Sterol Ring System Oxidation Pattern in Marine Sponges

Abstract
:Introduction

General biosynthetic Reactions in marine sponges
- epoxidation (generally αα and rarely ββ) followed by its opening in different pathways,
- oxidation of the allylic C-7 and C-4 carbons to give simple alcohols of the preferred configuration, and
- isomerisation of the double bond(s).
- oxidation to a carbonyl,
- dehydration producing unsaturation which will create new active allylic positions for further oxidation,
- retro Diels-Alder reaction in the case of vicinal diols, and
- condensation reactions involving OH, CH2OH, CHO, and COOH groups at appropriate locations.
I. Δ5-3β-Hydroxy Steroids
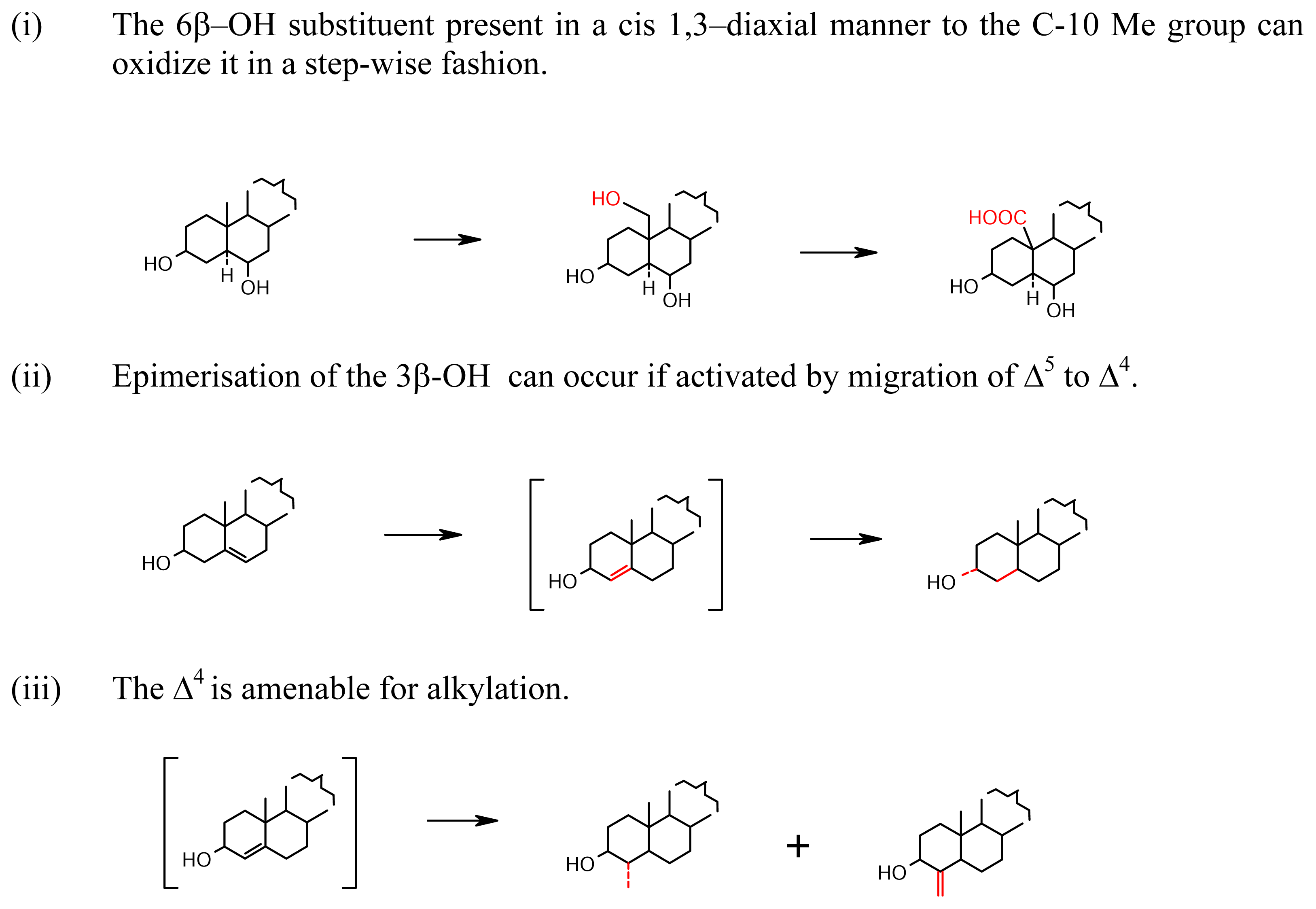
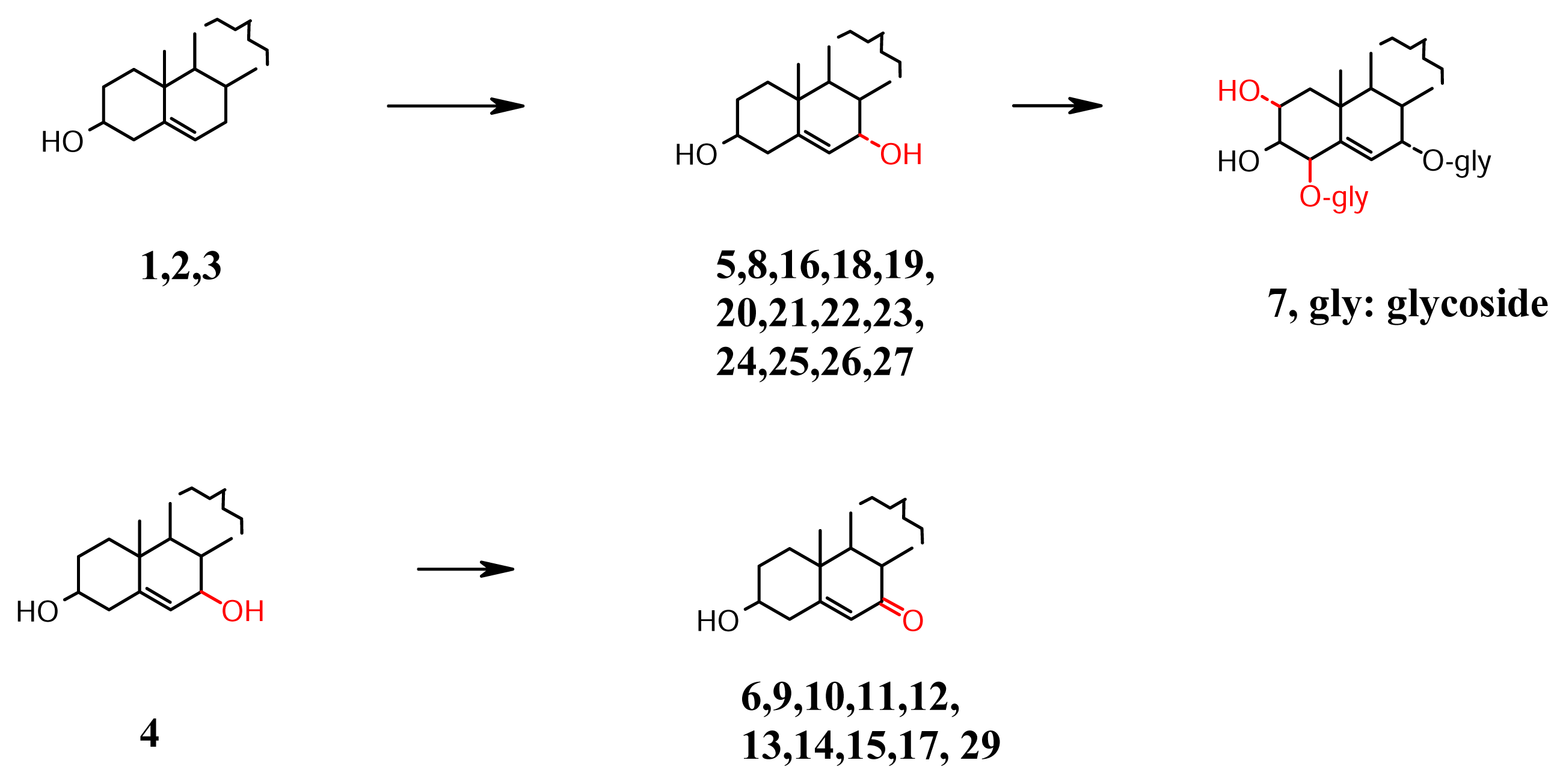
- Oxidation at C-7 (Scheme 3): The epimeric alcohols 4 and 5 and their ketone 6 are from Corallistes undulatus [19] and Cliona copiosa [20] by the allylic C-7 oxidation. It was for some time suspected that the C-7 oxidation might be resulting from auto-oxidation during isolation procedure. Hence, the isolation of the 7α-glycoside 7 (paschastrelloside A) from Paschastrella sp. confirms a biotic origin of the 7α oxygen [21]. The novel feature of 7 is its 2α-OH; the sterol inhibits cell division of fertilized starfish eggs. The presence of the 7α-OH and 7-keto group naturally in the sponge has received further proof from the recent isolation of a number of steroids 8–11 (gelliusterols A-D) from Gellius sp. of the Panaman Caribbean coast [22]. Sterol 12 was isolated from Polymastia sobustia from South China Sea [23], as well as 14 [24], 13 from Geodia japonica also from South China Sea [25]. Sterols 15 and 16 are from a Japanese specimen of Strongylophora corticata [26], and 17 (polysterol A) from Epipolasis sp. [27], and 18–27 topsentinols A to J from an Okinawan species of Topsentia [28]. The sterol containing the nucleus 28 (polysterol B belonging to another sterol subclass, the 3α-hydroxy sterol sulfates: Scheme 13) co-occurs with 29 (polysterol A). Although for the 3α-oxygenated sterols also, the parent is the 3β-Δ5-sterol nucleus, the biogenetic pathway is somewhat different. The isolation of sterols belonging to different biogenetic pathways may be due to symbionts causing species heterogeneity or artifact formation on preservation and the subsequent isolation procedure. Hence, it is necessary to know these factors well for rationalizing the co-occurrence of sterols belonging to different biogenetic classes. The Strongylophora corticata sterols may exemplify this dimension.
