
Generation, Characterisation and Identification of Bioactive Peptides from Mesopelagic Fish Protein Hydrolysates Using In Silico and In Vitro Approaches
 ,
,  ,
,  , ,
, ,  , , ,
, , ,
Abstract
1. Introduction
2. Results and Discussion
2.1. Hydrolysate Generation from Irish and Norwegian Mesopelagic Trawls and Proximate Compositional Analysis of Resulting Hydrolysates
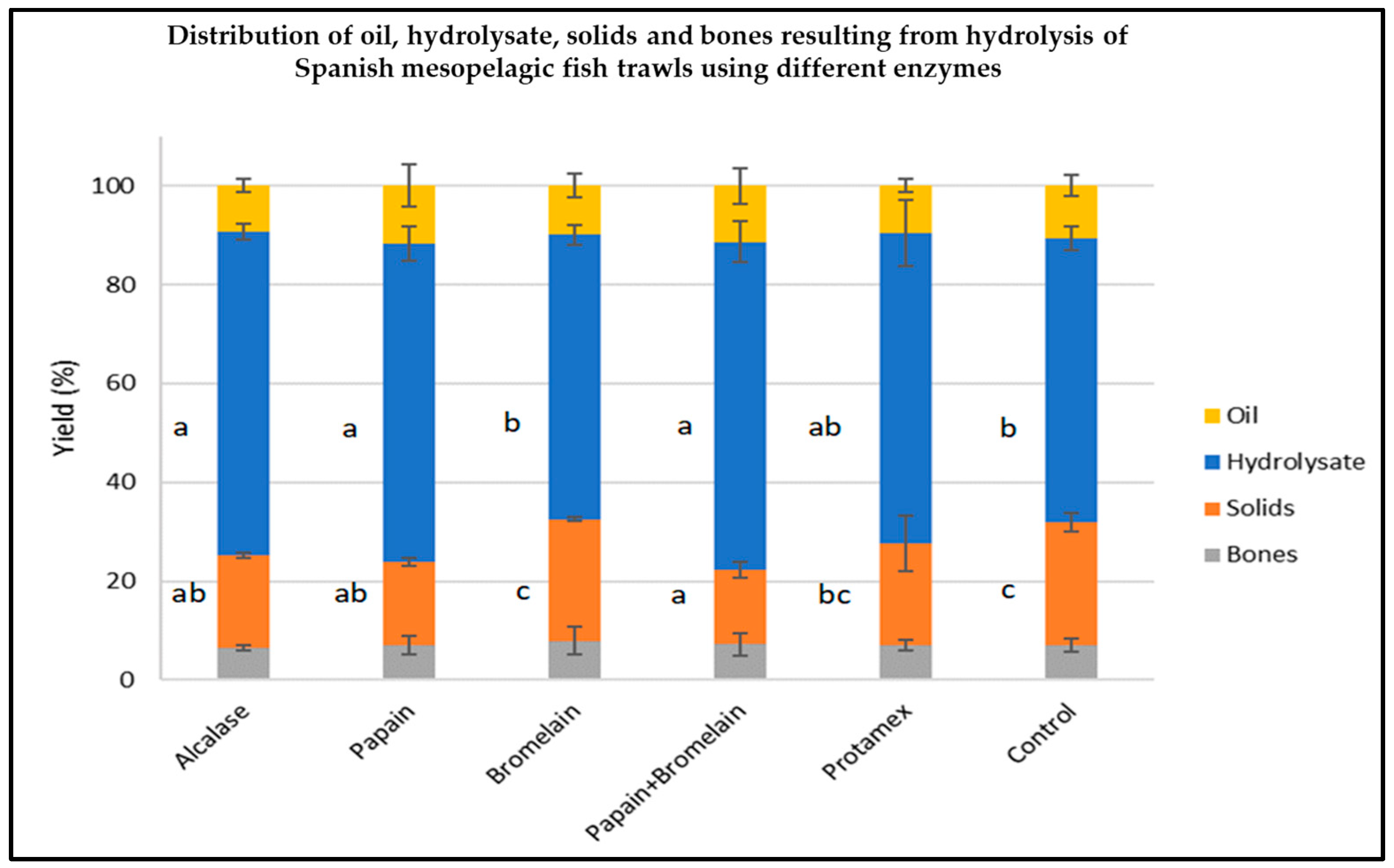
2.2. Hydrolysate Generation from Spanish Mesopelagic Fish Trawl
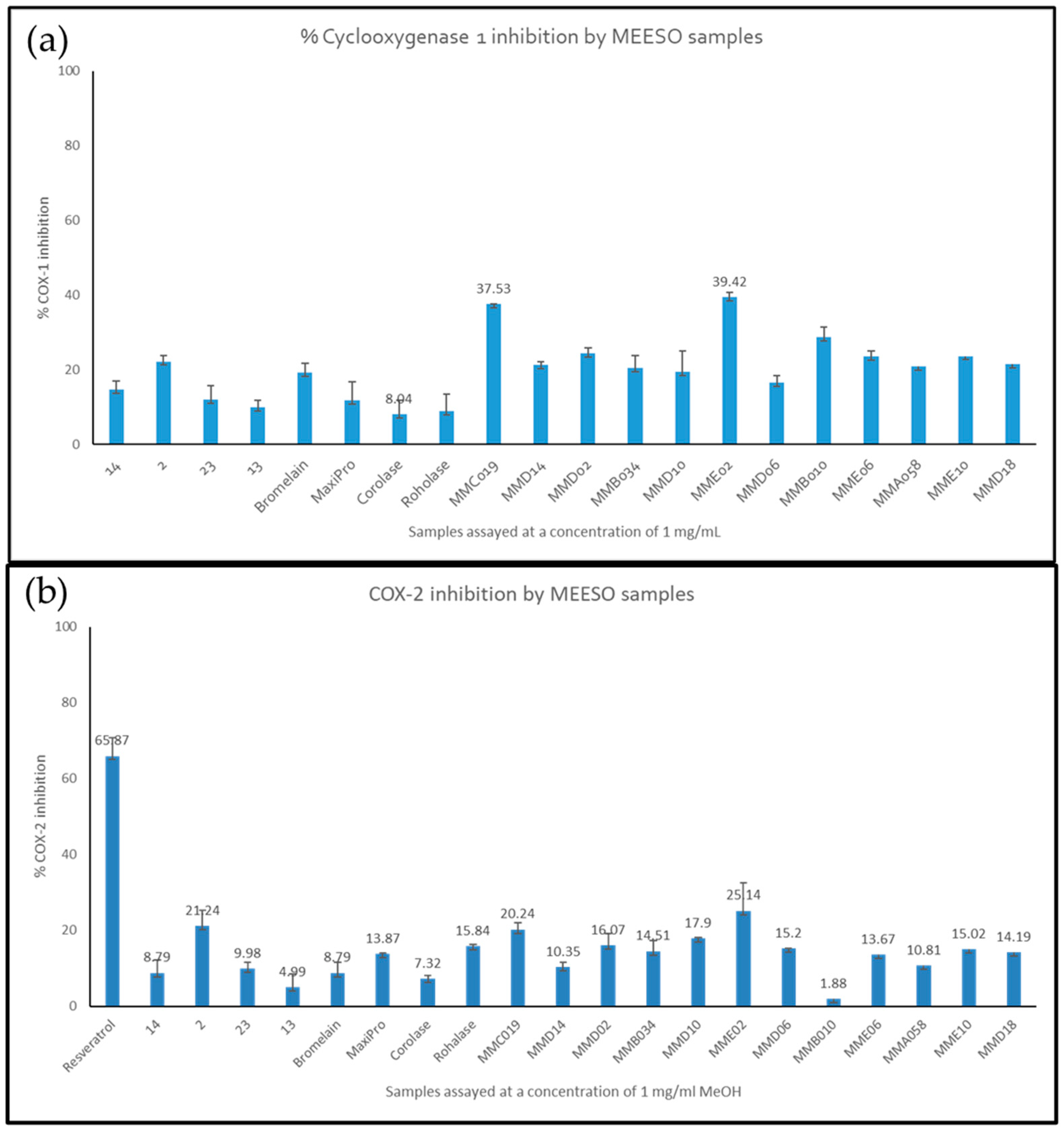
2.3. Cyclooxygenase (COX) Enzyme Inhibition by Generated Hydrolysates
2.4. Monoacylglycerol Lipase (MAGL) Enzyme Inhibition by Generated Hydrolysates
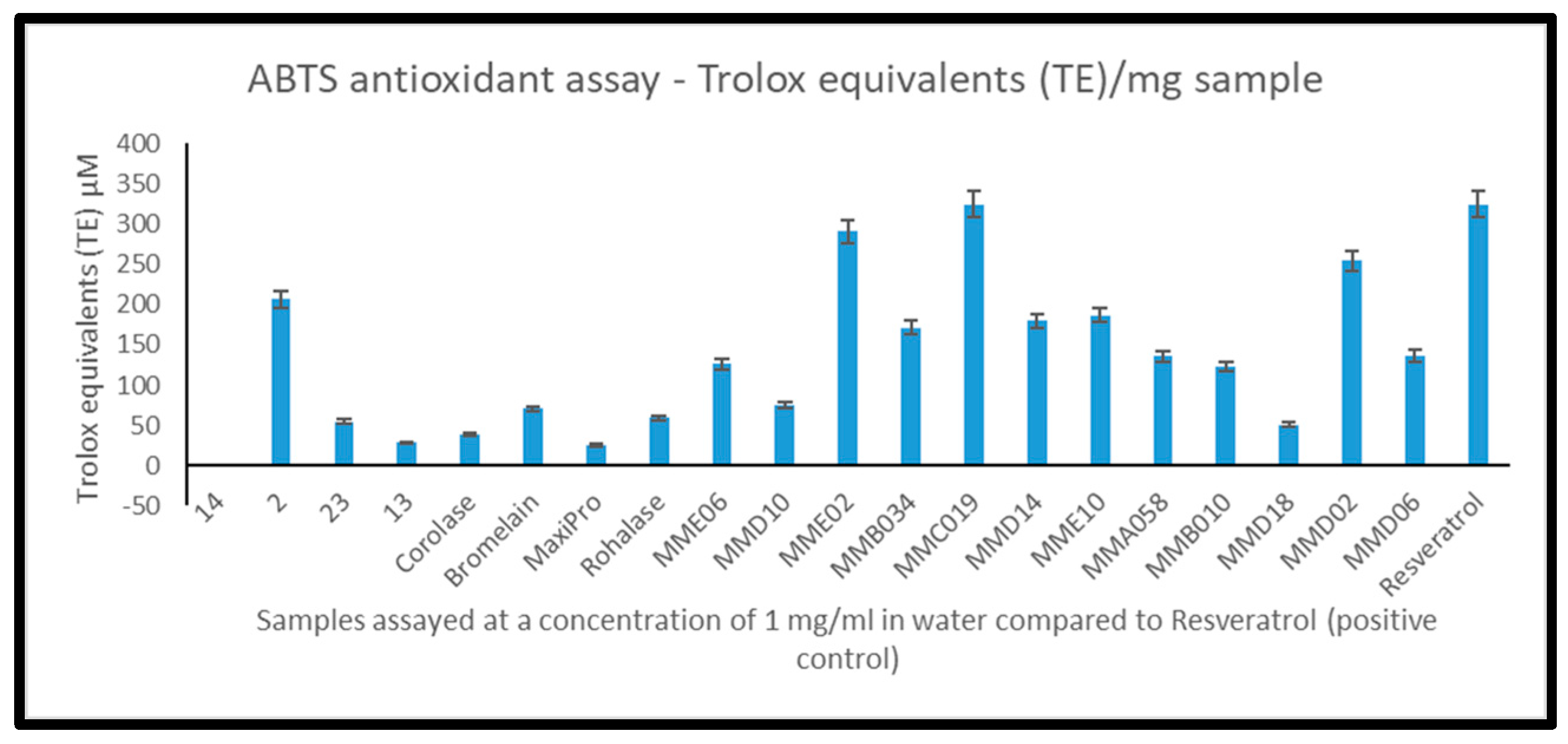
2.5. 2,2′-Azino-bis-3-ethylbenzthiazoline-6-sulphonic Acid Total Antioxidant Capacity (ABTS) of Mesopelagic Hydrolysates
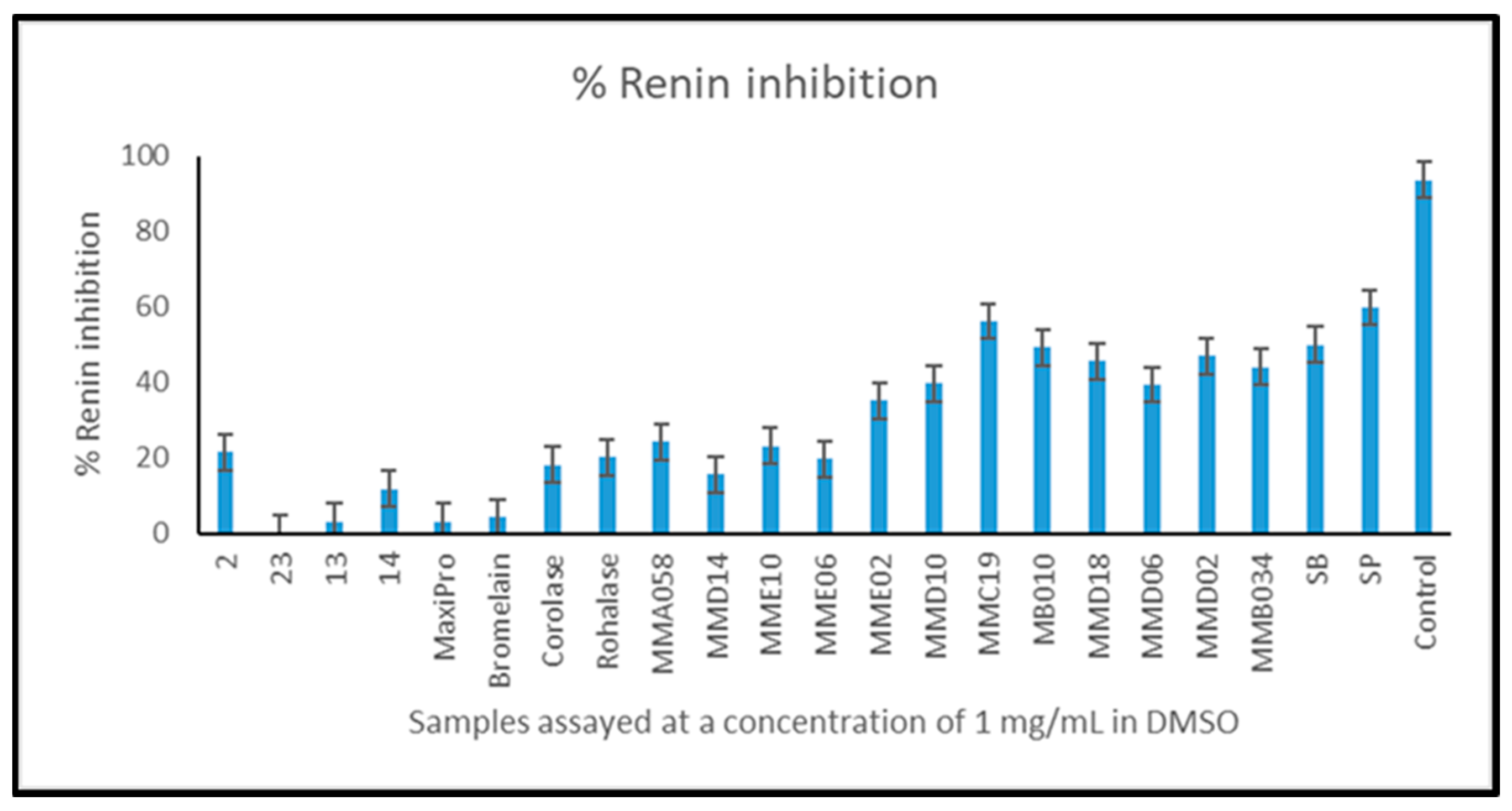
2.6. Angiotensin-1-Converting Enzyme (ACE-1) and Renin Inhibition by Mesopelagic Hydrolysates
2.7. Dipeptidyl Peptidase IV Inhibition by Mesopelagic Hydrolysates
2.8. Identification of Bioactive Peptides Using Mass Spectrometry and In Silico Analysis
2.9. Chemical Synthesis and Confirmation of Anti-Inflammatory Activity of Peptides Using In Vitro COX and MAGL Inhibition Bioassays
3. Materials and Methods
3.1. Supply and Processing of Raw Materials
3.2. Enzymatic Hydrolysis of Irish, Norwegian and Spanish Samples
3.3. Hydrolysate Enrichment Using Molecular Weight Cut-Off (MWCO) Filtration
3.4. Proximate Compositional Analysis of Mesopelagic Species
3.5. Bioactivity Assessments of Hydrolysates and Peptides with In Vitro Screening Assays
3.5.1. Cyclooxygenase (COX; EC E.C. 1.14. 99.1) Inhibition COX-1 and COX-2
activity)/(Corrected 100% initial activity) × 100
3.5.2. Monoacylglycerol Lipase (MAGL; EC 3.1. 1. 23) Inhibition
3.5.3. ACE-I Inhibition Assay
3.5.4. DPP-IV Inhibition Assay
3.5.5. Antioxidant Capacity: ABTS Assay
3.5.6. Renin Inhibition Activity
3.6. Peptides Identified Using Mass Spectrometry
3.7. Assessment of Bioactive Potential of Identified Peptides
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Staby, A.; Røstad, A.; Kaartvedt, S. Long-term acoustical observations of the mesopelagic fish Maurolicus muelleri reveal novel and varied vertical migration patterns. Mar. Ecol. Prog. Ser. 2011, 441, 241–255. [Google Scholar]
- Naik, A.S.; Whitaker, R.D.; Albrektsen, S.; Solstad, R.G.; Thoresen, L.; Hayes, M. Mesopelagic fish protein hydrolysates and extracts: A source of novel anti-hypertensive and anti-diabetic peptides. Front. Mar. Sci. 2021, 8, 1–9. [Google Scholar] [CrossRef]
