Abstract
Verrucarin A is an inhibitor of protein synthesis. In this study, we examined the inhibitory action of verrucarin A on signal molecules. Verrucarin A partially inhibited the IL-8 production of a PMA-stimulated promyelocytic leukemia cell line (HL-60 cells), and the effect was related to the inhibition of NF-κB activation at non-cytotoxic concentrations. Moreover, the inhibition of mitogen activated protein (MAP) kinase by verrucarin A was especially strong with p38- and JNK-phosphorylation. The findings show a new action of verrucarin A, and it is expected that this action relaxes the signal activation in response to stress.
Introduction
Marine natural products are important sources of biologically active agents [1]. Microorganisms isolated from marine environments have been recognized as a rich source of biologically active metabolites [2–9]. In the search for new anti-cancer materials, we have screened marine-derived fungal products for natural killer activity. One of about 100 culture broths of marine-derived fungi assayed, strongly inhibited natural killer (NK) activation. Namikoshi et al [10] isolated a new macrocyclic trichothecene, 12,13-deoxyrodidin E, and three known compounds (verrucarin A, verrucarin J, and roridin E) from a culture broth of Myrothecium roridum. The three known compounds showed strong cytotoxicity in human and murine leukemia cell lines (HL-60 and L1210, respectively), but 12,13-deoxyroridin E was about 80-fold less cytotoxic than roridin E, the epoxide variant. Many natural bioactive compounds are toxic to mammalian cells but at specific concentrations are expected to have a specific activity.
In this study, we demonstrated that verrucarin A inhibited the activation of p38 and JNK in the cytosol fraction. From this result we anticipate the discovery of a new compound that confers resistance to the excessive stress reaction in mammalian cells.
Materials and Methods
Material
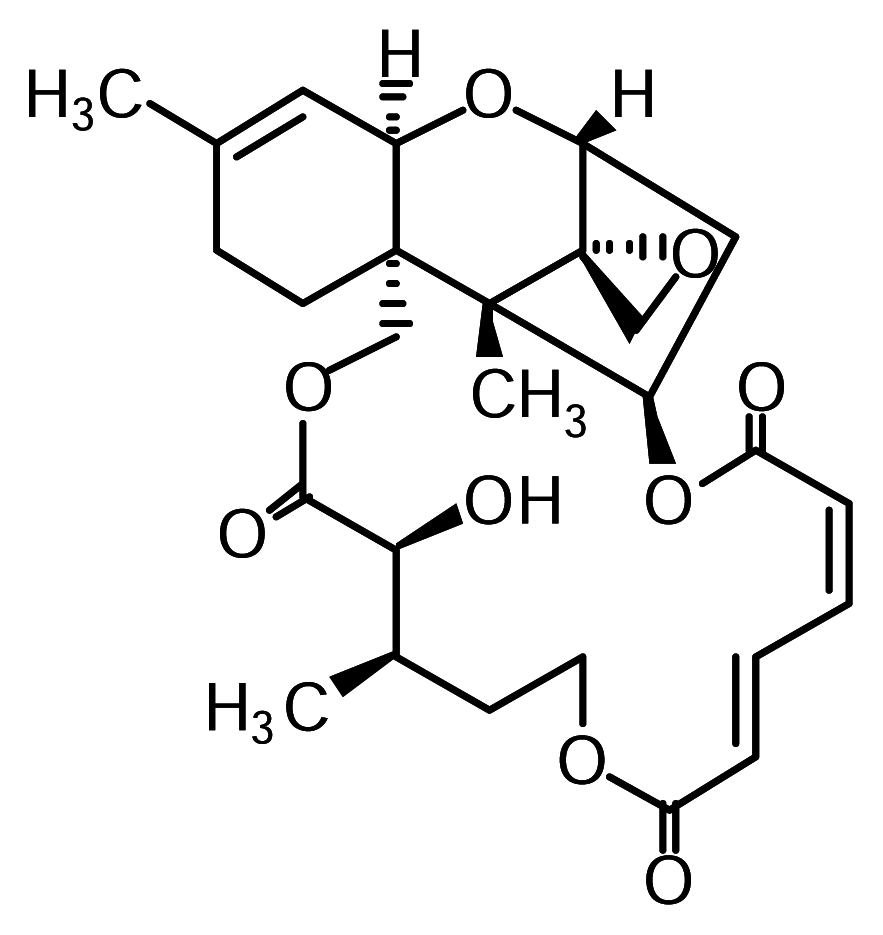
Verrucarin A, verrucarin J and roridin E were prepared as reported [10]. The structure of verrucarin A is shown in Figure 1. 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) and 12-O-tetradecanoyl phorbol 13-acetate (PMA) were purchased from Sigma (St. Louis, MO, USA). Sodium [51Cr]chromate was obtained from Amersham Biosciences K.K. (Tokyo, Japan) Antibodies to IκB-α, phospho-p38, phospho-JNK, phospho-ERK1 and actin were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). All other reagents and chemicals used were of the highest grade available commercially.