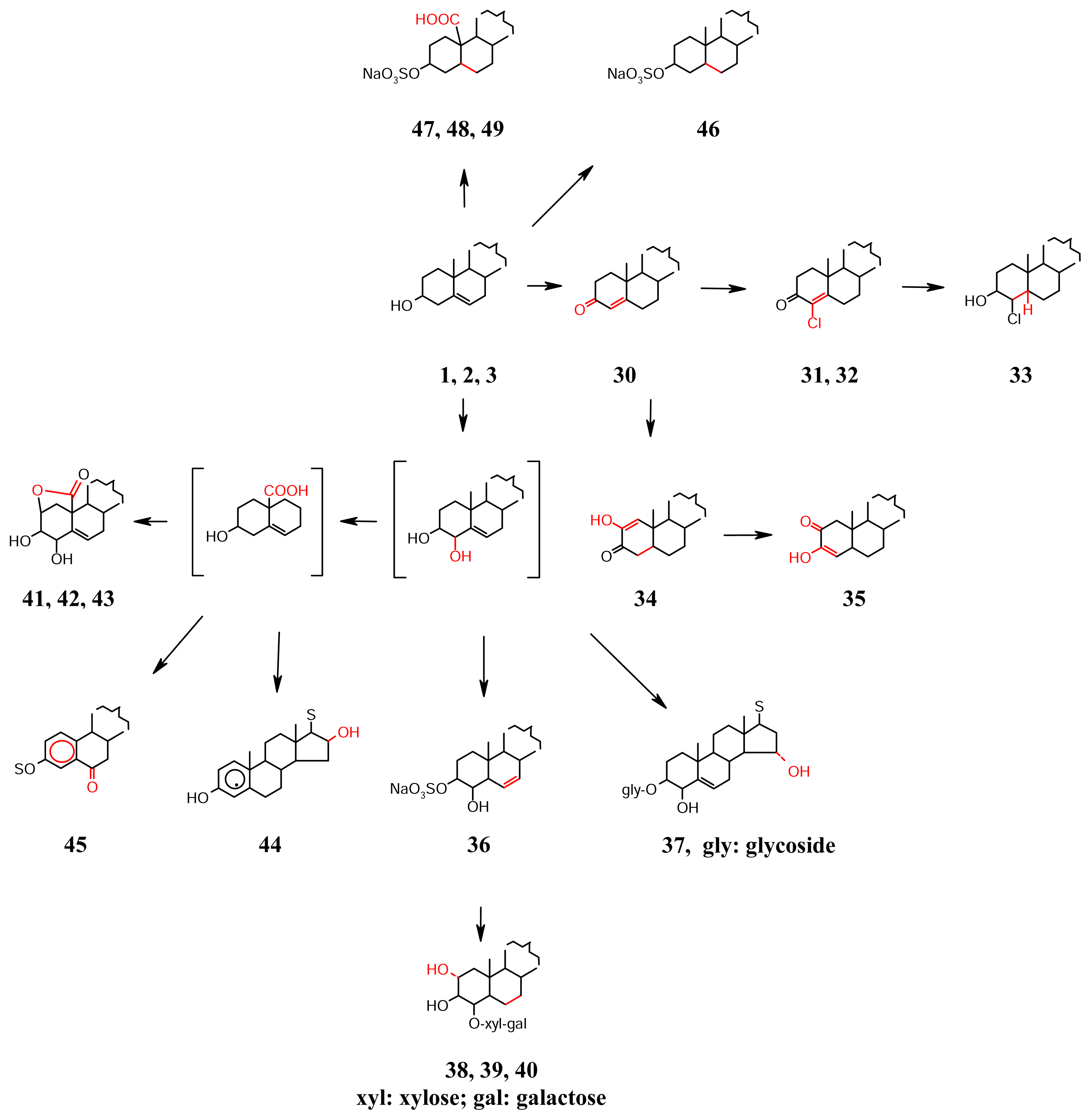
- Oxidation confined to Ring A (Scheme 4): The alcoholic C-3 and allylic C-4 are active sites for oxidation. The formation of 3-ketone can facilitate migration of the Δ5 to the conjugated Δ4 position, as found in 30, mycalone from Mycale sp. of Southern Australia [29]. The chloroketones 31 and 32, kiheisterones C and E present in Strongylacedon sp. from Maui along with the chlorohydrin D 33 are the only halogenated sterols isolated from sponges even though halogenated, particularly brominated natural products are common in marine sponges being derived from red algal symbionts. The products of C-2 activation are the diosphenols 34 (kiheisterone A) and 35 (kiheisterone B) of the same sponge [30]. A hydroxylation of the allylic C-4 is demonstrated by the 3β-sulfoxypregnane 36 isolated from Stylopus australis [31], and the glycoside 37 from Mycale laxissima [32]. The C-1 is activated via the Δ2 formation by dehydration of the 3β-OH. The Δ2, not so far observed in sponge sterols is a routine feature in the highly oxygenated sterol classes of withanolides and physalins from land plants belonging to the Solanaceae family [33]. The intermediate Δ2 can then indulge in vicinal 2α,3β-diol and 2β,3β-diol formation, e.g., the glycosides 38–40, the wondosterols A, B and C isolated from a two sponge association of Poecillastra wondoensis and Japsis wondoensis [34]. With the ring A becoming oxygen rich, the 19β-Me becomes amenable for oxidation to –COOH group and consequent lactonisation with the 2β-OH, as seen in the pregnane γ-carbolactones 41–43 isolated from the Hawaiian sponge Strongylophora sp. [35]. The free COOH group can also disappear by loss of CO2 leading to ring-A aromatisation found in the sterols 44 geodisterol isolated from Geodia sp. from Papua New Guinea [36], and the 19-nor pregnane glycoside 45 from Cribrochalina olemda from Pohnpei, Micronesia [37]. Oxidative elimination of the 19-Me takes place rather easily in sponges belonging to Axinellideae, e.g., Axinella polypoides, which contains 46 as the important sterol [38–40]. Its precursor A/B ring structure containing the 19-COOH group is present in the sterols 47–49 isolated from Toxadocia zumi [41].
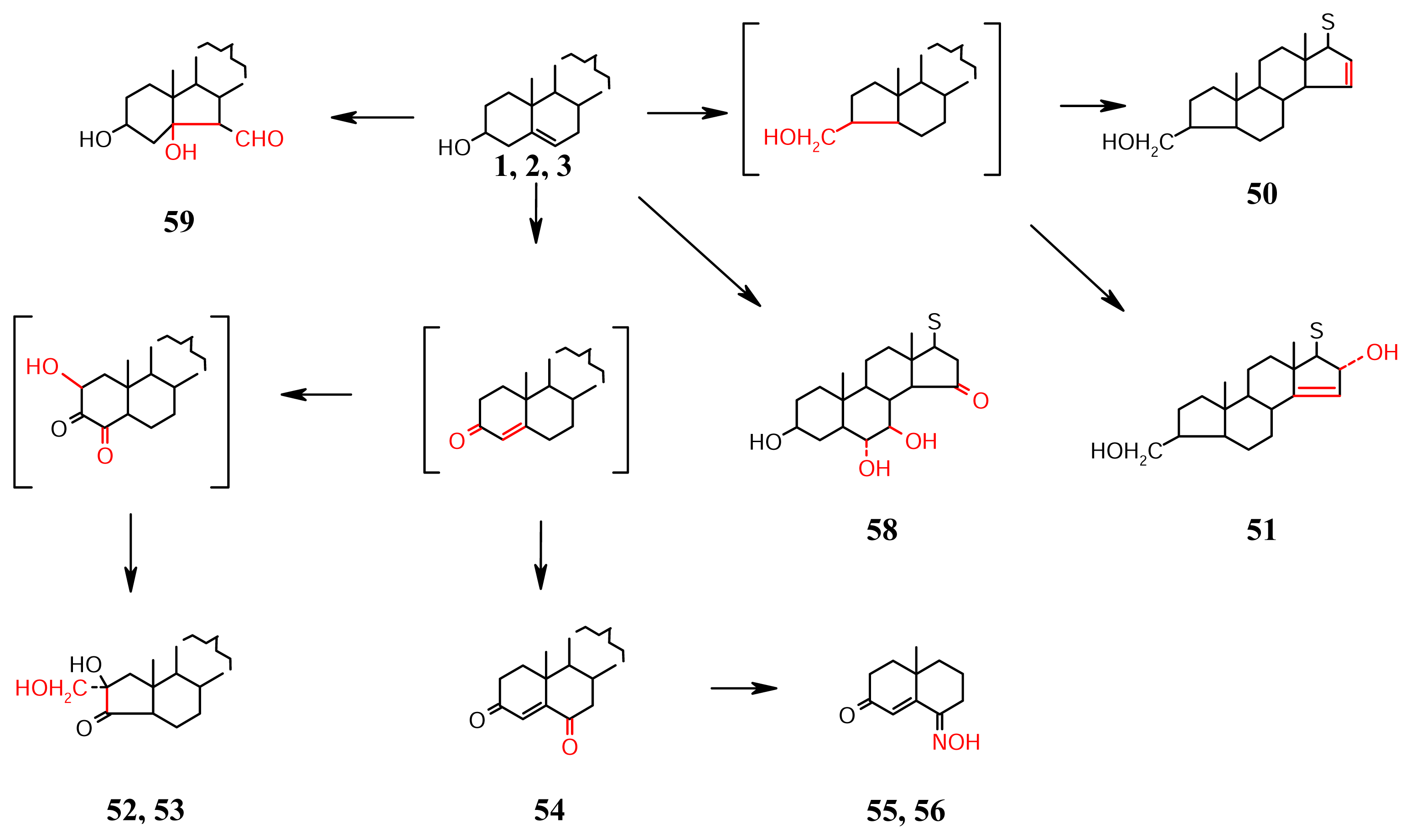
- Oxidations and rearrangements in theA/B ring system (Scheme 5): In sponges, ring Arearranged sterols cooccur with 3-keto sterols, and 3,6-diones, a phenomenon that is particularly unique in sponges belonging to the families Axinellideae and Hymeniacidonidae. This is attributed to an efficient enzyme system due to which the A/B ring reaction precedes oxidation at other centres, e.g., Δ15 introduction in 50, and Δ14-16α-OH system in 51 found in the sterols of Axinella proliferans from Reunion island in the Indian Ocean [42]. The biosynthesis of the unique 52 and 53 (anthosterones A and B respectively) of Anthoracurata gracia is suggested to take place by a benzilic acid rearrangement of a 2,3-diketo precursor as a new type of ring A contraction step [43].The Δ4-3,6-diketosterols 54, with several conventional side chains are also from Anthoracurata gracia, the sponge from which anthosterones 52 and 53 are isolated [43]. The 3,6-diketones of Geodia cydonium [44] and Cinachyra tarentina [45] co-occur with the more common 3-ketones [46]. The 6-oximino-3-ketones 55 and 56 were obtained from a mixture of Cinchyrella alloclada and C. apion [47]. The 5α,6α-dihydroxylation is seen in 57 from Spirastrella inconstans from India [48], and the 6α,7β-dihydroxylation is seen in 58 clathriol from Clathria lissosclera [49] of New Zealand. The former seems to be the precursor of ring B rearranged 59 orostanal isolated from Stellata hiwasaensis of Japan [50]. The sterol 59 is cytotoxic and apoptosis-inducing.
