Bioactive Sesterterpenes and Triterpenes from Marine Sponges: Occurrence and Pharmacological Significance




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Sesterterpenes (C25)
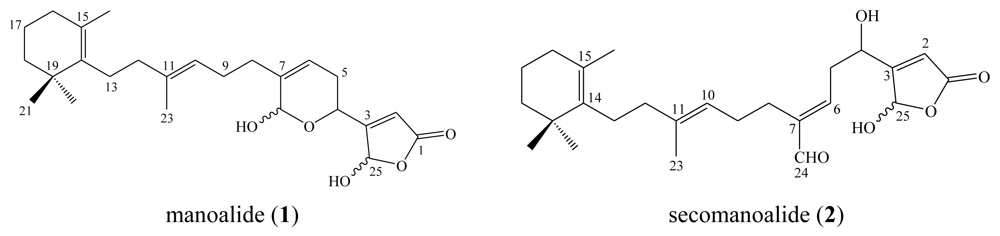

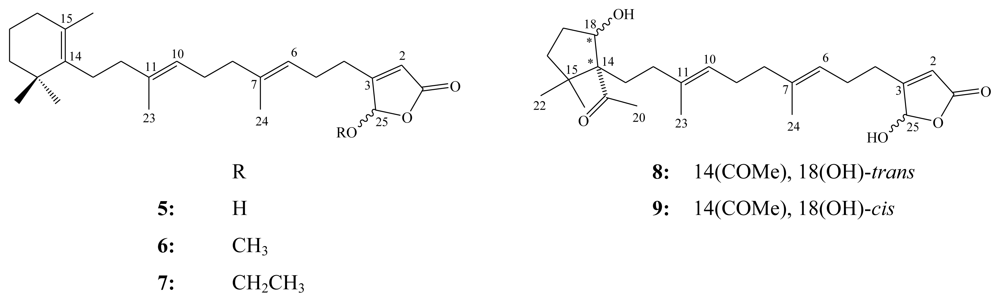

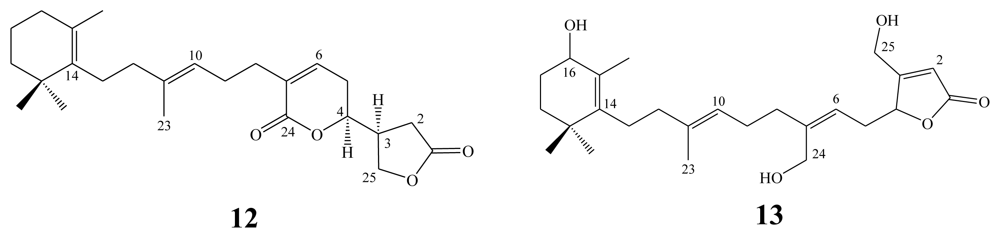
