
Possible Mechanisms of Stiffness Changes Induced by Stiffeners and Softeners in Catch Connective Tissue of Echinoderms
Abstract
1. Introduction
2. Tensilin
2.1. Tensilins Purified from the Dermis of Sea Cucumbers
2.2. Genes of Tensilin and Related Protein from Sea Cucumbers and Other Echinoderms

{kind=link}
{kind=link}
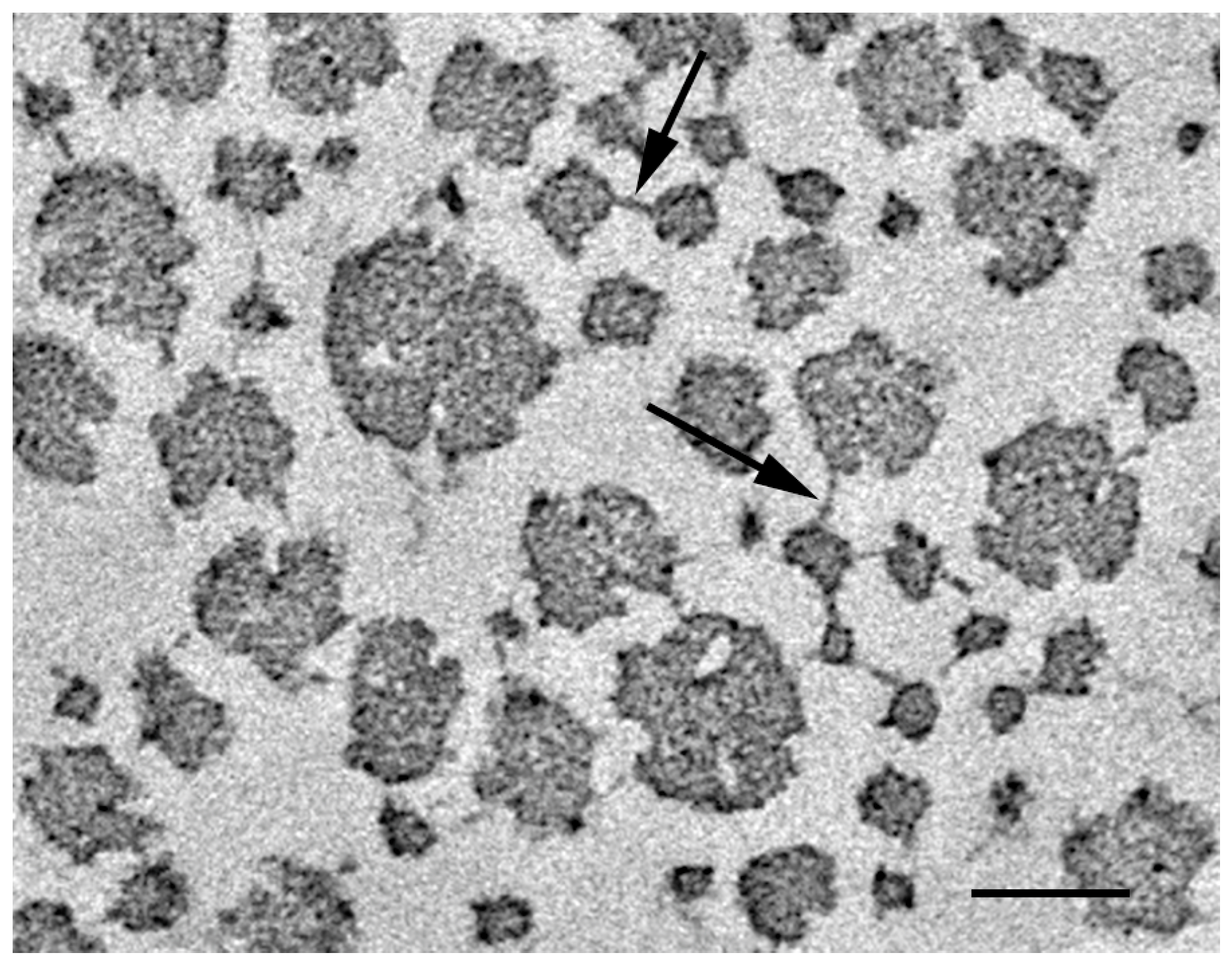{kind=link}
| Species | Body Parts (Protein or mRNA) | * Stiffening | Note | NCBI Accession | Reference |
|---|---|---|---|---|---|
| Cucumaria frondosa | Inner dermis (protein and mRNA) | + | AAK61535 | [17] | |
| Holothuria leucospilota | Dermis (protein) | + | Not yet cloned | [18] | |
| Apostichopus japonicus | Not shown (mRNA) | ND | ALD83456 | [33] | |
| Holothuria forskali | Cuvierian tubules (mRNA) | ND | Possible stiffener | AQR59058 | [29] |
| Dermis (mRNA) | + (** Recombinant) | MZ561455 | [19] | ||
| Cladolabes schmeltzii | Body wall (mRNA) | ND | GFWR01009215 | [27] | |
| ND | GFWR01009682 | ||||
| Eupentacta faudatrix | Regenerating gut (mRNA) | ND | Possible inhibitor of MMP16 (tensilin3) | GHCL01023186 | [31] |
2.3. Recombinant Tensilin and Related Protein
3. Novel Stiffening Factor
4. Softenin and Other Softeners
5. Ultrastructure of the Sea Cucumber Dermis
5.1. Extracellular Matrix
5.2. Tensilin-Containing Cell
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Motokawa, T. Connective tissue catch in echinoderms. Biol. Rev. 1984, 59, 255–270. [Google Scholar] [CrossRef]
- Wilkie, I.C. Mutable collagenous tissue: Extracellular matrix as mechano-effector. In Echinoderm Studies; Jangoux, M., Lawrence, J.M., Eds.; A.A. Balkema: Rotterdam, The Netherlands, 1996; Volume 5, pp. 61–102. [Google Scholar]
- Wilkie, I.C. Mutable collagenous tissue: Overview and biotechnological perspective. Prog. Mol. Subcell. Biol. 2005, 39, 221–250. [Google Scholar]
- Wilkie, I.C.; Sugni, M.; Gupta, H.S.; Candia Carnevali, M.D.; Elphick, M.R. The mutable collagenous tissue of echinoderms: From biology to biomedical applications. In Soft Matter for Biomedical Applications; Azeveno, H.S., Mano, J.F., Borges, J., Eds.; The Royal Society of Chemistry: Cambridge, UK; London, UK, 2021; pp. 3–33. [Google Scholar]