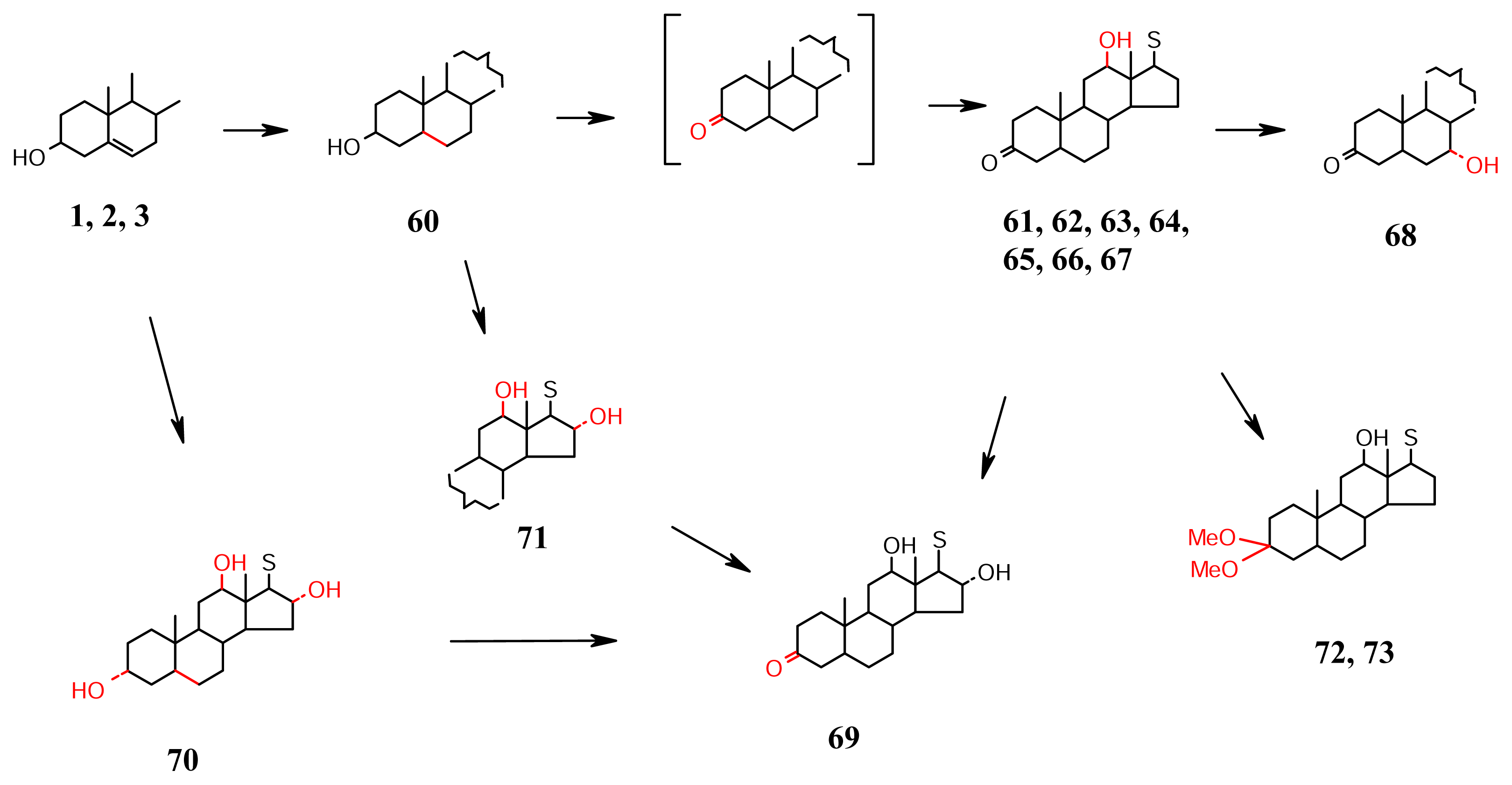
- Ring C oxidation (Scheme 6): The ring C site of oxidation at C-12 may not be requiring activation offered by a Δ5, a Δ7 or a 3β-OH. The saturated sterol 60 is in fact isolated in this group from Rhizochalina incrustata [51]. The activation seems to be coming from the heavily oxygenated (cyclopropane ring containing) side chains, c.f., the potent antitumour 61 [52], and 62–64 [53] from Xestospongia sp., which are named aragusterols A to D, and 65 and 66 [54] and 67 [55] named as xestosterol A, xestosterol B and aragusterol E respectively, from another Xestospongia sp. collected from Okinawa. In rare cases, a further hydroxylation occurs at C-7, e.g., 68 xestokerol B; [54] isolated along with xestokerols A, B and D and C-16, e.g., 69 [55], another aragusterol (aragusterol F) of the Xestospongia sp from Okinawa. The skeletons 70 and 71 are of aragusterols G and H respectively, also isolated from this collection [55]. The sterols 72, and 73 are aragusteroketals A and C respectively that are also from the same sponge [56], and perhaps artifacts of the isolation procedure.
II. Δ7-Sterols
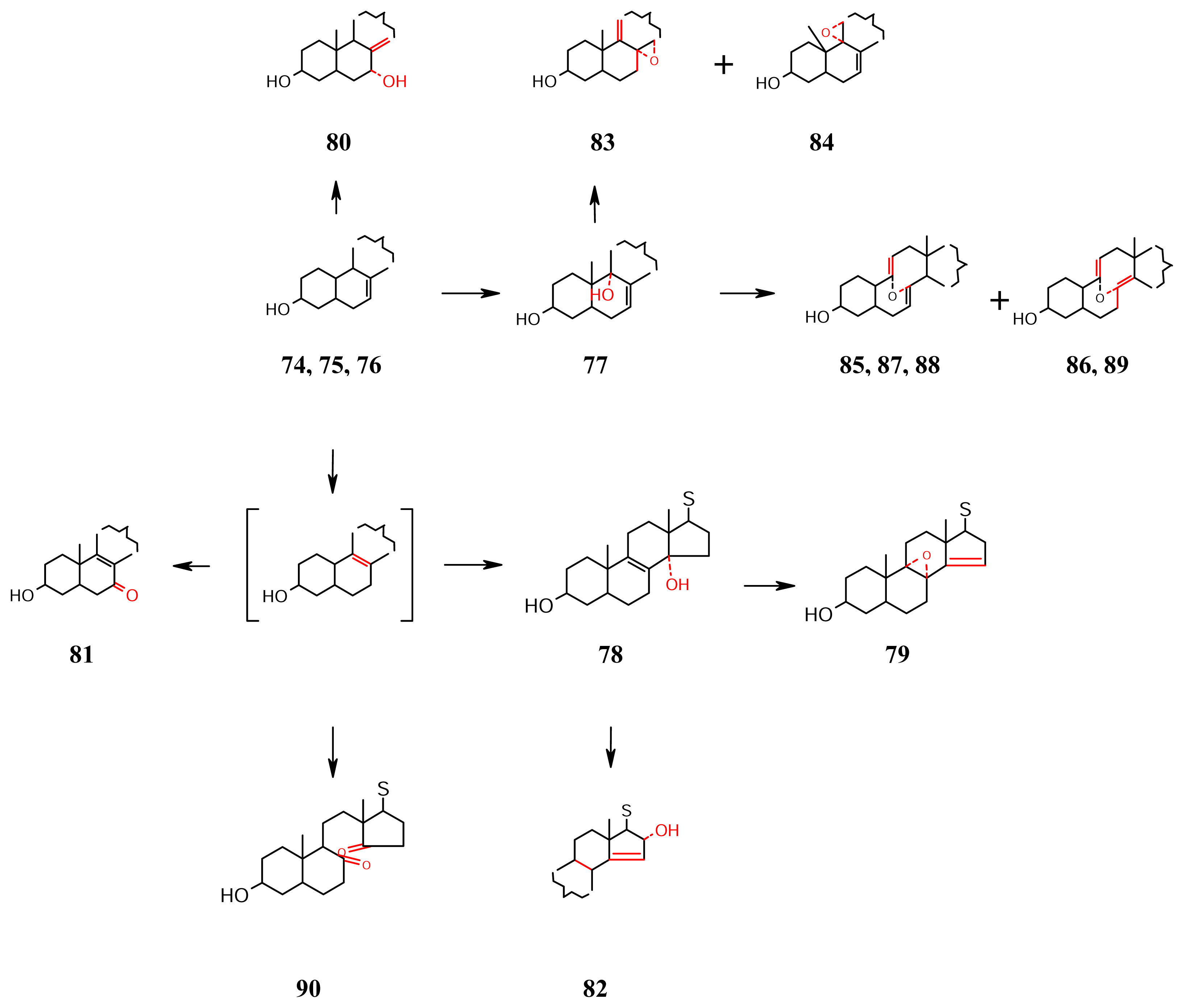
- Oxidation involving C-7, C-8, C-9, C-11 and C-14 (Scheme 7): The 3β-hydroxy-5,6-dihydro-Δ7 sterol nucleus seems to be undergoing allylic C-9 and C-14 (of the isomerised Δ8 nucleus) oxidation pathways. The C-9 oxidized 77 from Jericopsis graphidiophora [59] co-occurs with the C-14 oxidized 78 and 79 [60].The Δ8 migrated to Δ8(14) while 8α-OH is formed in 80 isolated from Pellina semitubulosa [20]. The Δ8 -7-ketone 81 is from Jereicopsis graphidiophora [59]. The Δ14-16-α-hydroxy sterol 82 is from the Mediterranean sponge Topsentia aurantiaca [61]. Extension of unsaturation to Δ9(11) followed by epoxidation is behind 83 and 84 [62]. The products of retro Diels-Alder reaction followed by cyclic ether formation, viz., 85–87, and their 3-methyl ethers 88, and 89 are from Microscleroderma spirophora from Senegal [60] that cooccur with the 8,14-seco-8,14-dione 90.
