
Algae-Based Nanoparticles for Oral Drug Delivery Systems

 , ,
, ,  and
and 
Abstract
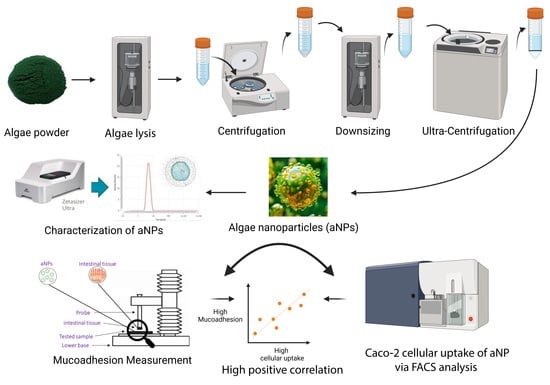
1. Introduction
2. Results and Discussion
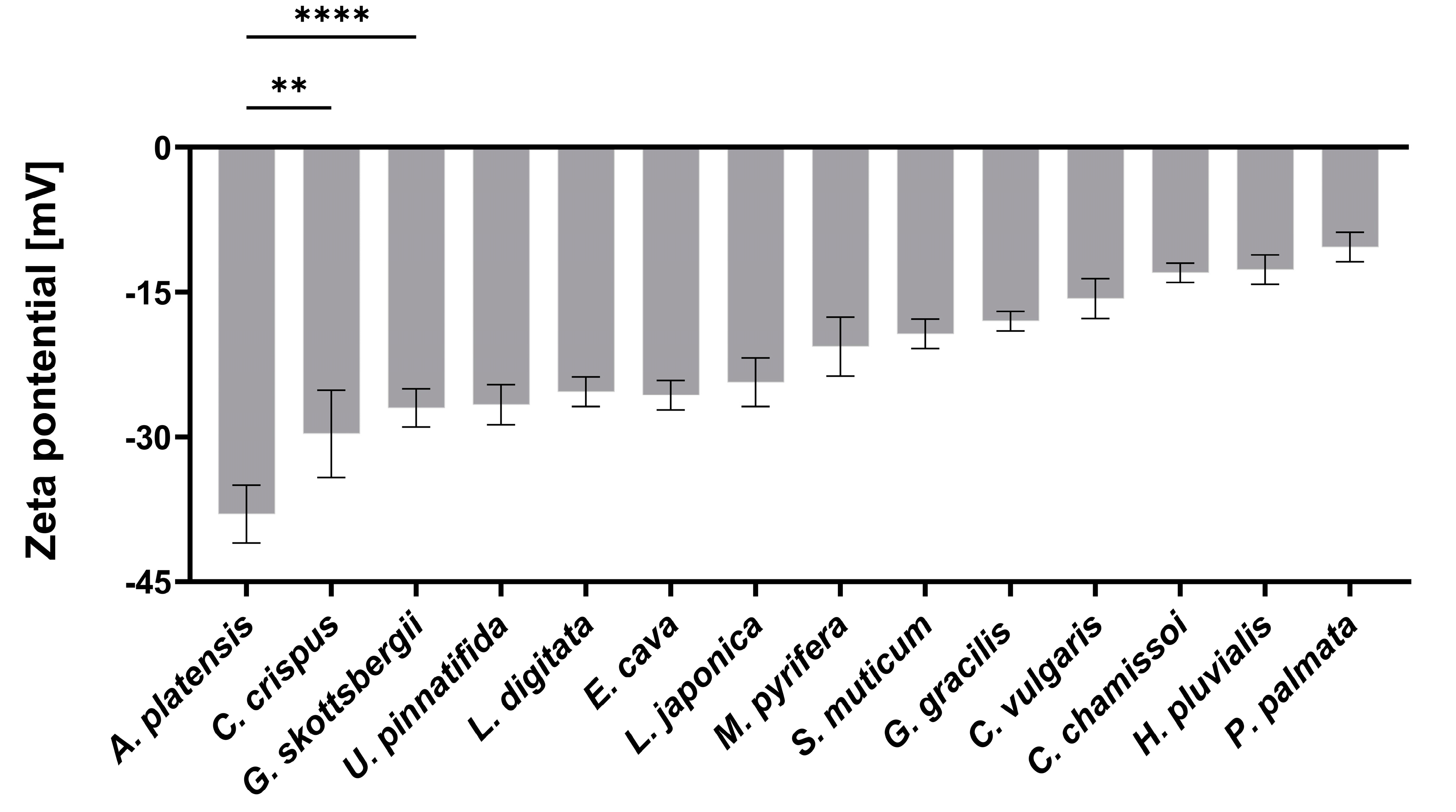
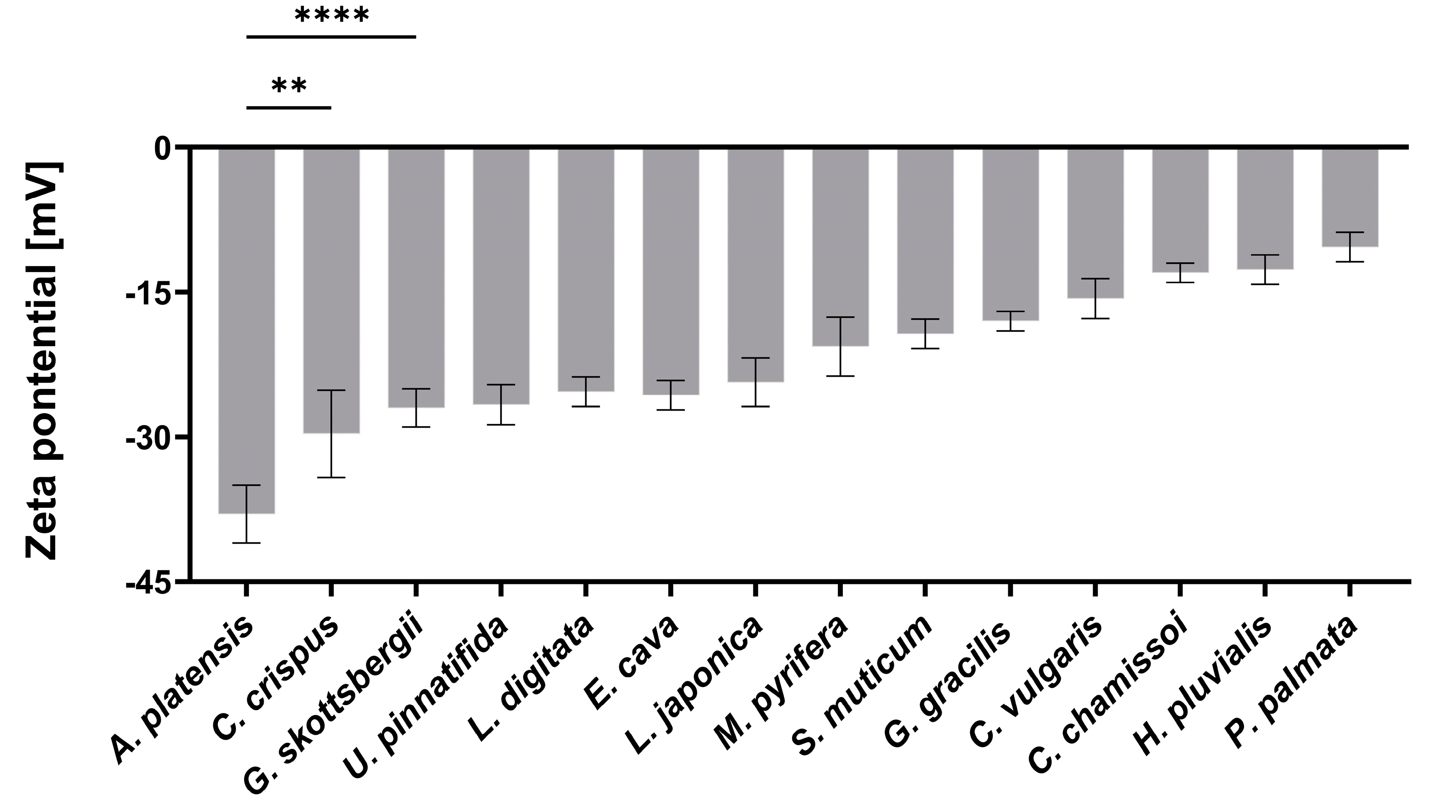
2.1. Characterization of aNPs
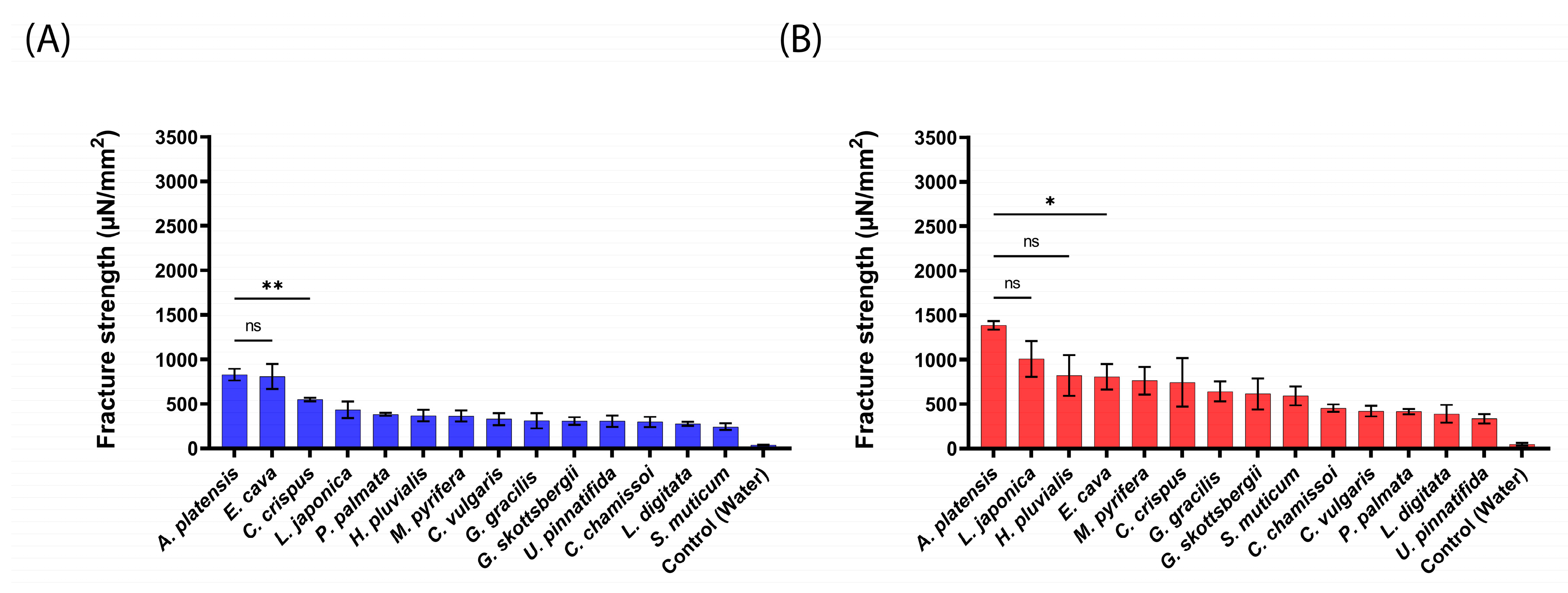
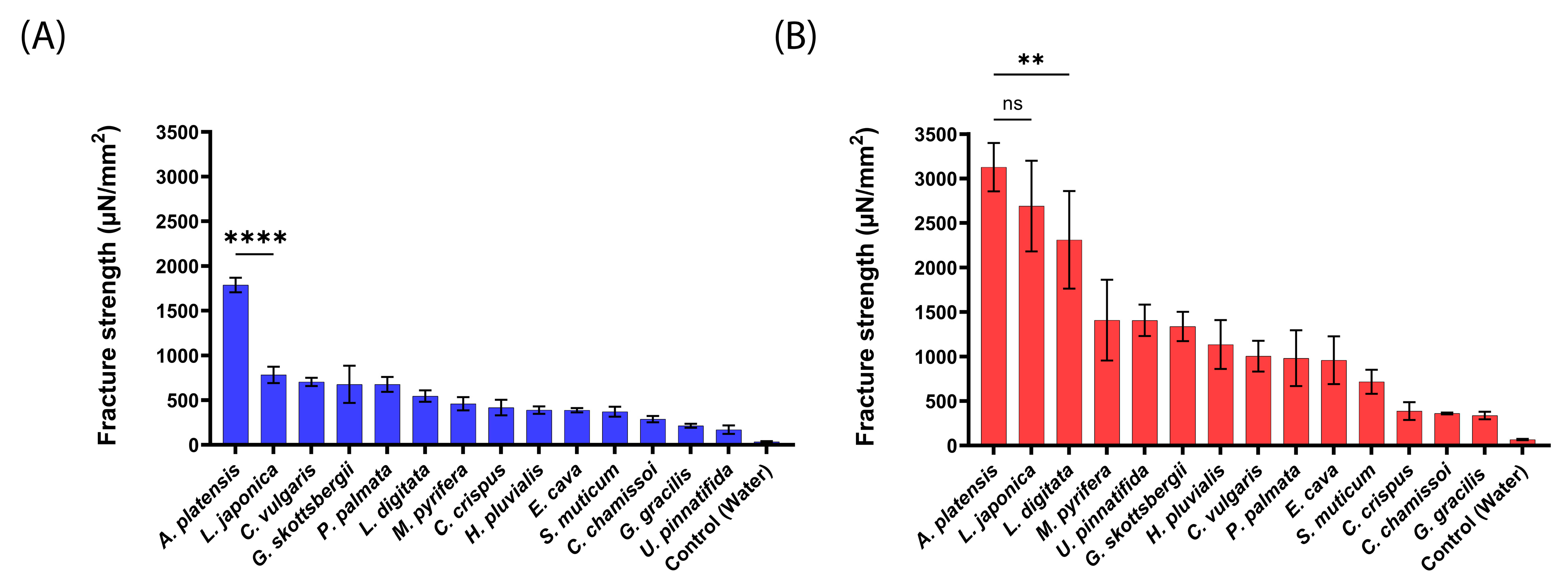
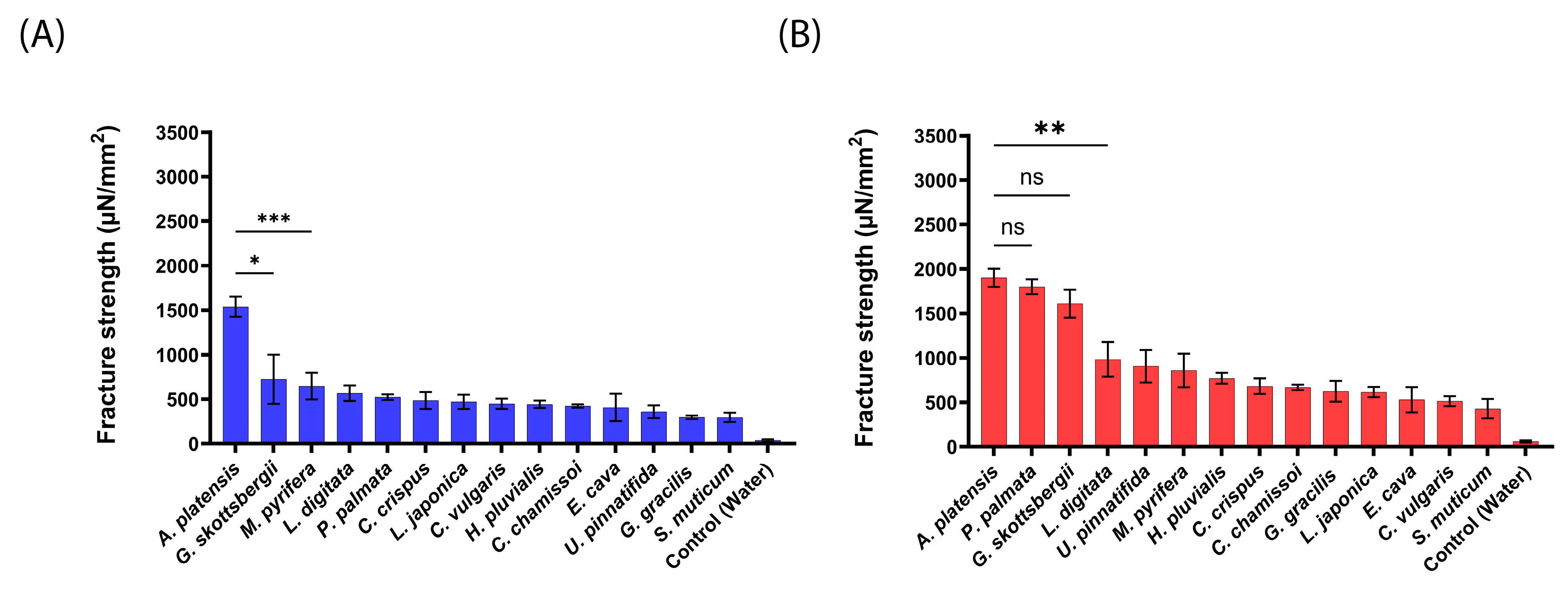
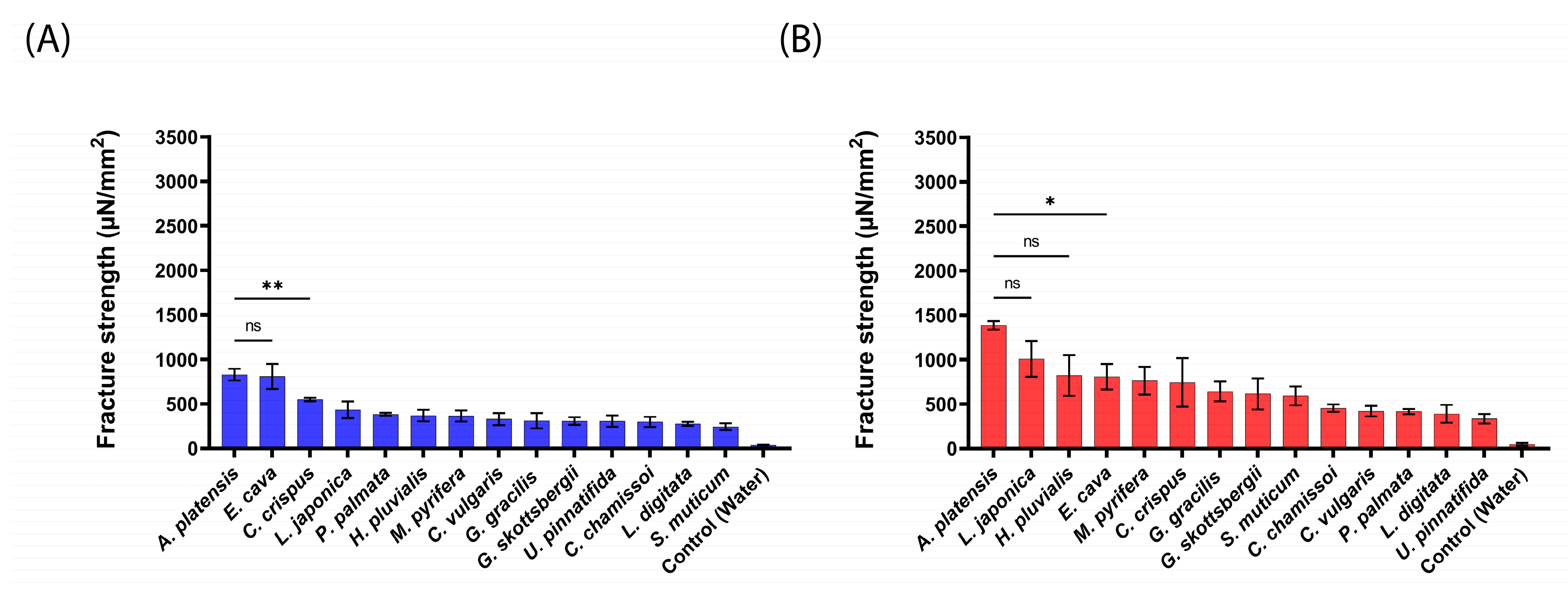
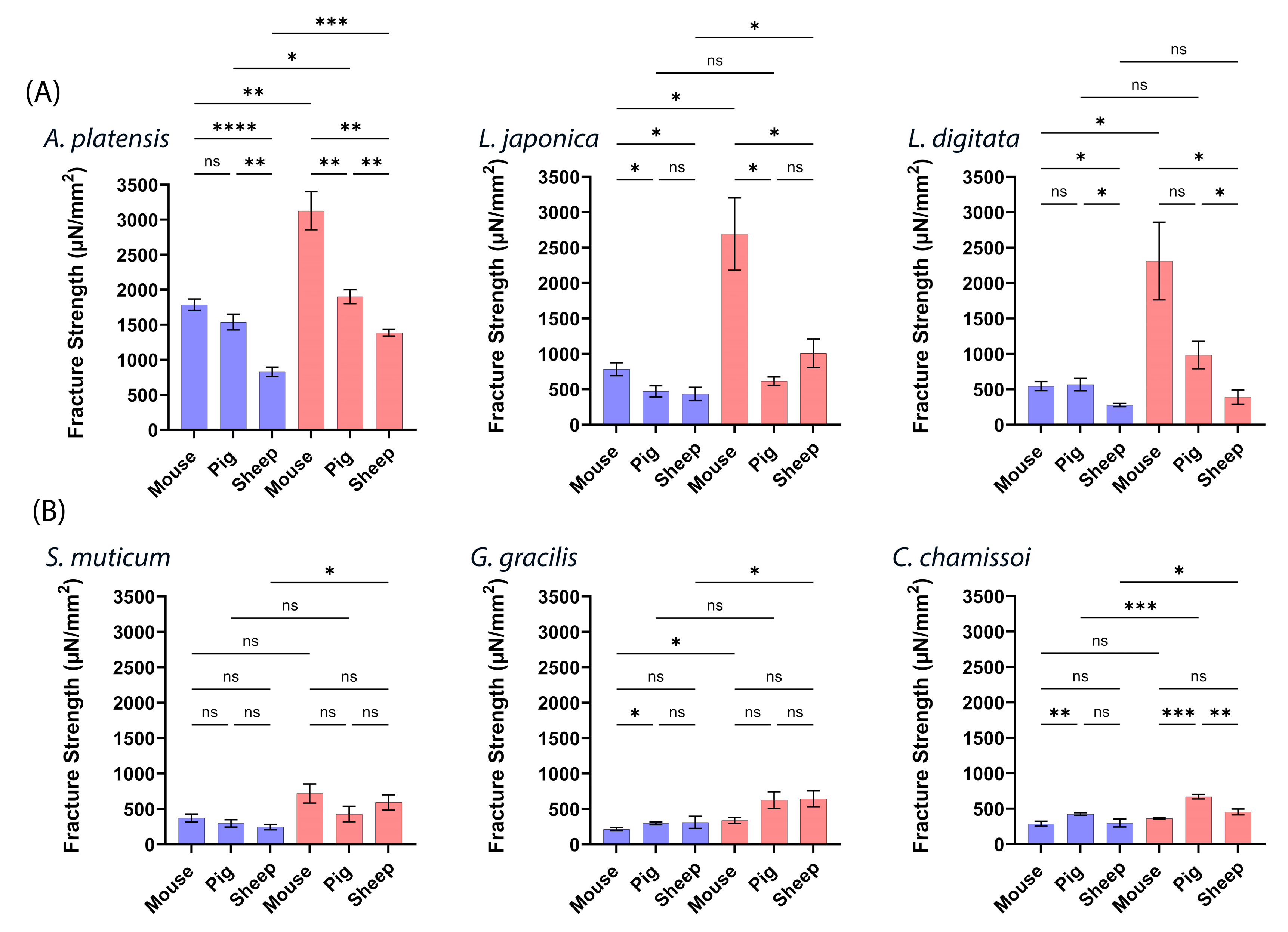
2.2. Mucoadhesive Force of the aNPs
2.3. Protein BLAST of Mucin2
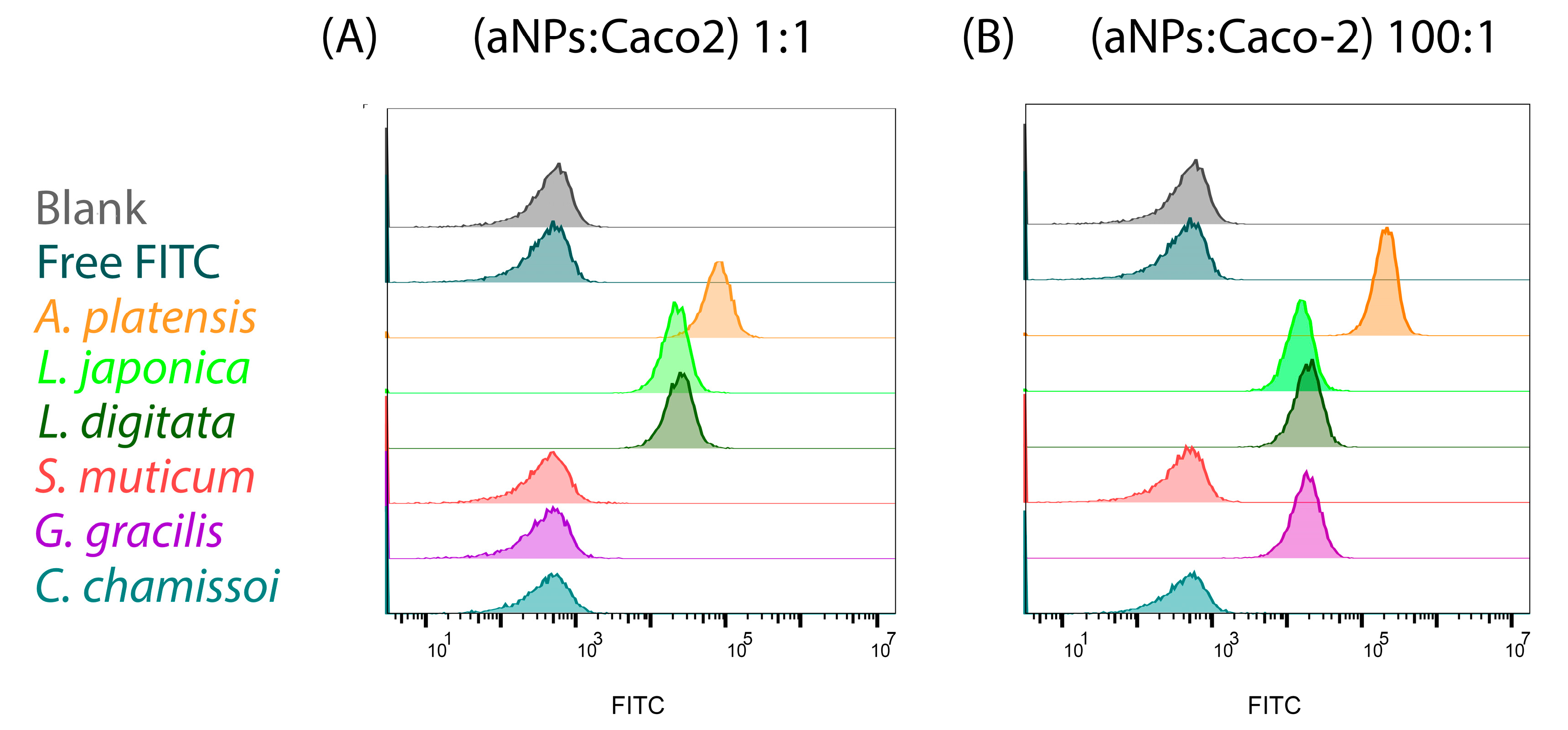
2.4. Cellular Uptake of aNPs
3. Conclusions
4. Materials and Methods
4.1. Materials
4.2. Preparation of aNPs
4.3. Characterization of aNPs
4.4. Mucoadhesion Measurement of aNPs
4.5. Encapsulation of Fluorescent Molecule in aNPs
4.6. Release profile of FD40 from A. platensis NPs
4.7. Protein BLAST Analysis
4.8. Caco-2 Uptake of aNPs
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Jesmeen, T.; Uddin, R. Orodispersible Tablets: A Short Review. Stamford J. Pharm. Sci. 2011, 4, 96–99. [Google Scholar] [CrossRef]
- Murdan, S.; Wei, L.; van Riet-Nales, D.A.; Gurmu, A.E.; Usifoh, S.F.; Tăerel, A.E.; Yıldız-Peköz, A.; Krajnović, D.; Azzopardi, L.M.; Brock, T.; et al. Association between Culture and the Preference for, and Perceptions of, 11 Routes of Medicine Administration: A Survey in 21 Countries and Regions. Explor. Res. Clin. Soc. Pharm. 2023, 12, 100378. [Google Scholar] [CrossRef] [PubMed]
