

Dinophysis acuminata or Dinophysis acuta: What Makes the Difference in Highly Stratified Fjords?


 ,
,  , , ,
, , ,  , ,
, ,
Abstract
1. Introduction
2. Results
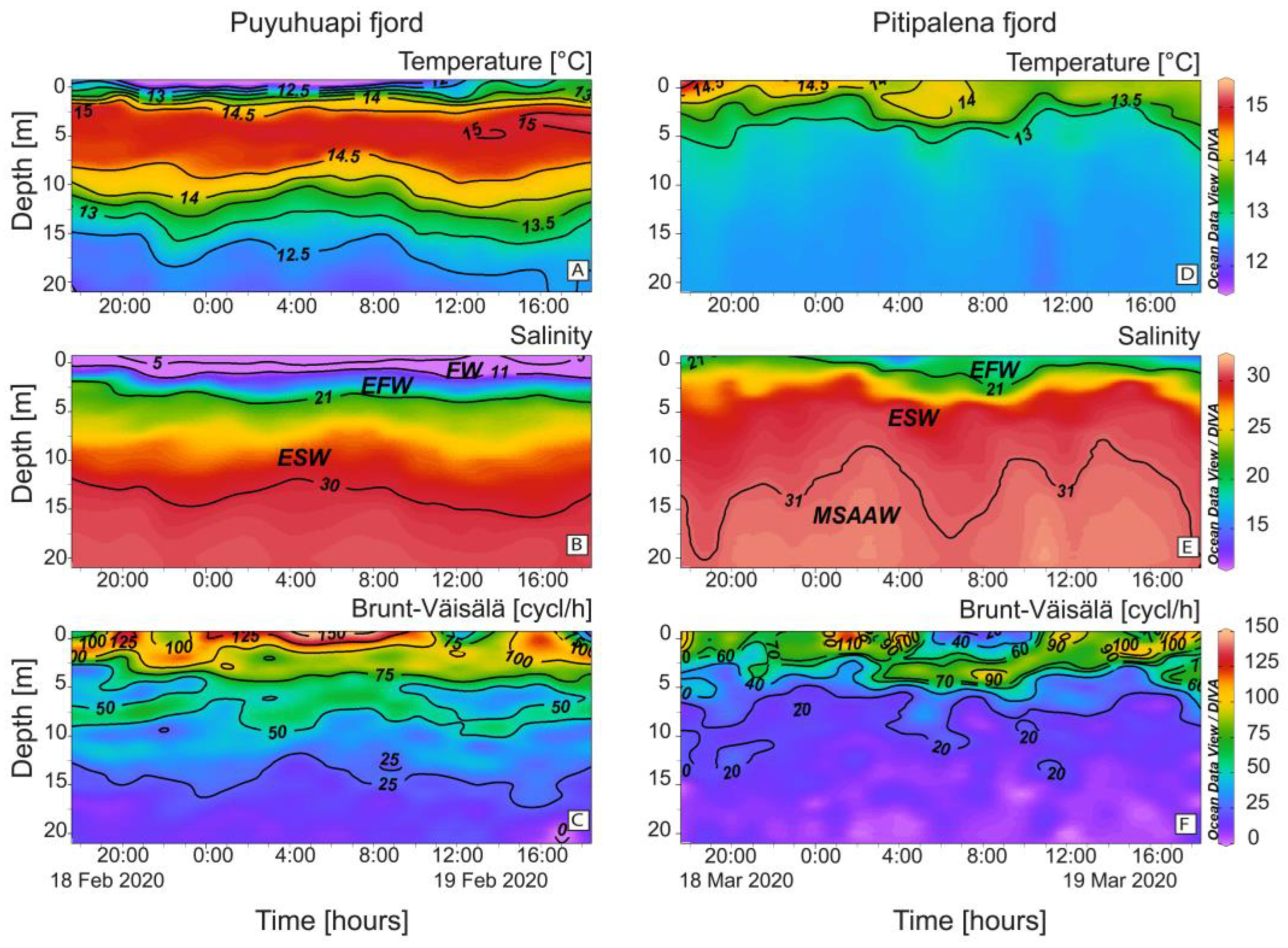
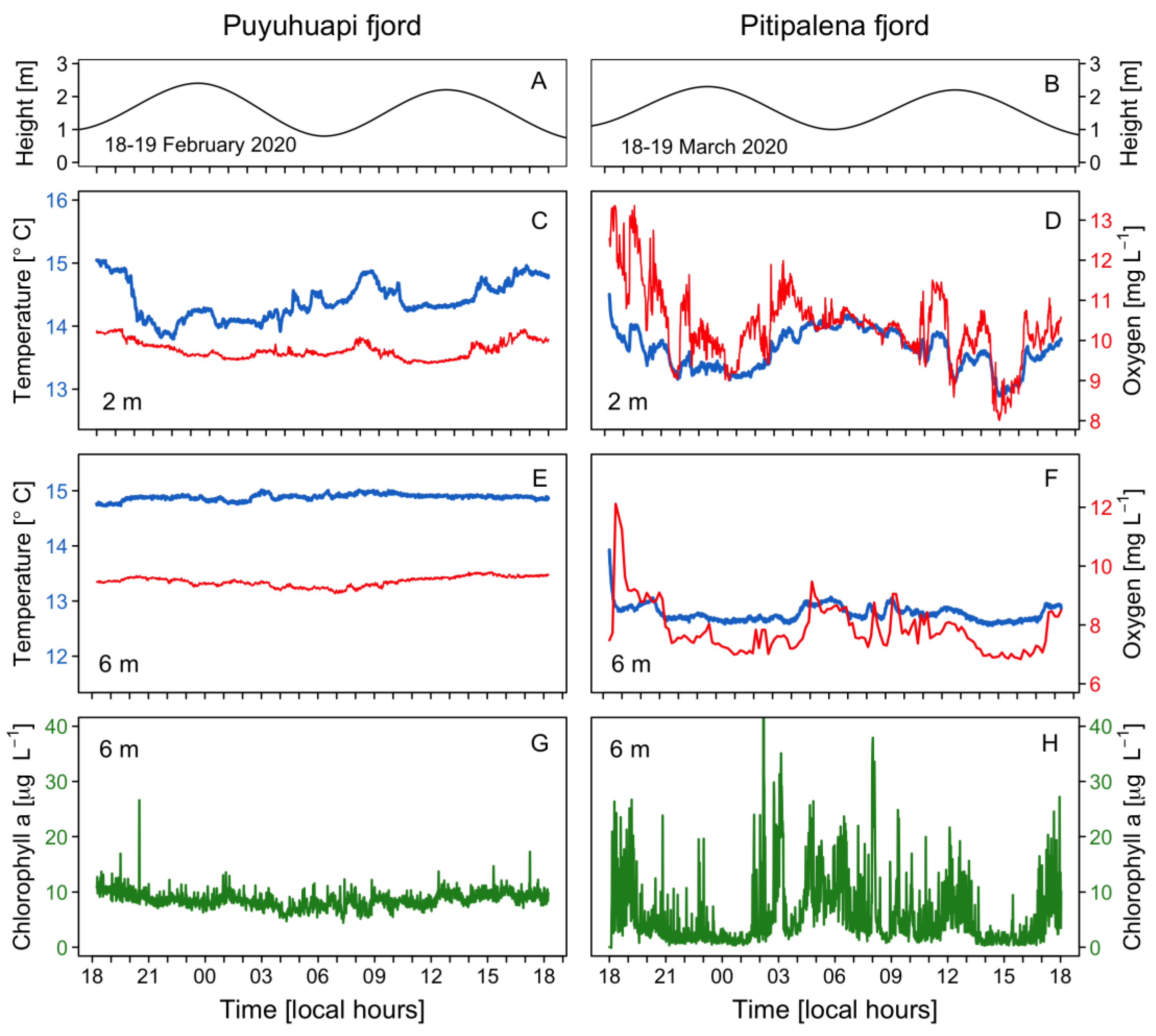
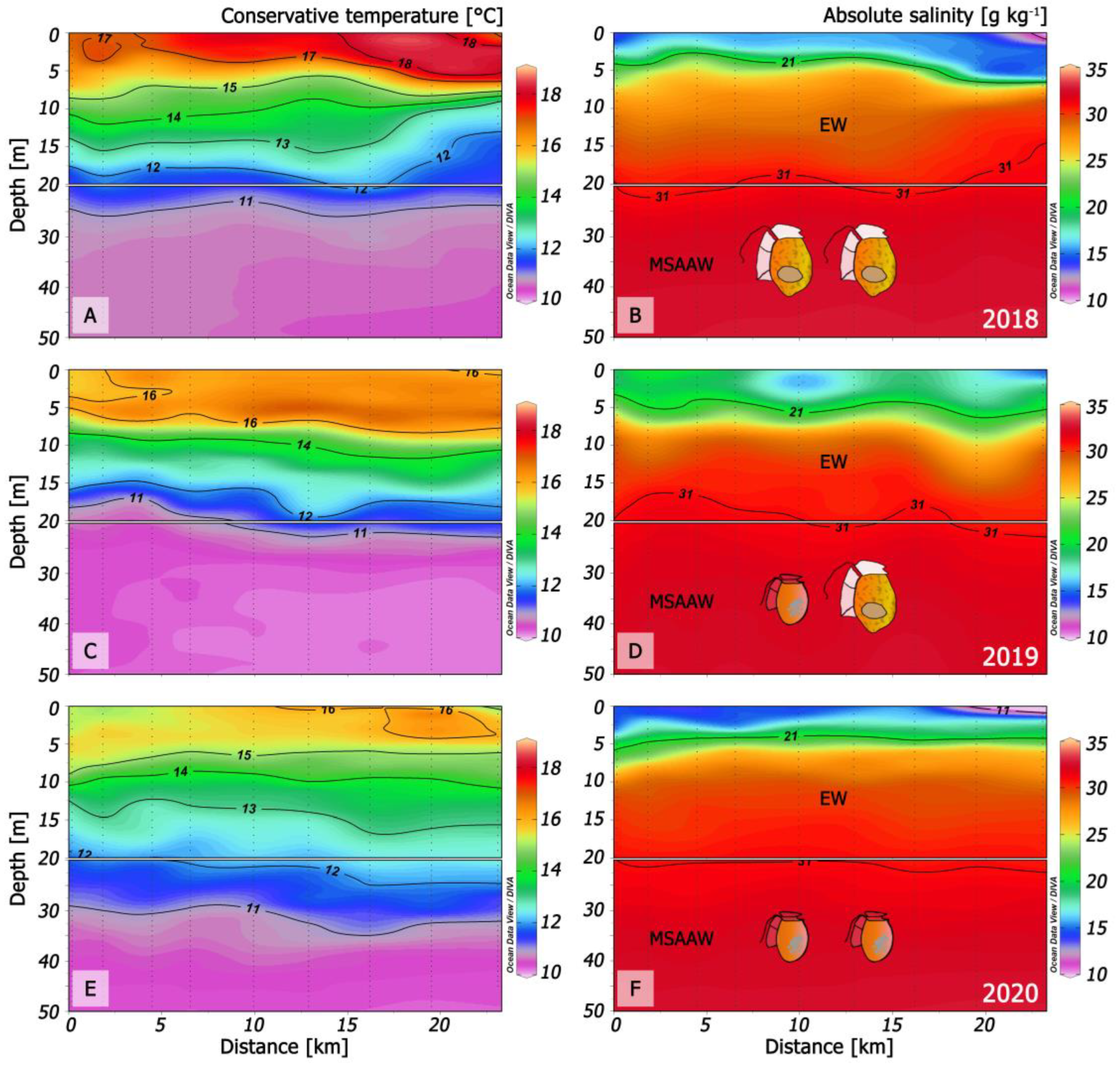
2.1. Vertical Distribution of Thermo-Haline Properties
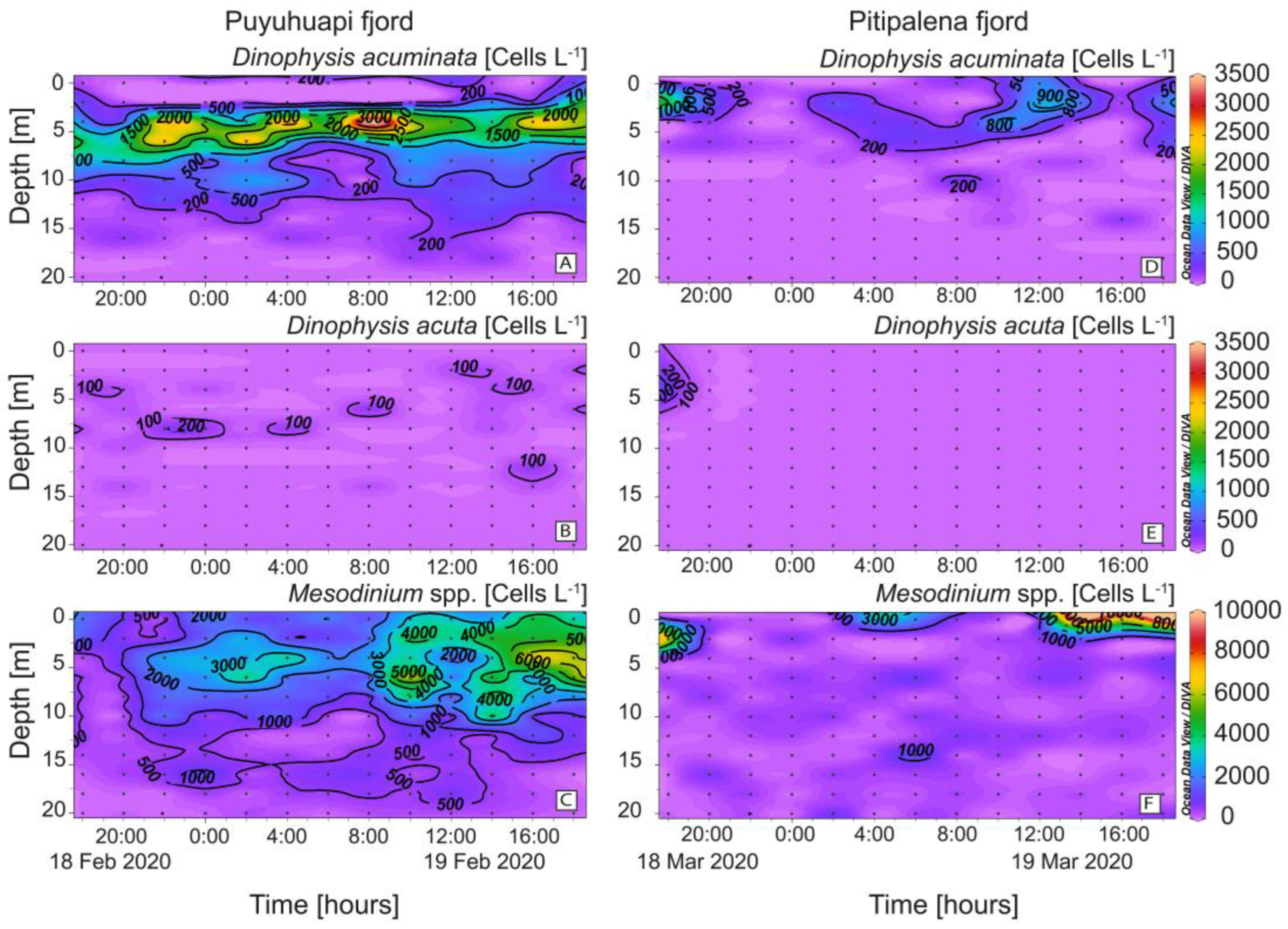
2.2. Distribution of Dinophysis acuminata, D. acuta, Mesodinium, and Other Observations
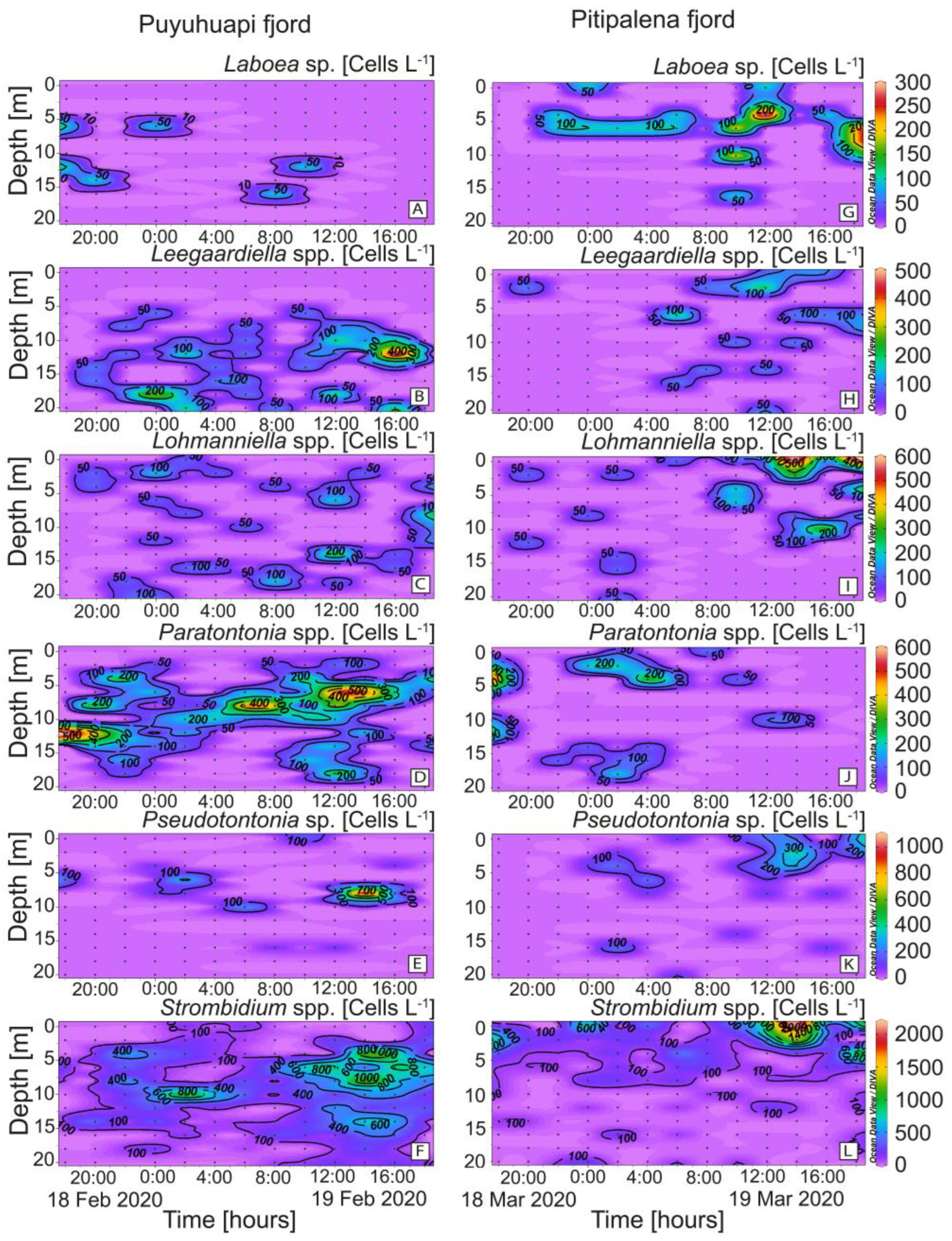
2.3. Distribution of Other Potential Ciliate Prey
