
Ishophloroglucin A Isolated from Ishige okamurae Protects Glomerular Cells from Methylglyoxal-Induced Diacarbonyl Stress and Inhibits the Pathogenesis of Diabetic Nephropathy


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
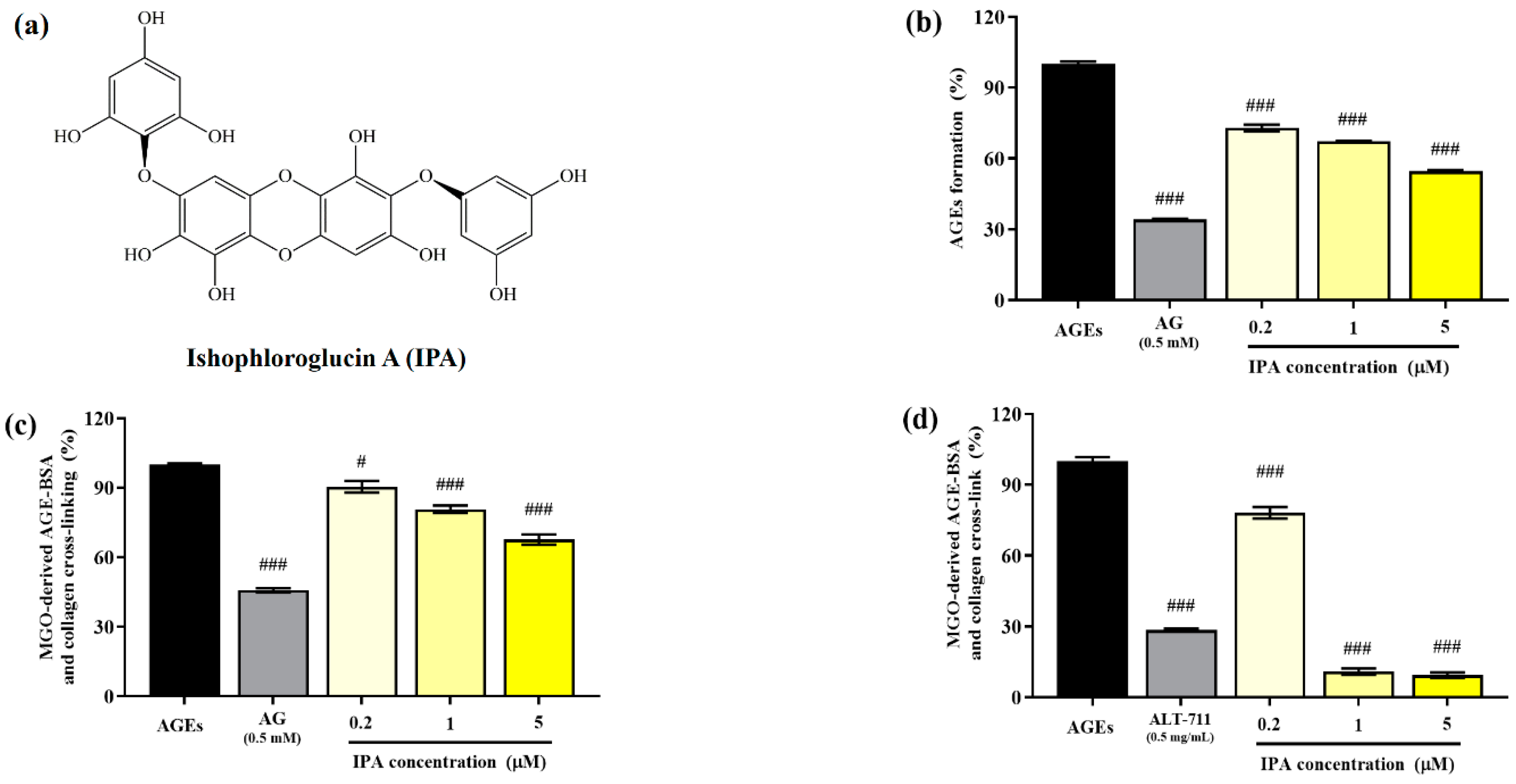
2.1. Anti-Glycation Properties of IPA
2.2. Renoprotective Effects of IPA Against MGO-Induced Oxidative Stress
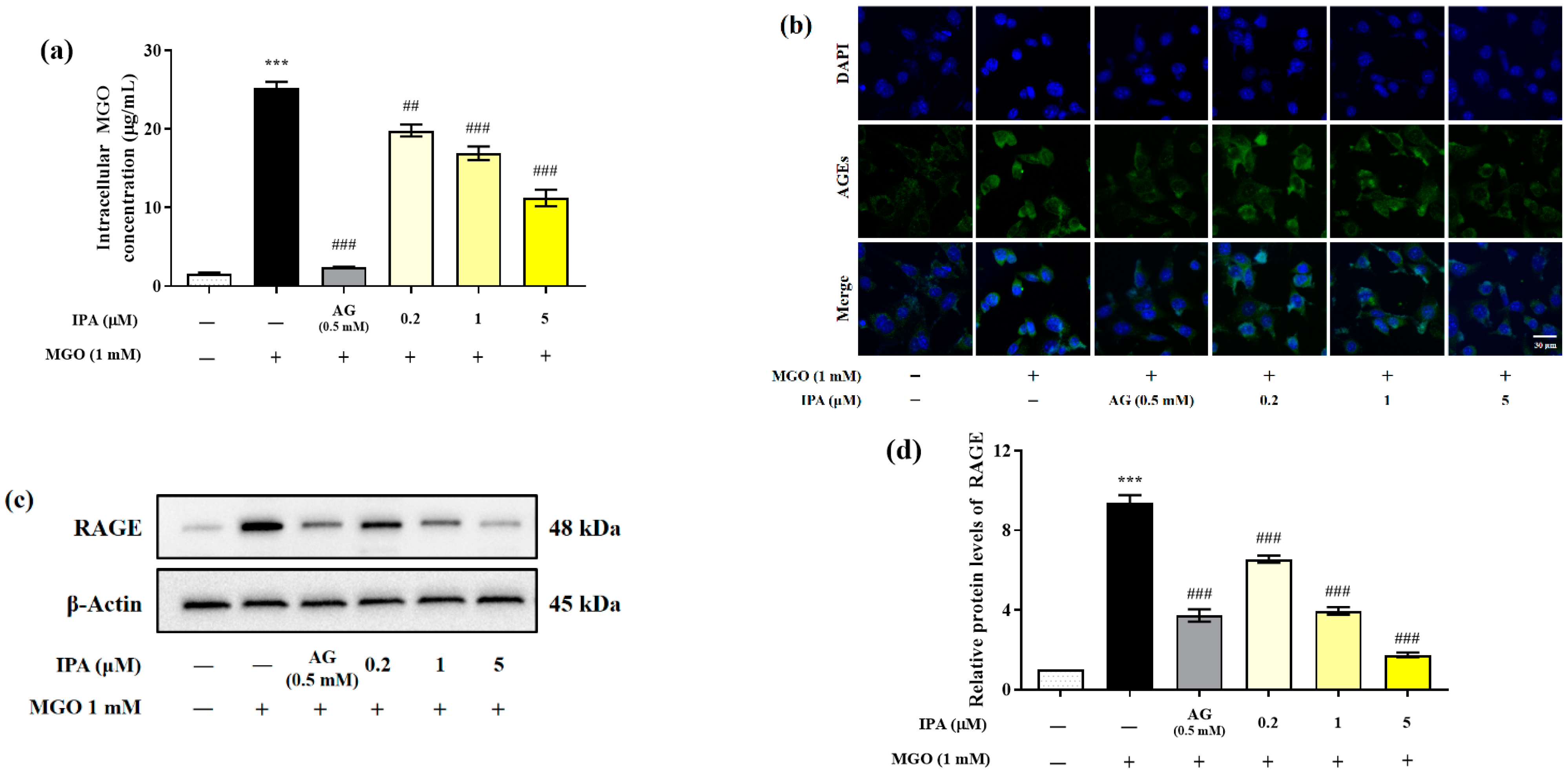
2.3. Inhibitory Effects of IPA on the Generation of Intracellular AGEs
2.4. Preventive Effects of IPA Against MGO-Induced Renal Cell Apoptosis
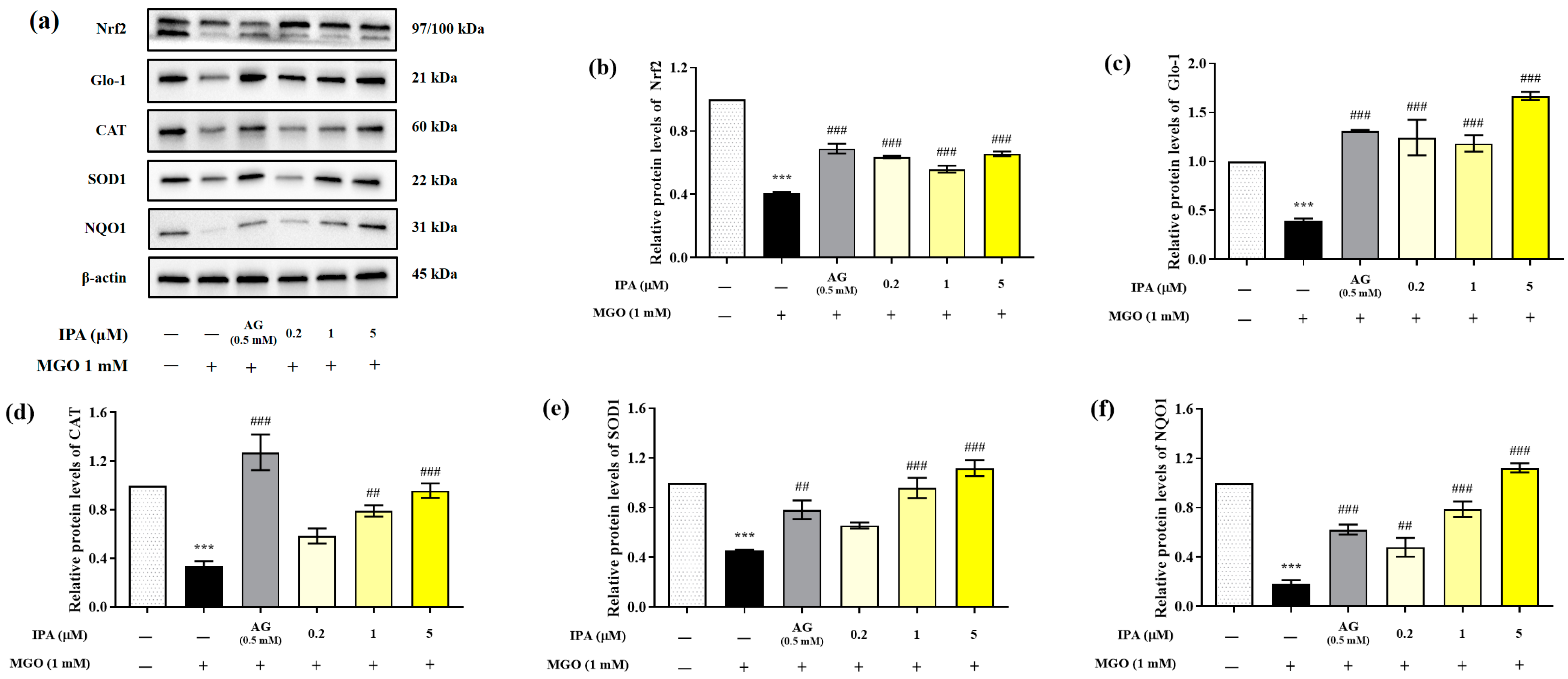
2.5. Effects of IPA on the Nrf2/ARE Signaling Pathway
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Sample Preparation
3.3. Determination of the Inhibitory Effect on AGE Formation
3.4. Determination of the MGO-Derived AGEs–Collagen Cross-Linking Inhibitory Ability
3.5. Determination of the MGO-Derived AGEs–Collagen Cross-Link Breaking Ability
3.6. Cell Culture
3.7. Assessment of Cell Viability
3.8. Intracellular ROS Production
3.9. Intracellular MGO Accumulation
3.10. Immunofluorescence Analysis for Intracellular MGO-Derived AGE Accumulation
3.11. Hoechst 33342/PI Double Staining
3.12. Apoptosis Analysis
3.13. Western Blotting
3.14. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rabbani, N.; Thornalley, P.J. Advanced glycation end products in the pathogenesis of chronic kidney disease. Kidney Int. 2018, 93, 803–813. [Google Scholar] [CrossRef]
- Vlassara, H.; Palace, M.R. Diabetes and advanced glycation endproducts. J. Intern. Med. 2002, 251, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Rwan, S.; Bejarano, E.; Taylor, A. Mechanistic targeting of advanced glycation end-products in age-related disease. BBA Mol. Basis Dis. 2018, 1864, 3631–3643. [Google Scholar] [CrossRef]
