
Identification and Evaluation of Antioxidant and Anti-Aging Peptide Fractions from Enzymatically Hydrolyzed Proteins of Spirulina platensis and Chlorella vulgaris

 , ,
, ,
Abstract
1. Introduction
2. Results and Discussion
2.1. Microalgae Growth
2.2. Protein Isolation, Enzymatic Hydrolysis, and Peptide Purification
2.3. Antioxidant Activity of Peptide Fractions
2.4. Anti-Aging Skin Activities
2.5. Identification of Peptide Using High-Resolution Tandem Mass Spectrometry and Molecular Networking
3. Materials and Methods
3.1. Algae Cultivation and Species Selection
3.2. Chemical Analysis
3.3. Bioactive Peptides Production
3.4. Hydrolysis and Peptide Chain Length Measurements
3.5. Peptide Purification
3.6. Antioxidant Features
3.6.1. DPPH Assay
3.6.2. ABTS Assay
3.6.3. FRAP Assay
3.7. Evaluation of Skin Anti-Aging Potential
3.7.1. Elastase Inhibition Assay
3.7.2. Collagenase Inhibition Assay
3.7.3. Tyrosinase Inhibition Assay
3.8. High-Pressure Liquid Chromatography Tandem Mass Spectrometry (HPLC-MS/MS)
3.9. Molecular Networking
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, K.L.; Gao, M.; Wang, Y.Z.; Li, X.R.; Wang, P.; Wang, B. Antioxidant peptides from protein hydrolysate of marine red algae Eucheuma cottonii: Preparation, identification, and cytoprotective mechanisms on H2O2 oxidative damaged HUVECs. Front. Microbiol. 2022, 13, 791248. [Google Scholar] [CrossRef] [PubMed]
- Hayes, M.; Aluko, R.E.; Aurino, E.; Mora, L. Generation of bioactive peptides from Porphyridium sp. and assessment of their potential for use in the prevention of hypertension, inflammation, and pain. Mar. Drugs 2023, 21, 422. [Google Scholar] [CrossRef] [PubMed]
- Zwander, S.; Chaturvedi, P.; Ghatak, A.; Weckwerth, W.; Marko, D.; Castejón, N. Integrating eco-friendly approaches to produce protein extracts and hydrolysates with antioxidant properties from Microchloropsis gaditana. Algal Res. 2024, 77, 103368. [Google Scholar] [CrossRef]
- Jannel, S.; Caro, Y.; Bermudes, M.; Petit, T. Novel insights into the biotechnological production of Haematococcus pluvialis-derived astaxanthin: Advances and key challenges to allow its industrial use as novel food ingredient. J. Mar. Sci. Eng. 2020, 8, 789. [Google Scholar] [CrossRef]
- Cunha, S.A.; Pintado, M.E. Bioactive peptides derived from marine sources: Biological and functional properties. Trends Food Sci. Technol. 2022, 119, 348–370. [Google Scholar] [CrossRef]