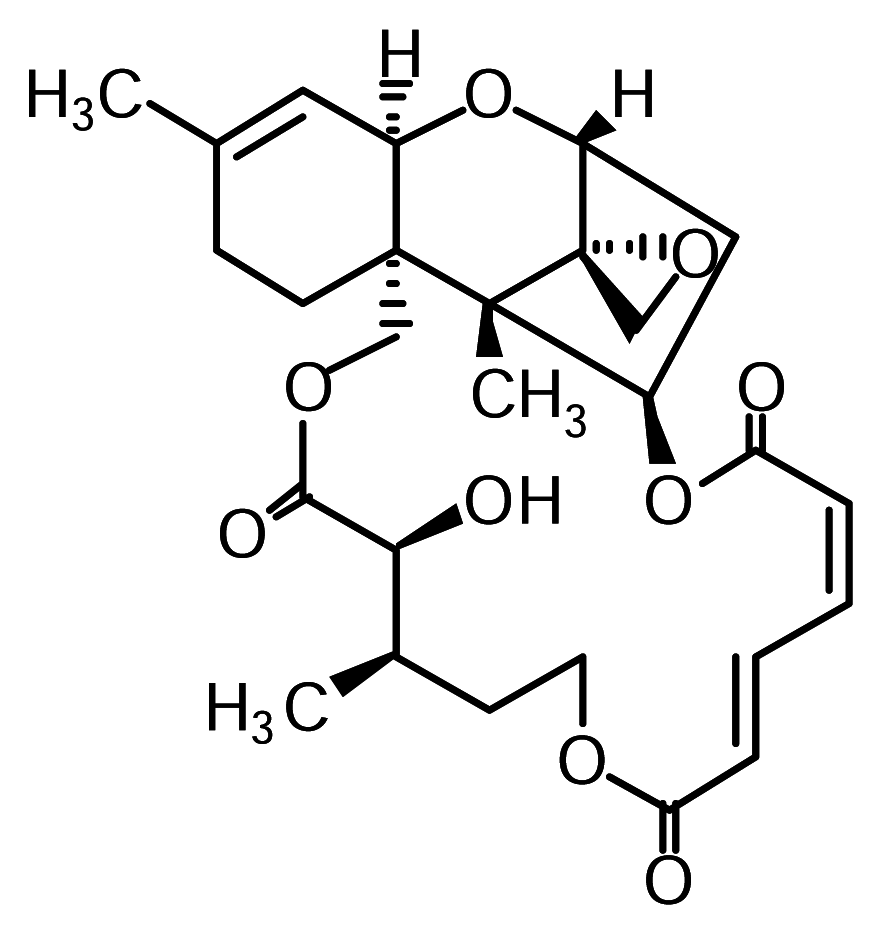
Figure 1.
Structure of verrucarin A.
Cell lines and culture conditions
The human promyelocytic cell line HL-60 and human erythroleukemia cell line K562 were obtained from the Japanese Cancer Research Resources Bank (JCRB, Kamiyoga, Tokyo, Japan). These cell lines were maintained in tissue culture dishes in RPMI 1640 medium (Nissui Seiyaku, Tokyo, Japan), supplemented with 10% heat-inactivated fetal calf serum (FCS), 2 mM glutamine, 100 U/ml of penicillin G and 100 μg/ml of streptomycin.
NK assay
The NK-susceptible target K562 tumor cells were labeled with 51Cr. Peripheral blood leukocytes (PBLs) were prepared from human blood. The PBLs (2 x 105 cells/100 μl medium) were plated, and IL-2 (to a final concentration of 10 ng/ml) was added. Then, the test compound was dissolved in DMSO and added to the PBL suspension (1 μl) at a final volume of 1 μl, and the cells were cultured overnight. 51Cr-labeled K562 cells were added to each well and incubated at 37°C for 4.5 hr. The plate was centrifuged, and 100 μl of supernatant was used for the analyses in a gamma counter [11].
The cytotoxicity was calculated as follows.
Detection of human IL-8 by ELISA
The interleukin 8 (IL-8) concentration of the culture supernatants under control and various test conditions was measured by ELISA using a combination of monoclonal and biotinylated monoclonal antibodies [12]. All samples were assayed at least in duplicate. Data are presented as the mean ± SE of three independent experiments.
Determination of cytotoxicity
Lethal cell injury was assessed by measuring lactate dehydrogenase (LDH) release [13], while proliferation was evaluated by enumerating viable cells using the MTT formazan production method [14]. HL-60 cells (1 x 106 cells/ml) were treated with PMA (with or without verrucarin A) and then transferred to 96-well microtiter plates. After a 24 h incubation, the LDH activity in the supernatants was determined, and 20 μl of MTT reagent (5 mg/ml in PBS) was then added to each well. After incubation for 3 h, formazan production was assessed by measuring the optical density (OD570 nm).
Western blot analysis
HL-60 cells were treated with verrucarin A and lysed in a buffer. The amount of protein in each sample was determined using the Bradford assay. Equal amounts of lysates were subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). The proteins were transferred onto nitrocellulose filters. After blocking with 5% non-fat milk in tris-buffered saline at room temperature for 1 h, the membranes were probed with antibodies to pp38, pJNK, and pERK1. They were then incubated for 30 min with anti-rabbit Ig (Amersham Biosciences K.K., Tokyo, Japan) and for another 30 min with peroxidase-streptavidin conjugate. Blots were developed using an enhanced chemiluminescence detection system (Amersham Biosciences K.K., Tokyo, Japan).
Plasmid construction, cell transfection and luciferase assay
Plasmid construction, cell transfection and the luciferase assay were carried out as described previously [15]. Luciferase reporter plasmids containing the luciferase gene under the control of promoters containing the recognition sites for nuclear factor (NF)-κB were constructed. Oligonucleotides corresponding to the recognition site for each of these factors were synthesized using a DNA synthesizer (RIKAKEN CO., LTD., Tokyo, Japan). The sequences were: 5′-AAGGGACTTTCCGCTGGGGATTCCAG-3′ (NF-κB, four copies). This resulted in a reporter plasmid in which the expression of the luciferase gene was under the control of the relevant transcription factors.
For cell transfections and luciferase assays, a modification of a previously described method [16] was used. Transfection was performed by the electroporation method. HL-60 cells (2.5 x 107) were washed with RPMI 1640 medium and resuspended in 700 μl of the medium containing 20 μg of reporter plasmid DNA. Electroporation was performed at 960 mF and 340 V with a Gene Pulser and a Gene Pulser Cuvette with a 0.4 cm electrode gap (Bio-Rad, Hercules, CA, USA). The cells were suspended in 40 ml of RPMI 1640 medium containing 10% FBS, divided between six 6-cm dishes and incubated for 24 h before being treated with the stimulants and/or test compounds. Cell extracts were prepared 8 h after treatment with PMA, and their luciferase activity was measured using a Pica Gene detection kit (Tokyo Inc. Tokyo, Japan) and a MiniLumat LB 9506 (Berthold Japan K.K., Tokyo, Japan).