- Thurmond, F.A.; Trotter, J.A. Morphology and biomechanics of the microfibrillar network of sea cucumber dermis. J. Exp. Biol. 1996, 199, 1817–1828. [Google Scholar] [CrossRef]
- Thurmond, F.A.; Koob, T.J.; Bowness, J.M.; Trotter, J.A. Partial biochemical and immunologic characterization of fibrillin microfibrils from sea cucumber dermis. Connect. Tissue Res. 1997, 36, 211–222. [Google Scholar] [CrossRef]
- Takahashi, K. The catch apparatus of the sea-urchin spine II. Response to stimuli. J. Fac. Sci. Univ. Tokyo Sect. IV 1967, 11, 121–130. [Google Scholar]
- Motokawa, T. The stiffness change of the holothurian dermis caused by chemical and electrical stimulation. Comp. Biochem. Physiol. 1981, 70, 41–48. [Google Scholar] [CrossRef]
- Motokawa, T. Viscoelasticity of holothurian body wall. J. Exp. Biol. 1984, 109, 63–75. [Google Scholar] [CrossRef]
- Motokawa, T.; Hayashi, Y. Calcium dependence of viscosity change caused by cations in holothurian catch connective tissue. Comp. Biochem. Physiol. 1987, 87, 579–582. [Google Scholar]
- Trotter, J.A.; Koob, T.J. Evidence that calcium-dependent cellular processes are involved in the stiffening response of holothurian dermis and that dermal cells contain an organic stiffening factor. J. Exp. Biol. 1995, 198, 1951–1961. [Google Scholar] [CrossRef]
- Trotter, J.A.; Chino, K. Regulation of cell-dependent viscosity in the dermis of the sea cucumber Actinopyga agassizi. Comp. Biochem. Physiol. 1997, 118, 805–811. [Google Scholar] [CrossRef]
- Koob, T.J.; Koob-Emunds, M.M.; Trotter, J.A. Cell-derived stiffening and plasticizing factors in sea cucumber (Cucumaria frondosa) dermis. J. Exp. Biol. 1999, 202, 2291–2301. [Google Scholar] [CrossRef]
- Szulgit, G.K.; Shadwick, R.E. Dynamic mechanical characterization of a mutable collagenous tissue: Response of sea cucumber dermis to cell lysis and dermal extracts. J. Exp. Biol. 2000, 203, 1539–1550. [Google Scholar] [CrossRef]
- Motokawa, T.; Tsuchi, A. Dynamic mechanical properties of body-wall dermis in various mechanical states and their implications for the behavior of sea cucumbers. Biol. Bull. 2003, 205, 261–275. [Google Scholar] [CrossRef]