- Marthandam Asokan, S.; Wang, T.; Su, W.T.; Lin, W.T. Antidiabetic effects of a short peptide of potato protein hydrolysate in STZ-induced diabetic mice. Nutrients 2019, 11, 779. [Google Scholar] [CrossRef]
- Liu, C.; Chen, G.; Rao, H.; Xiao, X.; Chen, Y.; Wu, C.; Bian, F.; He, H. Novel antioxidant peptides identified from Arthrospira platensis hydrolysates prepared by a marine bacterium Pseudoalteromonas sp. JS4-1 extracellular protease. Mar. Drugs 2023, 21, 133. [Google Scholar] [CrossRef]
- Maqsoudlou, A.; Mahoonak, A.S.; Mora, L.; Mohebodini, H.; Toldrá, F.; Ghorbani, M. Peptide identification in alcalase hydrolysated pollen and comparison of its bioactivity with royal jelly. Food Res. Int. 2019, 116, 905–915. [Google Scholar] [CrossRef]
- Gharehbeglou, P.; Sarabandi, K.; Akbarbaglu, Z. Insights into enzymatic hydrolysis: Exploring effects on antioxidant and functional properties of bioactive peptides from chlorella proteins. J. Agric. Food Res. 2024, 16, 101129. [Google Scholar] [CrossRef]
- Aurino, E.; Mora, L.; Marzocchella, A.; Kuchendorf, C.M.; Ackermann, B.; Hayes, M. Functional and Bioactive Benefits of Selected Microalgal Hydrolysates Assessed In Silico and In Vitro. Mar. Drugs 2025, 23, 53. [Google Scholar] [CrossRef]
- Safi, C.; Zebib, B.; Merah, O.; Pontalier, P.Y.; Vaca-Garcia, C. Morphology, composition, production, processing and applications of Chlorella vulgaris: A review. Renew. Sustain. Energy Rev. 2014, 35, 265–278. [Google Scholar] [CrossRef]
- Manlusoc, J.K.T.; Hsieh, C.L.; Hsieh, C.Y.; Salac, E.S.N.; Lee, Y.T.; Tsai, P.W. Pharmacologic application potentials of sulfated polysaccharide from marine algae. Polymers 2019, 11, 1163. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.Y.; Wang, Y.M.; Zhao, Y.Q.; Chi, C.F.; Wang, B. Cytoprotective effect of antioxidant pentapeptides from the protein hydrolysate of swim bladders of miiuy croaker (Miichthys miiuy) against H2O2-mediated human umbilical vein endothelial cell (HUVEC) injury. Int. J. Mol. Sci. 2019, 20, 5425. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, G.X.; Zhao, Y.Q.; Qiu, Y.T.; Chi, C.F.; Wang, B. Identification and active evaluation of antioxidant peptides from protein hydrolysates of skipjack tuna (Katsuwonus pelamis) head. Antioxidants 2019, 8, 318. [Google Scholar] [CrossRef]
- Sheng, Y.; Qiu, Y.T.; Wang, Y.M.; Chi, C.F.; Wang, B. Novel antioxidant collagen peptides of Siberian sturgeon (Acipenser baerii) cartilages: The preparation, characterization, and cytoprotection of H2O2-damaged human umbilical vein endothelial cells (HUVECs). Mar. Drugs 2022, 20, 325. [Google Scholar] [CrossRef]