Results
The effects of marine-derived fungal products on human NK activity
In order to identify new anti-cancer compounds, an NK assay that uses the T cell fraction of NK cells isolated from human blood, was performed with about 100 culture broths of marine-derived fungi. K562 as a target cell was labeled with 51Cr, and [51Cr] K562 and T cells (contained in NK cells) were mixed at a fixed ratio in the presence of IL-2; the radioactivity in the supernatant correlated with NK activity. As the control, we determined the radioactivity when DMSO was added to a mixture of T and [51Cr]K562 cells. The other samples were used for the change of DMSO. The results for about 100 samples are summarized in Table 1. They showed that no fungal broth strengthened NK activation, as expected, but that strong inhibition was shown by some broths. Notably, No. 42 was of interest, because no inhibition was obtained when IL-2 was not added, but a strong inhibition was detected when IL-2 was present (Table 1). Many natural bioactive compounds are toxic to mammalian cells, but specific concentrations are expected to have a specific action.

Table 1.
Effect of filamentous fungal broths on NK activity by human peripheral blood lymphocytes.a
The effects of roridin E, verrucarin A and verrucarin J on IL-8 production by PMA-stimulated HL-60 cells
Namikoshi et al. isolated a new macrocyclic trichothecene, 12,13-deoxyrodidin E, and three known compounds (verrucarin A, verrucarin J, and roridin E) from a culture broth (No.42) of Myrothecium roridum (2002). The three known compounds showed strong cytotoxicity in human and murine leukemia cell lines (HL-60 and L1210, respectively), but 12,13-deoxyroridin E was about 8 fold less cytotoxic than roridin E, the epoxide variant.
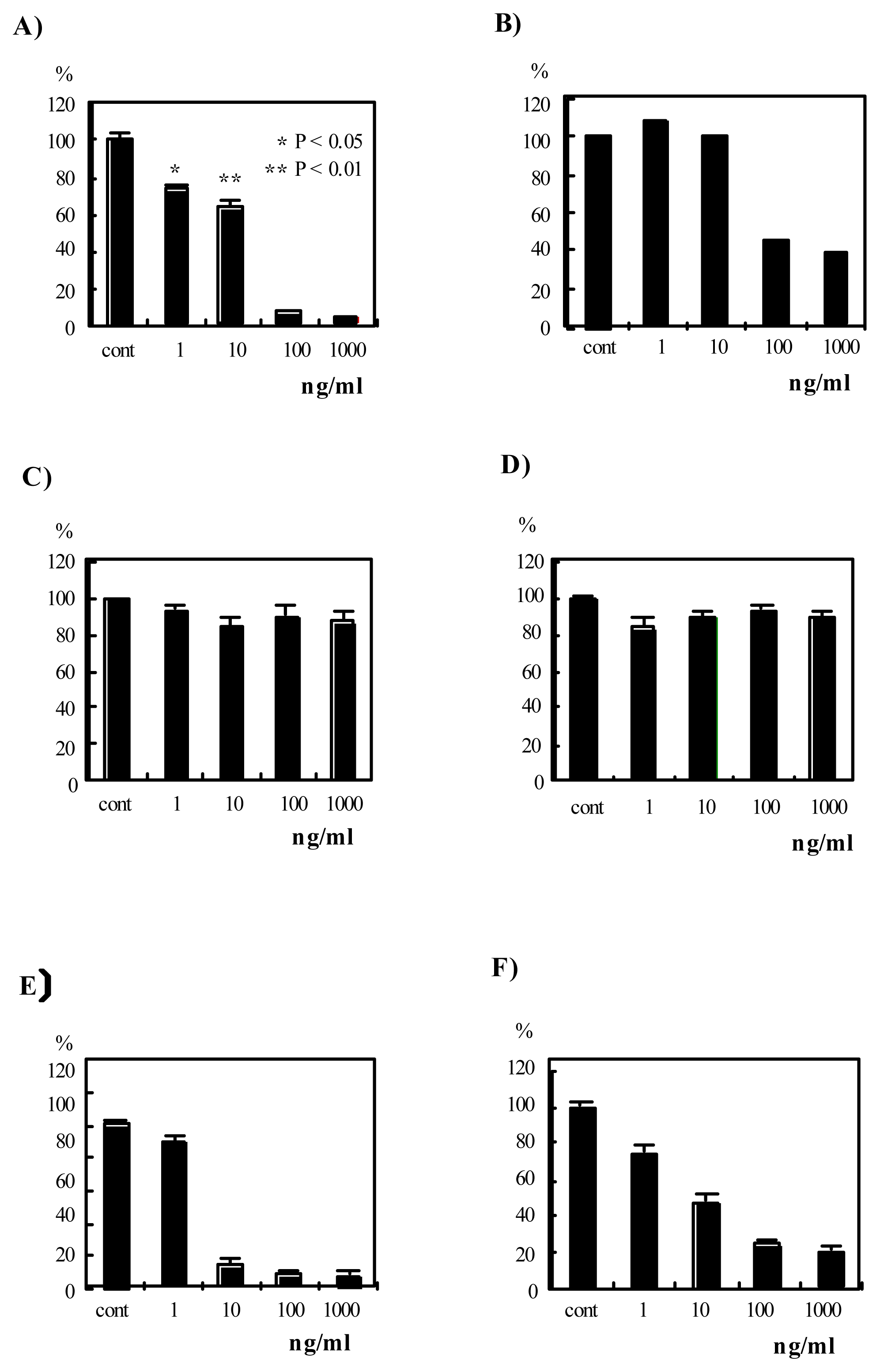
Therefore, the effects of verrucarin A, verrucarin J and roridin E on IL-8 production by PMA-stimulated HL-60 cells were examined, and the results are shown in Figure 2A, C and E. Verrucarin A and roridin E inhibited IL-8 production by PMA-stimulated HL-60 cells, but the inhibition by roridin E and verrucarin A (0.1 and 1 μg/ml) correlated with cell proliferation in the MTT assay. Therefore, it is estimated that the inhibition by roridin E and verrucarin A (0.1 and 1 μg/ml) is due to their cytotoxicity. On the other hand, verrucarin A (1 and 10 ng/ml) did not affect cell proliferation. Therefore, verrucarin A was of interest.