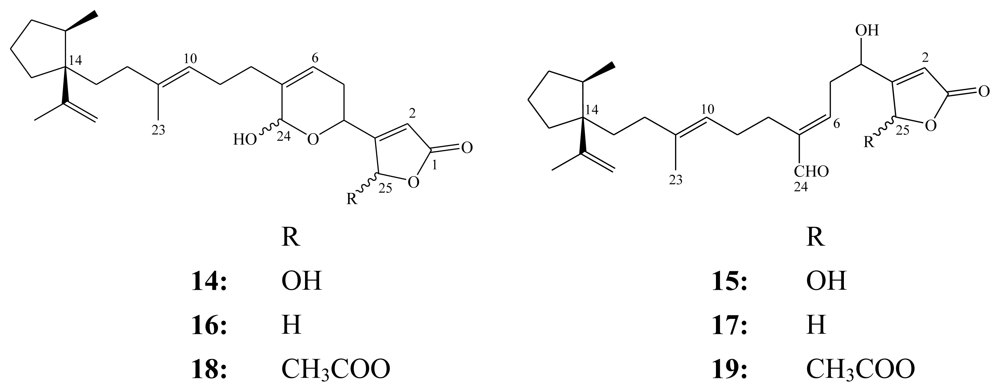

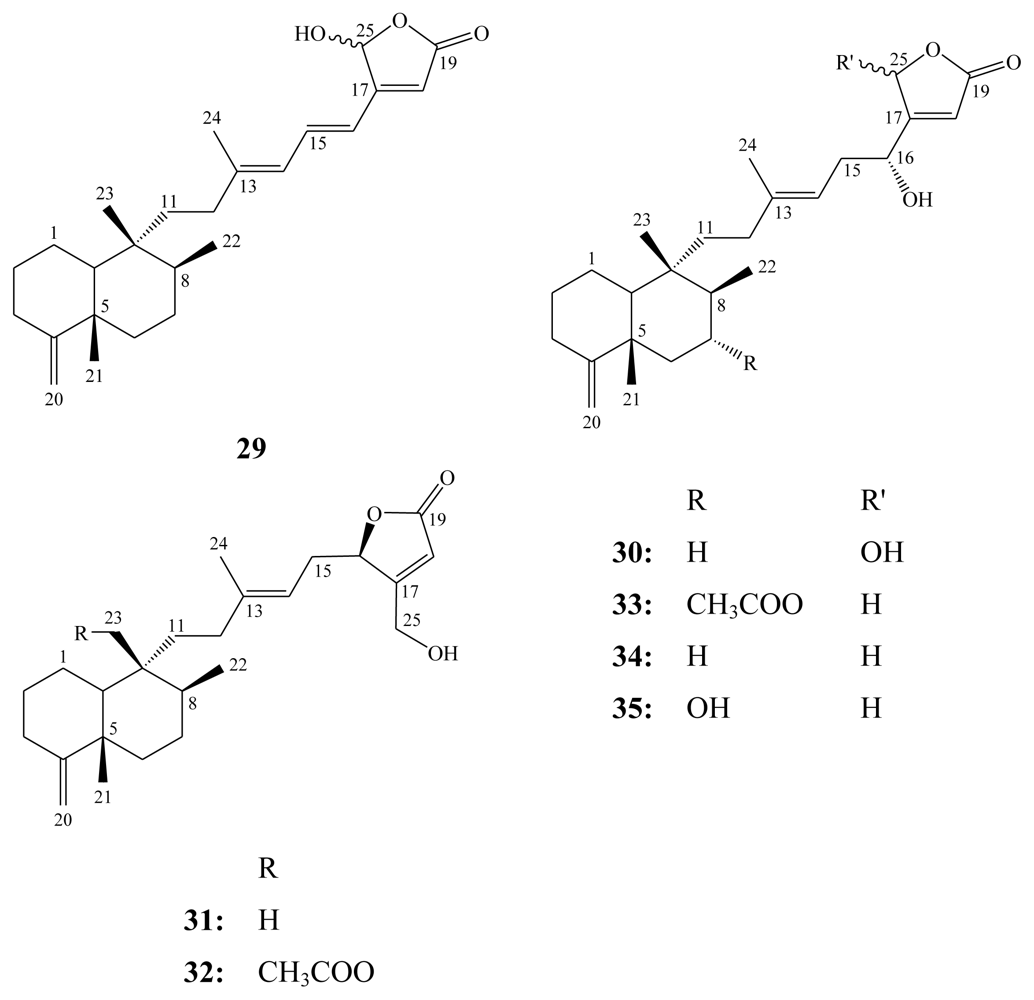


3. Triterpenes (C30)
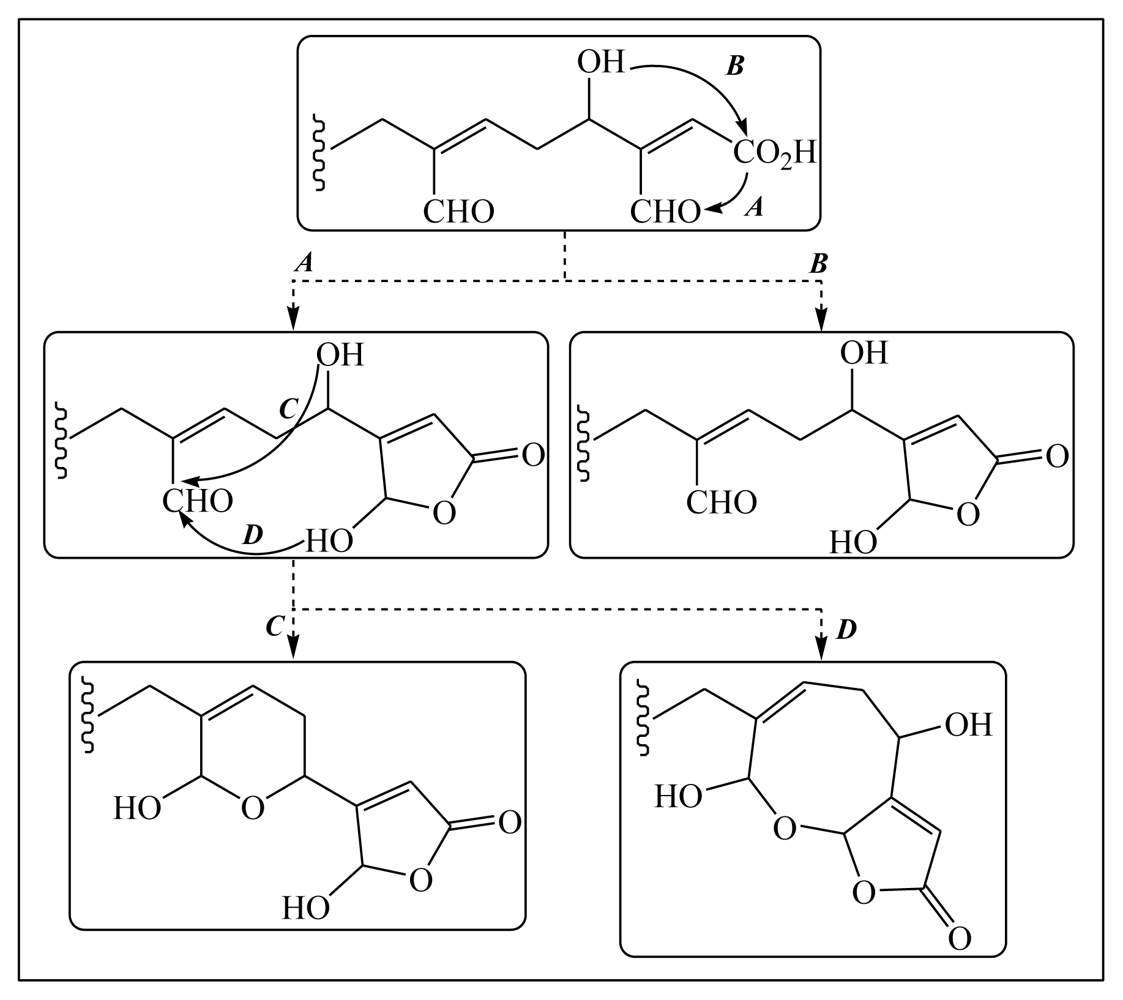
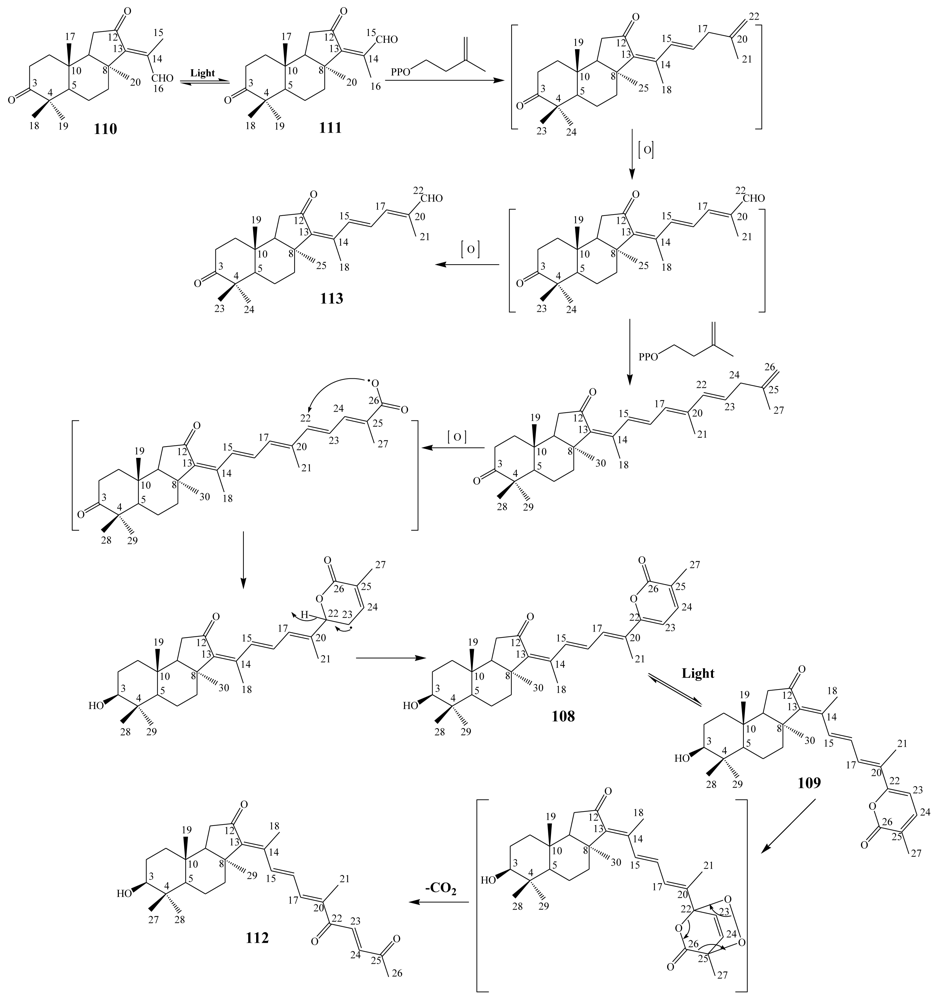
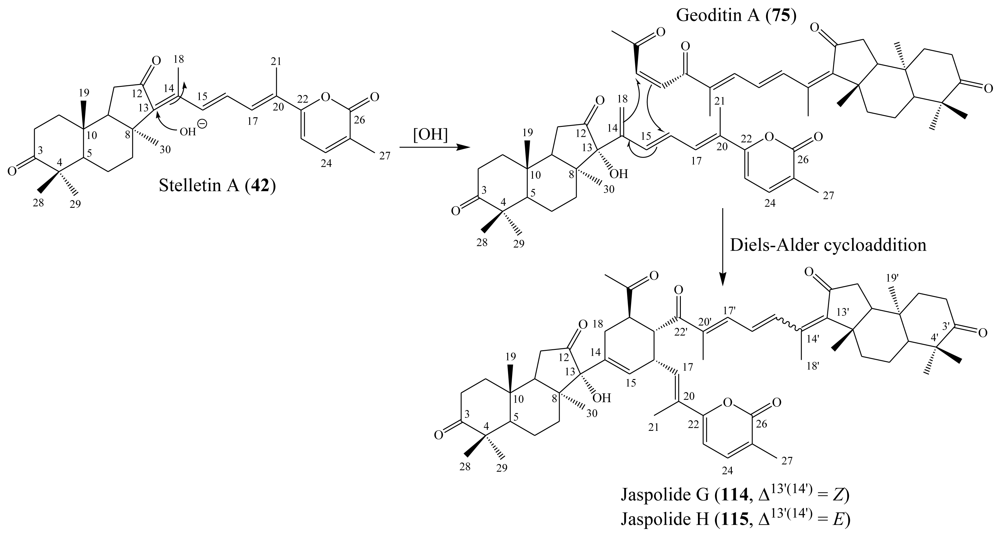
3.1. Isomalabaricane triterpenes