- Lisa, R.; Paulo, M.; Joana, C.; Daniela, S.-S.; Cristina, S.; Paulo, P.; Raquel, S. Advanced glycation end products and diabetic nephropathy: A comparative study using diabetic and normal rats with methylglyoxal-induced glycation. J. Physiol. Biochem. 2014, 70, 173–184. [Google Scholar]
- Lugt, T.V.D.; Opperhuizen, A.; Bast, A.; Vrolijk, M.F. Dietary advanced glycation end products and the gastrointestinal tract. Nutrients 2020, 12, 2814. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, Y.; Fu, L. Dietary advanced glycation end-products: Perspectives linking food processing with healyh implications. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2559–2587. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N. Advanced glycation endproducts-role in pathology of diabetic complications. Diabetes Res. Clin. Pract. 2005, 67, 3–21. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Thornalley, P.J. Advanced glycation endproducts: What is their releveance to diabetic complications? Diabetes Obes. Metab. 2007, 9, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Sireesh, D.; Bhakkiyalakshmi, E.; Ponjayanthi, B.; Rajaguru, P.; Ramkumar, K.M. Pathophysiological insights of methylglyoxal induced type-2 diabetes. Chem. Res. Toxicol. 2015, 28, 1666–1674. [Google Scholar]
- Rabbani, N.; Thornalley, P.J. Methylglyoxal, glyoxalase 1 and the dicarbonyl proteome. Amino Acids 2012, 42, 1133–1142. [Google Scholar] [CrossRef]
- Tan, A.L.; Forbes, J.M.; Cooper, M.E. AGE, RAGE, and ROS in diabetic nephropathy. Semin. Nephrol. 2007, 27, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Davoud, S.; Amir, G.H.; Hassan, A.; Somayeh, A. AGE-RAGE axis blockade in diabetic nephropathy: Current status and future directions. Eur. J. Pharmacol. 2018, 833, 158–164. [Google Scholar]
- Lo, C.-Y.; Hsiao, W.-T.; Chen, X.-Y. Efficiency of trapping methylglyoxal by phenols and phenolic acids. J. Food Sci. 2011, 76, H90–H96. [Google Scholar] [CrossRef]
- Zhu, D.; Wang, L.; Zhou, Q.; Yan, S.; Li, Z.; Sheng, J.; Zhang, W. (+)-Catechin ameliorates diabetic nephropathy by trapping methylglyoxal in type 2 diabetic mice. Mol. Nutr. Food Res. 2014, 58, 2249–2260. [Google Scholar] [CrossRef] [PubMed]