- Mo, J.; Prévost, S.F.; Blowes, L.M.; Egertová, M.; Terrill, N.J.; Wang, W.; Elphick, M.R.; Gupta, H.S. Interfibrillar stiffening of echinoderm mutable collagenous tissue demonstrated at the nanoscale. Proc. Natl. Acad. Sci. USA 2016, 113, E6362–E6371. [Google Scholar] [CrossRef]
- Tipper, J.P.; Lyons-Levy, G.; Atkinson, M.A.L.; Trotter, J.A. Purification, characterization and cloning of tensilin, the collagen-fibril binding and tissue-stiffening factor from Cucumaria frondosa dermis. Matrix Biol. 2003, 21, 625–635. [Google Scholar] [CrossRef]
- Tamori, M.; Yamada, A.; Nishida, N.; Motobayashi, Y.; Oiwa, K.; Motokawa, T. Tensilin-like stiffening protein from Holothuria leucospilota does not induce the stiffest state of catch connective tissue. J. Exp. Biol. 2006, 209, 1594–1602. [Google Scholar] [CrossRef]
- Bonneel, M.; Hennebert, E.; Aranko, A.S.; Hwang, D.S.; Lefevre, M.; Pommier, V.; Wattiez, R.; Delroisse, J.; Flammang, P. Molecular mechanisms mediating stiffening in the mechanically adaptable connective tissues of sea cucumbers. Matrix Biol. 2022, 108, 39–54. [Google Scholar] [CrossRef]
- Takehana, Y.; Yamada, A.; Tamori, M.; Motokawa, T. Softenin, a novel protein that softens the connective tissue of sea cucumbers through inhibiting interaction between collagen fibrils. PLoS ONE 2014, 9, e85644. [Google Scholar] [CrossRef]
- Matsuno, A.; Motokawa, T. Evidence for calcium translocation in catch connective tissue of the sea cucumber Stichopus chloronotus. Cell Tissue Res. 1992, 267, 307–312. [Google Scholar] [CrossRef]
- Birenheide, R.; Tamori, M.; Motokawa, T.; Ohtani, M.; Iwakoshi, E.; Muneoka, Y.; Fujita, T.; Minakata, H.; Nomoto, K. Peptides controlling stiffness of connective tissue in sea cucumbers. Biol. Bull. 1998, 194, 253–259. [Google Scholar] [CrossRef]
- Angerer, L.; Husssain, S.; Wei, Z.; Livingston, B.T. Sea urchin metalloproteases: A genomic survey of the BMP-1/tolloid-like, MMP and ADAM families. Dev. Biol. 2006, 300, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Tricarico, S.; Barbaglio, A.; Burlini, N.; Del Giacco, L.; Ghilardi, A.; Suguni, M.; Di Benedetto, C.; Bonasoro, F.; Wilkie, I.C.; Candia Carnevali, M.D. New insights into the mutable collagenous tissue of Paracentrotus lividus: Preliminary results. Zoosymposia 2012, 7, 279–285. [Google Scholar] [CrossRef]
