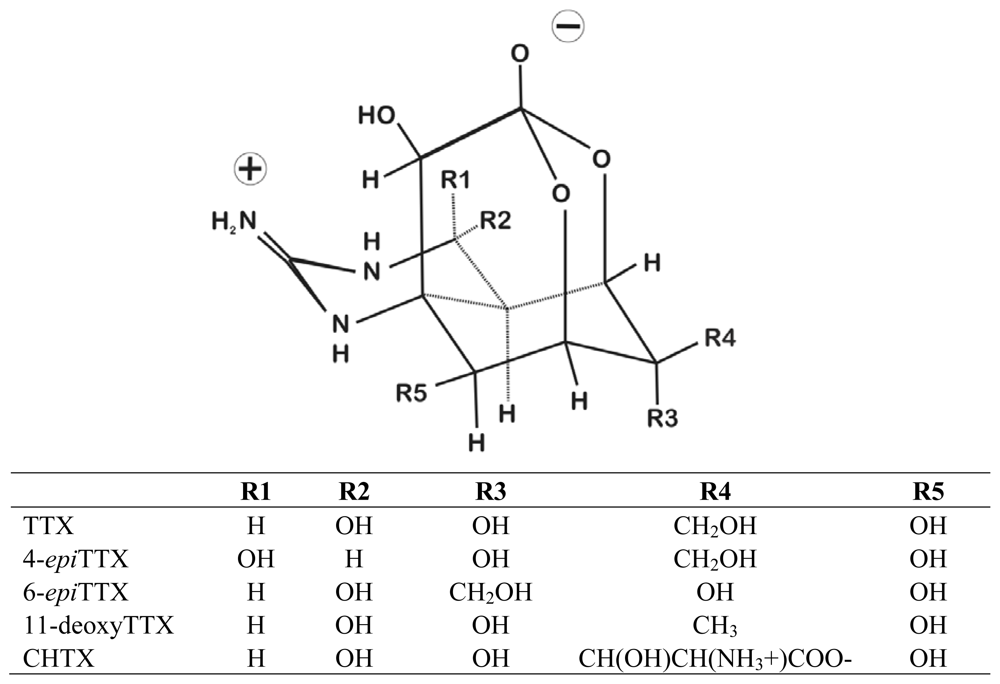{kind=link}
| Order | Family, Species | Primary Toxin | Estimated amount of TTX (or equivalents) per individual (ug) | Additional Analogs | References |
|---|---|---|---|---|---|
| Caudata | Ambystomatidae | ||||
| Ambystoma tigrinum | TTX | 12.6–17.6 | 6-epiTTX, 11-deoxyTTX | [13] | |
| Salamandridae | |||||
| Cynops ensicauda | TTX | 9.6–1540 | 6-epiTTX, 11-deoxyTTX | [12,13,26,59] | |
| Cynops pyrrhogaster | TTX | 8–616 | 6-epiTTX, 11-deoxyTTX | [13,26,55,59–61] | |
| Notophthalmus viridescens | TTX | 9.6–220 | 6-epiTTX, 11-deoxyTTX | [17,18,26,51,59,60,62] | |
| Paramesotriton hongkongensis | TTX | 8–11 | [13,60] | ||
| Taricha granulosa | TTX | <1–14,000 | 6-epiTTX, 11-deoxyTTX | [8,9,13,24–26,45,54,59,60,63–67] | |
| Taricha rivularis | TTX | 96–550 | [56,59,60,68] | ||
| Taricha torosa | TTX | <1–3000 | 6-epiTTX | [59,60,66,67] | |
| Triturus alpestris | TTX | 0–41 | 6-epiTTX | [13,18,59] | |
| Triturus cristatus | TTX | 0–9 | 6-epiTTX | [18,59] | |
| Triturus helveticus | TTX | 0–8 | 6-epiTTX | [18] | |
| Triturus marmoratus | TTX | 0.16–0.66 | [26,59] | ||
| Triturus vulgaris | TTX | 0–8 | 6-epiTTX, 11-deoxyTTX | [13,18,59] | |
| Anura | Brachycephalidae | ||||
| Brachycephalus ephippium | TTX | <1–22.4 | 6-epiTTX, 11-deoxyTTX | [20,52,53,69] | |
| Brachycephalus pernix | TTX | 5 | [53] | ||
| Dendrobatidae | |||||
| Colostethus inquinalis | TTX | 0.1–1.4 | [70] | ||
| Bufonidae | |||||
| Atelopus chiriquiensis | CHTX | 33 (TTX), 77 (CHTX) | TTX | [14,19,46,47,71] | |
| Atelopus glyphus | CHTX | 34–79 | [19] | ||
| Atelopus ignescens | TTX | <1.0–1.5 | [70] | ||
| Atelopus limosus | CHTX | 8–19 | [19] | ||
| Atelopus oxyrhynchus | TTX | 32–198 | [48,72] | ||
| Atelopus peruensis | TTX | 3.2–4.4 | [73] | ||
| Atelopus spumarius | TTX | 1.6–3.5 | [70] | ||
| Atelopus spurelli | TTX | <1–1.1 | [70] | ||
| Atelopus subornatus | TTX | 3.2–17.6 | [73] | ||
| Atelopus varius | TTX | 16–26 | [19,46,70,74] | ||
| Atelopus zeteki | ZTX | <1–264 | [19,46,58,70,75] | ||
| Rhacophoridae | |||||
| Polypedates sp. | TTX | 4.8–198 | [76] |
| Order | Family, Species | Reference |
|---|---|---|
| Caudata | Ambystomatidae | |
| Ambystoma tigrinum | [59] | |
| Amphiumadae | ||
| Amphiuma means | [59] | |