- Anwar, S.; Khan, S.; Almatroudi, A.; Khan, A.A.; Alsahli, M.; Almatroodi, S.A.; Rahmani, A.H. A review on mechanism of inhibition of advanced glycation end products formation by plant derived polyphenolic compounds. Mol. Biol. Rep. 2021, 48, 787–805. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tang, S.; Chen, Y.; Zhang, R.; Zhou, M.; Wang, C.; Feng, N.; Wu, Q. Structure-activity relationship of procyanidins on advanced glycation end products formation and corresponding mechanisms. Food Chem. 2019, 272, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.-H.; Yoo, G.; Kim, M.; Kurniawati, U.D.; Choi, I.-W.; Lee, S.-H. Dieckol, derived from the edible brown algae Ecklonia cava, attenuates methylglyoxal-associated diabetic nephropathy by suppressing AGE-RAGE interaction. Antioxidants 2023, 12, 593. [Google Scholar] [CrossRef]
- Cho, C.H.; Yoo, G.; Kim, M.; Lee, C.J.; Choi, I.-W.; Ryu, B.; Kim, B.-M.; Lee, S.-H. Diphlorethohydroxycarmalol, a phlorotannin contained in brown edible seaweed Ishige okamurae, prevents AGE-related diabetic nephropathy by suppression of AGE-RAGE interaction. Food Biosci. 2023, 53, 102659. [Google Scholar] [CrossRef]
- Sharifuddin, Y.; Chin, Y.-X.; Lim, P.-E.; Phang, S.-M. Potential bioactive compounds from seaweed for diabetes management. Mar. Drugs 2015, 13, 5447–5491. [Google Scholar] [CrossRef]
- Cho, C.H.; Youm, G.H.; Kim, M.; Kim, S.; Song, E.-J.; Nam, Y.-D.; Lee, S.-H. Evaluation of the relationship between bioactive components in seaweeds and advanced glycation end-products inhibitory activities using principal component analysis. Plant Foods Hum. Nutr. 2021, 76, 326–333. [Google Scholar] [CrossRef]