- Abdulbasit, A. Biochemical composition and bioactive properties of mesopelagic fish-derived protein hydrolysate from Northern Krill (Meganyctiphanes norvegica) and Muller’s Pearlside (Maurolicus muelleri) for possible utilization of novel marine resources. Master’s Thesis in Biotechnology, Norwegian University of Science and Technology, Trondheim, Norway, 2022. [Google Scholar]
- Madina, M.A.; Grimaldo, E.; Grimsmo, L.; Toldnes, B.; Slizyte, R.; Carvajal, A.K.; Schei, M.; Selnes, M.; Falch, E. Exploring the Potential of Atlantic Mesopelagic Species Processed on Board Commercial Fishing Vessels as a Source of Dietary Lipids. Foods 2024, 13, 1094. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.H.; Kriest, I.; Getzlaff, J. Diving deeper: Mesopelagic fish biomass estimates comparison using two different models. Front. Mar. Sci. 2023, 10, 1121569. [Google Scholar] [CrossRef]
- Calcinai, L.; Bonomini, M.G.; Leni, G.; Faccini, A.; Puxeddu, I.; Giannini, D.; Petrelli, F.; Prandi, B.; Sforza, S.; Tedeschi, T. Effectiveness of enzymatic hydrolysis for reducing the allergenic potential of legume by-products. Sci. Rep. 2022, 12, 16902. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Rao, Q. Effect of Processing on Fish Protein Antigenicity and Allergenicity. Foods 2021, 10, 969. [Google Scholar] [CrossRef] [PubMed]
- Gianfranceschi, G.L.; Gianfranceschi, G.; Quassinti, L.; Bramucci, M. Biochemical requirements of bioactive peptides for nutraceutical efficacy. J. Funct. Foods 2018, 47, 252–263. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef]
- Lund, A.; Knop, F.K.; Vilsbøll, T. Glucagon-like peptide-1 receptor agonist for the treatment of type 2 diabetes: Differences and similarities. Eur. J. Intern. Med. 2014, 25, 407–414. [Google Scholar] [CrossRef]
- Magon, N. Gonadotropin releasing hormone agonists: Expanding vistas. Indian J. Endocrinol. Metab. 2011, 15, 261–267. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Rossino, G.; Marchese, E.; Galli, G.; Verde, F.; Finizio, M.; Serra, M.; Linciano, P.; Collina, S. Peptides as Therapeutic Agents: Challenges and Opportunities in the Green Transition Era. Molecules 2023, 28, 7165. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Placha, D.; Jampilek, J. Chronic Inflammatory Diseases, Anti-Inflammatory Agents and Their Delivery Nanosystems. Pharmaceutics 2021, 13, 64. [Google Scholar] [CrossRef] [PubMed]