- Azagury, A.; Fonseca, V.C.; Cho, D.Y.; Perez-rogers, J.; Baker, M.; Steranka, E.; Goldenshtein, V.; Calvao, D.; Eric, M. Single Step Double-Walled Nanoencapsulation (SSDN ). J. Control. Release 2018, 280, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Azagury, A.; Amar-Lewis, E.; Appel, R.; Hallak, M.; Kost, J. Amplified CPEs Enhancement of Chorioamnion Membrane Mass Transport by Encapsulation in Nano-Sized PLGA Particles. Eur. J. Pharm. Biopharm. 2017, 117, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Bozzuto, G.; Molinari, A. Liposomes as Nanomedical Devices. Int. J. Nanomed. 2015, 10, 975–999. [Google Scholar] [CrossRef]
- Rahamim, V.; Azagury, A. Bioengineered Biomimetic and Bioinspired Noninvasive Drug Delivery Systems. Adv. Funct. Mater. 2021, 31, 2033. [Google Scholar] [CrossRef]
- Chabria, Y.; Duffy, G.P.; Lowery, A.J.; Dwyer, R.M. Hydrogels: 3d Drug Delivery Systems for Nanoparticles and Extracellular Vesicles. Biomedicines 2021, 9, 1694. [Google Scholar] [CrossRef]
- Sasaki, D.; Kusamori, K.; Takayama, Y.; Itakura, S.; Todo, H.; Nishikawa, M. Development of Nanoparticles Derived from Corn as Mass Producible Bionanoparticles with Anticancer Activity. Sci. Rep. 2021, 11, 22818. [Google Scholar] [CrossRef]
- Wang, Q.; Ren, Y.; Mu, J.; Egilmez, N.K.; Zhuang, X.; Deng, Z.; Zhang, L.; Yan, J.; Miller, D.; Zhang, H.G. Grapefruit-Derived Nanovectors Use an Activated Leukocyte Trafficking Pathway to Deliver Therapeutic Agents to Inflammatory Tumor Sites. Cancer Res. 2015, 75, 2520–2529. [Google Scholar] [CrossRef]