- Fernando, K.H.N.; Yang, H.-W.; Jiang, Y.; Jeon, Y.-J.; Ryu, B. Ishige okamurae extract and its constituent ishophloroglucin A attenuated in vitro and in vivo high glucose-induced angiogenesis. Int. J. Mol. Sci. 2019, 20, 5542. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, H.-W.; Jiang, Y.; Oh, J.-Y.; Jeon, Y.-J.; Ryu, B. Ishophloroglucin A isolated from Ishige okamurae suppresses melanogenesis induced by α-MSH: In vitro and in vivo. Mar. Drugs 2020, 18, 470. [Google Scholar] [CrossRef]
- Yang, H.-W.; Oh, S.; Chung, D.-M.; Seo, M.; Park, S.J.; Jeon, Y.-J.; Byun, J.; Ryu, B. Ishophloroglucin A, isolated from Ishige okamurae, alleviates dexamethasone-induced muscle atrophy through muscle protein metabolism in vivo. Mar. Drugs 2020, 20, 280. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.-W.; Son, M.; Choi, J.; Oh, S.; Jeon, Y.-J.; Byun, K.; Ryu, B. Effect of ishophloroglucin A, a component of Ishige okamurae, on glucose homeostasis in the pancreas and muscle of high fat diat-fed mice. Mar. Drugs 2019, 17, 608. [Google Scholar] [CrossRef]
- Kang, N.; Oh, S.; Kim, H.-S.; Ahn, H.; Choi, J.; Heo, S.-J.; Byun, K.; Jeon, Y.-J. Ishophloroglucin A, derived from Ishige okamurae, regulates high-fat-diet-induced fat accumulation via the leptin signaling pathway, associated with peripheral metabolism. Algal Res. 2020, 50, 101974. [Google Scholar] [CrossRef]
- Cho, S.-H.; Kim, H.-S.; Jung, H.-Y.; Park, J.-I.; Jang, Y.-J.; Ahn, J.; Kim, K.-N. Effect of ishophloroglucin A isolated from Ishige okamurae on in vitro osteoclastogenesis and osteoblastogenesis. Mar. Drugs 2023, 21, 377. [Google Scholar] [CrossRef]
- Kim, M.; Cho, C.; Lee, C.; Ryu, B.; Kim, S.; Hur, J.; Lee, S.-H. Ishige okamurae ameliorates methylglyoxal-induced nephrotoxicity via reducing oxidative stress, RAGE protein expression, and modulating MAPK, Nrf2/ARE signaling pathway in mouse glomerular mesangial cells. Foods 2021, 10, 2000. [Google Scholar] [CrossRef] [PubMed]
- Edelstein, D.; Brownlee, M. Mechanistic studies of advanced glycosylation end product inhibition by aminoguanidine. Diabetes 1992, 41, 26–29. [Google Scholar] [CrossRef]