- Bu, J.; Ding, R.; Zhou, L.; Chen, X.; Shen, E. Epidemiology of Psoriasis and Comorbid Diseases: A Narrative Review. Front. Immunol. 2022, 13, 880201. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhou, W.B.S.; Meng, J.; Zang, J. Does low-grade systemic inflammation have a role in chronic pain? Front. Mol. Neurosci. 2021, 14, 785214. [Google Scholar] [CrossRef] [PubMed]
- Bindu, S.; Mazumder, S.; Bandyopadhyay, U. Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A current perspective. Biochem. Pharmacol. 2020, 180, 114147. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Smith, C.J.; Zhang, Y.; Koboldt, C.M.; Muhammad, J.; Zweifel, B.S.; Shaffer, A.; Talley, J.J.; Masferrer, J.L.; Seibert, K.; Isakson, P.C. Pharmacological analysis of cyclooxygenase-1 in inflammation. Proc. Natl. Acad. Sci. USA 1998, 95, 13313–13318. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Curtis, E.; Fuggle, N.; Shaw, S.; Spooner, L.; Ntani, G.; Parsons, C.; Corp, N.; Honvo, G.; Baird, J.; Maggi, S.; et al. Safety of Cyclooxygenase-2 Inhibitors in Osteoarthritis: Outcomes of a Systematic Review and Meta-Analysis. Drugs Aging 2019, 36, 25–44. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Momin, R.A.; Nair, M.G.; Bourquin, L.D. Cyclooxygenase inhibitory and antioxidant cyanidin glycosides in cherries and berries. Phytomedicine 2001, 8, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Allaire, M.; Al Sayegh, R.; Mabire, M.; Hammoutene, A.; Siebert, M.; Caër, C.; Cadoux, M.; Wan, J.; Habib, A.; Le Gall, M.; et al. Monoacylglycerol lipase reprograms hepatocytes and macrophages to promote liver regeneration. JHEP Rep. 2023, 8, 100794. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Liang, Q.; Chalamaiah, M.; Liao, W.; Ren, X.; Ma, H.; Wu, J. Zein hydrolysate and its peptides exert anti-inflammatory activity on endothelial cells by preventing TNF-α-induced NF-ĸβ activation. J. Funct. Foods 2020, 64, 103598. [Google Scholar] [CrossRef]
- Dia, V.P.; Wang, W.; Oh, V.L.; de Lumen, B.O.; Gonzalez de Mejia, E. Isolation, purification and characterisation of lunasin from defatted soybean flour and in vitro evaluation of its anti-inflammatory activity. Food Chem. 2009, 114, 108–115. [Google Scholar] [CrossRef]