- Joshi, S.; Kumari, R.; Upasani, V.N. Applications of algae in cosmetics: An overview. Int. J. Innov. Res. Sci. Eng. Technol. 2018, 7, 1269. [Google Scholar] [CrossRef]
- Madan, K.; Nanda, S. In-vitro evaluation of antioxidant, anti-elastase, anti-collagenase, anti-hyaluronidase activities of safranal and determination of its sun protection factor in skin photoaging. Bioorg. Chem. 2018, 77, 159–167. [Google Scholar] [CrossRef]
- Jiratchayamaethasakul, C.; Ding, Y.; Hwang, O.; Im, S.T.; Jang, Y.; Myung, S.W.; Lee, J.M.; Kim, H.-S.; Ko, S.-C.; Lee, S.H. In vitro screening of elastase, collagenase, hyaluronidase, and tyrosinase inhibitory and antioxidant activities of 22 halophyte plant extracts for novel cosmeceuticals. Fish. Aquat. Sci. 2020, 23, 6. [Google Scholar] [CrossRef]
- Podolskiy, D.I.; Gladyshev, V.N. Intrinsic versus extrinsic cancer risk factors and aging. Trends Mol. Med. 2016, 22, 833–834. [Google Scholar] [CrossRef]
- Ricard-Blum, S.; Salza, R. Matricryptins and matrikines: Biologically active fragments of the extracellular matrix. Exp. Dermatol. 2014, 23, 457–463. [Google Scholar] [CrossRef]
- Pai, V.; Bhandari, P.; Shukla, P. Topical peptides as cosmeceuticals. Indian J. Dermatol. Venereol. Leprol. 2017, 83, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Daim, M.M.; Farouk, S.M.; Madkour, F.F.; Azab, S.S. Anti-inflammatory and immunomodulatory effects of Spirulina platensis in comparison to Dunaliella salina in acetic acid-induced rat experimental colitis. Immunopharmacol. Immunotoxicol. 2015, 37, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Razali, A.N.; Amin, A.M.; Sarbon, N.M. Antioxidant activity and functional properties of fractionated cobia skin gelatin hydrolysate at different molecular weight. Int. Food Res. J. 2015, 22, 651. [Google Scholar]
- Saallah, S.; Ishak, N.H.; Sarbon, N.M. Effect of different molecular weight on the antioxidant activity and physicochemical properties of golden apple snail (Ampullariidae) protein hydrolysates. Food Res. 2020, 4, 1363–1370. [Google Scholar] [CrossRef]
- Montalvo, G.E.B.; Thomaz-Soccol, V.; Vandenberghe, L.P.; Carvalho, J.C.; Faulds, C.B.; Bertrand, E.; Prado, M.R.; Bonatto, S.J.; Soccol, C.R. Arthrospira maxima OF15 biomass cultivation at laboratory and pilot scale from sugarcane vinasse for potential biological new peptides production. Bioresour. Technol. 2019, 273, 103–113. [Google Scholar] [CrossRef]