- Thormalley, P.J. Use of aminoguanidine (Pimagedine) to prevent the formation of advanced glycation endproducts. Arch. Biochem. Biophys. 2003, 419, 31–40. [Google Scholar]
- Vasan, S.; Foiles, P.; Founds, H. Therapeutic potential of breakers of advanced glycation end product-protein crosslinks. Arch. Biochem. Biophys. 2003, 419, 89–96. [Google Scholar] [PubMed]
- Zhu, H.; Poojary, M.M.; Andersen, M.L.; Lund, M.N. The effect of molecular structure of polyphenols on the kinetics of the trapping reactions with methylglyoxal. Food Chem. 2020, 319, 126500. [Google Scholar] [CrossRef]
- Tezuka, Y.; Nakaya, I.; Nakayama, K.; Nakayama, M.; Yahata, M.; Soma, J. Methylglyoxal as a prognostic factor in patients with chronic kidney disease. Nephrology 2019, 24, 943–950. [Google Scholar] [CrossRef]
- Reyaz, A.; Alam, S.; Chandra, K.; Kohli, S.; Agarwal, S. Methylglyoxal and soluble RAGE in type 2 diabetes mellitus: Association with oxidative stress. J. Diabetes Metab. Disord. 2020, 19, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Beisswenger, P.J.; Miller, M.E.; Howell, S.K.; Rich, S.S.; Russell, G.B.; Mauer, M. Early progression of diabetic nephropathy correlates with methylglyoxal-derived advanced glycation end products. Diabetes Care 2013, 36, 3234–3239. [Google Scholar] [CrossRef]
- Lu, J.; Randell, E.; Han, Y.C.; Adeli, K.; Krahn, J.; Meng, Q.H. Increased plasma methylglyoxal level, inflammation, and vascular endothelial dysfunction in diabetic nephropathy. Clin. Biochem. 2011, 44, 307–311. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Han, Y.-P.; Yu, H.-T.; Pu, X.-P.; Du, G.-H. Protocatechualdehyde prevents methylglyoxal-induced mitochondrial dysfunction and AGEs-RAGE axis activation in human lens epithelial cells. Eur. J. Pharmacol. 2014, 738, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, E.; Morita, A.; Mori, A.; Sakamoto, K.; Nakahara, T. The process of methylglyoxal-induced retinal capillary endothelial cell degeneration in rat. Mcrovasc. Res. 2023, 146, 104455. [Google Scholar] [CrossRef]