- Zhang, M.; Viennois, E.; Prasad, M.; Zhang, Y.; Wang, L.; Zhang, Z.; Han, M.K.; Xiao, B.; Xu, C.; Srinivasan, S.; et al. Edible Ginger-Derived Nanoparticles: A Novel Therapeutic Approach for the Prevention and Treatment of Inflammatory Bowel Disease and Colitis-Associated Cancer. Biomaterials 2016, 101, 321–340. [Google Scholar] [CrossRef]
- Alqahtani, M.S.; Kazi, M.; Alsenaidy, M.A.; Ahmad, M.Z. Advances in Oral Drug Delivery. Front. Pharmacol. 2021, 12, 618411. [Google Scholar] [CrossRef] [PubMed]
- Mudie, D.M.; Amidon, G.L.; Amidon, G.E. Physiological Parameters for Oral Delivery and in Vitro Testing. Mol. Pharm. 2010, 7, 1388–1405. [Google Scholar] [CrossRef]
- Blynskaya, E.V.; Tishkov, S.V.; Vinogradov, V.P.; Alekseev, K.V.; Marakhova, A.I.; Vetcher, A.A. Polymeric Excipients in the Technology of Floating Drug Delivery Systems. Pharmaceutics 2022, 14, 2779. [Google Scholar] [CrossRef]
- Gómez-Guillén, M.C.; Montero, M.P. Enhancement of Oral Bioavailability of Natural Compounds and Probiotics by Mucoadhesive Tailored Biopolymer-Based Nanoparticles: A Review. Food Hydrocoll. 2021, 118, 106772. [Google Scholar] [CrossRef]
- Rosso, A.; Andretto, V.; Chevalier, Y.; Kryza, D.; Sidi-Boumedine, J.; Grenha, A.; Guerreiro, F.; Gharsallaoui, A.; La Padula, V.; Montembault, A.; et al. Nanocomposite Sponges for Enhancing Intestinal Residence Time Following Oral Administration. J. Control. Release 2021, 333, 579–592. [Google Scholar] [CrossRef]