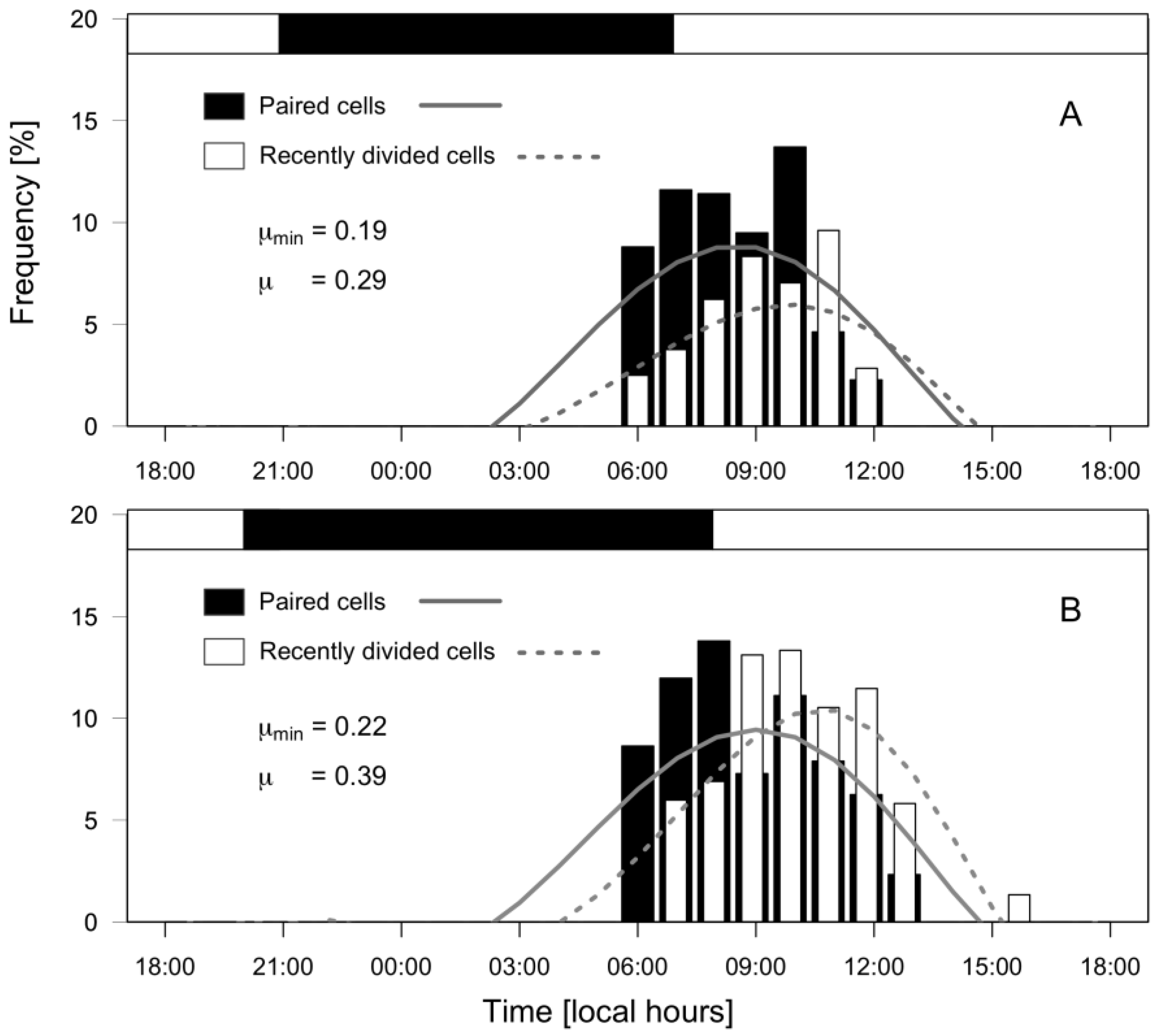
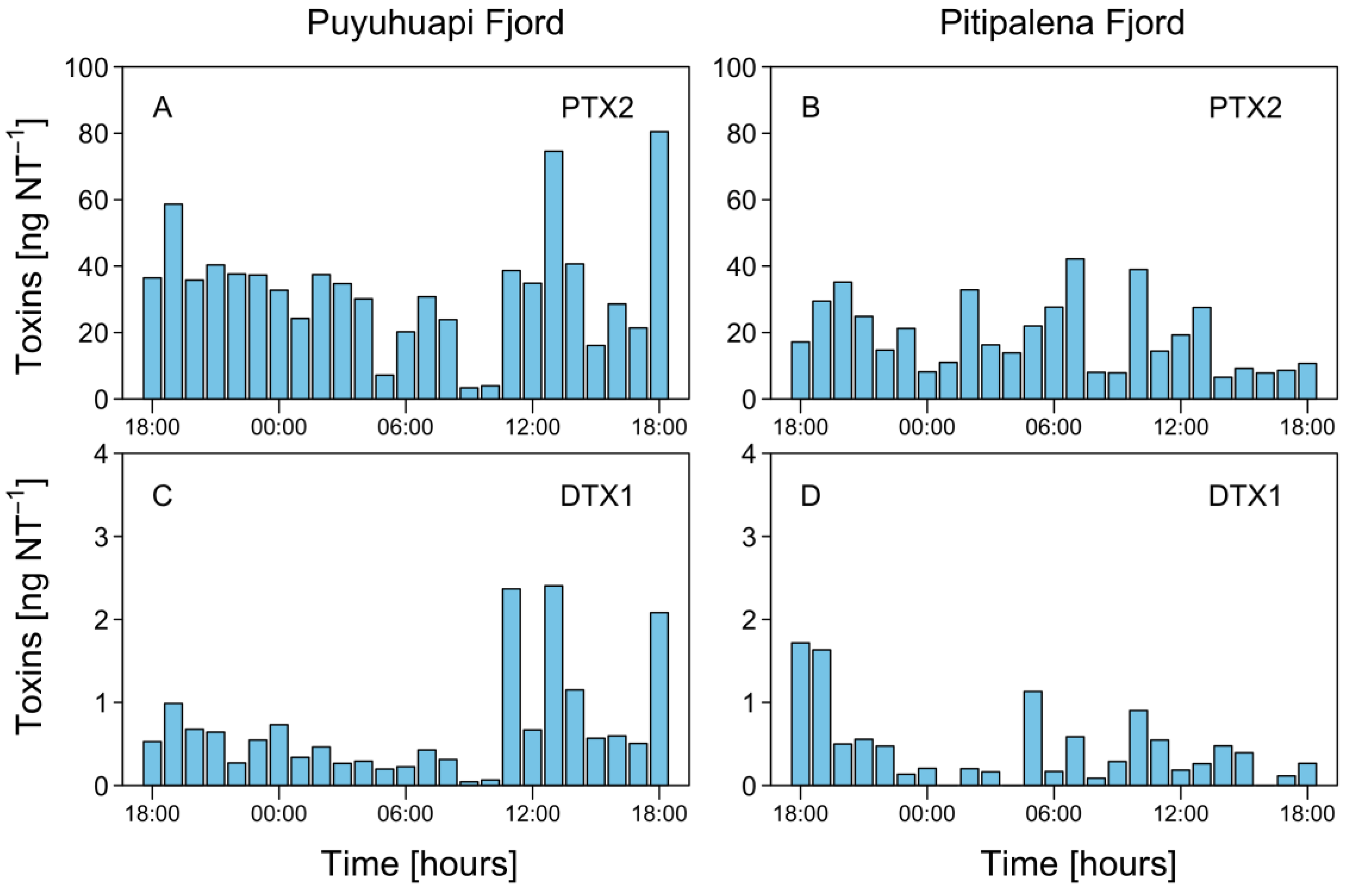
2.4. Physiological Status of Dinophysis acuminata: Division Rates and Cellular Toxin Content
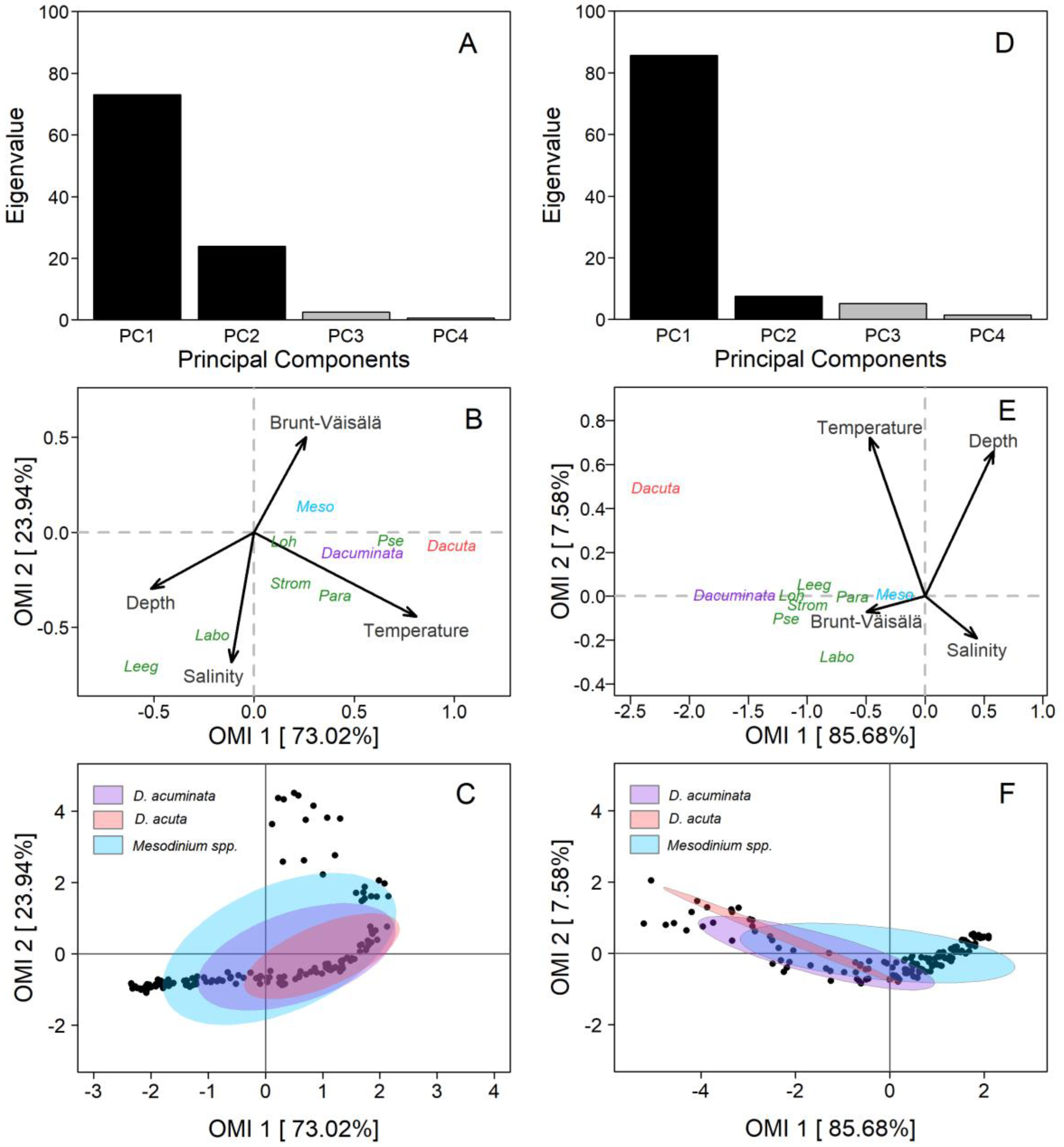
2.5. Niche Analysis
3. Discussion
3.1. Oceanographic Settings That Promote D. acuminata/D. acuta Dominance
3.2. Toxin Diversity
3.3. Niche Analysis
4. Materials and Methods
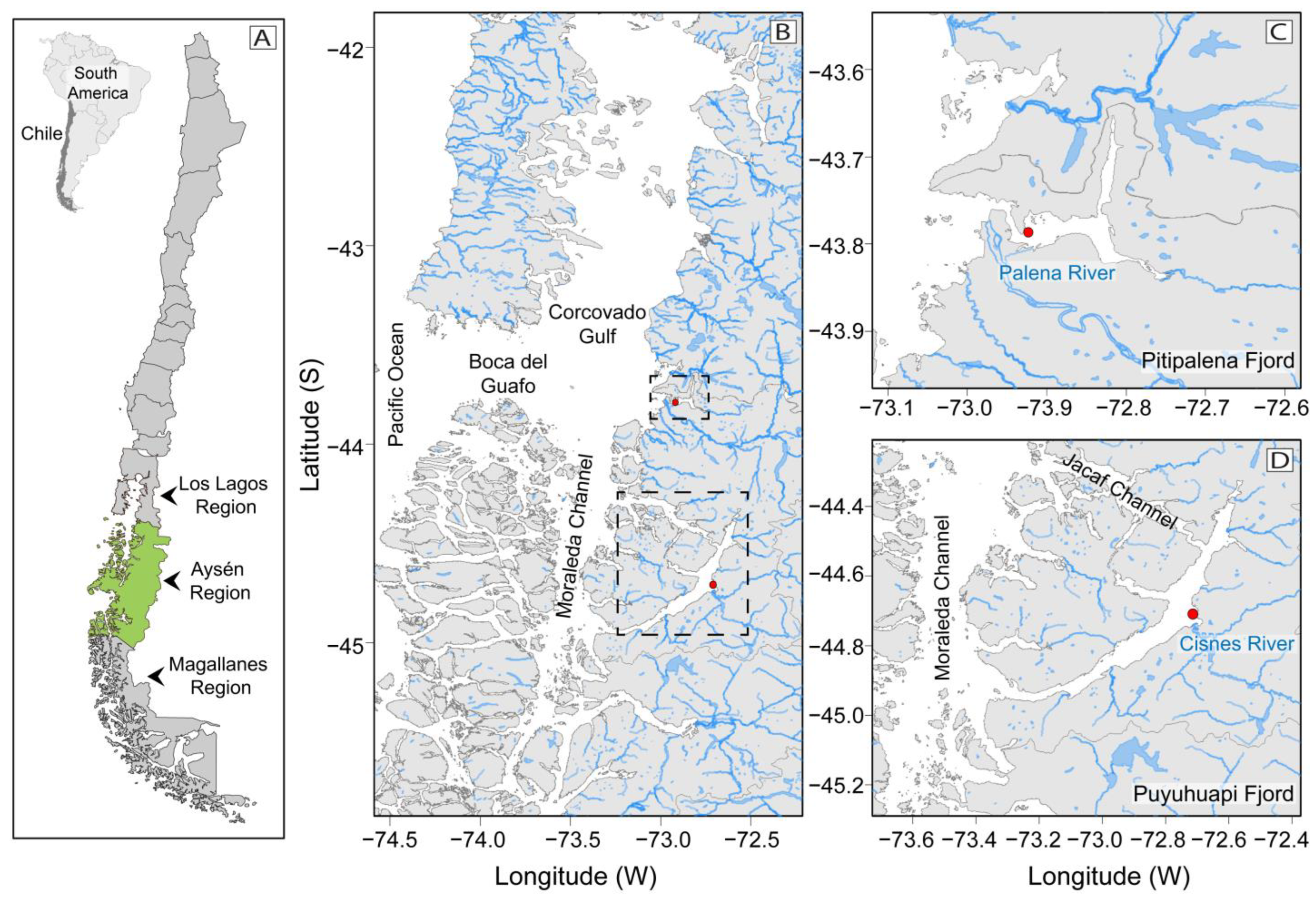
4.1. Study Area
4.2. Field Sampling
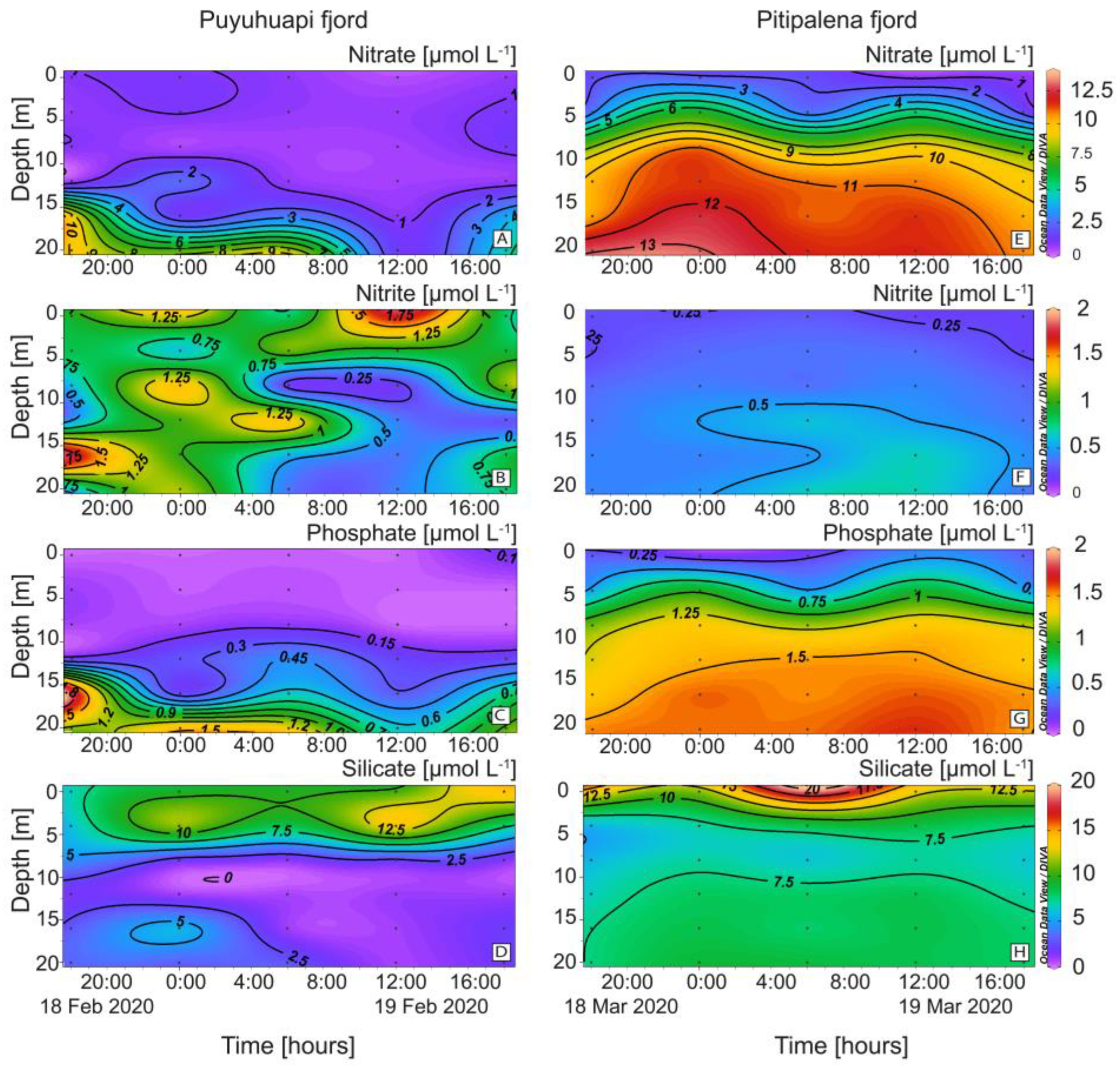
4.3. Nutrients
4.4. Phytoplankton Analysis
4.5. Division Rates