- Zhou, Q.; Gong, J.; Wang, M. Phloretin and its methylglyoxal adduct: Implications against advanced glycation end products-induced inflammation in endothelial cells. Food Chem. Toxicol. 2019, 129, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Bai, N.; He, K.; Ho, C.-T.; Yang, C.S.; Sang, S. Apple polyphenols, phloretin and phloridzin: New trapping agents of reactive dicarbonyl species. Chem. Res. Toxicol. 2008, 21, 2042–2050. [Google Scholar] [CrossRef]
- Li, X.; Zheng, T.; Sang, S.; Lv, L. Quercetin inhibits advanced glycation end product formation by trapping methylglyoxal and glyoxal. J. Agric. Food Chem. 2014, 62, 12152–12158. [Google Scholar] [CrossRef]
- Cho, C.-H.; Lee, C.-J.; Kim, M.-G.; Ryu, B.; Je, J.-G.; Kim, Y.; Lee, S.-H. Therapeutic potential of phlorotannin-rich Ecklonia cava extract on methylglyoxal-induced diabetic nephropathy in in vitro model. Mar. Drugs 2022, 20, 355. [Google Scholar] [CrossRef]
- Wang, G.; Wang, Y.; Yang, Q.; Xu, C.; Zheng, Y.; Wang, L.; Wu, J.; Zeng, M.; Luo, M. Metformin prevents methylglyoxal-induced apoptosis by suppressing oxidative stress in vitro and in vivo. Cell Death Dis. 2022, 13, 29. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.-F.; Miyata, S.; Hirota, Y.; Higo, S.; Miyazaki, H.; Fukunaga, M.; Hamada, Y.; Ueyama, S.; Muramoto, O.; Uriuhara, A.; et al. Methylglyoxal induces apoptosis through activation of p38 mitogen-activated protein kinase in rat mesangial cells. Kideny Int. 2003, 63, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, M.; Miyata, S.; Liu, B.F.; Miyazaki, H.; Hirota, Y.; Higo, S.; Hamada, Y.; Ueyama, S.; Kasuga, M. Methylglyoxal induces apoptosis through activation of p38 MAPK in rat Schwann cells. Biochem. Biophys. Res. Commun. 2004, 320, 689–695. [Google Scholar] [CrossRef]