- Ahn, C.B.; Cho, Y.S.; Je, J.Y. Purification and anti-inflammatory action of tripeptide from salmon pectoral fin byproduct protein hydrolysate. Food Chem. 2015, 1, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.L.; Shin, S.R.; Yoon, K.Y. Hydrolysis conditions for antioxidant peptides derived from enzymatic hydrolysates of sandfish (Arctoscopus japonicus). Food Sci. Biotechnol. 2017, 26, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Kemp, D.C.; Kwon, J.Y. Fish and Shellfish-Derived Anti-Inflammatory Protein Products: Properties and Mechanisms. Molecules 2021, 26, 3225. [Google Scholar] [CrossRef] [PubMed]
- Association of Official Analytical Chemists. Official Methods of Analysis of AOAC International; AOAC International: Rockville, MD, USA, 1998. [Google Scholar]
- Deng, H.; Li, W. Monoacylglycerol lipase inhibitors: Modulators for lipid metabolism in cancer malignancy, neurological and metabolic disorders. Acta Pharm. Sin. B 2020, 10, 582–602. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lafarga, T.; Aluko, R.E.; Rai, D.K.; O’Connor, P.; Hayes, M. Identification of bioactive peptides from a papain hydrolysate of bovine serum albumin and assessment of an antihypertensive effect in Spontaneously Hypertensive Rats. Food Res. Int. 2016, 81, 91–99. [Google Scholar] [CrossRef]
- Hayes, M.; Mora, L.; Lucakova, S. Identification of Bioactive Peptides from Nannochloropsis oculata Using a Combination of Enzymatic Treatment, in Silico Analysis and Chemical Synthesis. Biomolecules 2022, 12, 1806. [Google Scholar] [CrossRef] [PubMed]
- Mooney, C.; Haslam, N.; Pollastri, G.; Shields, D.C. Towards the improved discovery and design of functional peptides: Common features of diverse classes permit generalized prediction of bioactivity. PLoS ONE 2012, 7, e45012. [Google Scholar] [CrossRef]
- Khatun, M.S.; Hasan, M.M.; Kurata, H. PreAIP: Computational Prediction of Anti-inflammatory Peptides by Integrating Multiple Complementary Features. Front. Genet. 2019, 10, 00129. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Qi, L.; Du, J.; Sun, Y.; Xiong, Y.; Zhao, X.; Pan, D.; Zhi, Y.; Dang, Y.; Gao, X. Umami-MRNN: Deep learning based prediction of umami peptide using RNN and MLP. Food Chem. 2023, 405, 134935. [Google Scholar] [CrossRef]