- Rojo, E.M.; Piedra, I.; González, A.M.; Vega, M.; Bolado, S. Effect of process parameters on the valorization of components from microalgal and microalgal-bacteria biomass by enzymatic hydrolysis. Bioresour. Technol. 2021, 335, 125256. [Google Scholar] [CrossRef]
- Kristinsson, H.G.; Rasco, B.A. Biochemical and functional properties of Atlantic salmon (Salmo salar) muscle proteins hydrolyzed with various alkaline proteases. J. Agric. Food Chem. 2000, 48, 657–666. [Google Scholar] [CrossRef]
- Hamzeh, A.; Rezaei, M.; Khodabandeh, S.; Motamedzadegan, A.; Noruzinia, M. Antiproliferative and antioxidative activities of cuttlefish (Sepia pharaonis) protein hydrolysates as affected by degree of hydrolysis. J. Food Meas. Charact. 2018, 12, 721–727. [Google Scholar] [CrossRef]
- Nasir, S.N.A.M.; Sarbon, N.M. Angiotensin converting enzyme (ACE), antioxidant activity and functional properties of shortfin scad (Decapterus macrosoma) muscle protein hydrolysate at different molecular weight variations. Biocatal. Agric. Biotechnol. 2019, 20, 101254. [Google Scholar] [CrossRef]
- Akaberi, S.; Gusbeth, C.; Silve, A.; Senthilnathan, D.S.; Navarro-López, E.; Molina-Grima, E.; Frey, W. Effect of pulsed electric field treatment on enzymatic hydrolysis of proteins of Scenedesmus almeriensis. Algal Res. 2019, 43, 101656. [Google Scholar] [CrossRef]
- Rashidian, G.; Abedian Kenari, A.; Nikkhah, M. Evaluation of antioxidative and antibacterial activities of fractionated hydrolysate from shrimp Litopenaeus vannamei head wastes against aquatic pathogenic bacteria. Aquac. Res. 2021, 52, 3696–3704. [Google Scholar] [CrossRef]
- Pekkoh, J.; Ruangrit, K.; Pumas, C.; Duangjan, K.; Chaipoot, S.; Phongphisutthinant, R.; Srinuanpan, S. Transforming microalgal Chlorella biomass into cosmetically and nutraceutically protein hydrolysates using high-efficiency enzymatic hydrolysis approach. Biomass Convers. Biorefin. 2023, 13, 6299–6315. [Google Scholar] [CrossRef]
- Lisboa, C.R.; Pereira, A.M.; Costa, J.A.V. Biopeptides with antioxidant activity extracted from the biomass of Spirulina sp. LEB 18. Afr. J. Microbiol. Res. 2016, 10, 79–86. [Google Scholar] [CrossRef]
- Kim, S.-K.; Ravichandran, Y.D.; Khan, S.B.; Kim, Y.T. Prospective of the cosmeceuticals derived from marine organisms. Biochem. Eng. J. 2008, 13, 511–523. [Google Scholar] [CrossRef]
- Carrasco-Castilla, J.; Hernández-Álvarez, A.J.; Jiménez-Martínez, C.; Jacinto-Hernández, C.; Alaiz, M.; Girón-Calle, J.; Dávila-Ortiz, G. Antioxidant and metal chelating activities of peptide fractions from phaseolin and bean protein hydrolysates. Food Chem. 2012, 135, 1789–1795. [Google Scholar] [CrossRef]