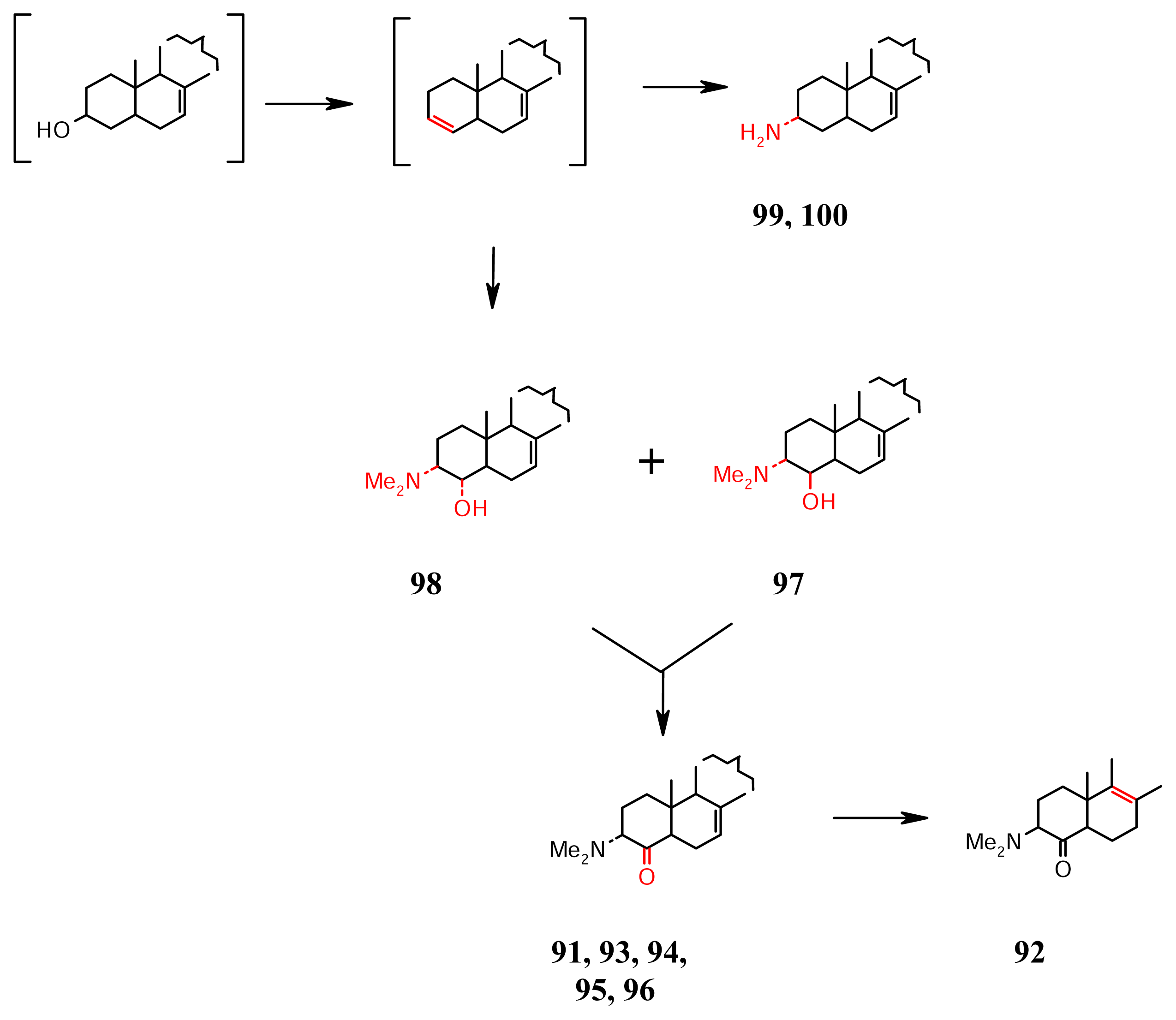
- Sterol amines (Scheme 8): The steroidal alkaloids, plakinamines 91–95 are α-amino ketones that are significantly cytotoxic from a Corticium sp. from Vanuatu [63]. Recently, it is found that the aminoketones (e.g., 96 plakinamine F) cooccur with the aminohydrins, e.g., 97 (plakinamine E) in the Corticium sp. of Guam [64], and 98 in a Vanuatuan collection of the same sponge [65]. The amines 96 and 97 have moderate cytotoxicity and antifungal activity, and nucleic acid-cleaving property. These aminohydrins probably formed via the addition of the elements of (CH3)2NOH across a Δ3 which may be responsible for the aminoketones cited above. The 3-amino steroids 99 and 100 that result from the addition of NH3 across Δ3 are also isolated from the Vanuatuan collection [65].
III.Δ5,7-Sterols
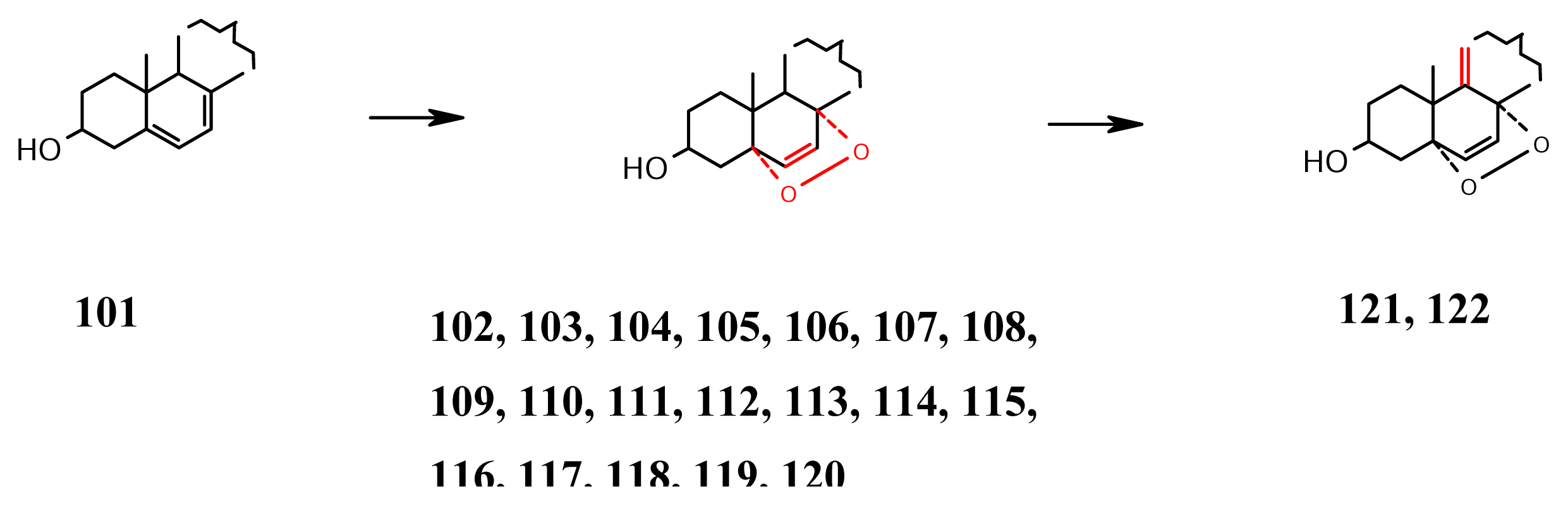
- Epidioxides (Scheme 9): Endoperoxides are routinely prepared in the laboratory by the action of singlet oxygen on cyclic conjugated dienes. Hence, when the endoperoxides 102–106 were isolated from Tethya aurantia [67] and 102, 107 and 108 from Axinella cannabina [68], it was suspected that they might be artifacts. However, such epidioxides continue to be isolated even when extreme care is taken to prevent their possible formation during extraction and isolation procedure. Thus, the Okinawan sponge Axinyssa sp. gave 109 axinysterol [69], and Lendenfeldia chondroides from Palau gave the antifouling sterols 110 and 111 [70]. The sponge species Luffariella cf. variabilis of Japan gave a mixture of the sterol epidioxides 112–120, accompanied with the cytotoxic 121, possessing extra Δ9(11) double bond [71], which system is also present in 122, recently isolated [72] from the same Axinyssa sp. that earlier gave 109 axinysterol [69] and which inhibits the growth of several human cancer cell lines.
- Epoxy derivatives of Δ5,7system (Scheme 10): The 1,2-oxides of the Δ5,7 sterols are predominantly α,α. The intact epoxide 123 and 124 its Δ8(14) isomer, both having cytotoxicity to a range of human and murine cell lines are isolated recently [73] from Polymastia tenax. These 7α-alcohols are associated with the dienone 125 in the sponge. This typical dienone structure containing steroids were earlier isolated as 126, 127 and 128 from Clathrina clathrus [74]. The 5α,6α-epoxy-7α-hydroxy-Δ8(14) system is also present in 129 isolated from an Indian specimen of Ircinia fasciculata [75] which should be the biogenetic precursor of 130 [76]. The 5α,6α-epoxy group opens up in a number of possible ways (see also Scheme 1), producing 5α,6α-dihydroxy system, 5α,6β-dihydroxy system, and the 5α-H,6α-hydroxy system. The 5β,6β-epoxide system also occurs in which the 3β-OH had epimerised to 3α-OH. The opening of this epoxide also proceeds in a number of ways, e.g., 5α,6β-dihydroxy system, 5β,6α-dihydroxy system and 5β-H,6β-hydroxy sterols. In each case, the Δ7 causes activation of sites for further modification of the sterol structure.The 5α,6α-dihydroxy system is evidenced in sterols 131 [77], 132 [78], and 133 [79] which are products of oxidation at extended sites. The sterol 131 is from Dysidea sp. from Northern Australia, and contains the additional 9α,11α-epoxide of a Δ9(11), itself made possible by action from Δ8. The sterols that co-occur with 131 in the sponge are 134 and 135, in which the C-11 activation is in evidence. The sterols 134 and 135 inhibit the binding of IL-8 to the human recombinant IL-8 receptor type A. The sterol 132, also containing the 9α,11α-epoxide is from an unidentified species of Dysidea collected from Guam [78]. In this sterol, the 19-Me is additionally hydroxylated. The sterol 133 is from D. herbaceae [79] from Ethiopia. This sponge is unique since each of the four sterols 136, 133, 137 and 138 isolated from it represents one type of 5,6-epoxide (or its opening), viz., a trans opening of the 5α,6α-epoxide, a cis opening of the 5α,6α-epoxide, a trans opening of the 5β,6β-epoxide of the 3α-hydroxy sterol and the 5β,6β-epoxy-4α-hydroxy sterol itself respectively.The 5α,6β-dihydroxy system is shown in addition to 136, in 139–148. The sterol 139 and 140 are from D. fragilis [80] collected in the Black Sea. The eight sterols 141–148 are from D. etheria from Bermuda [81]. The 5α-H,6α-hydroxy system is present in 149 [82] and 150 [83]. It is also present in 151 obtained from a Japanese Spongia sp. [84] which also gave 152–157 [85]. The unique feature of these six sterols is the presence of 4β-oxygen function. Further products of the 5β,6β-epoxide opening, in addition to 137 of Dysidea herbaceae [79] are the A/B cis 158–160 obtained from the same species of D. etheria that gave the A/B trans 141–148; hence, the unique ability of the two species of Dysidea. D. herbaceae is further unique for its 161 herbasterol [86], a 5β-H-9(11)-seco steroid, which is ichthyotoxic and antimicrobial. The cyclic ether 162 is from D. tupha of the Mediterranean [87].
- 9(11)-Seco Steroids: A Δ9(11) activation produces the 9α,11α-vicinal diol system which in turn appears to be responsible for the producion by retro-Diels Alder reaction, the 9,11-seco ketoaldehydes 163–165 luffasterols A, B and C present in Luffariella sp. from Palau [88], 166 [45] and 167 [89] isolated from the Mediterranean sponge Spongia officinalis. The keto aldehyde 166 goes to the keto alcohols 168 and 169 [45] in the sponge. The epoxy keto alcohol 170 glaciasterol B-3-acetate of Fasciospongia cavernosa which is toxic to brine shrimp, also from the Mediterranean [90], is however not associated with its corresponding aldehyde as also in the case of 171 blancasterol from the NE Pacific sponge Pleraplysilla sp. [91] from Vancouver and 172 from a Japanese species of Stelletta [92]. In the antihistaminic secosterols 173–182 of Euryspongia sp. from New Caledonia [93]; the 2-OH which is usually β in this series is epimerised to α-OH.