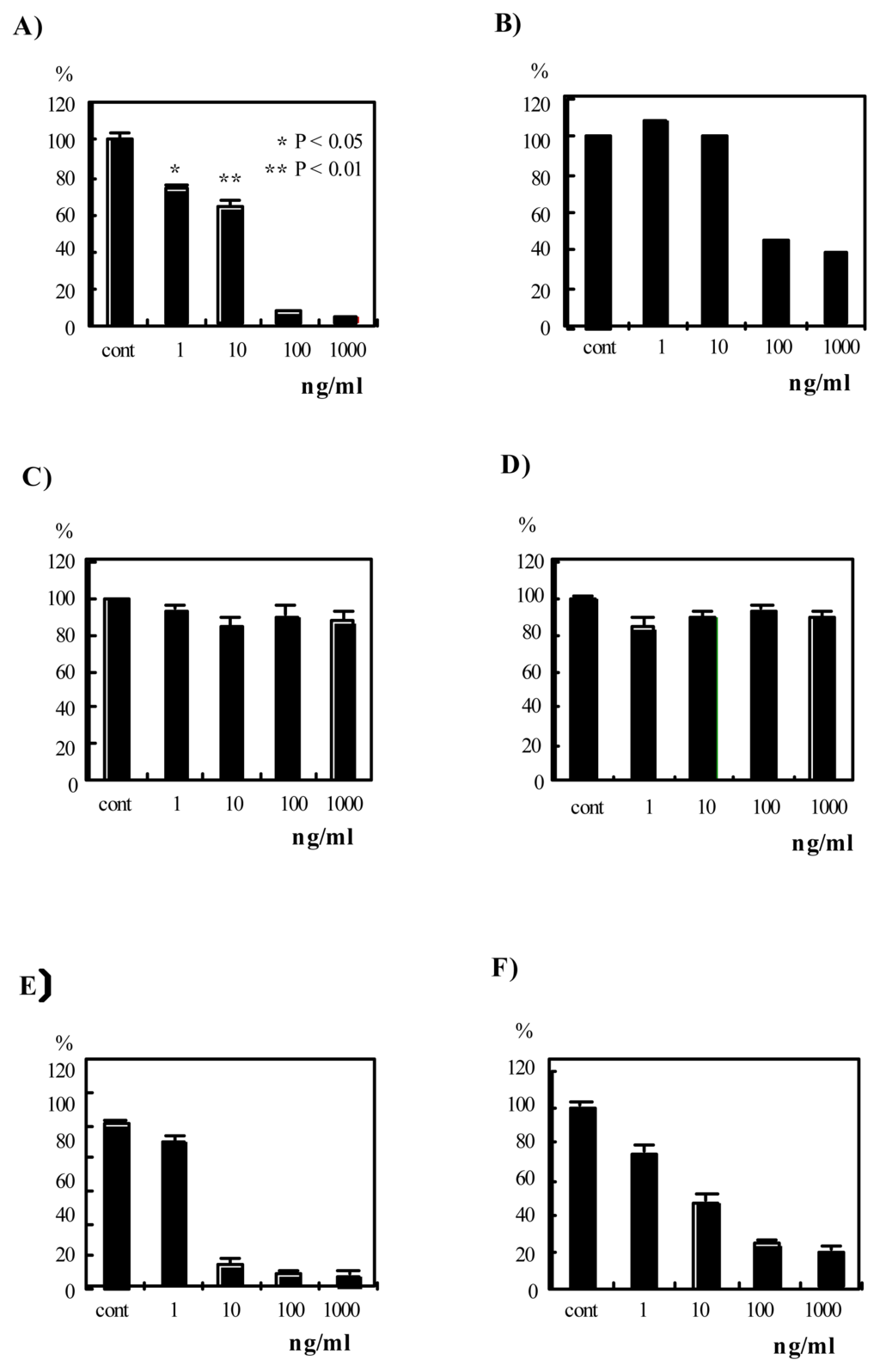
Figure 2.
Effects of verrucarin A, verrucarin J and roridin E on IL-8 production and cell proliferation in PMA-stimulated HL-60 cells. A), C) and E) show effects of verrucarin A, verrucarin J and roridin E, respectively, on IL-8 production. HL-60 cells (1 x 106 cells/ml) were treated with PMA (20 nM) and the indicated concentrations of the compounds for 24 h. The IL-8 concentration in the culture supernatant of the PMA-stimulated cells was determined to be ca. 24 ng/ml by ELISA as described in Materials and Methods. The data are shown as values relative (%) to the PMA-stimulated IL-8 production (24 ng/ml). The data are the mean values of three independent experiments. B), D) and F) show the effects of verrucarin A, verrucarin J and roridin E, respectively, on cell proliferation. The MTT assay was as described in Materials and Methods. The data are shown as values relative (%) to the value of PMA-stimulated condition. The data are the mean values of three independent experiments.