- Wu, H.; Liu, Z.; Zhao, Y.; Zeng, M. Enzymatic preparation and characterization of iron-chelating peptides from anchovy (Engraulis japonicus) muscle protein. Food Res. Int. 2012, 48, 435–441. [Google Scholar] [CrossRef]
- Bozkurt, F.; Bekiroglu, H.; Dogan, K.; Karasu, S.; Sagdic, O. Technological and bioactive properties of wheat glutenin hydrolysates prepared with various commercial proteases. LWT 2021, 149, 111787. [Google Scholar] [CrossRef]
- Shahidi, F.; Zhong, Y. Bioactive peptides. J. AOAC Int. 2008, 91, 914–931. [Google Scholar] [CrossRef]
- Nutho, B.; Tungmunnithum, D. Exploring major flavonoid phytochemicals from Nelumbo nucifera Gaertn. as potential skin anti-aging agents: In silico and in vitro evaluations. Int. J. Mol. Sci. 2023, 24, 16571. [Google Scholar] [CrossRef]
- Kunte, M.; Desai, K. The inhibitory effect of c-phycocyanin containing protein extract (c-pc extract) on human matrix metalloproteinases (mmp-2 and mmp-9) in hepatocellular cancer cell line (hepg2). Protein J. 2017, 36, 186–195. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Bandeira, N. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Lintner, P.; Peschard, O. Biologically active peptides: From a laboratory bench curiosity to a functional skin care product. Int. J. Cosmet. Sci. 2000, 22, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Hipkiss, A.R. Glycation, ageing, and carnosine: Are carnivorous diets beneficial? Mech. Ageing Dev. 2005, 126, 1034–1039. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, I.; Setoyama, O.; Jia, Y.; Fujita, N.; Waki, A.; Komiya, Y.; Arihara, K. Effects of the dipeptides comprising leucine and lysine on lifespan and age-related stress in Caenorhabditis elegans. Food Sci. Nutr. 2023, 11, 2776–2786. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, X.; Zhang, S.; Wang, Z.; Tian, S.; Wu, Q. Identification and free radical scavenging activity of oligopeptides from mixed-distillate fermented Baijiu grains and soy sauce residue. Metabolites 2024, 14, 298. [Google Scholar] [CrossRef]
- Aiba, S.; Ogawa, T. Assessment of growth yield of a blue—Green alga, Spirulina platensis, in axenic and continuous culture. Microbiology 1977, 102, 179–182. [Google Scholar] [CrossRef]
- Sorokin, C.; Krauss, R.W. The effects of light intensity on the growth rates of green algae. Plant Physiol. 1958, 33, 109. [Google Scholar] [CrossRef]