- Chen, X.; Huang, J.; He, B. AntiDMPpred: A web service for identifying anti-diabetic peptides. Peer J. 2022, 10, e13581. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ju, Z.; Li, M.; Xu, J.; Howell, D.C.; Li, Z.; Chen, F.E. Recent development on COX-2 inhibitors as promising anti-inflammatory agents: The past 10 years. Acta Pharm. Sin. B 2022, 12, 2790–2807. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Rivera-Jiménez, J.; Berraquero-García, C.; Pérez-Gálve, R.; García-Moreno, P.J.; Espejo-Carpio, F.J.; Guadix, A.; Guadix, E.M. Peptides and protein hydrolysates exhibiting anti-inflammatory activity: Sources, structural features and modulation mechanisms. Food Funct. 2022, 13, 12510. [Google Scholar] [CrossRef] [PubMed]
- Shin, T.-S.; Choi, K.S.; Chun, J.; Kho, K.H.; Son, S.A.; Shim, S.-Y. Anti-inflammatory effect of Abalone (aliotis discus Hannai) Viscera via inhibition of ROS production in LPS stimulated RAW 267.4 cells. Microbiol. Biotechnol. Lett. 2022, 50, 21–30. [Google Scholar] [CrossRef]
- Ghanbari, R.; Ebahimpour, M.; Zarei, A.; Ismail, A.; Abdul-Hamid, A.; Saari, N. Purification and Characterisation of Nitric Oxide Inhibitory peptides from Actinopyga lecanora through enzymatic hydrolysis. Food Biotechnol. 2016, 30, 263–277. [Google Scholar]
- Grancieri, M.; Martino, H.S.D.; Gonzalez de Mejia, E. Digested total protein and protein fractions from chia seed (Salvia hispanica L.) had high scavenging capacity and inhibited 5-LOX, COX-1-2, and iNOS enzymes. Food Chem. 2019, 289, 204–214. [Google Scholar] [PubMed]
- Shahi, Z.; Sayyed-Alangi, S.Z.; Najafian, L. Effects of enzyme type and process time on hydrolysis degree, electrophoresis bands and antioxidant properties of hydrolyzed proteins derived from defatted Bunium persicum Bioss. press cake. Heliyon 2020, 6, e03365. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Yu, R.; Choi, Y.; Ma, Z.Z.; Zhang, H.; Xiang, W.; Lee, D.Y.; Berman, B.M.; Moudgil, K.D.; Fong, H.H.; et al. Discovery of cyclooxygenase inhibitors from medicinal plants used to treat inflammation. Pharmacol. Res. 2010, 61, 519–524. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Jha, V.; Biagi, M.; Spinelli, V.; Di Stefano, M.; Macchia, M.; Minutolo, F.; Granchi, C.; Poli, G.; Tuccinardi, T. Discovery of Monoacylglycerol Lipase (MAGL) Inhibitors Based on a Pharmacophore-Guided Virtual Screening Study. Molecules 2021, 26, 78. [Google Scholar] [CrossRef]
- Zhang, Y.; Venkitasamy, C.; Pan, Z.; Liu, W.; Zhao, L. Novel Umami Ingredients: Umami peptides and their taste. J. Food Sci. 2017, 82, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Mei, S.; Yao, S.; Mo, J.; Wang, Y.; Tang, J.; Li, W.; Wu, T. Integration of cloud-based molecular networking and docking for enhanced umami peptide screening from Pixian douban. Food Chem. X 2023, 21, 101098. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Belloir, C.; Savistchenko, J.; Neiers, F.; Taylor, A.J.; McGrane, S.; Briand, L. Biophysical and functional characterization of the N-terminal domain of the cat T1R1 umami taste receptor expressed in Escherichia coli. PLoS ONE 2017, 12, e0187051. [Google Scholar] [CrossRef] [PubMed]