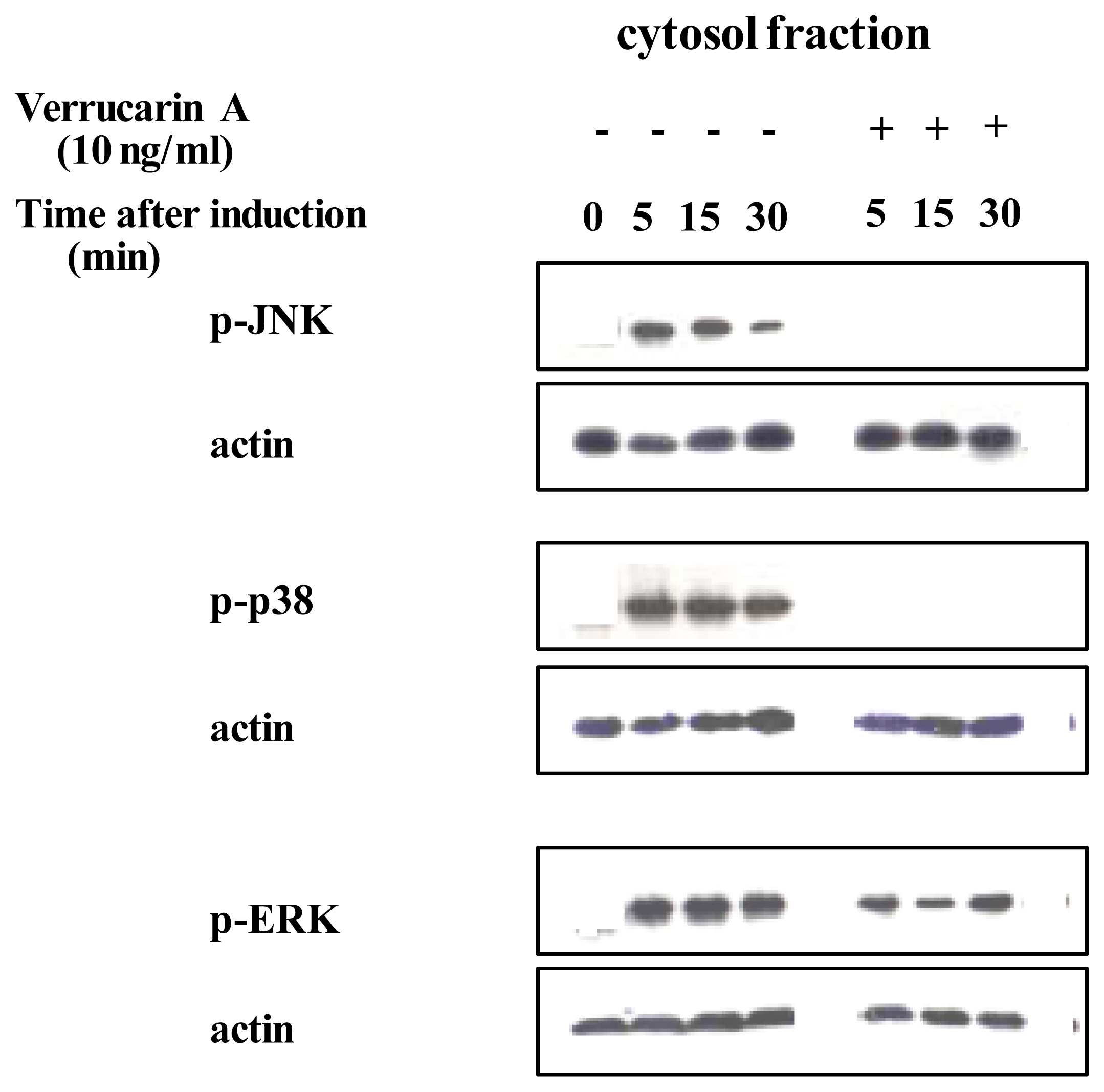
Protein in the cytosol protein preparations treated with or without verrucarin A was analyzed by western blotting.
Inhibitory mechanism of verrucarin A for PMA-stimulated IL-8 production
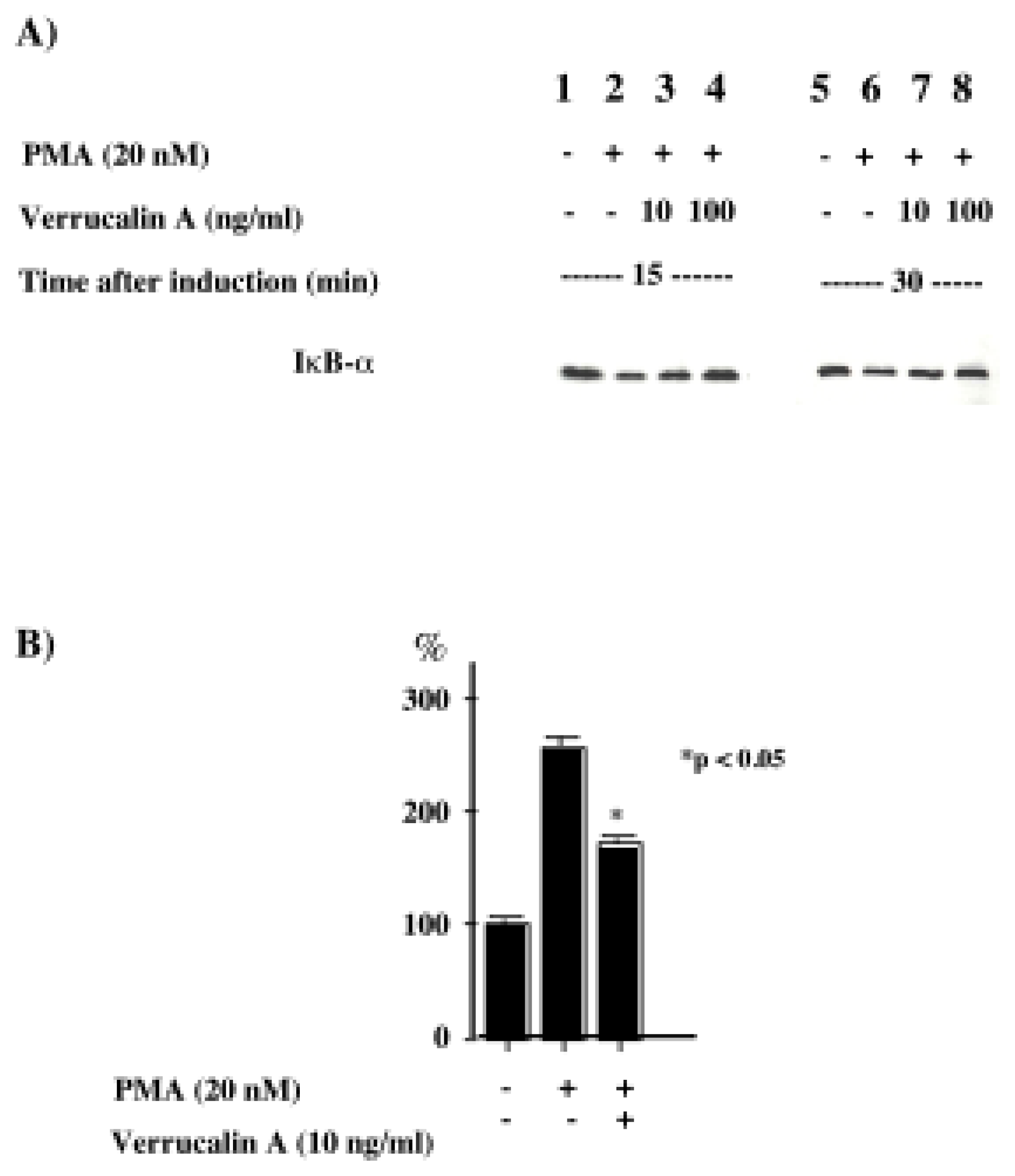
Recently, it was revealed that NF-κB and AP-1 are important for the production of IL-8 [17]. We first investigated whether verrucarin A affects the activation of NF-κB. Inactive NF-κB complexes are associated with inhibitory proteins of the IκB family, which sequester NF-κB in the cytoplasm. It is well known that in response to various stimuli, IκB proteins are first phosphorylated, then ubiquitinated and finally, rapidly degraded by the proteasome, allowing for the nuclear translocation of NF-κB and transcriptional initiation of NF-κB-dependent genes. The IκB family includes the widely distributed IκB-α and IκB-β proteins, together with IκB-γ, IκB-ɛ and Bcl-3. IκB proteins are characterized by a series of ankyrin repeats that interact with the DNA