- Do, M.H.; Lee, J.H.; Cho, K.; Knag, M.C.; Subedi, L.; Parveen, A.; Kim, S.Y. Therapeutic potential of Lespedeza bicolor to prevent methylglyoxal-induced glucotoxicity in familiar diabetic nephropathy. J. Clin. Med. 2019, 8, 1138. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Chaudhari, B.P.; Kakkar, P. Baicalin and chrysin mixture imparts cyto-protection against methylglyoxal induced cytotoxicity and diabetic tubular injury by modulating RAGE, oxidative stress and inflammation. Environ. Toxicol. Pharmacol. 2017, 50, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Ryu, B.; Jiang, Y.; Kim, H.-S.; Hyun, J.-M.; Lim, S.-B.; Li, Y.; Jeon, Y.-J. Ishophloroglucin A, a novel phlorotannin for standardizing the anti-α-glucosidase activity of Ishige okamurae. Mar. Drugs 2018, 16, 436. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, C.-H.; Kim, M.-G.; Ryu, B.; Lee, S.-H. Ishophloroglucin A Isolated from Ishige okamurae Protects Glomerular Cells from Methylglyoxal-Induced Diacarbonyl Stress and Inhibits the Pathogenesis of Diabetic Nephropathy. Mar. Drugs 2025, 23, 48. https://doi.org/10.3390/md23010048
Cho C-H, Kim M-G, Ryu B, Lee S-H. Ishophloroglucin A Isolated from Ishige okamurae Protects Glomerular Cells from Methylglyoxal-Induced Diacarbonyl Stress and Inhibits the Pathogenesis of Diabetic Nephropathy. Marine Drugs. 2025; 23(1):48. https://doi.org/10.3390/md23010048
Chicago/Turabian StyleCho, Chi-Heung, Min-Gyeong Kim, Bomi Ryu, and Sang-Hoon Lee. 2025. "Ishophloroglucin A Isolated from Ishige okamurae Protects Glomerular Cells from Methylglyoxal-Induced Diacarbonyl Stress and Inhibits the Pathogenesis of Diabetic Nephropathy" Marine Drugs 23, no. 1: 48. https://doi.org/10.3390/md23010048
APA StyleCho, C.-H., Kim, M.-G., Ryu, B., & Lee, S.-H. (2025). Ishophloroglucin A Isolated from Ishige okamurae Protects Glomerular Cells from Methylglyoxal-Induced Diacarbonyl Stress and Inhibits the Pathogenesis of Diabetic Nephropathy. Marine Drugs, 23(1), 48. https://doi.org/10.3390/md23010048