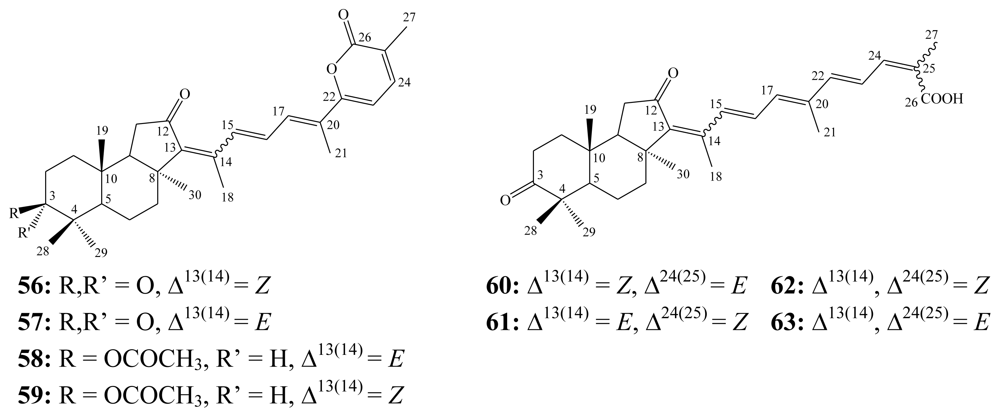
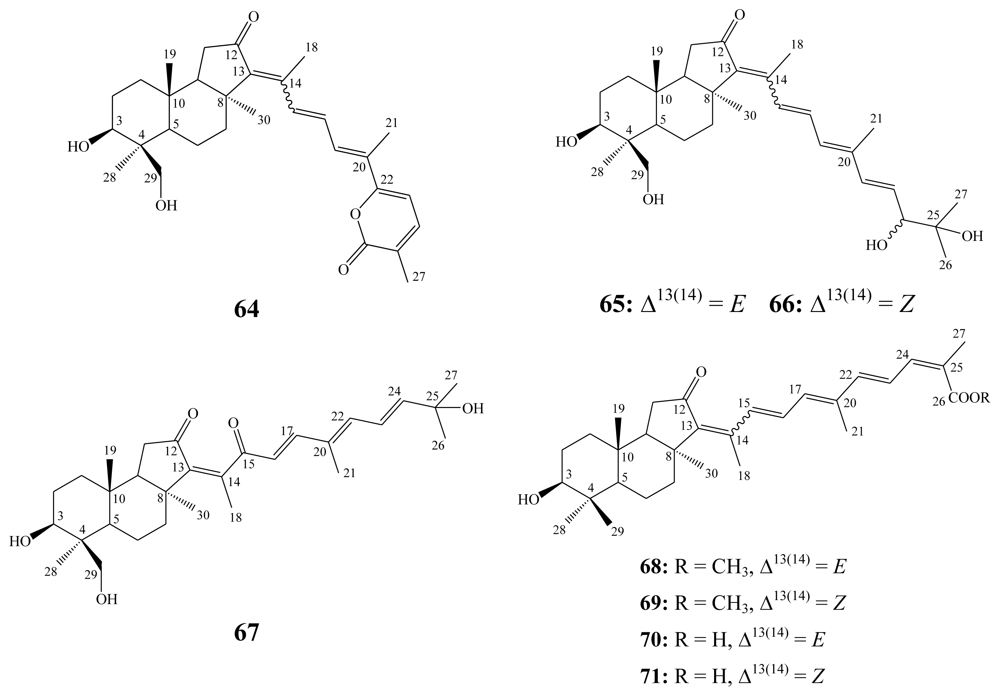

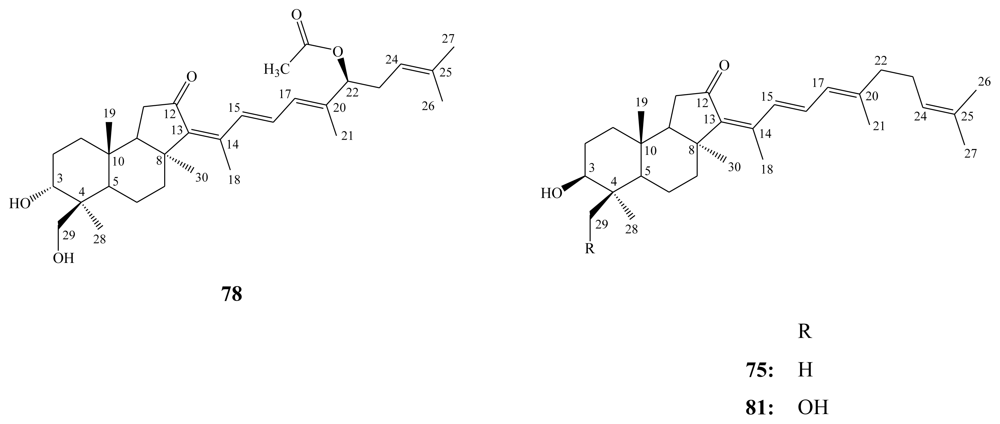
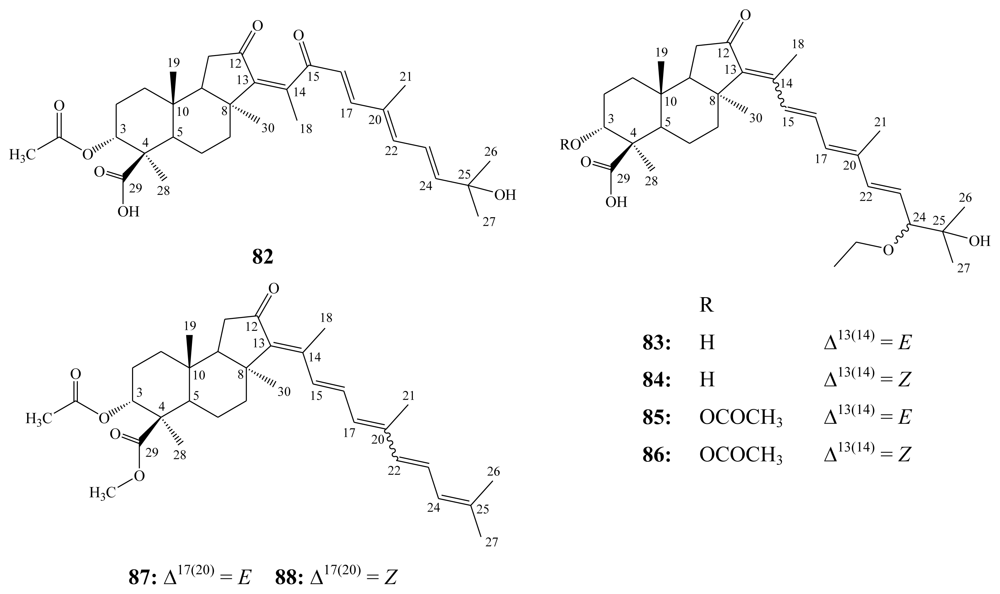

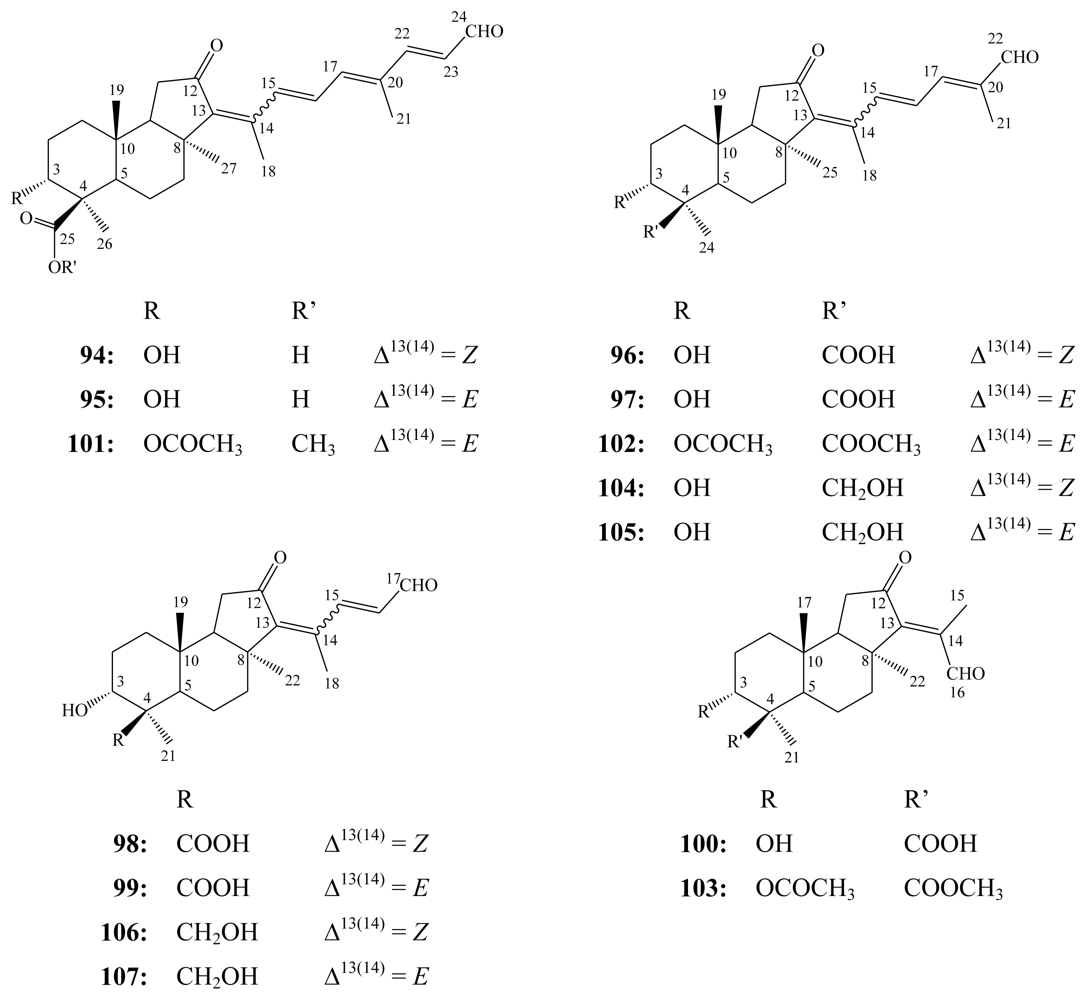
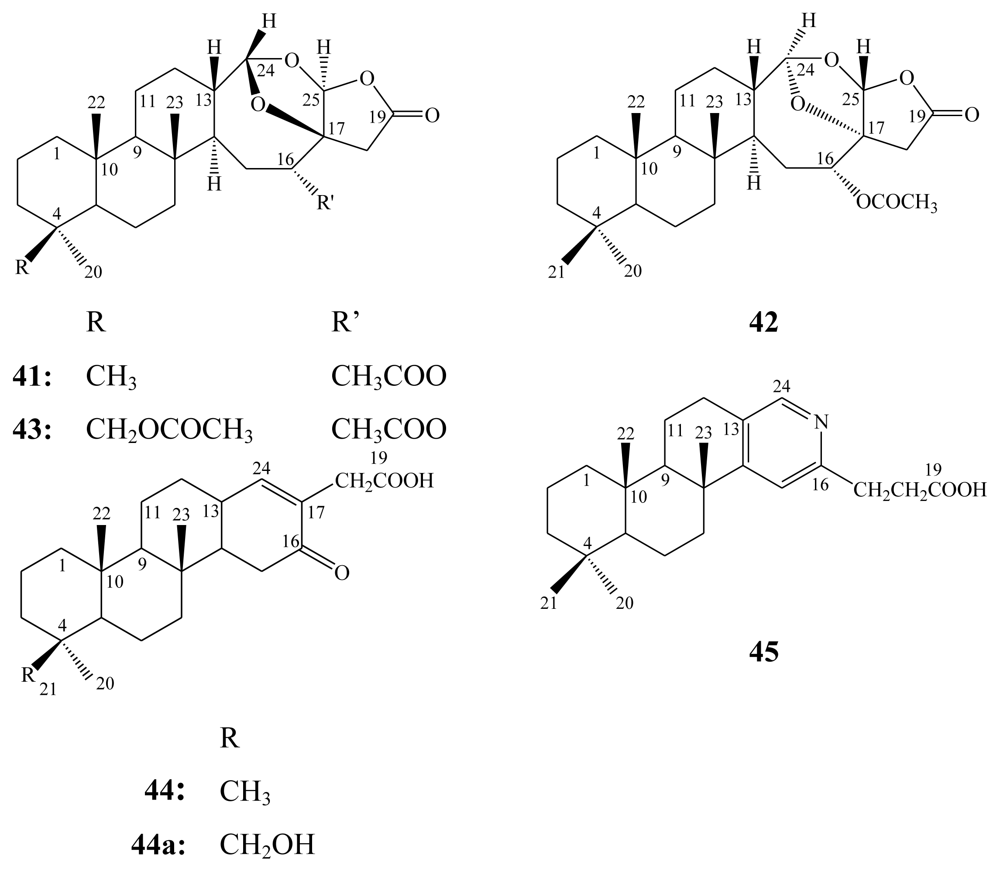
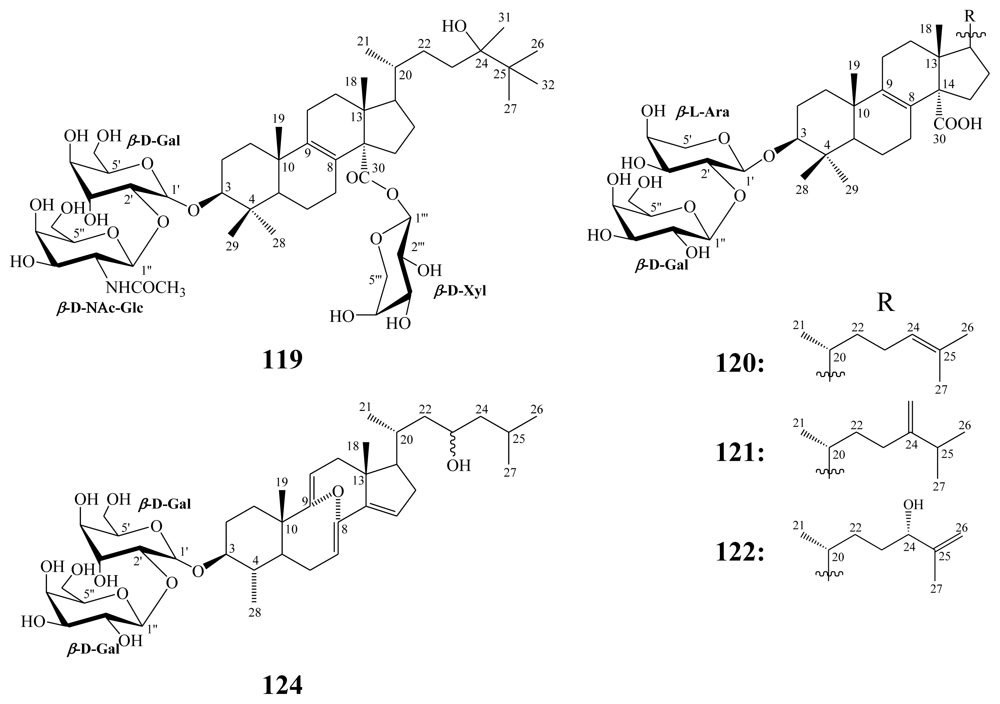
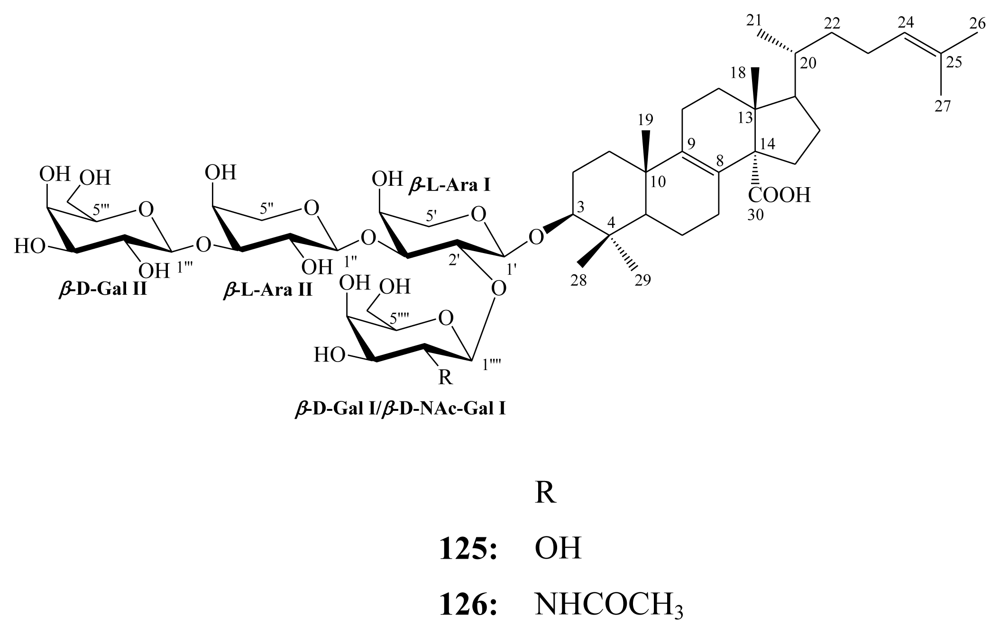