binding domain and nuclear localization signal of NF-κB. Therefore, the effect of verrucarin A on the activation of NF-κB was determined by western blotting with anti IκB-α The results, shown in Figure 3A, indicate that verrucarin A slightly inhibited the degradation of IκB-α. We also investigated the effect of verrucarin A on the transcriptional activity of the IL-8 promoter region using NF-κB binding of the promoter region. The results are shown in Figure 3B. The findings indicated that verrucarin A partially inhibited IL-8 production at the transcriptional level in PMA-stimulated HL-60 cells. Next, the effects of verrucarin A on the activation of MAP kinase were examined. We prepared cytosol fractions for each experimental conditions, and analyzed them by Western blotting with antibodies for phosphorylated p38, JNK and ERK. The results are shown in Figure 4. In the cytosol fraction, the activation of JNK and p38 was inhibited more than the activation of ERK-1.
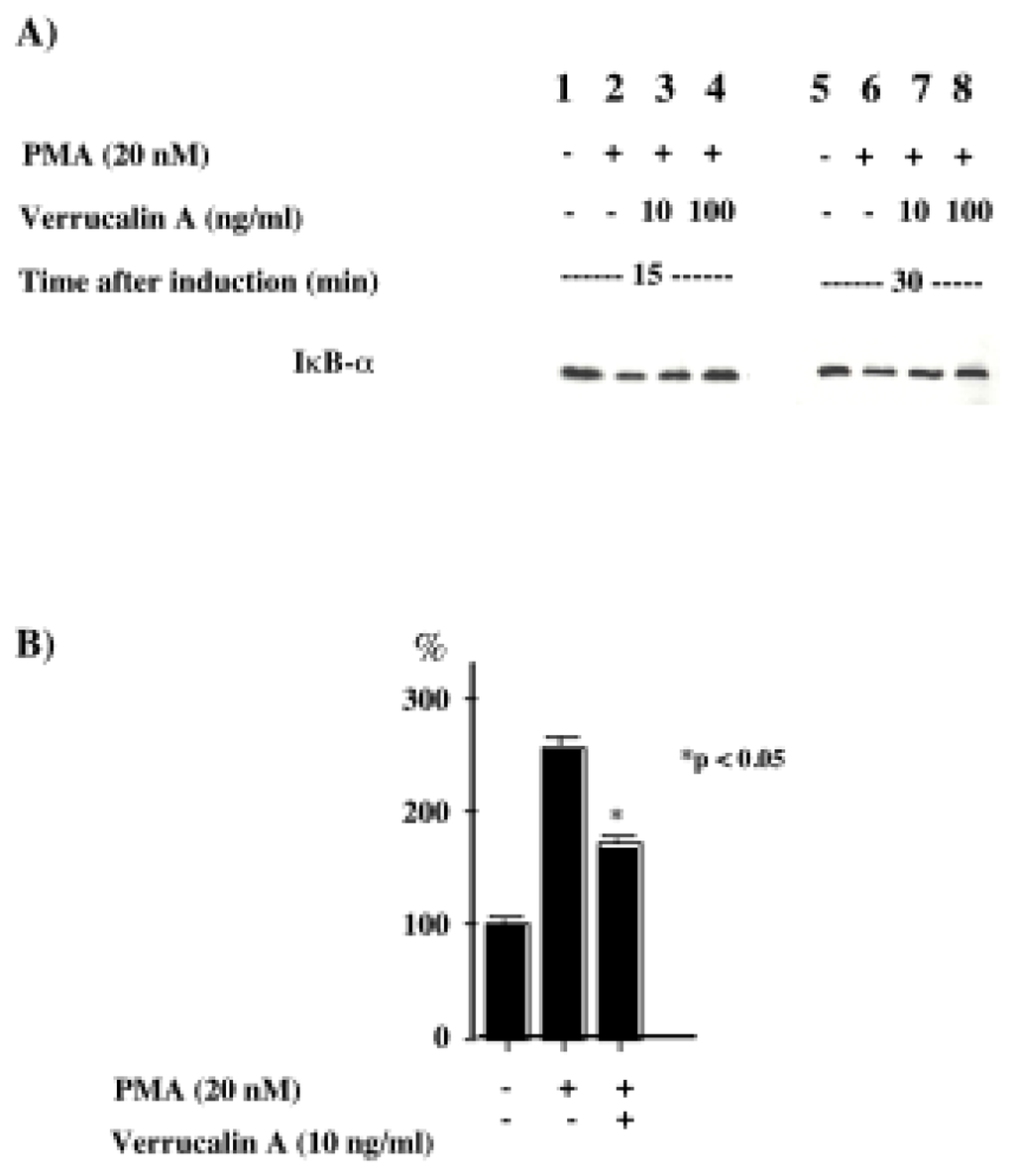
Figure 3.