- AACC (American Association of Cereal Chemists). Approved Method 56–30, 10th ed.; AACC: St. Paul, MN, USA, 2000. [Google Scholar]
- Kaur, M.; Singh, N. Characterization of protein isolates from different Indian chickpea (Cicer arietinum L.) cultivars. Food Chem. 2007, 102, 366–372. [Google Scholar] [CrossRef]
- Sanchez-Vioque, R.; Climente, A.; Vioque, J.; Bautista, J.; Millan, F. Protein isolate from chickpea (Cicer areitinum L.): Chemical composition, functional properties, and protein characterization. J. Food Chem. 1999, 64, 237–243. [Google Scholar] [CrossRef]
- Wasswa, J.; Tang, J.; Gu, X.H.; Yuan, X.Q. Influence of the extent of enzymatic hydrolysis on the functional properties of protein hydrolysate from grass carp (Ctenopharyngodon idella) skin. Food Chem. 2007, 104, 1698–1704. [Google Scholar] [CrossRef]
- Ovissipour, M.; Safari, R.; Motamedzadegan, A.; Rasco, B.; Pourgholam, R.; Mohagheghi, E.; Molla, A.E. Use of hydrolysates from yellowfin tuna Thunnus albacares fisheries by-product as a nitrogen source for bacteria growth media. Int. Aquat. Res. 2009, 1, 73–77. [Google Scholar]
- Christensen, M.; Andersen, E.; Christensen, L.; Andersen, M.L.; Baron, C.P. Textural and biochemical changes during ripening of salted herrings. J. Sci. Food Agric. 2011, 91, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Chen, J.; Cai, Y.; Lei, Y.; Chen, L.; Pei, L.; Zhou, D.; Liang, X.; Ruan, J. Antioxidant, free radical scavenging, anti-inflammatory and hepatoprotective potential of the extract from Parathelypteris nipponica (Franch. et Sav.) Ching. J. Ethnopharmacol. 2010, 130, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wu, Q.; Wu, Y.; Chen, G.; Yue, W.; Liang, Q. Response surface optimized ultrasonic-assisted extraction of flavonoids from Sparganii rhizoma and evaluation of their in vitro antioxidant activities. Molecules 2012, 17, 6769–6783. [Google Scholar] [CrossRef]
- Haddar, A.; Sellimi, S.; Ghannouchi, R.; Alvarez, O.M.; Nasri, M.; Bougatef, A. Functional, antioxidant, and film-forming properties of tuna-skin gelatin with a brown algae extract. Int. J. Biol. Macromol. 2012, 51, 477–483. [Google Scholar] [CrossRef]
- Honda, M. Z-Isomers of lycopene and β-carotene exhibit greater skin-quality improving action than their all-E-isomers. Food Chem. 2023, 421, 135954. [Google Scholar] [CrossRef]
- Sim, Y.Y.; Nyam, K.L. Application of Hibiscus cannabinus L.(kenaf) leaves extract as skin whitening and anti-aging agents in natural cosmetic prototype. Ind. Crops Prod. 2021, 167, 113491. [Google Scholar] [CrossRef]