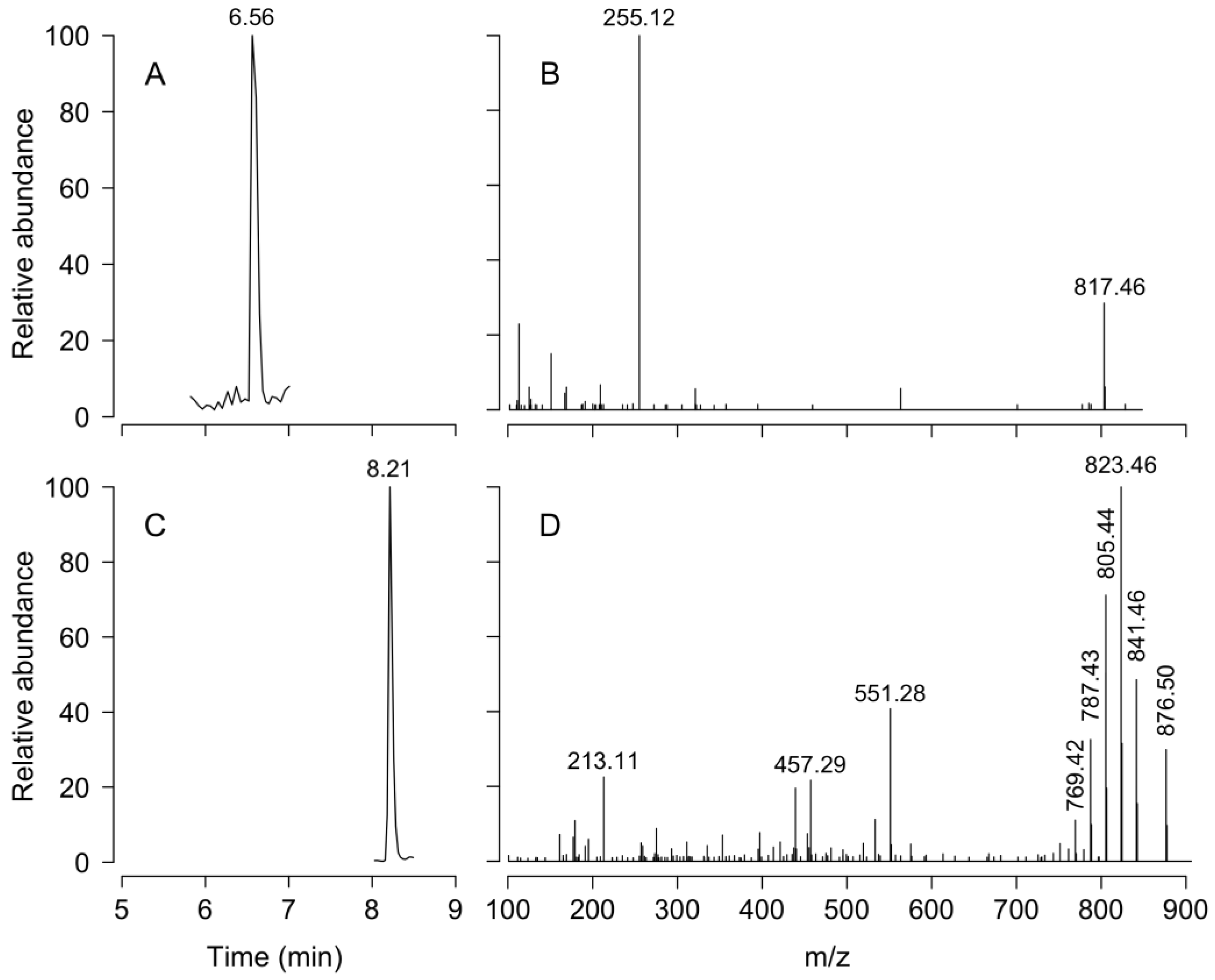
4.6. Lipophilic Toxin Analysis
4.6.1. Toxins Sample Extraction
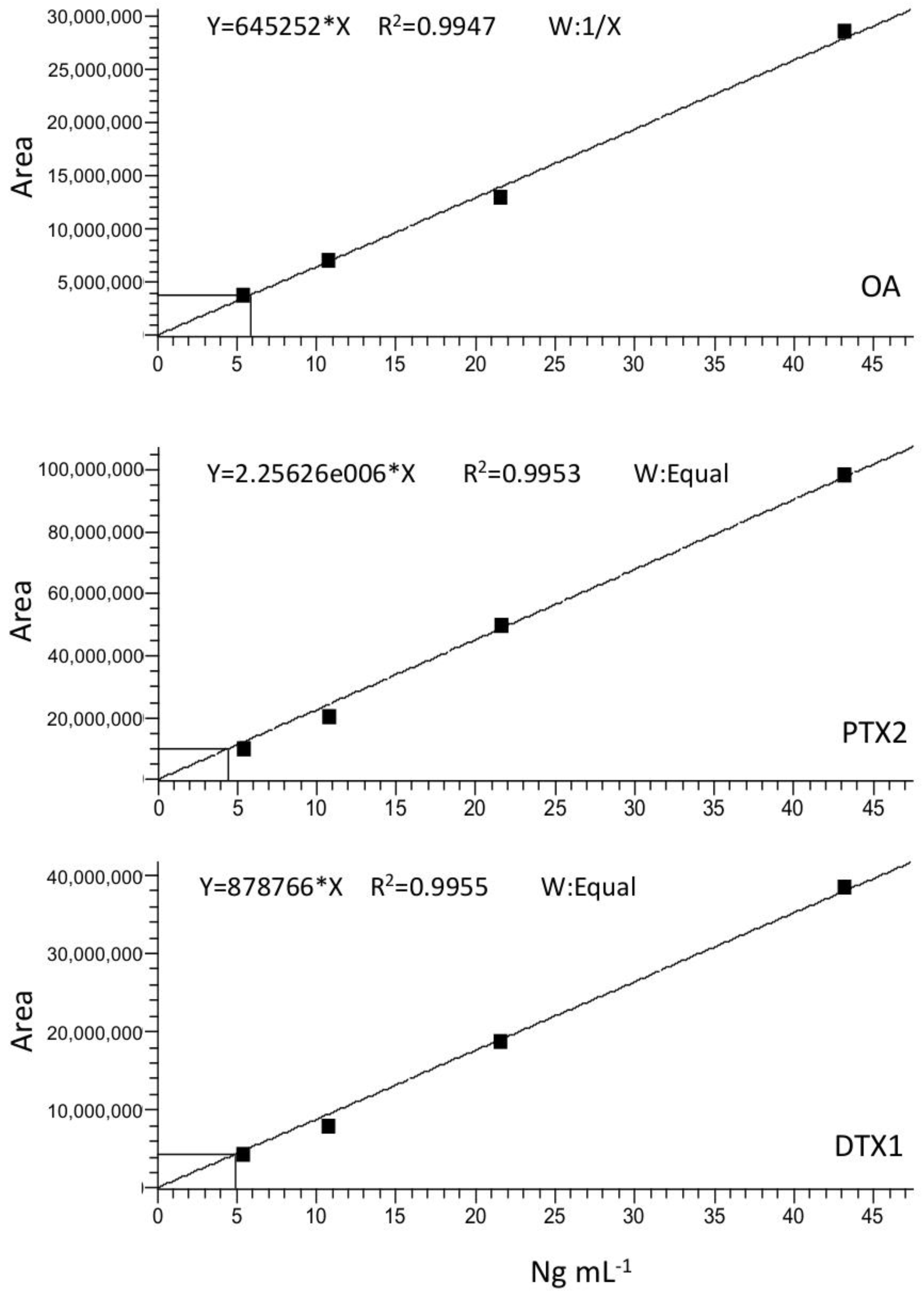
4.6.2. Toxin Detection and Quantification
4.7. Niche Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Appendix A


References
- Dickey, T.D. The emergence of concurrent high resolution physical and bio-optical measurements in the upper ocean and their applications. Rev. Geophys. 1991, 29, 383–413. [Google Scholar] [CrossRef]