| Cryptobranchidae | ||
| Cryptobranchus alleganiensis Plethidontidae | [59] | |
| Batrachoseps attenuatus | [59] | |
| Ensatina eschscholtzi | [59] | |
| Aneides lugubris | [59] | |
| Proteidae | ||
| Necturus maculosus | [59] | |
| Salamandridae | ||
| Salamandra salamandra | [59] | |
| Echinotriton andersoni | From Miyazawa, 2001[2] | |
| Anura | Bufonidae | |
| Atelopus certus | [19] | |
| Bufo boreas | [59] | |
| Bufo regularis | [70] | |
| Dendrophyryniscus minutus | [70,73] | |
| Melanophryniscus moreirae | [70] | |
| Melanophrynicus stelzneri | [73] | |
| Oreophrynella sp. | [73] | |
| Denrobatidae | ||
| Aromabates nocturnus | [70] | |
| Dendrobates pumilio | [70] | |
| Phyllobates bicolor | [70] | |
| Hylidae | ||
| Cyclorana australis | [70] | |
| Hemiphractus fasciatus | [70] | |
| Hyla cinera | [59] | |
| Litoria albuguttata | [70] | |
| Nyctimystes tympanocryptis | [70] | |
| Osteocephalus taurinus | [70] | |
| Phrynohyas venulosa Leptodactylidae | [70] | |
| Eleutherodactylus gollmeri Microhylidae | [70] | |
| Otophryne robusta | [70] | |
| Phrynomerus bifasciatus | [70] | |
| Scaphiophryne marmorata | [70] | |
| Mantellidae | ||
| Mantella aurantiaca Myobatrachidae | [70] | |
| Heleioporus albopunctatus | [70] | |
| Notaden nichollsi | [70] | |
| Pseudophryne corroboree | [70] | |
| Pipidae | ||
| Xenopus laevis | [59] | |
| Ranidae | ||
| Rana pipiens | [59] | |
| Rana rugulosa | [70] | |
| Rana septentrionalis | [70] |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hanifin, C.T. The Chemical and Evolutionary Ecology of Tetrodotoxin (TTX) Toxicity in Terrestrial Vertebrates. Mar. Drugs 2010, 8, 577-593. https://doi.org/10.3390/md8030577
Hanifin CT. The Chemical and Evolutionary Ecology of Tetrodotoxin (TTX) Toxicity in Terrestrial Vertebrates. Marine Drugs. 2010; 8(3):577-593. https://doi.org/10.3390/md8030577
Chicago/Turabian StyleHanifin, Charles T. 2010. "The Chemical and Evolutionary Ecology of Tetrodotoxin (TTX) Toxicity in Terrestrial Vertebrates" Marine Drugs 8, no. 3: 577-593. https://doi.org/10.3390/md8030577
APA StyleHanifin, C. T. (2010). The Chemical and Evolutionary Ecology of Tetrodotoxin (TTX) Toxicity in Terrestrial Vertebrates. Marine Drugs, 8(3), 577-593. https://doi.org/10.3390/md8030577