- Reineke, J.; Cho, D.Y.; Dingle, Y.L.; Cheifetz, P.; Laulicht, B.; Lavin, D.; Furtado, S.; Mathiowitz, E. Can Bioadhesive Nanoparticles Allow for More Effective Particle Uptake from the Small Intestine? J. Control. Release 2013, 170, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Cui, Z.; Guo, S.; Zhang, X.; Huo, Y.; Mao, S. Mucoadhesive versus Mucopenetrating Nanoparticles for Oral Delivery of Insulin. Acta Biomater. 2021, 135, 506–519. [Google Scholar] [CrossRef]
- Zhang, M.; Merlin, D. Nanoparticle-Based Oral Drug Delivery Systems Targeting the Colon for Treatment of Ulcerative Colitis. Inflamm. Bowel Dis. 2018, 24, 1401–1415. [Google Scholar] [CrossRef]
- Choukaife, H.; Seyam, S.; Alallam, B.; Doolaanea, A.A.; Alfatama, M. Current Advances in Chitosan Nanoparticles Based Oral Drug Delivery for Colorectal Cancer Treatment. Int. J. Nanomed. 2022, 17, 3933–3966. [Google Scholar] [CrossRef]
- Yermak, I.M.; Davydova, V.N.; Volod’ko, A.V. Mucoadhesive Marine Polysaccharides. Mar. Drugs 2022, 20, 522. [Google Scholar] [CrossRef]
- Zahariev, N.; Katsarov, P.; Lukova, P.; Pilicheva, B. Novel Fucoidan Pharmaceutical Formulations and Their Potential Application in Oncology—A Review. Polymers 2023, 15, 3242. [Google Scholar] [CrossRef] [PubMed]
- Parodi, A.; Molinaro, R.; Sushnitha, M.; Evangelopoulos, M.; Martinez, J.O.; Arrighetti, N.; Corbo, C.; Tasciotti, E. Bio-Inspired Engineering of Cell- and Virus-like Nanoparticles for Drug Delivery. Biomaterials 2017, 147, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Zhong, D.; Zhang, D.; Xie, T.; Zhou, M. Biodegradable Microalgae-Based Carriers for Targeted Delivery and Imaging-Guided Therapy toward Lung Metastasis of Breast Cancer. Small 2020, 16, 2000819. [Google Scholar] [CrossRef]
- Sambuy, Y.; De Angelis, I.; Ranaldi, G.; Scarino, M.L.; Stammati, A.; Zucco, F. The Caco-2 Cell Line as a Model of the Intestinal Barrier: Influence of Cell and Culture-Related Factors on Caco-2 Cell Functional Characteristics. Cell Biol. Toxicol. 2005, 21, 1–26. [Google Scholar] [CrossRef]
- Ji, N.; Hong, Y.; Gu, Z.; Cheng, L.; Li, Z.; Li, C. Chitosan Coating of Zein-Carboxymethylated Short-Chain Amylose Nanocomposites Improves Oral Bioavailability of Insulin in Vitro and in Vivo. J. Control. Release 2019, 313, 1–13. [Google Scholar] [CrossRef]
- Azagury, A.; Baptista, C.; Milovanovic, K.; Shin, H.; Morello, P.; Perez-Rogers, J.; Goldenshtein, V.; Nguyen, T.; Markel, A.; Rege, S.; et al. Biocoating—A Critical Step Governing the Oral Delivery of Polymeric Nanoparticles. Small 2022, 18, 2107559. [Google Scholar] [CrossRef]
- Takeuchi, H.; Yamamoto, H.; Kawashima, Y. Mucoadhesive Nanoparticulate Systems for Peptide Drug Delivery. Adv. Drug Deliv. Rev. 2001, 47, 39–54. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, P. Mucoadhesive Delivery System: A Smart Way to Improve Bioavailability of Nutraceuticals. Foods 2021, 10, 1362. [Google Scholar] [CrossRef]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. The Lipid Bilayer. In Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002. [Google Scholar]
