Expression, Purification and Bioactivities Analysis of Recombinant Active Peptide from Shark Liver

Abstract
:1. Introduction
2. Results
2.1. cDNA Cloning
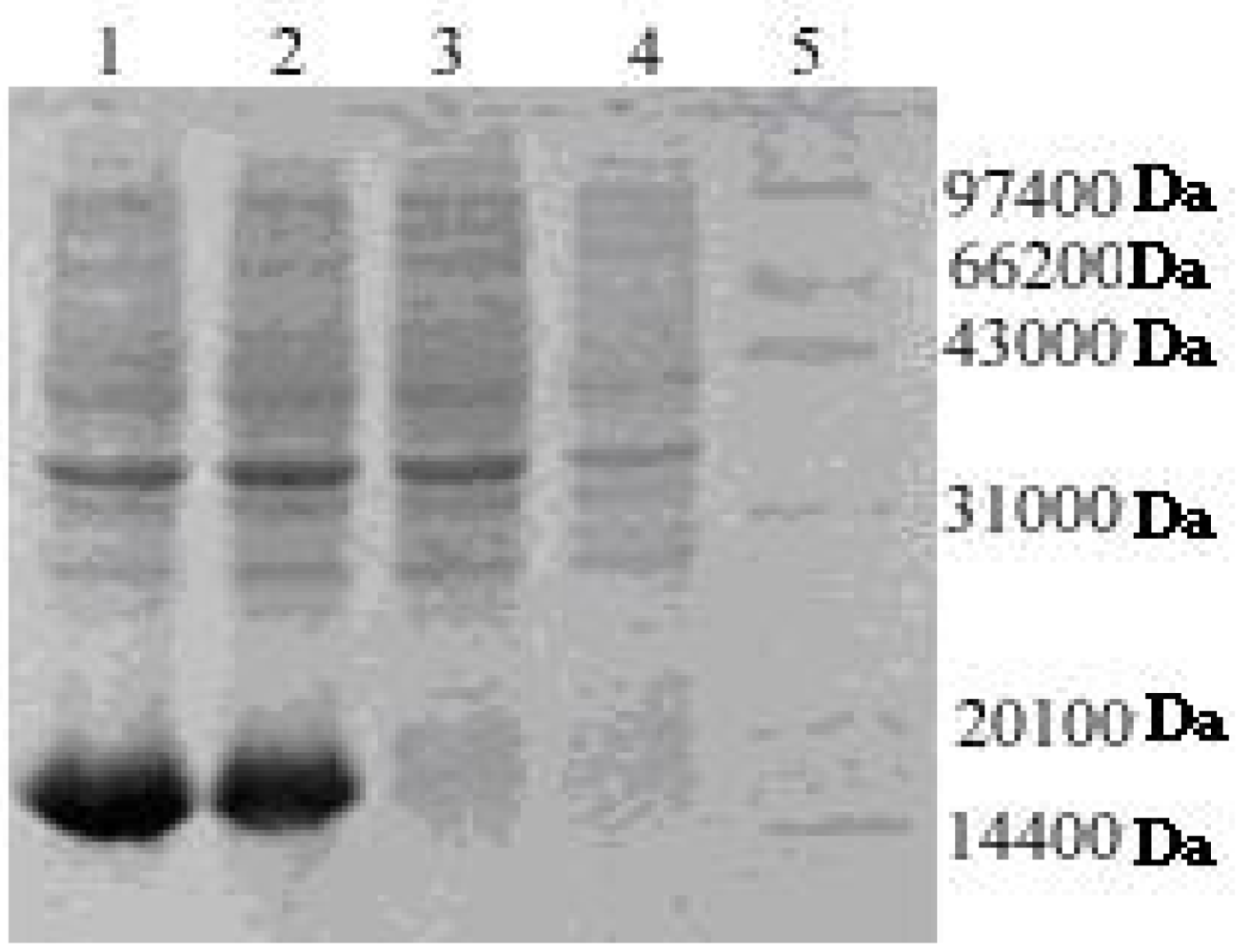
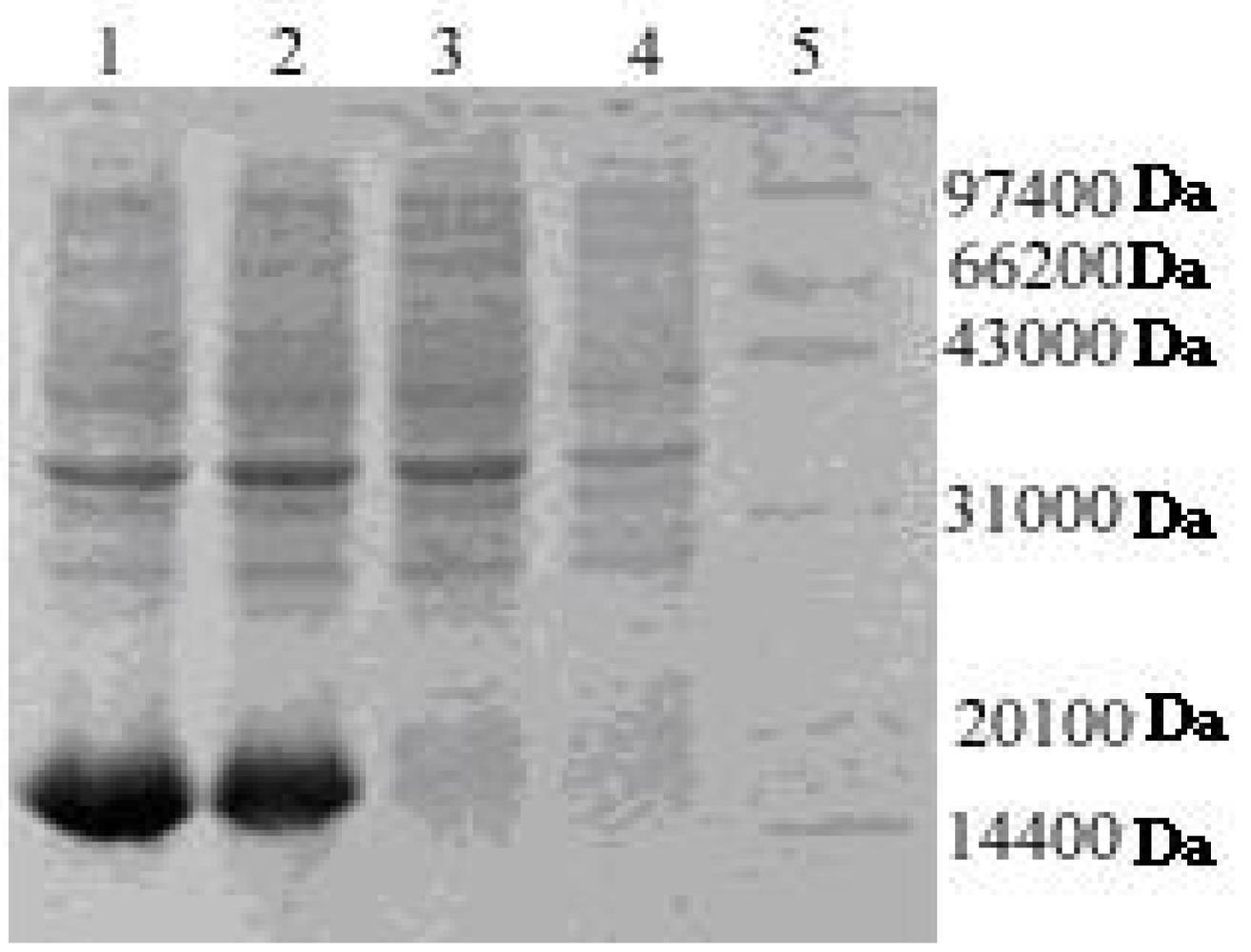
2.2. Expression and Purification of His-Tagged rAPSL
2.3. The Study of APSL Bioactivity
3. Discussion
4. Experimental
4.1. Materials
4.2 Cloning of the cDNA Fragment of APSL by RT-PCR
4.3. Overexpression and Purification of Recombinant APSL (rAPSL)
4.4. Improved Effects of rAPSL to the Proliferation of SMMC7721 Cells
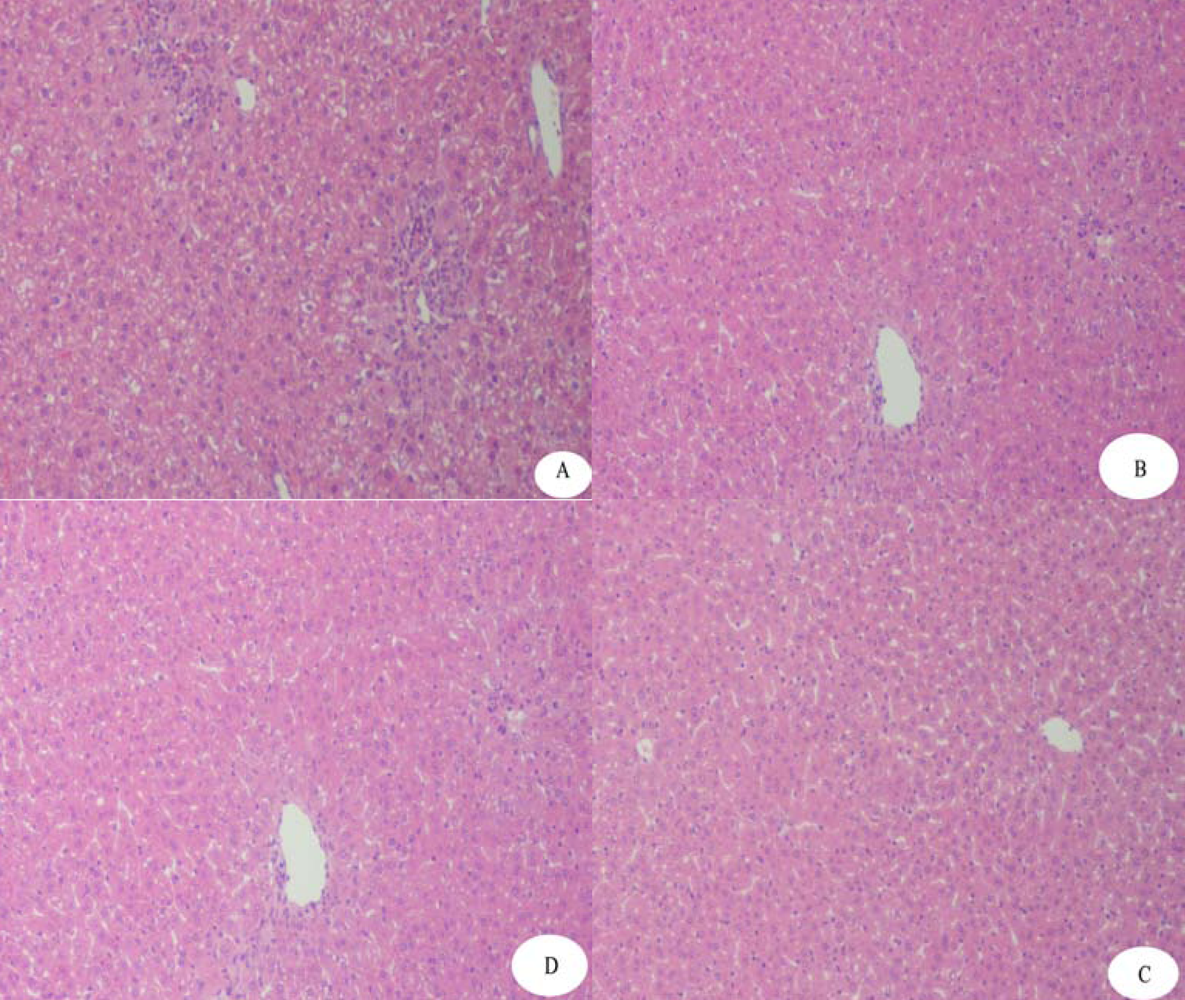
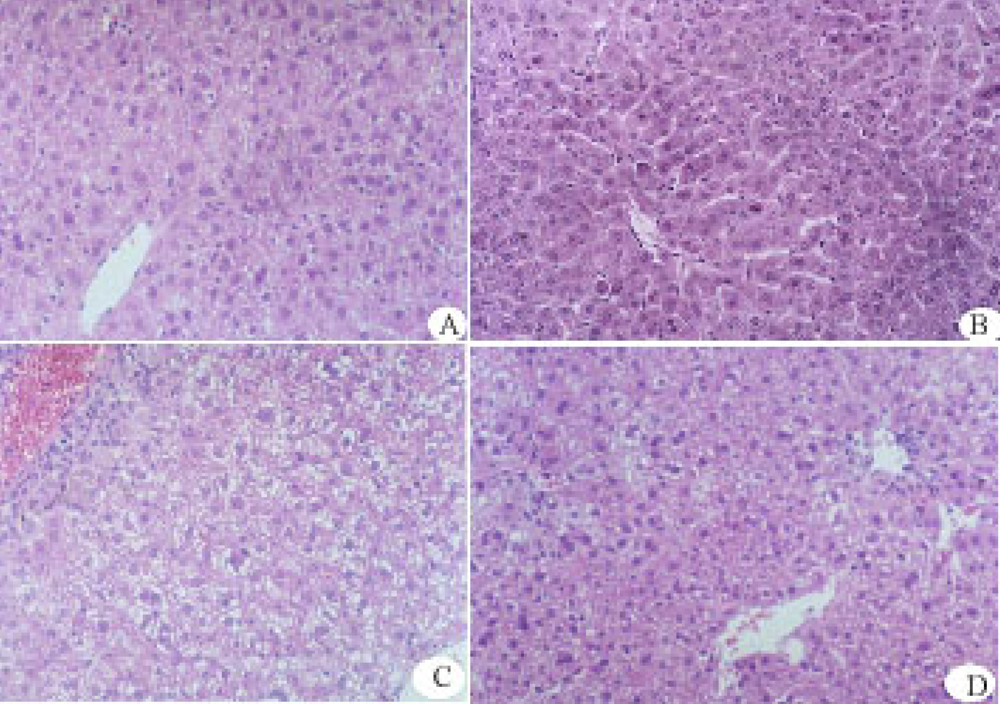
4.5. Protective Effects of rAPSL against Mice of Acute Hepatic Injury Induced by CCl4 or AAP
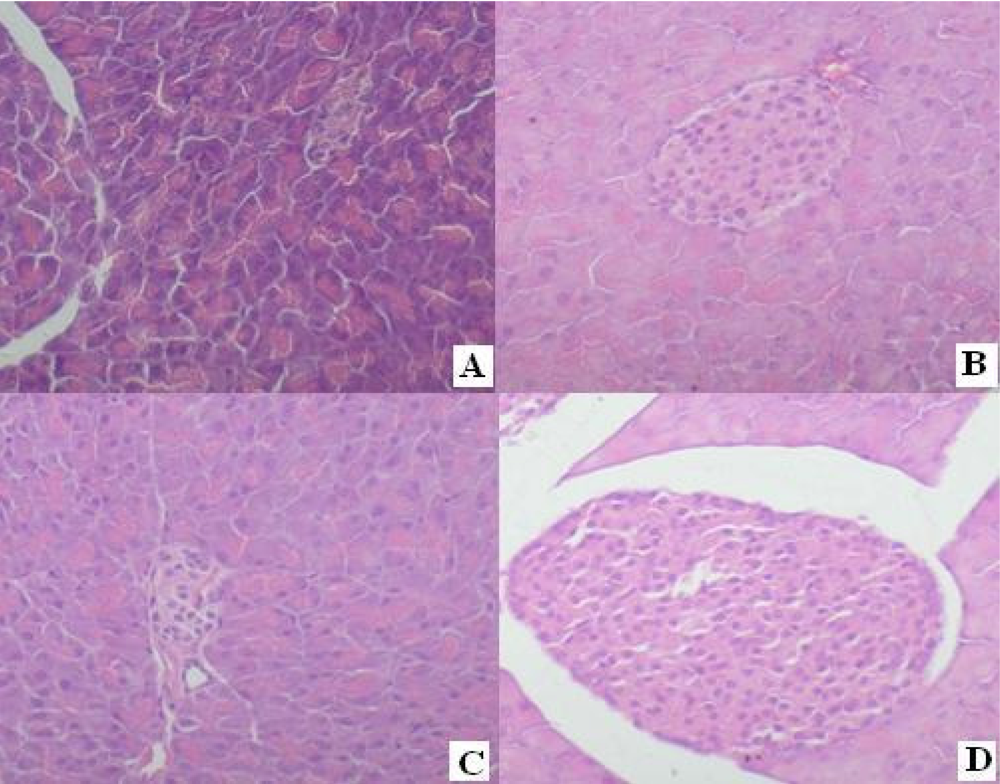
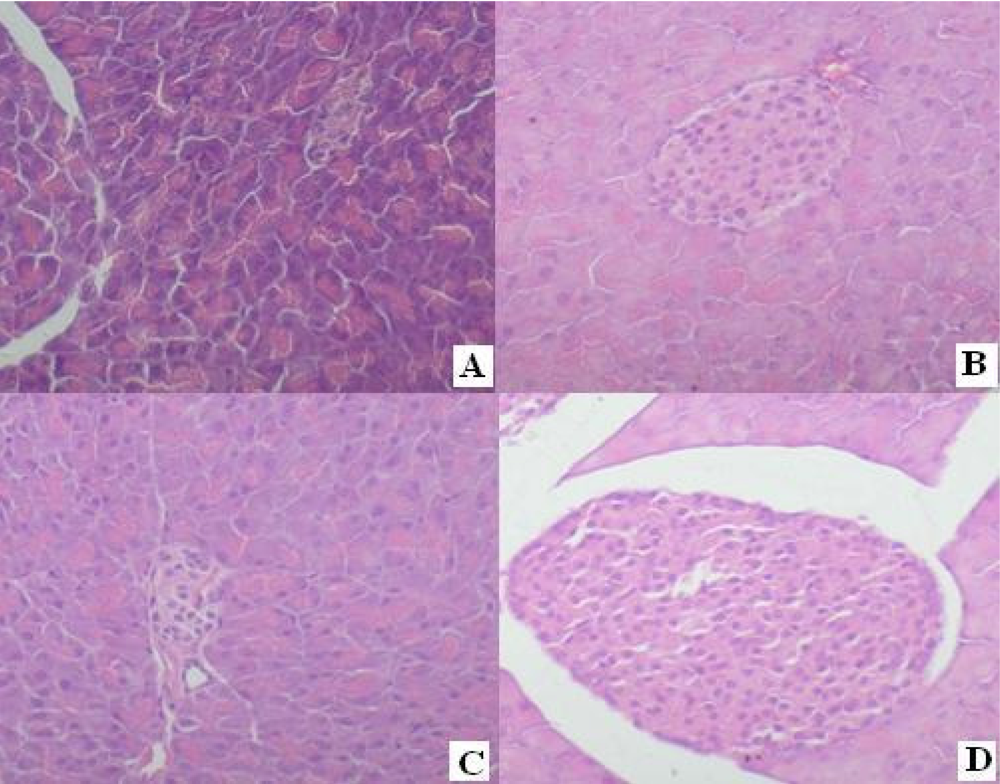
4.6. Protective Effects to the Diabetes Mellitus Mice Induced by Alloxan
4.7. Statistical Analysis of the Data
Acknowledgements