- Oxidation not involving 5α,6α-epoxide (Scheme 11): The reactions of the Δ5,7 system without the mediation of the 5α,6α-epoxide come under this group, e.g., 183 from an Indian specimen of Suberites carnosus [94]. Of particular significance is the methylation at C-4, activated by Δ5, as indicated by the occurrence of 184 polymastiamides A [95], and 185–189 polymastiamides B to F in Polymastia baletiformis from Norway, of which A, C, D and F have the 4α-Me substituent and B and E do not have substitution at C-4 [96]. The mildly cytotoxic 190 from Theonella swinhoei from Phillippines, has instead a C-4 methylene group, a group that also occurs in the sterols 191–193 from T. swinhoei from Okinawa [97]. In 191 and 193, the Δ8(14) underwent oxidation to give the 8–14 seco-8, 14-dione. The C-4 activation leading to a 4α-oxysulfate substitution is noticed in the ten sterols 194–203 acanthosterol sulfates A to J from Acanthodendrilla sp. from Japan [98]. Of these, 202 (acanthosterol sulfate I) and 203 (acanthosterol sulfate J) showed antifungal activity and cytotoxicity.

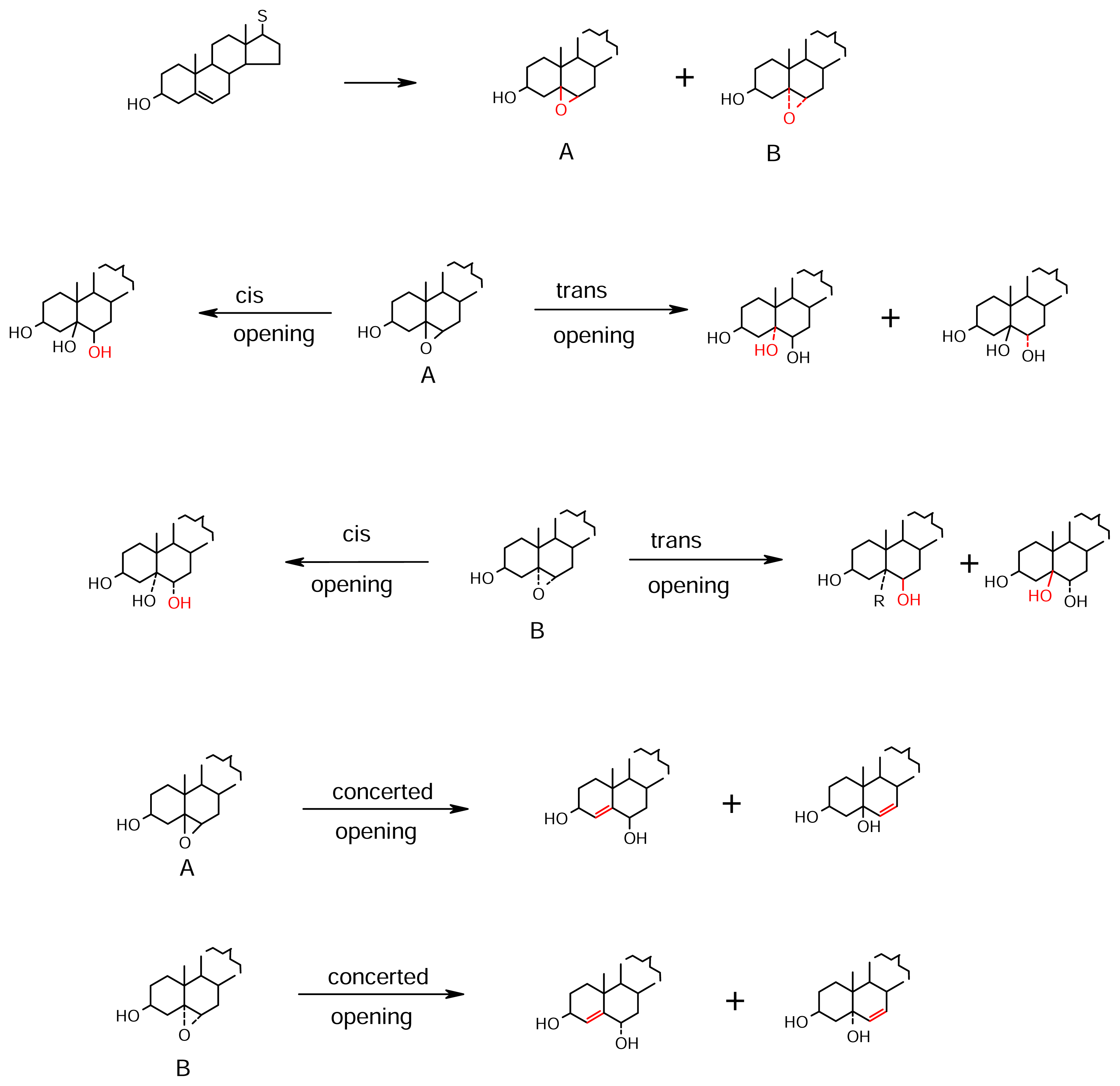{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
IV. 3α-Hydroxy Steroids
- Δ5-Origin (Scheme 12): The ring system of the sulfated steroids has a lone representative containing unsaturation in 204 [99]; all others are saturated, cf., 205 halistanol B sulfate from Pachastrella sp. [100] that inhibits endothelium converting enzyme. Weinbergsterols 206 (A) and 207 (C), have hydroxylation at C-16 while weinbergsterol B 208 has further hydroxylation at C-18; they are isolated from Petrosia weiinbergii [101,102]. The disulfates 209, 210 and 207 are sterol orthoesters involving 16β-OH (and 20-OH and 22-O-butyrate of the regular side chain), isolated from the same sponge. In this group, the 15α,16β-dihydroxylation is seen in 211 clathsterol with anti HIV-1 reverse transcriptase activity from an Eritrean sponge of genus Clathria [103]. The cytotoxic and antifungal 212 echinoclasterol with heavily oxygenated ring E is from the south Australian sponge Echinoclathria subhispida [104].
- Δ5,7-Origin (Scheme 13): The 3α-sulfate esterification is more prolific when the genesis is from the Δ5,7 sterol skeleton. The activation of ring carbons by Δ7 seems to extend to C-15α by migration of Δ7 to Δ8(14). In this group, 213 is halistanol sulfate from Halichondria moorei [99] which has potential activity against HIV virus. It is the forerunner of several halistanol sulfates, e.g., 214–217 halistanols A to D from Epipolasis sp. [105], and 218 to 220, in vitro HIV inhibiting halistanol sulfates F to H from Pseudoaxynissa digitata [106]. The sterol 221 which showed inhibition in guanosine diphosphate/G protein RAS exchange assay is ophirapstanol trisulfate from Topsentia ophiraphidites [107]. The sterol 222 is sokotrasterol sulfate isolated from two Halichondriidea species [108], and 223 is norsokotrasterol sulfate from Trachyposis halichondroides [109]. The sterol 224 is from a Japanese specimen of Topsentia sp. [110]. The trioxysulfate 29 polysterol B sulfate of a Japanese specimen of Epipolasis sp. is accompanied in the sponge with 28 polysterol A [27], a sterol that belongs to group 1 as mentioned earlier (Scheme 3).In Epipolasis sp. [105], the trioxysulfates are associated with the product 225 of further hydroxylation at C-15. The 15-ketosterol 226 xestobergsterol A, that inhibits the release of histamine from rat mast cells was isolated from Xestospongia bergquistii [111]. It was earlier isolated from Ircinia sp. from Okinawa [112]. The simultaneous activation of 6α by Δ4 and 7β and 15α by Δ8(14) followed by oxidation of the 15-OH to 15-ketone appears to be taking place in 227 contignasterol [113]. In these 15-keto sterols, the configuration at C-14 is 14βH as opposed to the usual 14αH configuration. The same 14βH configuration is noticed in 228 xestobergsterol C and the further 1β-hydroxylated steroid 229 xestobergsterol B from the above Ircinia sp. The sterol 230 with an additional 4α-OH from the Malaysian Haliclona sp. [114] and 231 from a new species of Oceanapia [115] also contain this ring structure. The sterol 231 is in fact accompanied with its 14αH epimer 232 in the sponge. Hence, a switchover of the original 14αH configuration to the more stable 14βH is indicated in these ketones. The reduction thereafter of the 15-ketone to 15β-OH should be responsible for 233 and 234 of two Philippine unidentified Haplosclerid sponges [116].
- 14α-Methylation (Scheme 14): The 14α-methylation is more common among tetracyclic triterpenes of land plants, e.g., lanosterol (VIII). This feature, together with the 4,4-dimethylation over the C19 cholesterol nucleus gives the usual C22 tetracyclic triterpene nucleus. The 14α-sterols of sponges all possess a Δ9(11)-unsaturation indicating that biological methylation in these sterols by 1,2-addition is facilitated in a homoannular-1,3-diene ring C as shown in the Scheme 14, cf., 235 lembehsterol B with Δ5 retained from the Indonesian Petrosia strongylata isolated together with the 6-O-sulfate ester viz., 236 lembehsterol A [117]. This steroid ring system was earlier found in 237 ibisterol sulfate (which is cytoprotective against the HIV-1 virus) from Topsentia sp. [118] and later also in 238 and 239, ibisterols B and C of a Phillippine sponge Xestospongia sp. These two sterols are associated with the ketoepoxide 240 [119]. The sterols 238, 239 and 240 are inhibitors of HIV-I integrase. In 241 to 245, topsentiasterol sulfates A to E isolated from an Okinawan Topsentia sp. have the additional 4β-OH group [120].