- Hägerström, H.; Edsman, K. Interpretation of Mucoadhesive Properties of Polymer. J. Pharm. Pharmacol. 2001, 53, 1589–1599. [Google Scholar] [CrossRef]
- Leal, J.; Smyth, H.D.C.; Ghosh, D. Physicochemical Properties of Mucus and Their Impact on Transmucosal Drug Delivery. Int. J. Pharm. 2017, 532, 555–572. [Google Scholar] [CrossRef]
- Fallingborg, J. Intraluminal PH of the Human Gastrointestinal Tract. Dan. Med. Bull. 1999, 46, 183–196. [Google Scholar]
- Shimizu, K.; Seiki, I.; Goto, Y.; Murata, T. Measurement of the Intestinal PH in Mice under Various Conditions Reveals Alkalization Induced by Antibiotics. Antibiotics 2021, 10, 180. [Google Scholar] [CrossRef]
- Stanford, A.H.; Gong, H.; Noonan, M.; Lewis, A.N.; Gong, Q.; Lanik, W.E.; Hsieh, J.J.; Lueschow, S.R.; Frey, M.R.; Good, M.; et al. A Direct Comparison of Mouse and Human Intestinal Development Using Epithelial Gene Expression Patterns. Pediatr. Res. 2020, 88, 66–76. [Google Scholar] [CrossRef]
- Nguyen, T.L.A.; Vieira-Silva, S.; Liston, A.; Raes, J. How Informative Is the Mouse for Human Gut Microbiota Research? DMM Dis. Models Mech. 2015, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kegade, P.; Gade, A.; Sawant, R.; Parkar, S. Liposomal Drug Delivery in Cancer. Asian J. Pharm. Res. 2020, 10, 293–298. [Google Scholar] [CrossRef]
- Faassen, F.; Vogel, G.; Spanings, H.; Vromans, H. Caco-2 Permeability, P-Glycoprotein Transport Ratios and Brain Penetration of Heterocyclic Drugs. Int. J. Pharm. 2003, 263, 113–122. [Google Scholar] [CrossRef]