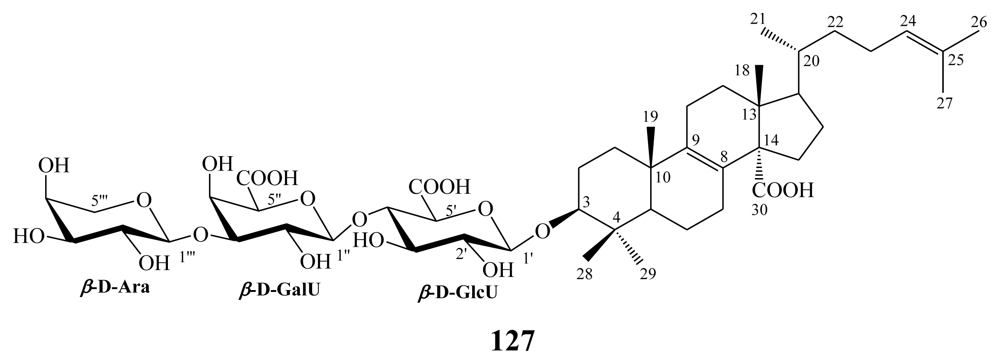
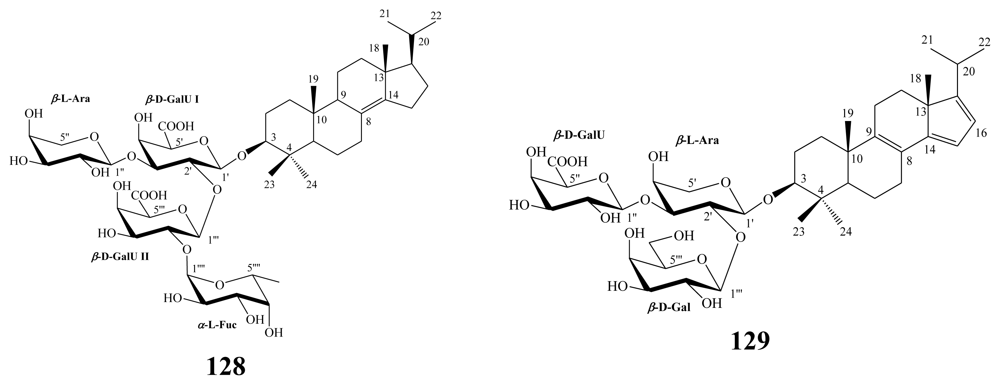
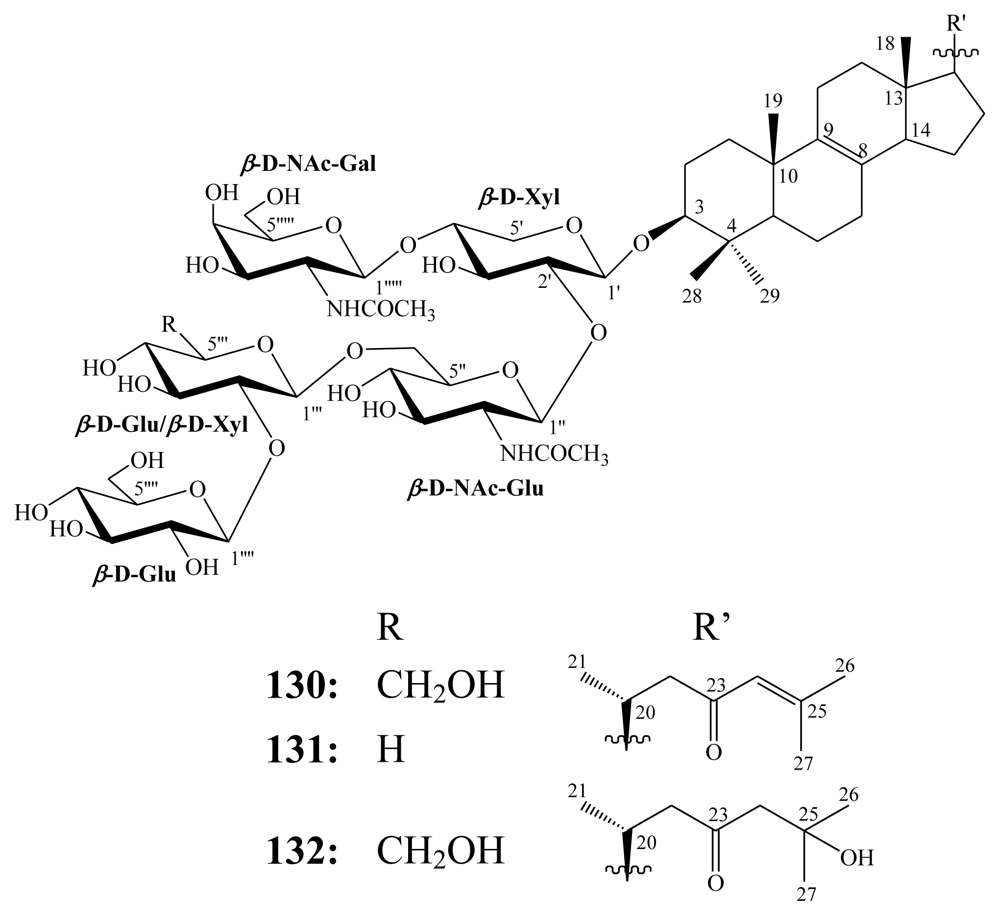
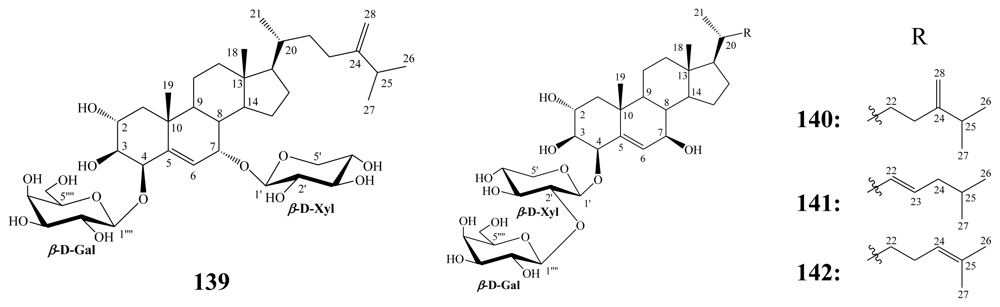
3.2. Steroidal saponins



4. Future Aspects
Acknowledgements
- Samples Availability: Available from the authors.