- Wilkie, I.C.; Fassini, D.; Cullorà, E.; Barbaglio, A.; Tricarico, S.; Suguni, M.; Del Giacco, L.; Candia Carnevali, M.D. Mechanical properties of the compass depressors of the sea-urchin Paracentrotus lividus (Echinodermata, Echinoidea) and the effects of enzymes, neurotransmitters and synthetic tensilin-like protein. PLoS ONE 2015, 10, e0120339. [Google Scholar] [CrossRef] [PubMed]
- Clouse, R.M.; Linchangco, G.V., Jr.; Kerr, A.M.; Reid, R.W.; Janies, D.A. Phylotranscriptomic analysis uncovers a wealth of tissue inhibitor of metalloproteinases variants in echinoderms. R. Soc. Open Sci. 2015, 2, 150377. [Google Scholar] [CrossRef]
- Dolmatov, I.Y.; Afanasyev, S.V.; Boyko, A.V. Molecular mechanisms of fission in echinoderms: Transcription analysis. PLoS ONE 2018, 13, e0195836. [Google Scholar] [CrossRef]
- Zhang, X.; Sun, L.; Yuan, J.; Sun, Y.; Gao, Y.; Zhang, L.; Li, S.; Dai, H.; Hamel, J.-F.; Liu, C.; et al. The sea cucumber genome provides insights into morphological evolution and visceral regeneration. PLoS Biol. 2017, 15, e2003790. [Google Scholar] [CrossRef]
- Demeuldre, M.; Hennebert, E.; Bonneel, M.; Lengerer, B.; Van Dyck, S.; Wattiez, R.; Ladurner, P.; Flammang, P. Mechanical adaptability of sea cucumber Cuvierian tubules involves a mutable collagenous tissue. J. Exp. Biol. 2017, 220, 2108–2119. [Google Scholar] [CrossRef]
- Dolmatov, I.Y.; Nizhnichenko, V.A.; Dolmatova, L.S. Matrix metalloproteinases and tissue inhibitors of metalloproteinases in echinoderms: Structure and possible functions. Cells 2021, 10, 2331. [Google Scholar] [CrossRef]
- Dolmatov, I.Y.; Kalacheva, N.V.; Tkacheva, E.S.; Shulga, A.P.; Zavalnaya, E.G.; Shamshurina, E.V.; Girich, A.S.; Boyko, A.V.; Eliseikina, M.G. Expression of Piwi, MMP, TIMP, and Sox during gut regeneration in holothurian Eupentacta faudatrix (Holothuroidea, Dendrochirotida). Genes 2021, 12, 1292. [Google Scholar] [CrossRef]
- Lv, Z.; Han, G.; Li, C. Tissue inhibitor of metalloproteinases 1 is involved in ROS-mediated inflammation via regulating matrix metalloproteinase 1 expression in the sea cucumber Apostichopus japonicus. Dev. Comp. Immunol. 2022, 127, 104298. [Google Scholar] [CrossRef]
- GenBank. 2015. Available online: https://www.ncbi.nim.nih.gov/protein/ALD83456.1 (accessed on 12 January 2023).