- Chang, T.S. An updated review of tyrosinase inhibitors. Int. J. Mol. Sci. 2009, 10, 2440–2475. [Google Scholar] [CrossRef]


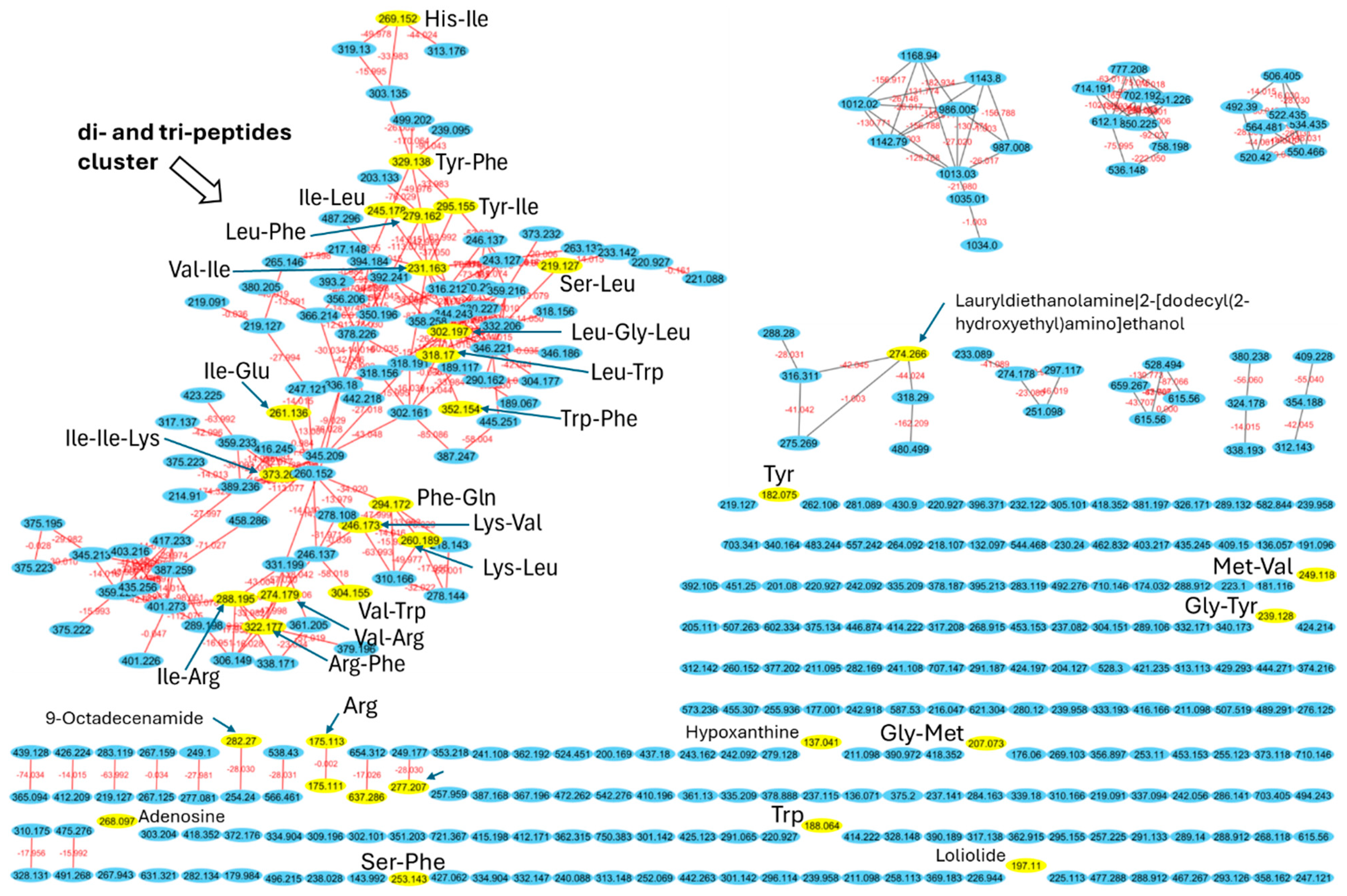
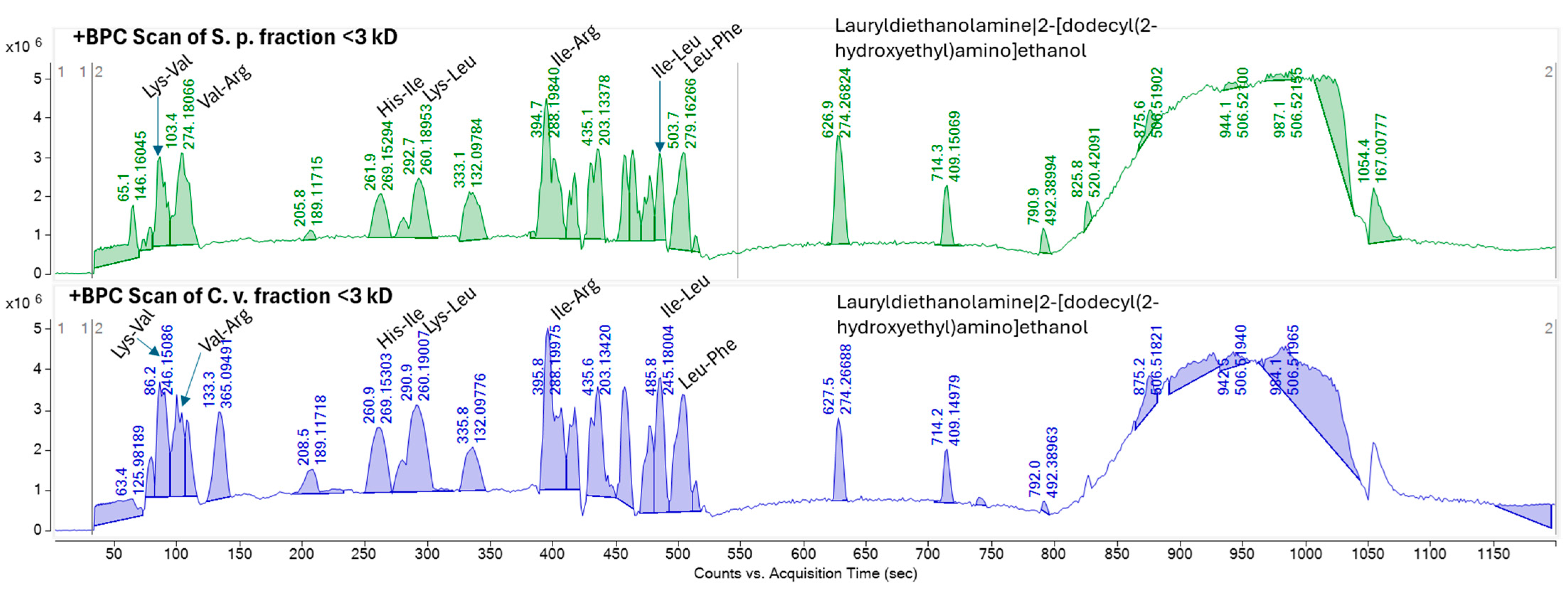


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. platensis | C. vulgaris | |||
|---|---|---|---|---|
| A | Dry weight (g/L) | 1 ± 0.08 a | 0.8 ± 0.12 a | |
| Dry weight (%) | 0.1 ± 0.08 a | 0.08 ± 0.12 a | ||
| Protein (%) | 64 ± 1.45 a | 54 ± 0.88 b | ||
| Lipid (%) | 10 ± 0.88 a | 9 ± 0.90 a | ||
| Moisture (%) | 9 ± 0.72 a | 5 ± 0.60 a | ||
| Ash (%) | 11 ± 0.50 a | 14 ±0.78 a | ||
| B | Lipid after defat (%) | 2.36 ± 0.23 a | 3.26 ± 0.35 a | |
| C | Isolated protein (%) | 82 ± 0.92 a | 72 ± 0.65 b | |
| D | Degree of hydrolysis (%) | 33 ± 0.70 a | 30 ± 1.19 a | |
| Length of the peptide chain | 3.07 ± 0.08 b | 3.33 ± 0.02 a | ||
| E | Peptide fraction (mg/g of hydrolyzed protein) | >10 kDa | 100 ± 7.50 c | 100 ± 3.78 c |
| 3–10 kDa | 200 ± 6.08 b | 150 ± 4.93 b | ||
| <3 kDa | 650 ± 9.82 a | 700 ± 6.17 a | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masoumifeshani, B.; Abedian Kenari, A.; Sottorff, I.; Crüsemann, M.; Amiri Moghaddam, J. Identification and Evaluation of Antioxidant and Anti-Aging Peptide Fractions from Enzymatically Hydrolyzed Proteins of Spirulina platensis and Chlorella vulgaris. Mar. Drugs 2025, 23, 162. https://doi.org/10.3390/md23040162
Masoumifeshani B, Abedian Kenari A, Sottorff I, Crüsemann M, Amiri Moghaddam J. Identification and Evaluation of Antioxidant and Anti-Aging Peptide Fractions from Enzymatically Hydrolyzed Proteins of Spirulina platensis and Chlorella vulgaris. Marine Drugs. 2025; 23(4):162. https://doi.org/10.3390/md23040162
Chicago/Turabian StyleMasoumifeshani, Baran, Abdolmohammad Abedian Kenari, Ignacio Sottorff, Max Crüsemann, and Jamshid Amiri Moghaddam. 2025. "Identification and Evaluation of Antioxidant and Anti-Aging Peptide Fractions from Enzymatically Hydrolyzed Proteins of Spirulina platensis and Chlorella vulgaris" Marine Drugs 23, no. 4: 162. https://doi.org/10.3390/md23040162
APA StyleMasoumifeshani, B., Abedian Kenari, A., Sottorff, I., Crüsemann, M., & Amiri Moghaddam, J. (2025). Identification and Evaluation of Antioxidant and Anti-Aging Peptide Fractions from Enzymatically Hydrolyzed Proteins of Spirulina platensis and Chlorella vulgaris. Marine Drugs, 23(4), 162. https://doi.org/10.3390/md23040162