References and Notes
- Braekman, JC; Daloze, D. Chemical defence in sponges. Pure Appl Chem 1986, 58, 357–364. [Google Scholar]
- Ruzicka, ZL. The isoprene rule and the biogenesis of terpenic compounds. Experientia 1953, 9, 357–367. [Google Scholar]
- Bergmann, W; Johnson, TB. The chemistry of marine animals. I. The sponge Microciona pralifera. Z Physiol Chem 1933, 222, 220–226. [Google Scholar]
- Fusetani, N. Biofouling and antifouling. Nat Prod Rep 2004, 21, 94–104. [Google Scholar]
- De Silva, ED; Scheuer, PJ. Manoalide, an antibiotic sesterterpenoid from the marine sponge Luffariella variabilis (Polejaeff). Tetrahedron Lett 1980, 21, 1611–1614. [Google Scholar]
- De Silva, ED; Scheuer, PJ. Three new sesterterpenoid antibiotics from the marine sponge Luffariella variabilis (Polejaeff). Tetrahedron Lett 1981, 22, 3147–3150. [Google Scholar]
- Kernan, MR; Faulkner, DJ; Jacobs, RS. The luffariellins, novel anti-inflammatory sesterterpenes of chemotaxonomic importance from the marine sponge Luffariella variabilis. J Org Chem 1987, 52, 3081–3083. [Google Scholar]
- Albizati, KF; Holman, T; Faulkner, DJ; Glaser, KB; Jacobs, RS. Luffariellolide, an anti-inflammatory sesterterpene from the marine sponge Luffariella sp. Experientia 1987, 43, 949–950. [Google Scholar]
- Kernan, MR; Faulkner, DJ; Parkanyi, L; Clardy, J; de Carvalho, MS; Jacobs, RS. Luffolide, a novel anti-inflammatory terpene from the sponge Luffariella sp. Experientia 1989, 45, 388–390. [Google Scholar]
- König, GM; Wright, AD; Sticher, O. Four new antibacterial sesterterpenes from a marine sponge of the genus Luffariella. J Nat Prod 1992, 55, 174–178. [Google Scholar]
- Potts, BCM; Capon, RJ; Faulkner, DJ. Luffalactone and (4E,6E)-dehydromanoalide from the sponge Luffariella variabilis. J Org Chem 1992, 57, 2965–2967. [Google Scholar]
- Butler, MS; Capon, RJ. The luffarins (A–Z), novel terpenes from an Australian marine sponge Luffariella geometrica. Aust J Chem 1992, 45, 1705–1743. [Google Scholar]
- Tsuda, M; Shigemori, H; Ishibashi, M; Sasaki, T; Kobayashi, J. Luffariolides A–E, new cytotoxic sesterterpenes from the Okinawan marine sponge Luffariella sp. J Org Chem 1992, 57, 3503–3507. [Google Scholar]
- Kobayashi, J; Zeng, CM; Ishibashi, M; Sasaki, T. Luffariolides F and G, new manoalide derivatives from the Okinawan marine sponge Luffariella sp. J Nat Prod 1993, 56, 436–439. [Google Scholar]
- Reddy, MVR; Harper, MK; Faulkner, DJ. Luffasterols A–C, 9,11-secosterols from the Palauan sponge Luffariella sp. J Nat Prod 1997, 60, 41–43. [Google Scholar]
- Tsuda, M; Endo, T; Mikami, Y; Fromont, J; Kobayashi, J. Luffariolides H and J, new sesterterpenes from a marine sponge Luffariella. J Nat Prod 2002, 65, 1507–1508. [Google Scholar]
- Namikoshi, M; Suzuki, S; Meguro, S; Nagai, H; Koike, Y; Kitazawa, A; Kobayashi, H; Oda, T; Yamada, J. Manoalide derivatives from a marine sponge Luffariella sp. collected in Palau. Fish Sci 2004, 70, 152–158. [Google Scholar]
- Ettinger-Epstein, P; Motti, CA; de Nys, R; Wright, AD; Battershill, CN; Tapiolas, DM. Acetylated sesterterpenes from the Great Barrier reef sponge Luffariella variabilis. J Nat Prod 2007, 70, 648–651. [Google Scholar]
- Gauvin-Bialecki, A; Aknin, M; Smadja, J. 24-O-Ethylmanoalide, a manoalide-related sesterterpene from the marine sponge Luffariella cf. variabilis. Molecules 2008, 13, 3184–3191. [Google Scholar]
- Kobayashi, M; Okamoto, T; Hayashi, K; Yokoyama, N; Sasaki, T; Kitagawa, I. Marine natural products. XXXII. Absolute configurations of C-4 of the manoalide family, biologically active sesterterpenes from the marine sponge Hyrtios erecta. Chem Pharm Bull 1994, 42, 265–270. [Google Scholar]
- Bourguet-Kondracki, ML; Debitus, C; Guyot, M. Biologically active sesterterpenes from a new Caledonian marine sponge Hyrtios sp. J Chem Res 1996, 192–193. [Google Scholar]
- Cambie, RC; Craw, PA; Bergquist, PR; Karuso, P. Chemistry of sponges, III. Manoalide monoacetate and thorectolide monoacetate, two new seterterpenoids from Thorectandra excavatus. J Nat Prod 1988, 51, 331–334. [Google Scholar]
- De Rosa, S; de Stefano, S; Zavodnik, N. Cacospongionolide: a new antitumoral sesterterpene, from the marine sponge Cacospongia mollior. J Org Chem 1988, 53, 5020–5023. [Google Scholar]
- Tasdemir, D; Concepción, GP; Mangalindan, GC; Harper, MK; Hajdu, E; Ireland, CM. New terpenoids from a Cacospongia sp. from the Philippines. Tetrahedron 2000, 56, 9025–9030. [Google Scholar]
- Montagnac, A; Païs, M; Debitus, C. Fasciospongides A, B and C, new manoalide derivatives from the sponge Fasciospongia sp. J Nat Prod 1994, 57, 186–190. [Google Scholar]
- De Rosa, S; Crispino, A; de Giulio, A; Iodice, C; Pronzato, R; Zavodnik, N. Cacospongionolide B, a new sesterterpene from the sponge Fasciospongia cavernosa. J Nat Prod 1995, 58, 1776–1780. [Google Scholar]
- De Rosa, S; Crispino, A; de Giulio, A; Iodice, C; Tommonaro, G. Cavernosolide, a new sesterterpene from a Tyrrhenian sponge. J Nat Prod 1997, 60, 844–846. [Google Scholar]
- De Rosa, S; Carbonelli, S. Two new luffarin derivatives from the Adriatic Sea sponge Fasciospongia cavernosa. Tetrahedron 2006, 62, 2845–2849. [Google Scholar]
- Elkhayat, E; Edrada, RA; Ebel, R; Wray, V; van Soest, R; Wiryowidagdo, S; Mohamed, MH; Müller, WEG; Proksch, P. New luffariellolide derivatives from the Indonesian sponge Acanthodendrilla sp. J Nat Prod 2004, 67, 1809–1817. [Google Scholar]
- Ueoka, R; Nakao, Y; Fujii, S; van Soest, RWM; Matsunaga, S. Aplysinoplides A–C, cytotoxic sesterterpnes from the marine sponge Aplysinopsis digitata. J Nat Prod 2008, 71, 1089–1091. [Google Scholar]
- De Freitas, JC; Blankemeier, LA; Jacobs, RS. In vitro inactivation of the neurotoxic action of β-bungarotoxin by the marine natural product, manoalide. Experientia 1984, 40, 864–865. [Google Scholar]
- Lombardo, D; Dennis, EA. Cobra venom phospholipase A2 inhibition by manoalide. A novel type of phospholipase inhibitor. J Biol Chem 1985, 260, 7234–7240. [Google Scholar]
- Glaser, KB; Jacobs, RS. Molecular pharmacology of manoalide. Inactivation of bee venom phospholipase A2. Biochem Pharmacol 1986, 35, 449–453. [Google Scholar]
- Glaser, KB; Jacobs, RS. Inactivation of bee venom phospholipase A2 by manoalide. A model based on the reactivity of manoalide with amino acids and peptide sequences. Biochem Pharmacol 1987, 36, 2079–2086. [Google Scholar]
- Bennett, CF; Mong, S; Clarke, MA; Kruse, LI; Crooke, ST. Differential effects of manoalide on secreted and intracellular phospholipases. Biochem Pharmacol 1987, 36, 733–740. [Google Scholar]
- Glaser, KB; de Carvalho, MS; Jacobs, RS; Kernan, MR; Faulkner, DJ. Manoalide: structure-activity studies and definition of the pharmacophore for phospholipase A2 inactivation. Mol Pharmacol 1989, 36, 782–788. [Google Scholar]
- Jacobson, PB; Marshall, LA; Sung, A; Jacobs, RS. Inactivation of human synovial fluid phospholipase A2 by the marine natural product, manoalide. Biochem Pharmacol 1990, 39, 1557–1564. [Google Scholar]
- Ortiz, AR; Pisabarro, MT; Gago, F. Molecular model of the interaction of bee venom phospholipase A2 with manoalide. J Med Chem 1993, 36, 1866–1879. [Google Scholar]
- Deems, RA; Lombardo, D; Morgan, BP; Mihelich, ED; Dennis, EA. The inhibition of phospholipase A2 by manoalide and manoalide analogues. Biochim Biophys Acta 1987, 917, 258–268. [Google Scholar]