- Wilkie, I.C.; Candia Carnevali, M.D.; Andrietti, F. Variable tensility of the peristomial membrane of the sea-urchin Paracentrotus lividus (Lamarck). Comp. Biochem. Physiol. 1993, 105, 493–501. [Google Scholar] [CrossRef]
- Yamada, A.; Tamori, M.; Iketani, T.; Oiwa, K.; Motokawa, T. A novel stiffening factor inducing the stiffest state of holothurian catch connective tissue. J. Exp. Biol. 2010, 213, 3416–3422. [Google Scholar] [CrossRef] [PubMed]
- Iwakoshi, E.; Ohtani, M.; Takahashi, T.; Muneoka, Y.; Ikeda, T.; Fujita, T.; Minakata, H.; Nomoto, K. Comparative aspects of structure and action of bioactive peptides isolated from the sea cucumber Stichopus japonicus. In Peptide Chemistry 1994; Ohno, M., Ed.; Protein Research Foundation: Osaka, Japan, 1995; pp. 261–264. [Google Scholar]
- Elphick, M.R. The protein precursors of peptides that affect the mechanics of connective tissue and/or muscle in the echinoderm Apostichopus japonicus. PLoS ONE 2012, 7, e44492. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.-L.; Hu, Y.-Q.; Shen, J.-D.; Cai, Q.-F.; Liu, G.-M.; Su, W.-J.; Cao, M.-J. Identification of a novel gelatinolytic metalloproteinase (GMP) in the body wall of sea cucumber (Stichopus japonicus) and its involvement in collagen degradation. Process Biochem. 2013, 48, 871–877. [Google Scholar] [CrossRef]
- Junqueira, L.C.U.; Montes, G.S.; Murão, P.A.S.; Carneiro, J.; Salles, L.M.M.; Bonetti, S.S. Collagen–proteoglycans interaction during autotomy in the sea cucumber Stichopus badionotus. Rev. Can. Biol. 1980, 39, 157–164. [Google Scholar]
- Tamori, M.; Ishida, K.; Matsuura, E.; Ogasawara, K.; Hanasaka, T.; Takehana, Y.; Motokawa, T.; Osawa, T. Ultrastructural changes associated with reversible stiffening in catch connective tissue of sea cucumbers. PLoS ONE 2016, 11, e0155673. [Google Scholar] [CrossRef]
- Holland, N.D.; Grimmer, J.C. Fine structure of the cirri and a possible mechanism for their motility in stalkless crinoids (Echinodermata). Cell Tissue Res. 1981, 214, 207–217. [Google Scholar] [CrossRef]
- Holland, N.D.; Grimmer, J.C. Fine structure of syzygial articulations before and after arm autotomy in Florometra serratissima (Echinodermata: Crinoidea). Zoomorphology 1981, 98, 169–183. [Google Scholar] [CrossRef]
- Motokawa, T. Fine structure of the dermis of the body wall of the sea cucumber, Stichopus chloronotus, a connective tissue which changes its mechanical properties. Galaxea 1982, 1, 55–64. [Google Scholar]
- Wilkie, I.C. Design for disaster: The ophiuroid intervertebral ligament as a typical mutable collagenous structure. In Echinoderm Biology; Burke, R.D., Mladenov, P.V., Lambert, P., Parsley, R.L., Eds.; A.A. Balkema: Rotterdam, The Netherlands, 1988; pp. 25–38. [Google Scholar]
- Del Castillo, J.; Smith, D.S.; Vidal, A.M.; Sierra, C. Catch in the primary spine of the sea urchin Eucidaris tribuloides: A brief review and a new interpretation. Biol. Bull. 1995, 188, 120–127. [Google Scholar] [CrossRef]