Conclusions














Acknowledgements
References and Notes
- Goad, L.J. The sterols of marine invertebrates: composition, biosynthesis and metabolites. In Marine Natural Products, Chemical and Biological Perspectives, Vol. II; 1978; Scheuer, P. J., Ed.; Academic Press: New York; pp. 76–172. [Google Scholar]
- Schmitz, F.J. Uncommon marine steroids. In Marine natural products, chemical and biological perspectives Vol, I; 1978; Scheuer, P. J., Ed.; Academic Press: New York; Volume I, pp. 241–297. [Google Scholar]
- D’Auria, M.V.; Minale, L.; Ricco, R. Polyoxygenated steroids of marine origin. Chem. Rev 1993, 93, 1839–1895. [Google Scholar]
- Stonik, V.A. Marine polar steroids. Russ. Chem. Rev. 2001, 70, 673–715. [Google Scholar]
- Kanazawa, A. Sterols in marine invertebrate. Fish. Sci. 2001. [Google Scholar]
- Garson, M.J. Biosynthetic studies on marine natural products. Nat. Prod. Rep 1989, 6, 143–170. [Google Scholar]
- Garson, M.J. The biosynthesis of marine natural products. Chem. Rev. 1993, 93, 1699–1733. [Google Scholar]
- Thomas, R. Biogenic speculation and biosynthetic advances. Nat. Prod. Rep 2004, 21, 224–248, , and earlier reviews.. [Google Scholar]
- De Luca, P.; De Rosa, M.; Minale, L.; Sodano, G. Marine sterols with a new pattern of side chain alkylation from the sponge Aplysina(Verongia) aerophoba. J. Chem. Soc. Perkin Trans. I. 1972, 2132–2135. [Google Scholar]
- Fattorusso, E.; Megno, S.; Mayol, L.; Santacroce, C.; Sica, D. Calysterol: A C29 cyclopropenecontaining marine sterol from the sponge Calyx nicaensis. Tetrahedron 1975, 31, 1715–1716. [Google Scholar]
- Sica, D.; Zollo, F. Petrosterol, the major sterol with a cyclopropane side chain in the sponge Petrosia ficiformis. Tetrahedron Lett 1978, 19, 837–838. [Google Scholar]
- Ravi, B. N.; Kokke, W.C.M.C.; Delseth, C.; Djerassi, C. Isolation and structure of 26,27-cycloaplysterol (petrosterol) a cyclopropane-containing marine sterol. Tetrahedron Lett 1978, 45, 4379–4380. [Google Scholar]
- Bortolotto, M.; Braekman, J.C.; Daloze, D.; Tursch, B. Chemical studies of marine invertebrates XXXVI, strongylosterol, a novel C-30 sterol from the sponge Strongylophora durissima dendy. Bull. Soc. Chim. Belg. 1978, 87, 539–543. [Google Scholar]
- Kokke, W.C.M.C.; Tarchini, C.; Stierle, D.B.; Djerassi, C. Isolation, structure elucidation and partial synthesis of xestosterol, biosynthetically significant sterol from the sponge Xestospongia muta. J. Org. Chem 1979, 44, 3385–3388. [Google Scholar]
- Kerr, R.G.; Kerr, S.L.; Pettit, G.R.; Harald, D.L.; Groy, T.L.; Djerassi, C. Sterols of marine invertebrates. 63. Isolation and structure elucidation of sutinasterol, the major sterol of the marine sponge Xestospongia sp. J. Org. Chem. 1991, 56, 58–62. [Google Scholar]
- Li, L.N.; Li, H.T.; Lang, R.W.; Itoh, T.; Sica, D.; Djerassi, C. Minor and trace sterols in marine invertebrates. 31. Isolation and structure elucidation of 23H-isocalysterol, a naturally occurring cyclopropene. Some comparative observations on the course of hydrogenolytic ring opening of steroidal cyclopropenes and cyclopropanes. J. Am. Chem. Soc 1982, 104, 6726–6732. [Google Scholar]
- Doss, G.A.; Djerassi, C. Sterols in marine invertebrates. 60. Isolation and structure elucidation of four new steroidal cyclopropenes from the sponge Calyxpodatypa. J. Am. Chem. Soc 1988, 110, 8124–8128. [Google Scholar]
- Reddy, N. S.; Ramesh, P.; Venkateswarlu, Y. A novel C−27 nor steroid from the marine sponge Petrosia testudinaria. Nat. Prod. Lett 1999, 14, 131–134. [Google Scholar]
- Guerriero, A.; D’Ambrosio, M.; Fransesco, P.; Debitus, C.; Ribes, O. Pteridines, Sterols, and indole derivatives from the Lithistid sponge Corallistes undulatus of the coral sea. J. Nat. Prod 1991, 56, 1962–1970. [Google Scholar]
- Notaro, G.; Piccialli, V.; Sica, V.; Pronzato, R. New Δ8(14)-3β,7α-dihydroxysterols from the marine sponge Pellina semitubulosa. J. Nat. Prod 1992, 55, 773–779. [Google Scholar]
- Hirota, S.H.; Takayama, S.; Miyashiro, Y.; Ozaki, S.I. Structure of a novel steroidal saponin, pachastrelloside A, obtained from a marine sponge of the genus Pachastrella. Tetrahedron Lett 1990, 31, 3321–3324. [Google Scholar]
- Gallimore, W.A.; Kelly, M.; Scheuer, P.J. Gelliusterols A-D, new acetylenic sterols from a sponge, Gellius Species. J. Nat. Prod 2001, 64, 741–744. [Google Scholar]
- Xu, S.H.; Zeng, L.M. Study on the chemical constituents of marine sponge Polymastia sobustia. Youji Huaxue 2001, 21, 45–48. [Google Scholar]
- Xu, S.H.; Zeng, L.M. The identification of two new sterols from marine organism Polymastia sobustia. Chin. Chem. Lett 2000, 11, 531–534. [Google Scholar]
- Zhang, W.; Che, C. Isomalabaricane-type nortriterpenoids and other constituents of the marine sponge Geodia japonica. J. Nat. Prod 2001, 64, 1489–1492. [Google Scholar]
- Umeyama, A.; Ito, S.; Yoshigaki, A.; Arihara, S. Two new 26,27-cyclosterols from the marine sponge Strongylophora corticata. J. Nat. Prod 2000, 63, 1540–1542. [Google Scholar]
- Umeyama, A.; Adachi, K.; Ito, S.; Arihara, S. New 24-isopropylcholesterol and 24-isopropenylcholesterol sulfate from the marine sponge Epipolasis species. J. Nat. Prod 2000, 63, 1175–1177. [Google Scholar]
- Ishibashi, M.; Yamagishi, E.; Kobayashi, J. Topsentionols A~J, new sterols with highly branched side chains from marine sponge Topsentia sp. Chem. Pharm. Bull 1997, 45, 1435–1438. [Google Scholar]
- Rochfort, S.J.; Gable, R.W.; Capon, R.J. Mycalone: a new steroidal lactone from a southern Australian marine sponge, Mycale sp. Aust. J. Chem 1996, 49, 715–718. [Google Scholar]
- Carney, J.R.; Scheuer, P.J.; Kelley-Borges, M. Three unprecedented chloro steroids from the Maui sponge Strongylacidon sp.: kiheisterones C, D, and E. J. Org. Chem 1993, 58, 3460–3462. [Google Scholar]
- Prinsep, M.R.; Blunt, J.W.; Murray, H.; Munro, G. A new sterol sulfate from the marine sponge Stylopus australis. J. Nat. Prod 1989, 52, 657–659. [Google Scholar]
- Kalinovsky, A.I.; Antonov, A.S.; Afiyatullov, S.S.; Dmitrenok, P.S.; Evtuschenko, E.V.; Stonik, V.A. Mycaloside A, a new steroid oligoglycoside with an unprecedented structure from the Caribbean sponge Mycale laxissima. Tetrahedron Lett 2002, 43, 523–525. [Google Scholar]
- Kirson, I.; Glotter, E. Recent developments in naturally occurring ergostane-type steroids. A review. J. Nat. Prod 1981, 44, 633–647. [Google Scholar]
- Ryu, G.; Choi, B.W.; Lee, B.H.; Hwang, K.H.; Lee, U.C.; Jeong, D.S.; Lee, N.H. Wondosterols A-C, three steroidal glycosides from a Korean marine two-sponge association. Tetrahedron 1999, 55, 13171–13178. [Google Scholar]
- Corgiat, J.M.; Scheuer, P.J.; Rios Steiner, J.L.; Clardy, J. Three pregnane-10,2-carbolactones from a sponge, Strongylophora sp. Tetrahedron 1993, 49, 1557–1562. [Google Scholar]
- Wang, G.Y.S.; Crews, P. Geodisterol, a novel polyoxygenated sterol with an aromatic A ring from the tropical marine sponge Geodia sp. Tetrahedron Lett 1996, 37, 8145–8146. [Google Scholar]
- Yeung, B.K.S.; Hamann, M.T.; Scheuer, P.J.; Kelly-Borges, M. Hapaioside: A 19-norpregnane glycoside from the sponge Cribrochalina olemda. Tetrahedron 1994, 50, 12593–12598. [Google Scholar]
- De Rosa, M.; Minale, L.; Sodano, G. Metabolism in porifera. V. Biosynthesis of 19-norstanols. Conversion of cholesterol into 19-norcholestanols by the sponge Axinella polypoides. Experientia 1975, 31, 758–759. [Google Scholar]
- Minale, L.; Persico, D.; Sodano, G. Metabolism in porifera. IX. Studies on the biological conversion of cholesterol into 19-nor-cholestanol by the sponge Axinella polypoides. Experentia 1979, 35, 296–297. [Google Scholar]
- Crist, B.V.; Djerassi, C. Minor and trace sterols in marine invertebrates 47. A re-investigation of the 19-nor stanols isolated from the sponge Axinella polypoides. Steroids 1983, 42, 331–343. [Google Scholar]
- Nakatsu, T.; Walker, R.P.; Thompson, J.E.; Faulkner, D.J. Biologically-active sterol sulfates from the marine sponge Toxadocia zumi. Experientia 1983, 39, 759–761. [Google Scholar]
- Aknin, M.; Gaydou, E.M.; Boury-Esnault, N.; Costantino, V.; Fattorusso, E.; Mangoni, A. norsterols in Axinella proliferans sponge from Indian Ocean. Comp. Biochem. Physiol 1996, 113B, 845–848. [Google Scholar]
- Tischler, M.; Ayer, S.W.; Andersen, R.J.; Mitchell, J.F.; Clardy, J. Anthosterones A and B, ring A-contracted steroids from the sponge Anthoracuata graceae. Can. J. Chem 1988, 66, 1173–1178. [Google Scholar]