- Salatin, S.; Maleki Dizaj, S.; Yari Khosroushahi, A. Effect of the Surface Modification, Size, and Shape on Cellular Uptake of Nanoparticles. Cell Biol. Int. 2015, 39, 881–890. [Google Scholar] [CrossRef]
- Sherovski, P.; Stojković, G.; Ristovska, N. Development, Validation and Application of First Derivative Spectroscopy Ratio Method for Estimation of Bradford Assay. Anal. Biochem. 2018, 558, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Estrellas, K.M.; Fiecas, M.; Azagury, A.; Laulicht, B.; Cho, D.Y.; Mancini, A.; Reineke, J.; Furtado, S.; Mathiowitz, E. Time-Dependent Mucoadhesion of Conjugated Bioadhesive Polymers. Colloids Surf. B Biointerfaces 2019, 173, 454–469. [Google Scholar] [CrossRef]
- Rehan, M.; Al-Bahadly, I.; Thomas, D.G.; Avci, E. Measurement of Peristaltic Forces Exerted by Living Intestine on Robotic Capsule. IEEE/ASME Trans. Mechatron. 2021, 26, 1803–1811. [Google Scholar] [CrossRef]
- Grondin, J.A.; Kwon, Y.H.; Far, P.M.; Haq, S.; Khan, W.I. Mucins in Intestinal Mucosal Defense and Inflammation: Learning From Clinical and Experimental Studies. Front. Immunol. 2020, 11, 2054. [Google Scholar] [CrossRef] [PubMed]
- Mucin 2 [Homo Sapiens]. Available online: https://www.ncbi.nlm.nih.gov/protein/AZL49145.1 (accessed on 1 September 2023).
- Protein BLAST. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastp&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome (accessed on 1 September 2023).