- Hoyle, N.T.; Merritt, J.H. Quality of fish protein hydrolysates from herring (Clupea harengus). J. Food Sci. 1994, 59, 76–79. [Google Scholar] [CrossRef]
- Nielsen, P.H.; Petersen, D.; Dambmann, C.J.J.O.F.S. Improved method for determining food protein degree of hydrolysis. J. Food Sci. 2001, 66, 642–646. [Google Scholar] [CrossRef]
- Hayes, M.; Aluko, R.E.; Aurino, E.; Mora, L. Generation of Bioactive Peptides from Porphyridium sp. and Assessment of Their Potential for Use in the Prevention of Hypertension, Inflammation and Pain. Mar. Drugs 2023, 21, 422. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Bao, J.; Lao, X.; Zheng, H. Novel 3D Structure Based Model for Activity Prediction and Design of Antimicrobial Peptides. Sci. Rep. 2018, 8, 11189. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.K.; Lee, B.; Kim, S.T.; Yoo, J.S.; Sung, J.-S. Designing a novel functional peptide with dual antimicrobial and anti-inflammatory activities via in silico methods. Front. Immunol. 2022, 13, 821070. [Google Scholar] [CrossRef]
- Iwaniak, A.; Minkiewicz, P.; Darewicz, M. BIOPEP-UWM database—Present and future. Curr. Opin. Food Sci. 2024, 55. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hydrolysate | Peptide Sequence | Peptide Ranker Value | PreAIP RF Combined Values | Anti-Diabetic Prediction (AntiDMPpred) | BIOPEP-UWM | Umami |
|---|---|---|---|---|---|---|
| CE21009 Haul 23 Maurolicus muelleri (Code 23) Alcalase hydrolysate. Irish sample. Freeze-dried. High Confidence AIP (0.482). | KTLRKMGKWCCHCFPCCRGSGKSNVGAW | 0.999 | High confidence AIP (0.731) | Low probability | Novel | umami, predicted threshold: 25.47 mmol/L |
| DGINVLGLIVFCLVLGIVIGRKWEKGQIL | 0.996 | High confidence AIP (0.560) | Low probability | Novel | umami, predicted threshold: 12.65 mmol/L | |
| Origin—Thin-lipped Mullet | FDAFLPM | 0.955 | Medium confidence AIP (0.392) | Low probability | Novel | Non-umami |
| GLGGMLF | 0.939 | Low confidence AIP (0.370) | Low probability | Novel | umami, predicted threshold: 35.46 mmol/L | |
| Origin—Salmo salar—Atlantic Salmon | QCPLHRPWAL | 0.932 | High confidence AIP (0.499) | Low probability | Novel | Non-umami |