- Reynolds, LJ; Morgan, BP; Hite, GA; Mihelich, ED; Dennis, EA. Phospholipase A2 inhibition and modification by manoalogue. J Am Chem Soc 1988, 110, 5172–5177. [Google Scholar]
- Potts, BCM; Faulkner, DJ; Jacobs, RS. Phospholipase A2 inhibitors from marine organisms. J Nat Prod 1992, 55, 1701–1717. [Google Scholar]
- Reynolds, LJ; Mihelich, ED; Dennis, EA. Inhibition of venom phospholipase A2 by manoalide and manoalogue. J Biol Chem 1991, 266, 16512–16517. [Google Scholar]
- Potts, BCM; Faulkner, DJ; de Carvalho, MS; Jacobs, RS. Chemical mechanism of inactivation of bee venom phospholipase A2 by the marine natural products manoalide, luffariellolide, and scalaradial. J Am Chem Soc 1992, 114, 5093–5100. [Google Scholar]
- De Rosa, M; Giordano, S; Scettri, A; Sodano, G; Soriente, A; Pastor, PG; Alcaraz, MJ; Payá, M. Synthesis and comparison of the anti-inflammatory activity of manoalide and cacospongionolide B analogues. J Med Chem 1998, 41, 3232–3238. [Google Scholar]
- Mann, J. Sponges to wipe away pain. Nature 1992, 358, 540. [Google Scholar]
- Blanchard, JL; Epstein, DM; Boisclair, MD; Rudolph, J; Pal, K. Dysidiolide and related γ-hydroxybutenolide compounds as inhibitors of the protein tyrosine phosphatase, CDC25. Bioorg Med Chem Lett 1999, 9, 2537–2538. [Google Scholar]
- Charan, RD; McKee, TC; Boyd, MR. Thorectandrols A and B, new cytotoxic sesterterpenes from the marine sponge Thorectandra species. J Nat Prod 2001, 64, 661–663. [Google Scholar]
- Charan, RD; McKee, TC; Boyd, MR. Thorectandols C, D, and E, new sesterterpenes from the marine sponge Thorectandra sp. J Nat Prod 2002, 65, 492–495. [Google Scholar]
- De Rosa, S; de Giulio, A; Crispino, A; Iodice, C; Tommonaro, G. Further bioactive sesterterpenes from the Tyrrhenian sponge Fasciospongia cavernosa. Nat Prod Res 1997, 10, 267–274. [Google Scholar]
- Barrero, AF; Alvarez-Manzaneda, EJ; Chahboun, R; Cuerva, JM; Segovia, A. Synthetic applications of the thermal rearrangement of ozonides: first enantiospecific synthesis of marine metabolite Luffarin W. Synlett 2000, 9, 1269–1272. [Google Scholar]
- Boukouvalas, J; Robichaud, J; Maltais, F. A unified strategy for the regiospecific assembly of homoallayl-substituted butenolides and γ-hydroxybutenolides: first synthesis of luffariellolide. Synlett 2006, 16, 2480–2482. [Google Scholar]
- Gross, H; König, GM. Terpenoids from marine organisms: unique structures and their pharmacological potential. Phytochem Rev 2006, 5, 115–141. [Google Scholar]
- Kernan, MR; Barrabee, EB; Faulkner, DJ. Variation of the metabolites of Chromodoris funerea: comparison of specimen from a Palauan marine lake with those from adjacent waters. Comp Biochem Physiol 1988, 89B, 275–278. [Google Scholar]
- Sullivan, B; Faulkner, DJ. An antimicrobial sesterterpene from a Palauan sponge. Tetrahedron Lett 1982, 23, 907–910. [Google Scholar]
- Schmidt, EW; Faulkner, DJ. Palauolol, a new anti-inflammatory sesterterpene from the sponge Fascaplysinopsis sp. from Palau. Tetrahedron Lett 1996, 37, 3951–3954. [Google Scholar]
- De Rosa, S; Puliti, R; Crispino, A; de Giulio, A; de Sena, C; Iodice, C; Mattia, CA. 25-Deoxycacospongionolide B and cacospongionolide C, two new terpenoids from the sponge Fasciospongia cavernosa. Tetrahedron 1995, 51, 10731–10736. [Google Scholar]
- Lal, AR; Cambie, RC; Rickard, CEF; Bergquist, PR. Sesterterpene lactones from a sponge species of the genus Dactylospongia. Tetrahedron Lett 1994, 35, 2603–2606. [Google Scholar]
- Cambie, CR; Lal, AR; Rickard, CEF. A sesterterpene lactone from Petrosaspongia nigra sp. nov. Acta Cryst 1996, C52, 709–711. [Google Scholar]
- Gomez-Paloma, L; Randazzo, A; Minale, L; Debitus, C; Roussakis, C. New cytotoxic sesterterpenes from the New Caledonian marine sponge Petrosaspongia nigra (Bergquist). Tetrahedron 1997, 53, 10451–10458. [Google Scholar]
- Randazzo, A; Debitus, C; Minale, L; Pastor, PG; Alcaraz, MJ; Payá, M; Gomez-Paloma, L. Petrosaspongiolodes M–R: new potent and selective phosphoplipase A2 inhibitors from the New Caledonian marine sponge Petrosaspongia nigra. J Nat Prod 1998, 61, 571–575. [Google Scholar]
- De Marino, S; Iorizzi, M; Zollo, F; Debitus, C; Menou, J.-L; Ospina, LF; Alcaraz, MJ; Payá, M. New pyridinium alkaloids from a marine sponge of the genus Spongia with a human phospholipase A2 inhibitor profile. J Nat Prod 2000, 63, 322–326. [Google Scholar]
- Balsinde, J; Balboa, MA; Insel, PA; Dennis, EA. Regulation and inhibition of phospholipase A2. Ann Rev Pharmacol Toxicol 1999, 39, 175–189. [Google Scholar]
- Dal Piaz, FD; Casapullo, A; Randazzo, A; Riccio, R; Pucci, P; Marino, G; Gomez-Paloma, L. Molecular basis of phospholipase A2 inhibition by petrosaspongiolide M. ChemBioChem 2002, 3, 664–671. [Google Scholar]
- Monti, MC; Casapullo, A; Riccio, R; Gomez-Paloma, L. Further insights on the structural aspects of PLA2 inhibition by γ-hydroxybutenolide-containing natural products: a comparative study of petrosaspongiolides M–R. Bioorg Med Chem 2004, 12, 1467–1474. [Google Scholar]
- Monti, MC; Casapullo, A; Riccio, R; Gomez-Paloma, L. PLA2-mediated catalytic activation of its inhibitor 25-acetyl-petrosaspongiolide M: serendipitous identification of a new PLA2 suicide inhibitor. FEBS Lett 2004, 578, 269–274. [Google Scholar]
- Gomez-Paloma, L; Monti, MC; Terracciano, S; Casapullo, A; Riccio, R. Chemistry and biology of anti-inflammatory marine natural products. Phospholipase A2 inhibitors. Curr Org Chem 2005, 9, 1419–1427. [Google Scholar]
- Monti, MC; Riccio, R; Casapullo, A. Effects of petrosaspongiolide R on the surface topology of bee venom PLA2: a limited proteolysis and mass spectrometry analysis. Bioorg Chem 2009, 37, 6–10. [Google Scholar]
- Monti, MC; Casapullo, A; Cavasotto, CN; Tosco, A; Dal Piaz, F; Ziemys, A; Margarucci, L; Riccio, R. The binding mode of petrosaspongiolide M to the human group IIA phospholipase A2: exploring the role of covalent and noncovalent interactions in the inhibition process. Chem Eur J 2009, 15, 1155–1163. [Google Scholar]
- Ferreiro-Mederos, L; Lanners, S; Henchiri, H; Fekih, A; Hanquet, G. Hemisynthesis of two marine cheilantane sesterterpenes from (−)-sclareol: first enantioselective synthesis of petrosaspongiolide R. Nat Prod Res 2009, 23, 256–263. [Google Scholar]
- Chawla, A; Dev, S. A new class of triterpenoids from Ailanthus malabarica DC. Derivatives of malabaricane. Tetrahedron Lett 1967, 8, 4837–4843. [Google Scholar]
- Sobti, RR; Dev, S. A direct correlation of (+)-malabaricol with (+)-ambreinolide. Tetrahedron Lett 1968, 9, 2215–2217. [Google Scholar]
- Paton, WF; Paul, IC; Bajaj, AG; Dev, S. The structure of malabaricol. Tetrahedron Lett 1979, 20, 4153–4154. [Google Scholar]
- Ravi, BN; Wells, RJ; Croft, KD. Malabaricane triterpenes from a Fijian collection of the sponge Jaspis stellifera. J Org Chem 1981, 46, 1998–2001. [Google Scholar]
- McCabe, T; Clardy, J; Minale, L; Pizza, C; Zollo, F; Riccio, R. A triterpenoid pigment with the isomalabaricane skeleton from the marine sponge Stelletta sp. Tetrahedron Lett 1982, 23, 3307–3310. [Google Scholar]
- Rao, Z; Deng, S; Wu, H; Jiang, S. Rhabdastrellic acid–A, a novel triterpenoid from the marine sponge Rhabdastrella globostellata. J Nat Prod 1997, 60, 1163–1164. [Google Scholar]