Effect of verrucarin A on the activation of NF-κB in PMA-stimulated HL-60 cells. A) Western blot analysis with polyclonal antiserum to IκB-α. Total protein extracted from the control (1 and 5) and PMA-stimulated HL-60 cells, with or without 15 or 30 min incubation with verrucarin A (2–4 and 6–8), was electrophoresed on 10% SDS-PAGE gels and examined by western blotting using polyclonal antiserum. B) Effect of verrucarin A on the transcriptional activation of the NF-κB binding region in PMA-stimulated HL-60 cells. HL-60 cells (2.5 x 107) were transfected with the reporter plasmid including the NF-κB binding region and treated with PMA (20 nM) and verrucarin A. The luciferase activities were determined as described in Materials and Methods. The activity of the cells treated with PMA (20 nM) was set at 100%. Each value is the average of the results from three independent experiments.
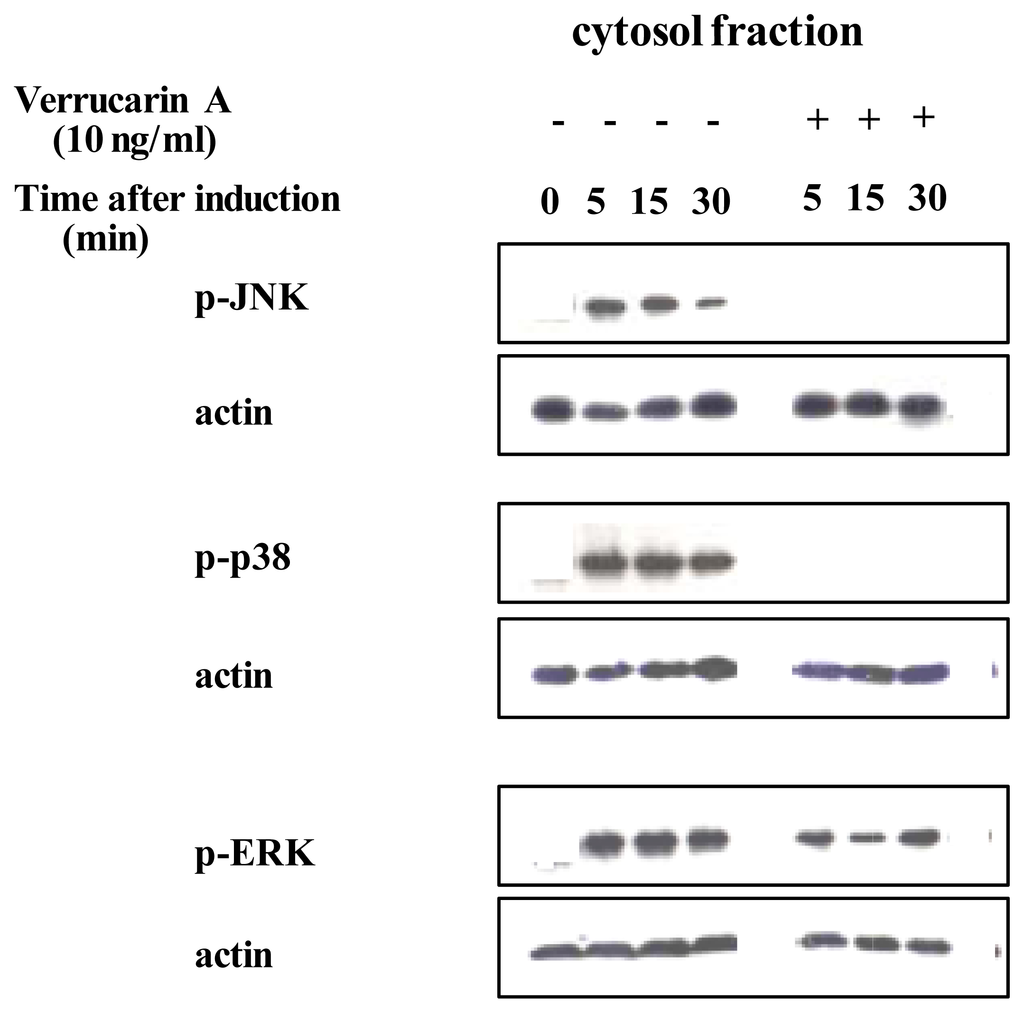
Figure 4.
Effect of verrucarin A on the activation of the MAP kinase in PMA-stimulated HL-60 cells as determined by western blotting using phosphorylated anti-p38, -JNK and -ERK antibodies.