- Sample Availability: Available from the authors.
References
- Xie, ZF; Cheng, GM; Huang, Y; Xing, CX. Study on immune activity of shark liver extracts. Chin J Biochem Pharm 1999, 20, 126–129. [Google Scholar]
- Ou, Y; Liao, GY; Wu, WT. Cloning and expression of a new gene from Shark Liver and its inhibitory effects on Hepatoma cells. Chin J Nat Med 2009, 7, 65–70. [Google Scholar] [CrossRef]
- Hagiya, M; Francavilla, A; Polomege, L. Cloning and sequence analysis of the augmenter of liver regeneration (ALR) gene: expression of biologically active recombinant ALR and demonstration of tissue distribution. Proc Natl Acad Sci U S A 1994, 91, 8142–8146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, ZH; Yang, XM; Xie, L. Cloning and expression of rat augmenter of liver regeneration. Chin J Biochem Pharm 1997, 13, 130–135. [Google Scholar]
- Qiu, ZH; He, FC; Yang, XM. The alteration of hepatic stimulatory substance and its mRNA in the process of regenerating liver in rat. Chin Appl Physiol 1996, 12, 236–238. [Google Scholar]
- Hong, G; Wu, GZ; Liu, GQ. Protective effects of shark hepatic stimulator substance on alloxan-induced diabetes in mice. J Chin Pharm Univ 2003, 34, 155–159. [Google Scholar]
- Ou, Y; Li, Q; Lv, ZB; Wu, WT. Purification and characterization of hepatocyte regenerational stimulatory factor from shark liver. J Chin Pharm Sci 2003, 12, 175–180. [Google Scholar]
- Ye, BP; Xi, T; Lv, ZB. Cloning and sequence analysis of cDNA fragment of shark hepatic stimulator substance’s similarity. Chin J Nat Med 2003, 1, 111–115. [Google Scholar]
- Sambrool, J; Fritsch, EF; Maniatis, T. Molecular Cloning: A Laboratory Manual, third ed; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; Chapter 15; p. 1228. [Google Scholar]
- Liu, A; Guo, Y; Wu, WT. Optimization and improvement of the MTT assay for HSS bio-activity in vitro. Pharm Biotechnol 2000, 9, 344–348. [Google Scholar]