- Guo, J.-F; Zhou, J.-M; Zhang, Y; Deng, R; Liu, J.-N; Feng, G.-K; Liu, Z.-C; Xiao, D.-J; Deng, S.-Z; Zhu, X.-F. Rhabdastrellic acid–A inhibited PI3K/Akt pathway and induced apoptosis in human leukemia HL–60 cells. Cell Biol Int 2008, 32, 48–54. [Google Scholar]
- Su, JY; Meng, YH; Zeng, LM; Fu, X; Schmitz, FJ. Stellettin A, a new triterpenoid pigment from the marine sponge Stelletta tenuis. J Nat Prod 1994, 57, 1450–1451. [Google Scholar]
- McCormick, JL; McKee, TC; Cardellina, JH, II; Leid, M; Boyd, MR. Cytotoxic triterpenes from a marine sponge, Stelletta sp. J Nat Prod 1996, 59, 1047–1050. [Google Scholar]
- McKee, TC; Bokesch, HR; McCormick, JL; Rashid, MA; Spielvogel, D; Gustafson, KR; Alavanja, MM; Cardellina, JH, II; Boyd, MR. Isolation and characterization of new anti-HIV and cytotoxic leads from plants, marine, and microbial organisms. J Nat Prod 1997, 60, 431–438. [Google Scholar]
- Tasdemir, D; Mangalindan, GC; Concepción, GP; Verbitski, SM; Rabindran, S; Miranda, M; Greenstein, M; Hooper, JNA; Harper, MK; Ireland, CM. Bioactive isomalabaricane triterpenes from the marine sponge Rhabdastrella globostellata. J Nat Prod 2002, 65, 210–214. [Google Scholar]
- Tang, SA; Deng, ZW; Li, J; Fu, HZ; Pei, YH; Zhang, S; Lin, WH. A new isomalabaricane triterpenoid from sponge Jaspis sp. Chin Chem Lett 2005, 16, 353–355. [Google Scholar]
- Clement, JA; Li, M; Hecht, SM; Kingston, DGI. Bioactive isomalabaricane triterpenoids from Rhabdastrella globostellata that stabilize the binding of DNA polymerase β to DNA. J Nat Prod 2006, 69, 373–376. [Google Scholar]
- Liu, WK; Cheung, FWK; Che, C.-T. Stellettin A induces oxidative stress and apoptosis in HL–60 human leukemia and LNCaP prostate cancer cell lines. J Nat Prod 2006, 69, 934–937. [Google Scholar]
- Krysko, DV; Roels, F; Leybaert, L; D’Herde, K. Mitochrondrial transmembrane potential changes support the concept of mitochondrial heterogeneity during apoptosis. J Histochem Cytochem 2001, 49, 1277–1284. [Google Scholar]
- Lin, H.-W; Wang, Z.-L; Wu, J.-H; Shi, N; Zhang, H.-J; Chen, W.-S; Morris-Natschke, SL; Lin, A.-S. Stellettins L and M, cytotoxic isomalabaricane-type triterpenes, and sterols from the marine sponge Stelletta tenuis. J Nat Prod 2007, 70, 1114–1117. [Google Scholar]
- Lv, F; Xu, M; Deng, Z; de Voogd, NJ; van Soest, RWM; Proksch, P; Lin, WH. Rhabdastrellins A – F, isomalabaricane triterpenes from the marine sponge Rhabdastrella aff. distinca. J Nat Prod 2008, 71, 1738–1741. [Google Scholar]
- Tsuda, M; Ishibashi, M; Agemi, K; Sasaki, T; Kobayashi, J. Stelliferins A–F, new antineoplastic isomalabaricane triterpenes from the Okinawan marine sponge Jaspis stellifera. Tetrahedron 1991, 47, 2181–2194. [Google Scholar]
- Oku, N; Matsunaga, S; Wada, SI; Watabe, S; Fusetani, N. New isomalabaricane triterpenes from the marine sponge Stelletta globostellata that induce morphological changes in rat fibroblasts. J Nat Prod 2000, 63, 205–209. [Google Scholar]
- Meragelman, KM; McKee, TC; Boyd, MR. New cytotoxic isomalabaricane triterpenes from the sponge Jaspis species. J Nat Prod 2001, 64, 389–392. [Google Scholar]
- Tabudravu, JN; Jaspars, M. Stelliferin riboside, a triterpene monosaccharide isolated from the Fijian sponge Geodia globostellifera. J Nat Prod 2001, 64, 813–815. [Google Scholar]
- Raeppel, F; Weibel, J.-M; Heissler, D. Synthesis of the trans-syn-trans perhydrobenz[e]indene moiety of the stellettins and the stelliferins. Tetrahedron Lett 1999, 40, 6377–6381. [Google Scholar]
- Ryu, G; Matsunaga, S; Fusetani, N. Globostellatic acids A–D, new cytotoxic isomalabaricane triterpenes from the marine sponge Stelletta globostelleta. J Nat Prod 1996, 59, 512–514. [Google Scholar]
- Fouad, M; Edrada, RA; Ebel, R; Wray, V; Müller, WEG; Lin, WH; Proksch, P. Cytotoxic isomalabaricane triterpenes from the marine sponge Rhabdastrella globostellata. J Nat Prod 2006, 69, 211–218. [Google Scholar]
- Aoki, S; Sanagawa, M; Watanabe, Y; Setiawan, A; Arai, M; Kobayashi, M. Novel isomalabaricane triterpenes, exhibiting selective anti-proliferative activity against vascular endothelial cells, from marine sponge Rhabdastrella globostellata. Bioorg Med Chem 2007, 15, 4818–4828. [Google Scholar]
- Zhang, WH; Che, C.-T. Isomalabaricane-type nortriterpenoids and other constituents of the marine sponge Geodia japonica. J Nat Prod 2001, 64, 1489–1492. [Google Scholar]
- Lv, F; Deng, Z; Li, J; Fu, H; van Soest, RWM; Proksch, P; Lin, WH. Isomalabaricane-type compounds from the marine sponge Rhabdastrella aff. distinca. J Nat Prod 2004, 67, 2033–2036. [Google Scholar]
- Liu, WK; Ho, JCK; Che, CT. Apoptotic activity of isomalabaricane triterpenes on human promyelocytic leukemia HL-60 cells. Cancer Lett 2005, 230, 102–110. [Google Scholar]
- Kobayashi, J; Yuasa, K; Kobayashi, T; Sasaki, T; Tsuda, M. Jaspiferals A–G, new cytotoxic isomalabaricane-type nortriterpenoids from Okinawan marine sponge Jaspis stellifera. Tetrahedron 1996, 52, 5745–5750. [Google Scholar]
- Zampella, A; D’Auria, MV; Debitus, C; Menou, J.-L. New isomalabaricane derivatives from a new species of Jaspis sponge collected at the Vanuatu Islands. J Nat Prod 2000, 63, 943–946. [Google Scholar]
- Bourguet-Kondracki, M-LA; Debitus, C; Guyot, M. New cytotoxic isomalabaricane-type sesterterpenes from the new Caledonian marine sponge Rhabdastrella globostellata. Tetrahedron Lett 2000, 41, 3087–3090. [Google Scholar]
- Tang, S; Pei, Y; Fu, H; Deng, Z; Li, J; Proksch, P; Lin, WH. Jaspolides A–F, six new isomalabaricane-type terpenoids from the sponge Jaspis sp. Chem Pharm Bull 2006, 54, 4–8. [Google Scholar]
- Tang, S; Deng, Z; Proksch, P; Lin, WH. Jaspolides G and H, unique bisisomalabaricanes from the Chinese marine sponge Jaspis sp. Tetrahedron Lett 2007, 48, 5443–5447. [Google Scholar]
- Li, M; Wei, S.-Y; Tang, S.-A; Lin, W.-H; Cui, J.-R. Antileukemic activity of jaspolide B, an isomalabaricance-type triterpene from marine sponge Jaspis sp. on human promyeloleukemic HL-60 cells. J Ch Pharm Sci 2008, 17, 11–15. [Google Scholar]
- Wei, S.-Y; Li, M; Tang, SA; Sun, W; Xu, B; Cui, J.-R; Lin, W.-H. Induction of apoptosis accompanying with G1 phase arrest and microtubule disassembly in human hepatoma cells by jaspolide B, a new isomalabaricane-type triterpene. Cancer Lett 2008, 262, 114–122. [Google Scholar]
- Cheng, JF; Kobayashi, J; Nakamura, H; Ohizumi, Y; Hirata, Y; Sasaki, T. Penasterol, a novel antileukemic sterol from the Okinawan marine sponge Penares sp. J Chem Soc Perkin Trans I 1988, 8, 2403–2406. [Google Scholar]
- Shoji, N; Umeyama, A; Motoki, S; Arihara, S; Ishida, T; Nomoto, K; Kobayashi, J; Takei, M. Potent inhibitors of histamine release, two novel triterpenoids from the Okinawan marine sponge Penares incrustans. J Nat Prod 1992, 55, 1682–1685. [Google Scholar]
- Carmely, S; Roll, M; Loya, Y; Kashman, Y. The structure of eryloside A, a new antitumor and antifungal 4-methylated steroidal glycoside from the sponge Erylus lendenfeldi. J Nat Prod 1989, 52, 167–170. [Google Scholar]
- D’Auria, MV; Paloma, LG; Minale, L; Riccio, R; Debitus, C. Structure characterization by two-dimensional NMR spectroscopy, of two marine triterpene oligosaccharides from a Pacific sponge of the genus. Erylus Tetrahedron 1992, 48, 491–498. [Google Scholar]