- Migliuolo, A.; Piccialli, V.; Sica, D. Steroidal ketones from the sponge Geodia cydonium J. Nat Prod 1990, 53, 1262–1266. [Google Scholar]
- Migliuolo, A.; Piccialli, V.; Sica, D. Structure elucidation and synthesis of 3β,6α-dihydroxy-9-oxo-9, 11-seco-5α-cholest-7-en-11-al, a novel 9,11-secosterol from the sponge Spongia officinalis. Tetrahedron 1991, 47, 7937–7950. [Google Scholar]
- Aiello, A.; Fattorusso, E.; Magno, S.; Menna, M.; Pansini, M. Steroids of the marine sponge Cinachyra tarentina: Isolation of cholest-4-ene-3,6-dione and (24R)-24-ethylcholest-4-ene-3,6-dione. J. Nat. Prod. 1991, 54, 281–285. [Google Scholar]
- Rodriguez, J.; Nunez, L.; Peixinho, S.; Jimenez, C. Isolation and synthesis of the first natural 6-hydroximino 4-en-3-one-steroids from the sponges Cinachyrella spp. Tetrahedron Lett 1997, 38, 1833–1836. [Google Scholar]
- Das, B.; Rao, S.P.; Srinivas, K.V.M.S. Studies on marine chemicals, Part VI. A new clionasterol derivative from the marine sponge Spirastrella inconstans. J. Nat. Prod 1993, 56, 2210–2211. [Google Scholar]
- Keyzers, R.A.; Northcote, P.T.; Webb, V. Clathriol, a novel polyoxygenated 14β steroid isolated from the New Zealand marine sponge Clathria lissosclera. J. Nat. Prod 2002, 65, 598–600. [Google Scholar]
- Miyamoto, T.; Kodama, K.; Aramarki, Y.; Higuchi, R.; van Soest, R.W.M. Orostanal, a novel abeo-sterol inducing apoptosis in leukemia cell from a marine sponge, Stelletta hiwasaensis. Tetrahedron Lett 2001, 42, 6349–6351. [Google Scholar]
- Makar’eva, T.N.; Stonik, V.A.; Ponomarenko, L.P.; Kalinovsky, A.I. Isolation of (24R)-24,25-methylene-5α-cholestan-3β-ol, a new cyclopropane – containing sponge sterol. J. Chem. Res. (S) 1996, 10, 468–469. [Google Scholar]
- Iguchi, K.; Fujita, M.; Nagaoka, H.; Mitome, H.; Yamada, Y. Aragusterol a: A potent antitumor marine steroid from the okinawan sponge of the genus, Xestospongia. Tetrahedron Lett 1993, 34, 6277–6280. [Google Scholar]
- Iguchi, K.; Shimura, H.; Taira, S.; Yokoo, C.; Matsumoto, K.; Yamada, Y. Aragusterol B and D, new 26,27-cyclosterols from the Okinawan marine sponge of the genus Xestospongia. J. Org. Chem 1994, 59, 7499–7502. [Google Scholar]
- Kobayashi, J.; Ishida, K.; Naitoh, K.; Shigemori, H.; Mikami, Y.; Sasaki, T. Xestokerols A, B, and C, new C29 steroids with a cyclopropane ring from the Okinawan marine sponge Xestospongia sp. J. Nat. Prod 1993, 56, 1350–1355. [Google Scholar]
- Miyaoka, H.; Shinohara, M.; Shimomura, M.; Mitome, H.; Yano, A.; Iguchi, K.; Yamada, Y. Aragusterols E-H, new 26,27-cyclosterols from the Okinawan marine sponge of the genus Xestospongia and absolute configurations of xestokerols A and B. Tetrahedron 1997, 53, 5403–5412. [Google Scholar]
- Kobayashi, M.; Chen Higuchi, Y. J. K.; Aoki, S.; Kitagawa, I. Marine natural products. XXXVII. Aragusteroketals A and C, two novel cytotoxic steroids from a marine sponge of Xestospongia sp. Chem. Pharm. Bull. 1996, 44, 1840–1842. [Google Scholar]
- Bultel-Ponce, V.; Brouard, J.P; Vaeelet, J.; Guyot, M. Thymosiosterol and Δ24 thymosiosterol, new sterols from the sponge Thymosiopsis sp. Tetrahedron Lett. 1999, 40, 2955–2956. [Google Scholar]
- Gunasekera, S.P.; Kelly-Borges, M.; Longley, R.E. A new cytotoxic sterol methoxymethyl ether from a deep water marine sponge Scleritoderma sp. cf paccardi. J. Nat. Prod 1996, 59, 161–162. [Google Scholar]
- D’Auria, M.V.; Gomez, P.L.; Minale, L.; Riccio, R.; Debitus, C. Jereisterol A and B : Two 3β-methoxy-secosteroids from the pacific sponge Jereicopsis graphidiophora. Tetrahedron Lett 1991, 32, 2149–2152. [Google Scholar]
- Costantino, V.; Fattorusso, E.; Mangoni, A.; Aknin, M.; Gaydou, E.M. Novel 3β-methoxysteroids from the Senegalse sponge Microscleroderma spirophora. Steroids 1994, 59, 181–184. [Google Scholar]
- Ciminiello, P.; Fattorusso, E.; Magno, S.; Mangoni, A.; Pansini, M. Three new D-ring unsaturated sterols from the Mediterranean sponge Topsentia aurantiaca: Structure determination and complete nuclear magnetic resonance assignment. Steroids 1992, 57, 62–66. [Google Scholar]
- D’Auria, M.V.; Gomez, P.L.; Minale, L.; Riccio, R.; Debitus, C.; Levi, C. Unique 3β-Omethylsterols from the Pacific sponge Jereicopsis graphidiophora. J. Nat. Prod 1992, 55, 311–320. [Google Scholar]
- De Marino, S.; Iorizzi, M.; Zollo, F; Roussakis, C.; Debitus, C. Plakinamine C and D and three other new steroidal alkaloids from the sponge Corticium sp. Eur. J. Org. Chem 1999, 697–701. [Google Scholar]
- Lee, H.S.; Seo, Y.; Rho, J.R.; Shin, J.; Paul, V.J. New steroidal alkaloids from an undescribed sponge of the genus Corticium. J. Nat. Prod 2001, 64, 1474–1476. [Google Scholar]
- Borbone, N.; De Marino, S.; Iorizzi, M.; Zollo, F.; Debitus, C.; Esposito, G.; Iuvone, T. Minor steroidal alkaloids from the marine sponge Corticium sp. J. Nat. Prod 2002, 65, 1206–1209. [Google Scholar]
- Hu, J.F.; Kelly, M.; Hamann, M.T. 26-nor-25-isopropyl-ergosta-5,7,22E-trien-3β-ol: a new C29 sterol from the sponge Agelas sceptrum from Jamaica. Steroids 2002, 67, 743–747. [Google Scholar]
- Sheikh, Y.M.; Djerassi, C. Steroids from sponges. Tetrahedron 1974, 30, 4095–4103. [Google Scholar]
- Cafieri, F.; Fattorusso, E.; Frigerio, A.; Santacroce, C.; Sica, D. Sterols of Axinella cannabina, a marine sponge. Gazz Chim Ital 1975, 105, 595–602. [Google Scholar]
- Iguchi, K.; Shimura, H.; Zhi, Yang; Yamada, Y. A new 5α,8α-epidioxy sterol from the okinawan marine sponge of the Axinyssa genus. Steroids 1993, 58, 410–413. [Google Scholar]
- Sera, Y.; Adachi, K.; Shizuri, Y. A new epidioxy sterol as an antifouling substance from a Palauan marine sponge, Lendenfeldia chondrodes. J. Nat. Prod 1999, 62, 152–154. [Google Scholar]
- Gauvin, A.; Smadja, J.; Ankin, M.; Faure, R.; Gaydou, E.M. Isolation of bioactive 5α,8α-epidioxy sterols from the marine sponge Luffariella cf. variabilis. Can. J. Chem 2000, 78, 986–992. [Google Scholar]
- Iwashima, M.; Terada, I.; Iguchi, K; Yamori, T. New biologically active marine sesquiterpenoid and steroid from the Okinawan sponge of the genus Axinyssa. Chem. Pharm. Bull 2002, 50, 1286–1289. [Google Scholar]
- Santafe, G.; Paz, V.; Rodriguez, J.; Jimenez, C. Novel cytotoxic oxygenated C29 sterols from the Colombian marine sponge Polymastia tenax. J. Nat. Prod 2002, 65, 1161–1164. [Google Scholar]
- Aiello, A.; Ciminiello, P.; Fattorusso, E.; Magno, S. Three new 7-keto sterols from the Mediterranean sponge Clathrina clathrus. Steroids 1988, 52, 533–542. [Google Scholar]
- Venkateswarlu, Y.; Reddy, M.V.R.; Rao, M.R. A new epoxy sterol from the sponge Ircinia fasciculate. J. Nat. Prod 1996, 59, 876–877. [Google Scholar]
- Aiello, A.; Fattorusso, E.; Magno, S.; Mayol, L.; Menna, M. Isolation of two novel 5α,6α-epoxy-7-ketosterols from the encrusting demospongia Oscarella lobularis. J. Nat. Prod 1990, 53, 487–491. [Google Scholar]
- Leone, P.A.; Redburn, J.; Hooper, J.N.A.; Quinn, R.J. Polyoxygenated Dysidea sterols that inhibit the binding of [I125] IL-8 to the human recombinant IL-8 receptor Type A. J. Nat. Prod 2000, 63, 694–697. [Google Scholar]
- Gunasekera, S.P.; Schimtz, F.J. Marine natural products: 9.alpha.,11.alpha.-epoxycholest-7-ene-3.beta.,5.alpha.,6.beta.,19-tetraol 6-acetate from a sponge, Dysidea sp. J. Org. Chem 1983, 48, 885–886. [Google Scholar]
- Isaacs, S.; Berman, R.; Kashman, Y.; Gebreyesus, T.; Yosief, T. New polyhydroxy sterols, dysidamides, and a dideoxyhexose from the sponge Dysidea herbacea. J. Nat. Prod 1991, 54, 83–91. [Google Scholar]
- Milkova, S.Ts.; Mikhova, B.P.; Nikolov, N.M.; Popov, S.S.; Andreev, S.N. Two new polyhydroxylated sterols from the sponge Dysidea fragilis. J. Nat. Prod. 1992, 55, 974–978. [Google Scholar]
- West, R.R.; Cardellina, J.H. Isolation and identification of eight new polyhydroxylated sterols from the sponge Dysidea etheria. J. Org. Chem 1988, 53, 2782–2787. [Google Scholar]
- Piccialli, V.; Sica, D. Four new trihydroxylated sterols from the sponge Spongionella gracilis. J. Nat. Prod 1987, 50, 915–920. [Google Scholar]
- Zhong, Y.; Su, Jing-Yu; Zeng, L.; Shen, W.; Wang, Q.W. Structure of a new sterol from the South China sponge Dysidea fragilis. Chin. J. Chem. 1993, 11, 560–564. [Google Scholar]
- Aoki, S.; Yoshioka, Y.; Miyamoto, Y.; Higuchi, K.; Setiawan, A.; Murakami, N.; Chen, Z.S.; Sumizawa, T.; Akiyama, S.; Akiyama, K. Agosterol A, a novel polyhydroxylated sterol acetate reversing multidrug resistance from a marine sponge of Spongia sp. Tetrahedron Lett 1998, 39, 6303–6306. [Google Scholar]