- Xu, SY; Bian, RL; Chen, X. Methodology of Pharmacological Experiments, third ed; People’s Medical Publishing House: Beijing, China, 2002; pp. 1346–1349. [Google Scholar]
- Lv, ZB; Li, Q; Ye, BP; Ruan, QP; Wu, WT. Protective effects of shark hepatic stimulator substance against acute hepatic injury induced by acetaminophen in mice. Acta Pharm Sin 2004, 39, 17–21. (in Chinese). [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Dosage (μg/mL) | OD570 |
|---|---|---|
| Control | - | 0.36 ± 0.03 |
| APSL | 50 | 0.65 ± 0.12** |
| rAPSL | 50 | 0.45 ± 0.02** |
| 25 | 0.41 ± 0.04 | |
| 12.5 | 0.40 ± 0.03 |
| Group | Dosage mg/kg | AST (OD505) | AST (cal’s unit) | ALT (OD505) | ALT (cal’s unit) |
|---|---|---|---|---|---|
| Control | - | 0.141 ± 0.017** | 60.88 ± 10.11** | 0.087 ± 0.022** | 31.17 ± 10.53** |
| Model | - | 0.317 ± 0.046 | 164.49 ± 27.16 | 0.256 ± 0.079 | 111.4 ± 65.05 |
| APSL | 3 | 0.220 ± 0.029** | 107.43 ± 16.88** | 0.153 ± 0.034** | 62.60 ± 21.63** |