| LACNCNLHARRCRFNM | 0.908 | High confidence AIP (0.629) | Low probability | Novel | umami, predicted threshold: 37.66 mmol/L | |
| TFSWGFDDFSCC | 0.889 | High confidence AIP (0.496) | Low probability | Novel | umami, predicted threshold: 14.82 mmol/L | |
| GINVLGLIVFCLVLGI | 0.888 | High confidence AIP (0.620) | Low probability | Novel | umami, predicted threshold: 35.08 mmol/L | |
| LLSSELQSLLIATTCLRELISCC | 0.873 | High confidence (0.614) | Low probability | Novel | umami, predicted threshold: 13.25 mmol/L | |
| Origin—Makaira nigricans—Atlantic Blue Marlin | NVGEVVCIFLTAALGLPEALI | 0.868 | High confidence AIP (0.612) | Likely to be anti-diabetic (probability of 0.8) | Novel | umami, predicted threshold: 19.89 mmol/L |
| Maurolicus muelleri (MMC019) Endogenous enzyme autolysis (Spanish sample), spray dried. High confidence AIP (0.514) | SFVPNGASLEDCHCNLPCLA | 0.874 | High confidence AIP (0.506) | Low probability | Novel | umami, predicted threshold: 30.65 mmol/L |
| GFSAVNMRKFG | 0.797 | High confidence AIP (0.527) | Low probability | Novel | umami, predicted threshold: 30.85 mmol/L | |
| Origin—Cypinus carpio—Common Carp | IAGFEIFDFNSLEQLC | 0.734 | High confidence AIP (0.540) | Low probability | Novel | umami, predicted threshold: 36.00 mmol/L |
| NLFKDCNF | 0.693 | Medium confidence AIP (0.467) | Low probability | Novel | umami, predicted threshold: 18.79 mmol/L | |
| PFGAADQDPF | 0.677 | Low confidence AIP (0.370) | Low probability | Novel | Non-umami | |
| NSGAGILPSPSTPRFP | 0.621 | Medium confidence AIP (0.453) | Low probability | Novel | umami, predicted threshold: 25.95 mmol/L | |
| DVEFLPPQLPSDKFKDDPVG | 0.601 | Medium confidence AIP (0.433) | Low probability | Novel | umami, predicted threshold: 20.45 mmol/L | |
| Origin—Takifuga rubipes—Japanese Puffer Fish | GFAGDDAPR | 0.598 | Negative AIP (0.284) | Low probability | Novel | umami, predicted threshold: 1.68 mmol/L |
| FSPFGAAD | 0.58 | Low confidence AIP (0.346) | Low probability | Novel | umami, predicted threshold: 17.39 mmol/L | |
| PSRILYG | 0.574 | Medium confidence AIP (0.412) | Low probability | Novel | Non-umami | |
| Maurolicus muelleri (MME02—Spanish haul) (medium confidence AIP—0.446) | VFIPFNPL | 0.871 | Low confidence AIP (0.382) | Low probability | Novel | Non-umami |
| NDLPWEF | 0.861 | Low confidence AIP (0.349) | Low probability | Novel | Non-umami | |
| VLLFFYAPWCGQ | 0.846 | High confidence AIP (0.524) | Low probability | Novel | Non-umami | |