- Aoki, S.; Setiawan, A.; Yoshioka, Y.; Higuchi, K.; Fudetani, R.; Chen, Z.S.; Sumizawa, T.; Akiyama, S.; Kobayashi, M. Reversal of multidrug resistance in human carcinoma cell line by agosterols, marine spongean sterols. Tetrahedron 1999, 55, 13965–13972. [Google Scholar]
- Capon, R.J.; Faulkner, D.J. Herbasterol, an ichthyotoxic 9,11-secosterol from the sponge Dysidea herbacea. J. Org. Chem 1985, 50, 4771–4773. [Google Scholar]
- Braekman, J.C.; Daloze, D.; Moussiaux, B.; Vandevyver, G.; Riccio, G. Cholest-6-en-11β,19-epoxy-3β, 5α, 8α, 9α-tetrol, a novel polyhydroxylated steroid from the sponge Dysidea tupha. Bull. Soc. Chim. Belg 1988, 97, 293–296. [Google Scholar]
- Reddy, M.V.R.; Harper, M.K.; Faulkner, D.J. Luffasterols A-C, 9,11-Secosterols from the Palauan sponge Luffariella sp. J. Nat. Prod 1997, 60, 41–43. [Google Scholar]
- Adinolfi, R.; Migliuolo, A.; Piccialli, V.; Sica, D. Isolation and synthesis of a new 9,11-Secosterol from the sponge Spongia officinalis. J. Nat. Prod 1994, 57, 1220–1226. [Google Scholar]
- De Rosa, S.; De Giulio, A.; Crispino, A.; Iodice, C.; Tommonaro, G. New 9,11−secosterol from the Tyrrhenian sponge Fasciospongia cavernosa. Nat. Prod. Lett 1999, 13, 15–20. [Google Scholar]
- Pika, J.; Andersen, R.J. Blancasterol, a cytotoxic 9,11-secosteroid isolated from the northeastern pacific marine sponge Pleraplysilla sp. Tetrahedron 1993, 49, 8757–8760. [Google Scholar]
- Li, H.; Matsunaga, S.; Fusetani, N. Bioactive marine metabolites. 62. A new 9,11-secosterol, stellettasterol from a marine sponge Stelletta sp. Experientia 1994, 50, 771–773. [Google Scholar]
- Dopeso, J.; Quinoa, E.; Riguere, R.; Debitus, C.; Bergquist, P.R. Euryspongiols: ten new highly hydroxylated 9,11-secosteroids with antihistaminic activity from the sponge euryspongia sp. Stereochemistry and reduction. Tetrahedron 1994, 50, 3813–3828. [Google Scholar]
- Mishra, P.D.; Wahidulla, S.; D’Souza, L.; Kamat, S.Y. Lipid constituents of marine sponge Suberites carnosus. Indian J. Chem. Sect. B 1996, 35, 806–809. [Google Scholar]
- Kong, F.; Andersen, R.J. Polymastiamide A, a novel steroid/amino acid conjugate isolated from the Norwegian marine sponge Polymastia boletiformis (Lamarck, 1815). J. Org. Chem. 1993, 58, 6924–6927. [Google Scholar]
- Kong, F.; Andersen, R.J. Polymastiamides B-F, Novel steroid/amino acid conjugates isolated from the Norwegian marine sponge Polymastia boletiformis. J. Nat. Prod 1996, 59, 379–385. [Google Scholar]
- Umeyama, A.; Shoji, N.; Enoki, M.; Arihara, S. Swinhosterols A-C, 4-methylene secosteroids from the marine sponge Theonella swinhoei. J. Nat. Prod 1997, 60, 296–298. [Google Scholar]
- Tsukamoto, S.; Matsunaga, S.; Fusetani, N.; Van Soest, R.W.M. Acanthosterol sulfates A-J: ten new antifungal steroidal sulfates from a marine sponge Acanthodendrilla sp. J. Nat. Prod 1998, 61, 1374–1378. [Google Scholar]
- Fusetani, N.; Matsunaga, S.; Konosu, S. Bioactive marine metabolites II. Halistanol sulfate, an antimicrobial novel steroid sulfate from the marine sponge Halichondria cf. moorei Bergquist. Tetrahedron Lett 1981, 22, 1985–1988. [Google Scholar]
- Patil, A.D.; Freyer, A.J.; Breen, A.; Carte, B.; Johnson, R.K. Halistanol disulfate B, a novel sulfated sterol from the sponge Pachastrella sp.: Inhibitor of endothelin converting enzyme. J. Nat. Prod 1996, 59, 606–608. [Google Scholar]
- Sun, H.H.; Cross, S.S.; Gunasekera, M.; Kohen, F.E. Weinbersterol disulfates A and B, antiviral steroid sulfates from the sponge Petrosia weinbergi. Tetrahedron 1991, 47, 1185–1190. [Google Scholar]
- Koehn, F.E.; Gunasekera, M.; Cross, S.S. New antiviral sterol disulfate ortho esters from the marine sponge Petrosia weinbergi. J. Org. Chem 1991, 56, 1322–1325. [Google Scholar]
- Rudi, A.; Yosief, T.; Loya, S.; Hizi, A.; Schleyer, M.; Kashman, Y. Clathsterol, a novel anti-HIV-1 RT sulfated sterol from the sponge Clathria species. J. Nat. Prod 2001, 64, 1451–1453. [Google Scholar]
- Li, H.; Matsunaga, S.; Fusetani, N.; Fujiki, H.; Murphy, P.T.; Willis, R.H.; Baker, J.T. Echinoclasterol sulfate phenethylammonium salt, a unique steroid sulfate from the marine sponge, Echinoclathria subhispida. Tetrahedron Lett 1993, 34, 5733–5736. [Google Scholar]
- Kerr, R.G.; Baker, B.J. Marine sterols. Nat. Prod. Rep 1991, 8, 465–498. [Google Scholar]
- Bifulco, G.; Bruno, I.; Minale, L.; Riccio, R. Novel HIV-inhibitory halistanol sulfates F-H from a marine sponge, Pseudoaxinissa digitata. J. Nat. Prod 1994, 57, 164–167. [Google Scholar]
- Gunasekera, S.P.; Sennett, S.H.; Kelly-Borges, M.; Bryant, R.W. Ophirapstanol trisulfate, a new biologically active steroid sulfate from the deep water marine sponge Topsentia ophiraphidites. J. Nat. Prod 1994, 57, 1751–1754. [Google Scholar]
- Makar’eva, T.N.; Shubina, L.K.; Kalinovsky, A.I.; Stonik, V.A.; Elyakov, G.B. Steroids in porifera. II. Steroid derivatives from two sponges of the family Halichondriidae: Sokotrasterol sulfate, a marine steroid with a new pattern of side chain alkylation. Steroids 1983, 42, 267–281. [Google Scholar]
- Makar’eva, T.N.; Dmitrenok, P.S.; Shubina, L.K.; Stonik, V.A. Steroidal compounds of marine sponges. X. 26-norsokotrasterol sulfate, a new steroid from the sponge Trachyopsis halichondroides. Khim. Prir. Soedin 1988, 3, 371–375. [Google Scholar]
- Tsukamoto, S.; Kato, H.; Hirota, H.; Fusetani, N. Isolation of an unusual 2-aminoimidazolium salt of steroid trisulfate from a marine sponge Topsentia sp. Fish. Sci 1997, 63, 310–312. [Google Scholar]
- Shoji, N.; Umeyama, A.; Shin, K.; Takeda, K.; Arihara, S.; Kobayashi, J.; Takei, M. Two unique pentacyclic steroids with cis C/D ring junction from Xestospongia bergquistii Fromont, powerful inhibitors of histamine release. J. Org. Chem 1992, 57, 2996–2997. [Google Scholar]
- Kobayashi, J.; Shinonaga, H.; Shigemori, H.; Umeyama, A.; Shoji, N.; Arihara, S. Xestobergsterol C, a new pentacyclic steroid from the Okinawan marine sponge Ircinia sp. and absolute stereochemistry of xestobergsterol A. J. Nat. Prod 1995, 58, 312–318. [Google Scholar]
- Yang, L.; Andersen, R.J. Absolute configuration of the antiinflamatory sponge natural product contignasterol. J. Nat. Prod 2002, 65, 1924–1926. [Google Scholar]
- Sperry, S.; Crews, P. Haliclostanone sulfate and halistanol sulfate from an Indo-Pacific Haliclona sponge. J. Nat. Prod 1997, 60, 29–32. [Google Scholar]
- Fu, X.; Ferreira, M.L.G.; Schmitz, F.J.; Kelly, M. Tamosterone sulfates: A C-14 epimeric pair of polyhydroxylated sterols from a new Oceanapiid sponge genus. J. Org. Chem 1999, 64, 6706–6709. [Google Scholar]
- Qureshi, A.; Faulkner, D.J. Haplosamates A and B: New steroidal sulfamate esters from two haplosclerid sponges. Tetrahedron 1999, 55, 8323–8330. [Google Scholar]
- Aoki, S.; Naka, Y.; Itoh, T.; Furukawa, T.; Rachmat, R.; Akiyama, S.; Kobayashi, M. Lembehsterols A and B, novel sulfated sterols inhibiting thymidine phosphorylase, from the marine sponge Petrosia strongylata. Chem. Pharm. Bull 2002, 50, 827–830. [Google Scholar]
- McKee, T.C.; Cardellina, J.H., II; Tischler, M.; Snader, K.M.; Boyd, M.R. Ibisterol sulfate, a novel HIV-inhibitory sulfated sterol from the deep water sponge Topsentia sp. Tetrahedron Lett. 1993, 34, 389–392. [Google Scholar]
- Lerch, M.L.; Faulkner, D.J. Unusual polyoxygenated sterols from a Philippines sponge Xestospongia sp. Tetrahedron 2001, 57, 4091–4094. [Google Scholar]
- Fusetani, N.; Takahashi, M.; Matsunaga, S. Topsentiasterol sulfates, antimicrobial sterol sulfates possessing novel side chains, from a marine sponge, Topsentia sp. Tetrahedron 1994, 50, 7765–7770. [Google Scholar]
© 2005 by MDPI Reproduction is permitted for noncommercial purposes.
Share and Cite
Sarma, N.S.; Krishna, M.S.R.; Rao, S.R. Sterol Ring System Oxidation Pattern in Marine Sponges. Mar. Drugs 2005, 3, 84-111. https://doi.org/10.3390/md303084
Sarma NS, Krishna MSR, Rao SR. Sterol Ring System Oxidation Pattern in Marine Sponges. Marine Drugs. 2005; 3(3):84-111. https://doi.org/10.3390/md303084
Chicago/Turabian StyleSarma, Nittala S., M. Sri Rama Krishna, and S. Ramakrishna Rao. 2005. "Sterol Ring System Oxidation Pattern in Marine Sponges" Marine Drugs 3, no. 3: 84-111. https://doi.org/10.3390/md303084
APA StyleSarma, N. S., Krishna, M. S. R., & Rao, S. R. (2005). Sterol Ring System Oxidation Pattern in Marine Sponges. Marine Drugs, 3(3), 84-111. https://doi.org/10.3390/md303084