- Barbaglio, A.; Tricarico, S.; Ribeiro, A.; Ribeiro, C.; Sugni, M.; Di Benedetto, C.; Wilkie, I.; Barbosa, M.; Bonasoro, F.; Candia Carnevali, M.D. The mechanically adaptable connective tissue of echinoderms: Its potential for bio-innovation in applied technology and ecology. Mar. Environ. Res. 2012, 76, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Tamori, M.; Takemae, C.; Motokawa, T. Evidence that water exudes when holothurian connective tissue stiffens. J. Exp. Biol. 2010, 213, 1960–1966. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.R.; Barbaglio, A.; Benedetto, C.D.; Ribeiro, C.C.; Wilkie, I.C.; Candia Carnevali, M.D.; Barbosa, M.A. New insights into mutable collagenous tissue: Correlation between the microstructure and mechanical state of a sea-urchin ligament. PLoS ONE 2011, 6, e24822. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.R.; Barbaglio, A.; Oliveira, M.J.; Santos, R.; Coelho, A.V.; Ribeiro, C.C.; Wilkie, I.C.; Candia Carnevali, M.D.; Barbosa, M.A. Correlation between the biochemistry and mechanical states of a sea-urchin ligament: A mutable collagenous structure. Biointerphases 2012, 7, 38. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, I.C. The juxtaligamental cells of Ophiocomina nigra (Abildgaard) (Echinodermata: Ophiuroidea) and their possible role in mechano-effector function of collagenous tissue. Cell Tissue Res. 1979, 197, 515–530. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.S.; Wainwright, S.A.; Baker, J.; Cayer, M.L. Structural features associated with movement and ‘catch’ of sea-urchin spines. Tissue Cell 1981, 13, 299–320. [Google Scholar] [CrossRef]
- Hidaka, M.; Takahashi, K. Fine structure and mechanical properties of the catch apparatus of the sea-urchin spine, a collagenous connective tissue with muscle-like holding capacity. J. Exp. Biol. 1983, 103, 1–14. [Google Scholar] [CrossRef]
- Welsch, U.; Lange, A.; Bals, R.; Heinzeller, T. Juxtaligamental cells in feather stars and isocrinids. In Echinoderm Research 1995; Emson, R.H., Smith, A.B., Campbell, A.C., Eds.; A.A. Balkema: Rotterdam, The Netherlands, 1995; pp. 129–135. [Google Scholar]
- Charlina, N.A.; Dolmatov, I.Y.; Wilkie, I.C. Juxtaligamental system of the disc and oral frame of the ophiuroid Amphipholis kochii (Echinodermata: Ophiuroidea) and its role in autotomy. Invertebr. Biol. 2009, 128, 145–156. [Google Scholar] [CrossRef]
- Hennebert, E.; Haesaerts, D.; Dubois, P.; Flammang, P. Evaluation of the different forces brought into play during tube foot activities in sea stars. J. Exp. Biol. 2010, 213, 1162–1174. [Google Scholar] [CrossRef]
- Motokawa, T. Mechanical mutability in connective tissue of starfish body wall. Biol. Bull. 2011, 221, 280–289. [Google Scholar] [CrossRef]
- Wilkie, I.C. Functional morphology of the arm spine joint and adjacent structures of the brittlestar Ophiocomina nigra (Echinodermata, Ophiuroidea). PLoS ONE 2016, 11, e0167533. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamori, M.; Yamada, A. Possible Mechanisms of Stiffness Changes Induced by Stiffeners and Softeners in Catch Connective Tissue of Echinoderms. Mar. Drugs 2023, 21, 140. https://doi.org/10.3390/md21030140
Tamori M, Yamada A. Possible Mechanisms of Stiffness Changes Induced by Stiffeners and Softeners in Catch Connective Tissue of Echinoderms. Marine Drugs. 2023; 21(3):140. https://doi.org/10.3390/md21030140
Chicago/Turabian StyleTamori, Masaki, and Akira Yamada. 2023. "Possible Mechanisms of Stiffness Changes Induced by Stiffeners and Softeners in Catch Connective Tissue of Echinoderms" Marine Drugs 21, no. 3: 140. https://doi.org/10.3390/md21030140
APA StyleTamori, M., & Yamada, A. (2023). Possible Mechanisms of Stiffness Changes Induced by Stiffeners and Softeners in Catch Connective Tissue of Echinoderms. Marine Drugs, 21(3), 140. https://doi.org/10.3390/md21030140