- Portaccio, M.; Faramarzi, B.; Lepore, M. Probing Biochemical Differences in Lipid Components of Human Cells by Means of ATR-FTIR Spectroscopy. Biophysica 2023, 3, 524–538. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of aNPs | Size (nm) | PDI | Relative NP Concentration * | Protein Content (mg/mL) |
|---|---|---|---|---|
| C. chamissoi | 605 ± 67 | 0.22 ± 0.02 | 1.5 ± 0.3 | 0.04 ± 0.01 |
| G. gracilis | 232 ± 13 | 0.24 ± 0.02 | 9.1 ± 1.2 | 0.07 ± 0.001 |
| C. crispus | 275 ± 24 | 0.33 ± 0.03 | 4.1 ± 1.0 | 0.01 ± 0.001 |
| U. pinnatifida | 466 ± 18 | 0.29 ± 0.01 | 6.4 ± 0.5 | 0.44 ± 0.01 |
| S. muticum | 170 ± 07 | 0.24 ± 0.01 | 28.2 ± 9.9 | 0.68 ± 0.06 |
| L. digitata | 212 ± 20 | 0.27 ± 0.01 | 12.4 ± 7.8 | 0.11 ± 0.001 |
| L. japonica | 217 ± 10 | 0.19 ± 0.02 | 40.0 ± 8.7 | 0.68 ± 0.001 |
| E. cava | 235 ± 12 | 0.19 ± 0.01 | 27.3 ± 8.0 | 0.81 ± 0.01 |
| H. pluvialis | 251 ± 07 | 0.21 ± 0.01 | 30.2 ± 3.9 | 3.24 ± 0.02 |
| M. pyrifera | 252 ± 20 | 0.29 ± 0.03 | 6.7 ± 1.0 | 0.15 ± 0.02 |
| P. palmata | 245 ± 33 | 0.44 ± 0.12 | 7.1 ± 0.4 | 0.77 ± 0.09 |
| G. skottsbergii | 381 ± 13 | 0.48 ± 0.01 | 3.6 ± 0.2 | 0.01 ± 0.01 |
| A. platensis | 126 ± 02 | 0.14 ± 0.001 | 60.8 ± 2.9 | 2.61 ± 0.81 |
| C. vulgaris | 157 ± 11 | 0.25 ± 0.05 | 5.1 ± 0.5 | 0.80 ± 0.04 |
| Animal | Sequence ID | Length [aa] | Identities [%] | Positives [%] | Gaps [%] | Query Cover [%] | Score |
|---|---|---|---|---|---|---|---|
| Mouse | NP_076055.4 | 4576 | 80 | 87 | 0 | 43 | 2532 |
| Sheep | XP_042093899.1 | 5972 | 49 | 63 | 3 | 87 | 1495 |
| Pig | XP_020938146.1 | 5759 | 49 | 63 | 3 | 60 | 1474 |
| Type of aNPs | EE |
|---|---|
| A. platensis | 47% |
| L. japonica | 55% |
| L. digitata | 40% |
| S. muticum | 32% |
| G. gracilis | 35% |
| C. chamissoi | 34% |
| Type of aNP | MFIF 1:1 | MFIF 100:1 | Uptake 1:1 | Uptake 100:1 |
|---|---|---|---|---|
| Free FITC | 228 | 228 | 0.59% | 0.59% |
| A. platensis | 73,629 | 207996 | 100% | 100% |
| L. japonica | 20,984 | 14238 | 100% | 100% |
| L. digitata | 24,576 | 17838 | 100% | 100% |
| S. muticum | 229 | 212 | 0.59% | 0.54% |
| G. gracilis | 229 | 17103 | 0.59% | 100% |
| C. chamissoi | 255 | 270 | 0.65% | 0.69% |
| Common Name | Botanical Name | Catalog # | Group-Type |
|---|---|---|---|
| Chondracanthus chamissoi | Chondracanthus chamissoi | ncchchpwd | Red macroalgae |
| Gracilaria | Gracilaria gracilis | FRX730 | Red macroalgae |
| Irish moss | Chondrus crispus | FRX853 | Red macroalgae |
| Wakame | Undaria pinnatifida | FRX1419 | Brown macroalgae |
| Sargassum seaweed | Sargassum muticum | ncsasepwd | Brown macroalgae |
| Kelp laminaria digitata | Laminara digitata | FRX874PAC | Brown macroalgae |
| Kombu | Laminaria japonica | FRX886 | Brown macroalgae |
| Ecklonia cava | Ecklonia cava | FRX3544 | Brown macroalgae |
| Astaxanthin | Haematococcus pluvialis | FRX177 | Red microalgae |
| Giant kelp | Macrocytis pyrifera | ncgikepwd | Brown macroalgae |
| Dulse | Palmaria palmata | FRX576 | Red macroalgae |
| Gigartina red marine | Gigartina skottsbergii | FRX680 | Red macroalgae |
| Spirulina | Arthospira platensis | FRX1333 | Green microalgae |
| Chlorella vulgaris | Chlorella vulgaris | FRX454 | Green microalgae |
| Type of Algae | Initial Mass * (g) |
|---|---|
| C. chamissoi | 2.0 |
| G. gracilis | 2.0 |
| C. crispus | 0.5 |
| U. pinnatifida | 2.0 |
| S. muticum | 2.0 |
| L. digitata | 2.0 |
| L. japonica | 2.0 |
| E. cava | 2.0 |
| H. pluvialis | 2.0 |
| M. pyrifera | 2.0 |
| P. palmata | 2.0 |
| G. skottsbergii | 0.3 |
| A. platensis | 0.2 |
| C. vulgaris | 0.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drori, E.; Patel, D.; Coopersmith, S.; Rahamim, V.; Drori, C.; Jadhav, S.S.; Avital, R.; Anker, Y.; Azagury, A. Algae-Based Nanoparticles for Oral Drug Delivery Systems. Mar. Drugs 2024, 22, 98. https://doi.org/10.3390/md22030098
Drori E, Patel D, Coopersmith S, Rahamim V, Drori C, Jadhav SS, Avital R, Anker Y, Azagury A. Algae-Based Nanoparticles for Oral Drug Delivery Systems. Marine Drugs. 2024; 22(3):98. https://doi.org/10.3390/md22030098
Chicago/Turabian StyleDrori, Eliyahu, Dhaval Patel, Sarah Coopersmith, Valeria Rahamim, Chen Drori, Suchita Suryakant Jadhav, Roni Avital, Yaakov Anker, and Aharon Azagury. 2024. "Algae-Based Nanoparticles for Oral Drug Delivery Systems" Marine Drugs 22, no. 3: 98. https://doi.org/10.3390/md22030098
APA StyleDrori, E., Patel, D., Coopersmith, S., Rahamim, V., Drori, C., Jadhav, S. S., Avital, R., Anker, Y., & Azagury, A. (2024). Algae-Based Nanoparticles for Oral Drug Delivery Systems. Marine Drugs, 22(3), 98. https://doi.org/10.3390/md22030098