Discussion
We identified verrucarin A by a natural killer (NK) cell assay in marine-derived fungal products. Recently, verrucarin A was reported to be an inhibitor of protein synthesis, and to affect inflammatory cytokine production [18,19], but its target molecule for the inhibition of inflammation was not clear. Therefore, we examined the effects of verrucarin A on IL-8 production in PMA-stimulated HL-60 cells. The results indicated a weak inhibition of IL-8 production at concentrations that were not cytotoxic. Therefore, the target for inhibition was JNK and p38 MAP kinases. It was supposed that the fraction No.42 (increased verrucarin A) inhibited IL-2 dependent MAP kinase activation in NK cells. Although, p42 MAP kinase inhibitor (PD 09805) and p38 MAP kinase inhibitor (SB 203580) have recently been reported, varrucarin A inhibited both p38 and JNK MAP kinases. Generally, the activation of JNK and p38 MAP kinase is promoted by stress, and thus verrucarin A is expected to inhibit the activation of MAP kinase (JNK and p38).
- Sample availability: Not available.
References
- Faulkner, D.J. Marine natural products. Nat. Prod. Rep 2002, 19, 1–48. [Google Scholar]
- Jensen, P.R.; Fenical, W. Secondary metabolites from marine fungi. In Fungi In Marine Environments; Hyde, K.D., Ed.; Fungal Diversity Research Series; Volume 7, Fungal Diversity Press: Hong Kong, 2002; pp. 293–315. [Google Scholar]
- Liberra, K.; Lindequist, U. Marine fungi – a prolific resource of biologically active natural products? Pharmazie 1995, 50, 583–588. [Google Scholar]
- Kelecom, A. Secondary metabolites from marine microorganisms. Annals Brazil. Acad. Sci 2002, 74, 151–170. [Google Scholar]
- Proksch, P.; Edrada, R.A.; Ebel, R. Drugs from the seas - current status and microbiological implications. Appl. Microbiol. Biotechnol 2002, 59, 125–134. [Google Scholar]
- Jensen, P.R.; Fenical, W. In Drugs from the Sea; Fusetani, N., Ed.; Karger: Basel, 2000; pp. 6–29. [Google Scholar]
- Faulkner, D.J.; Harper, M.K.; Haygood, M.G.; Salomon, C.E.; Schmidt, E.W. Symbiotic bacteria in sponges: sources of bioactive substances. In Drugs from the Sea; Fusetani, N., Ed.; Karger: Basel, 2000; pp. 107–119. [Google Scholar]
- Pietra, F. Secondary metabolites from marine microorganisms: bacteria, protozoa, algae and fungi. Achievements and prospects. Nat. Prod. Rep 1997, 14, 453–464. [Google Scholar]
- Davidson, B.S. New dimensions in natural products research: cultured marine microorganisms. Curr. Opin. Biotechnol 1995, 6, 284–291. [Google Scholar]
- Namikoshi, M.; Akano, K.; Kasuga, I.; Mine, Y.; Takahashi, T.; Kobayashi, H. A new macrocyclic trichothecene, 12,13-deoxyroridin E, produced by the marine-derived fungus Myrothecium roridum collected in Palau. J. Nat. Prod 2001, 64, 396–398. [Google Scholar]
- Domzig, W.; Stadler, B.M.; Herberman, R.B. Interleukin 2 dependence of human natural killer (NK) cell activity. J. Immunol 1983, 130, 1970–1973. [Google Scholar]
- Kasahara, T.; Oda, T.; Hatake, K.; Akiyama, M.; Mukaida, N.; Matsushima, K. Interleukin-8 and monocyte chemotactic protein-1 production by a human glioblastoma cell line, T98G in coculture with monocytes: involvement of monocyte-derived interleukin-1alpha. Eur. Cytokine Netw 1998, 9, 47–55. [Google Scholar]
- Shimizu, S.; Nomoto, M.; Naito, S.; Yamamoto, T.; Momose, K. Stimulation of nitric oxide synthase during oxidative endothelial cell injury. Biochem. Pharmacol 1998, 55, 77–83. [Google Scholar]
- Carmichael, J.; DeGraff, W.G.; Gazdar, A.F.; Minna, J.D.; Mitchell, J.B. Evaluation of a tetrazolium-based semiautomated colorimetric assay: assessment of chemosensitivity testing. Cancer Res 1987, 47, 939–942. [Google Scholar]
- Hiramoto, M.; Shimizu, N.; Sugimoto, K.; Tang, J.; Kawakami, Y.; Ito, M.; Aizawa, S.; Tanaka, H.; Makino, I.; Handa, H. Nuclear targeted suppression of NF-kappa B activity by the novel quinone derivative E3330. J. Immunol 1998, 160, 810–819. [Google Scholar]
- Oda, T.; So, Y.; Sato, Y.; Shimizu, N.; Hand, H.; Yasukochi, Y.; Kasahara, T. Inhibition by (±)-indenestrol A of interferon gamma-stimulated nitric oxide formation in murine macrophge RAW 264.7 cells. Mut. Res 2003, 534, 187–195. [Google Scholar]
- Mukaida, N.; Morita, M.; Ishikawa, Y.; Rice, N.; Okamoto, S.; Kasahara, T.; Matsushima, K. Effects of satratoxins and other macrocyclic trichothecenes on IL-2 production and viability of EL-4 thymoma cells. J. Biol. Chem 1994, 269, 13289–13295. [Google Scholar]
- Lee, M.; Li, S.; Jarvis, B.B.; Pestka, J.J. J. Toxicol. Environ. Health A 1999, 57, 459–474.
- Li, G.; Bishop, K.J.; Hall, T.C. J. Biol. Chem. 2001, 276, 2062–2068.
© 2005 by MDPI Reproduction is permitted for noncommercial purposes.