- Gulavita, NK; Wright, AE; Kelly-Borges, M; Longley, RE; Yarwood, D; Sills, MA. Eryloside E from an Atlantic sponge Erylus goffrilleri. Tetrahedron Lett 1994, 35, 4299–4302. [Google Scholar]
- Stead, P; Hiscox, S; Robinson, PS; Pike, NB; Sidebottom, PJ; Roberts, AD; Taylor, NL; Wright, AE; Pomponi, SA; Langely, D. Eryoside F, a novel penasterol disaccharide possessing potent thrombin receptor antagonist activity. Bioorg Med Chem Lett 2000, 10, 661–664. [Google Scholar]
- Shin, J; Lee, H.-S; Woo, L; Rho, J.-R; Seo, Y; Cho, KW; Sim, CJ. New triterpenoid saponins from the sponge Erylus nobilis. J Nat Prod 2001, 64, 767–771. [Google Scholar]
- Fouad, M; Al-Trabeen, K; Badran, M; Wray, V; Edrada, R; Proksch, P; Ebel, R. New steroidal saponins from the sponge Erylus lendenfeldi. ARKIVOC 2004, xiii, 17–27. [Google Scholar]
- Antonov, AS; Kalinovsky, AI; Stonik, VA; Afiyatullov, SS; Aminin, DL; Dmitrenok, PS; Mollo, E; Cimino, G. Isolation and structures of erylosides from the Caribbean sponge Erylus formosus. J Nat Prod 2007, 70, 169–178. [Google Scholar]
- Afiyatullov, SS; Kalinovsky, AI; Antonov, AS; Ponomarenko, LP; Dmitrenok, PS; Aminin, DL; Krasokhin, VB; Nosova, VM; Kisin, AV. Isolation and structures of erylosides from the Caribbean sponge Erylus goffrilleri. J Nat Prod 2007, 70, 1871–1877. [Google Scholar]
- Jaspars, M; Crews, P. A triterpene tetrasaccharide, formoside, from the Caribbean Choristida sponge Erylus formosus. Tetrahedron Lett 1994, 35, 7501–7504. [Google Scholar]
- Kubanek, J; Pawlik, JR; Eve, TM; Fenical, W. Triterpene glycosides defend the Caribbean reef sponge Erylus formosus from predatory fishes. Mar Ecol Prog Ser 2000, 207, 69–77. [Google Scholar]
- Takada, K; Nakao, Y; Matsunaga, S; van Soest, RWM; Fusetani, N. Nobiloside, a new neuraminidase inhibitory triterpenoidal saponin from the marine sponge Erylus nobilis. J Nat Prod 2002, 65, 411–413. [Google Scholar]
- Okada, Y; Matsunaga, S; van Soest, RWM; Fusetani, N. Sokodosides, steroid glycosides with an isopropyl side chain, from the marine sponge Erylus placenta. J Org Chem 2006, 71, 4884–4888. [Google Scholar]
- Dasgupta, S; Pramanik, K; Mukhopadhyay, B. Oligosaccarides through reactivity tuning: convergent synthesis of the trisaccharides of the steroid glycoside sokodoside B isolated from marine sponge Erylus placenta. Tetrahedron 2007, 63, 12310–12316. [Google Scholar]
- Kitagawa, I; Kobayashi, M; Okamoto, Y; Yoshikawa, M; Hamamoto, Y. Structures of sarasinosides A1, B1, and C1; new norlanostane-triterpenoid oligosaccharides from the Palauan marine sponge Asteropus sarasinosum. Chem Pharm Bull 1987, 35, 5036–5039. [Google Scholar]
- Schmitz, FJ; Ksebati, MB; Gunasekera, SP; Agarwal, S. Sarasinoside A1: A saponin containing amino sugars isolated from a sponge. J Org Chem 1988, 53, 5941–5947. [Google Scholar]
- Kobayashi, M; Okamoto, Y; Kitagawa, I. Marine natural products. XXVIII. The structures of sarasinosides A1, A2, A3, B1, B2, B3, C1, C2, and C3, nine new norlanostane-triterpenoidal oligosaccharides from the Palauan marine sponge Asteropus sarasinosum. Chem Pharm Bull 1991, 39, 2867–2877. [Google Scholar]
- Espada, A; Jiménez, C; Rodríguez, J; Crews, P; Riguera, R. Sarasinosides D–G: four new triterpenoid saponins from the sponge Asteropus sarasinosum. Tetrahedron 1992, 48, 8685–8696. [Google Scholar]
- Lee, H.-S; Seo, Y; Cho, KW; Rho, J.-R; Shin, J; Paul, VJ. New triterpenoid saponins from the sponge Melophlus isis. J Nat Prod 2000, 63, 915–919. [Google Scholar]
- Dai, H.-F; Edrada, RA; Ebel, R; Nimtz, M; Wray, V; Proksch, P. Norlanostane triterpenoidal saponins from the marine sponge Melophlus sarassinorum. J Nat Prod 2005, 68, 1231–1237. [Google Scholar]
- Kalinovsky, AI; Antonov, AS; Afiyatullov, SS; Dmitrenok, PS; Evtuschenko, EV; Stonik, VA. Mycaloside A, a new steroid oligoglycoside with an unprecedented structure from the Caribbean sponge Mycale laxissima. Tetrahedron Lett 2002, 43, 523–525. [Google Scholar]
- Antonov, AS; Afiyatullov, SS; Kalinovsky, AI; Ponomarenko, LP; Dmitrenok, PS; Aminin, DL; Agafonosa, IG; Stonik, VA. Mycalosides B–I, eight new spermostatic steroid oligoglycosides from the sponge Mycale laxissima. J Nat Prod 2003, 66, 1082–1088. [Google Scholar]
- Afiyatullov, SS; Antonov, AS; Kalinovsky, AI; Dmitrenok, PS. Two new steroid oligoglycosides from the Caribbean sponge Mycale laxissima. Nat Prod Commun 2008, 3, 1581–1586. [Google Scholar]
- Cafieri, F; Fattorusso, E; Taglialatela-Scafati, O. Ectyoplasides A–B – unique triterpene oligoglycosides from the Caribbean sponge Ectyoplasia ferox. Eur J Org Chem 1999, 231–238. [Google Scholar]
- Campagnuolo, C; Fattorusso, E; Taglialatela-Scafati, O. Feroxosides A–B, two norlanostane tetraglycosides from the Caribbean sponge Ectyoplasia ferox. Tetrahedron 2001, 57, 4049–4055. [Google Scholar]
- Antonov, AS; Kalinovsky, AI; Stonik, VA. Ulososide B, a new unusual norlanostanetriterpene glycoside and its genuine aglycone from the Madagascar sponge Ulosa sp. Tetrahedron Lett 1998, 39, 3807–3808. [Google Scholar]
- Antonov, AS; Kalinovskii, AI; Dmitrenok, PS; Stonik, VA. New triterpene glycosides from Ulosa sp. sponge. Russ J Bioorg Chem 2002, 28, 183–188. [Google Scholar]
- Ryu, G; Choi, BW; Lee, BH; Hwang, K.-H; Lee, UC; Jeong, DS; Lee, NH. Wondosterols A–C, three steroidal glycosides from a Korean marine two-sponge association. Tetrahedron 1999, 55, 13171–13178. [Google Scholar]
- Hirota, H; Takayama, S; Miyashiro, S; Ozaki, Y; Ikegami, S. Structure of a novel steroidal saponin, pachastrelloside A, obtained from a marine sponge of the genus Pachastrella. Tetrahedron Lett 1990, 31, 3321–3324. [Google Scholar]
- Proksch, P; Edrada, RA; Ebel, R. Drugs from the sea: current status and microbiological applications. Appl Micobiol Biotechnol 2002, 59, 125–134. [Google Scholar]
- Olivera, BM. ω-Conotoxin MVIIA: from marine snail venom to analgesic drug. In Drugs from the Sea; Fusetani, N, Ed.; Basel: Karger, Switzerland, 2000; pp. 74–85. [Google Scholar]
- Mendola, D. Aquacultural production of bryostatin 1 and ecteinascidin 743. In Drugs from the Sea; Fusetani, N, Ed.; Karger: Basel, Switzerland, 2000; pp. 74–85. [Google Scholar]
- Fenical, W. Chemical studies of marine bacteria: developing a new resource. Chem Rev 1993, 93, 1673–1683. [Google Scholar]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ebada, S.S.; Lin, W.; Proksch, P. Bioactive Sesterterpenes and Triterpenes from Marine Sponges: Occurrence and Pharmacological Significance. Mar. Drugs 2010, 8, 313-346. https://doi.org/10.3390/md8020313
Ebada SS, Lin W, Proksch P. Bioactive Sesterterpenes and Triterpenes from Marine Sponges: Occurrence and Pharmacological Significance. Marine Drugs. 2010; 8(2):313-346. https://doi.org/10.3390/md8020313
Chicago/Turabian StyleEbada, Sherif S., WenHan Lin, and Peter Proksch. 2010. "Bioactive Sesterterpenes and Triterpenes from Marine Sponges: Occurrence and Pharmacological Significance" Marine Drugs 8, no. 2: 313-346. https://doi.org/10.3390/md8020313
APA StyleEbada, S. S., Lin, W., & Proksch, P. (2010). Bioactive Sesterterpenes and Triterpenes from Marine Sponges: Occurrence and Pharmacological Significance. Marine Drugs, 8(2), 313-346. https://doi.org/10.3390/md8020313