| CGRASCPVLCSG | 0.845 | High confidence AIP (0.480) | Low probability | Novel | umami, predicted threshold: 23.60 mmol/L | |
| Origin—Makaira nigricans—Atlantic Blue Marlin | GFNPPDLDIM | 0.828 | Low confidence AIP (0.382) | Low probability | Novel | non-umami |
| Origin—Cypinus carpio—Common Carp | SDNAYQFMLT | 0.72 | Medium confidence AIP (0.412) | Low probability | Novel | umami, predicted threshold: 34.06 mmol/L |
| CLGSPNPLDII | 0.687 | Medium confidence AIP (0.408) | Low probability | Novel | umami, predicted threshold: 36.98 mmol/L | |
| RCPEALF | 0.672 | High confidence AIP (0.556) | Low probability | Novel | non-umami | |
| ADDEDADGESSGEPPGAPKQEEAI | 0.667 | High confidence AIP (0.469) | Low probability | Novel | umami, predicted threshold: 8.32 mmol/L | |
| DSFGRLT | 0.662 | Low confidence AIP (0.387) | Low probability | Novel | umami, predicted threshold: 12.87 mmol/L |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayes, M.; Naik, A.; Mora, L.; Iñarra, B.; Ibarruri, J.; Bald, C.; Cariou, T.; Reid, D.; Gallagher, M.; Dragøy, R.; et al. Generation, Characterisation and Identification of Bioactive Peptides from Mesopelagic Fish Protein Hydrolysates Using In Silico and In Vitro Approaches. Mar. Drugs 2024, 22, 297. https://doi.org/10.3390/md22070297
Hayes M, Naik A, Mora L, Iñarra B, Ibarruri J, Bald C, Cariou T, Reid D, Gallagher M, Dragøy R, et al. Generation, Characterisation and Identification of Bioactive Peptides from Mesopelagic Fish Protein Hydrolysates Using In Silico and In Vitro Approaches. Marine Drugs. 2024; 22(7):297. https://doi.org/10.3390/md22070297
Chicago/Turabian StyleHayes, Maria, Azza Naik, Leticia Mora, Bruno Iñarra, Jone Ibarruri, Carlos Bald, Thibault Cariou, David Reid, Michael Gallagher, Ragnhild Dragøy, and et al. 2024. "Generation, Characterisation and Identification of Bioactive Peptides from Mesopelagic Fish Protein Hydrolysates Using In Silico and In Vitro Approaches" Marine Drugs 22, no. 7: 297. https://doi.org/10.3390/md22070297
APA StyleHayes, M., Naik, A., Mora, L., Iñarra, B., Ibarruri, J., Bald, C., Cariou, T., Reid, D., Gallagher, M., Dragøy, R., Galino, J., Deyà, A., Albrektsen, S., Thoresen, L., & Solstad, R. G. (2024). Generation, Characterisation and Identification of Bioactive Peptides from Mesopelagic Fish Protein Hydrolysates Using In Silico and In Vitro Approaches. Marine Drugs, 22(7), 297. https://doi.org/10.3390/md22070297