| rAPSL | 3 | 0.203 ± 0.061** | 97.79 ± 35.94** | 0.152 ± 0.027** | 62.12 ± 15.80** |
| 1 | 0.266 ± 0.035* | 134.71 ± 20.53* | 0.177 ± 0.045* | 73.96 ± 32.37* |
| Group | Dosage (mg/kg) | ALT(OD505) | ALT (cal’s unit) |
|---|---|---|---|
| Control | - | 0.067 ± 0.016** | 21.52 ± 7.94** |
| Model | - | 0.234 ± 0.081 | 101.17 ± 38.83 |
| APSL | 3 | 0.104 ± 0.022** | 39.26 ± 10.63** |
| rAPSL | 3 | 0.104 ± 0.026** | 38.90 ± 12.39** |
| 1 | 0.141 ± 0.062* | 56.58 ± 29.62* |
| Group | Dosage mg/kg | FPG(mmol/L)
| |||
|---|---|---|---|---|---|
| First week | Second week | Third week | Forth week | ||
| Control | - | 6.37 ± 1.10 | 6.42 ± 1..24 | 6.5 ± 1.24*** | 6.74 ± 1.31*** |
| Model | - | HI | HI | 22.73 ± 3.65 | 20.08 ± 5.18▴▴▴ |
| INS | 6★ | 4.25 ± 0.63 | 4.07 ± 0.73 | 4.23 ± 1.28*** | 4.32 ± 1.10*** |
| rAPSL | 3 | HI | HI | 12.28 ± 4.54*** | 11.44 ± 6.03*** |
| 1 | HI | HI | 20.21 ± 5.92 | 18.04 ± 6.71 | |
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lv, Z.; Ou, Y.; Li, Q.; Zhang, W.; Ye, B.; Wu, W. Expression, Purification and Bioactivities Analysis of Recombinant Active Peptide from Shark Liver. Mar. Drugs 2009, 7, 258-267. https://doi.org/10.3390/md7020258
Lv Z, Ou Y, Li Q, Zhang W, Ye B, Wu W. Expression, Purification and Bioactivities Analysis of Recombinant Active Peptide from Shark Liver. Marine Drugs. 2009; 7(2):258-267. https://doi.org/10.3390/md7020258
Chicago/Turabian StyleLv, Zhengbing, Yu Ou, Qian Li, Wenping Zhang, Boping Ye, and Wutong Wu. 2009. "Expression, Purification and Bioactivities Analysis of Recombinant Active Peptide from Shark Liver" Marine Drugs 7, no. 2: 258-267. https://doi.org/10.3390/md7020258
APA StyleLv, Z., Ou, Y., Li, Q., Zhang, W., Ye, B., & Wu, W. (2009). Expression, Purification and Bioactivities Analysis of Recombinant Active Peptide from Shark Liver. Marine Drugs, 7(2), 258-267. https://doi.org/10.3390/md7020258