- GEOHAB. Global Ecology and Oceanography of Harmful Algal Blooms, GEOHAB Core Research Project: HABs in Fjords and Coastal Embayments; Cembella, A., Guzma, L., Roy, S., Dioge, J., Eds.; IOC and SCOR: Paris, France; Newark, DE, USA, 2010; p. 57. [Google Scholar]
- Pitcher, G.C.; Figueiras, F.G.; Hickey, B.M.; Moita, M.T. The physical oceanography of upwelling systems and the development of harmful algal blooms. Prog. Oceanogr. 2010, 85, 5–32. [Google Scholar] [CrossRef] [PubMed]
- Díaz, P.A.; Pérez-Santos, I.; Basti, L.; Garreaud, R.; Pinilla, E.; Barrera, F.; Tello, A.; Schwerter, C.; Arenas-Uribe, S.; Soto-Riquelme, C.; et al. How local and climate change drivers shaped the formation, dynamics and potential recurrence of a massive fish-killer microalgal bloom in Patagonian fjord. Sci. Total Environ. 2023, 865, 161288. [Google Scholar] [CrossRef] [PubMed]
- Smayda, T.J.; Trainer, V. Dinoflagellate blooms in upwelling systems: Seeding, variability, and contrasts with diatom bloom behaviour. Prog. Oceanogr. 2010, 85, 92–107. [Google Scholar] [CrossRef]
- Velo-Suárez, L.; Reguera, B.; Garcés, E.; Wyatt, T. Vertical distribution of division rates in coastal dinoflagellate Dinophysis spp. populations: Implications for modelling. Mar. Ecol. Prog. Ser. 2009, 385, 87–96. [Google Scholar] [CrossRef]
- Díaz, P.A.; Ruiz-Villareal, M.; Mouriño-Carballido, B.; Fernández-Pena, C.; Riobó, P.; Reguera, B. Fine scale physical-biological interactions during a shift from relaxation to upwelling with a focus on Dinophysis acuminata and its potential ciliate prey. Prog. Oceanogr. 2019, 175, 309–327. [Google Scholar] [CrossRef]
- Baldrich, Á.M.; Pérez-Santos, I.; Álvarez, G.; Reguera, B.; Fernández-Pena, C.; Rodríguez-Villegas, C.; Araya, M.; Álvarez, F.; Barrera, F.; Karasiewicz, S.; et al. Niche differentiation of Dinophysis acuta and D. acuminata in a stratified fjord. Harmful Algae 2021, 103, 102010. [Google Scholar] [CrossRef] [PubMed]
- Díaz, P.A.; Ruiz-Villarreal, M.; Velo-Suárez, L.; Ramilo, I.; Gentien, P.; Lunven, M.; Fernand, L.; Raine, R.; Reguera, B. Tidal and wind-event variability and the distribution of two groups of Pseudo-nitzschia species in an upwelling-influenced Ría. Deep Sea Res. II 2014, 101, 163–179. [Google Scholar] [CrossRef]
- Dekshenieks, M.; Donaghay, P.; Sullivan, J.; Rines, J.; Osborn, T.; Twardowski, M. Temporal and spatial occurrence of thin phytoplankton layers in relation to physical processes. Mar. Ecol. Prog. Ser. 2001, 223, 61–71. [Google Scholar] [CrossRef]
- McManus, M.A.; Alldredge, A.L.; Barnard, A.H.; Boss, E.; Case, J.F.; Cowles, T.J.; Donaghay, P.L.; Eisner, L.B.; Gifford, D.J.; Greenlaw, C.F.; et al. Characteristics, distribution and persistence of thin layers over a 48 hour period. Mar. Ecol. Prog. Ser. 2003, 261, 1–19. [Google Scholar] [CrossRef]
- Broullón, E.; López-Mozos, M.; Reguera, B.; Chouciño, P.; Doval, M.D.; Fernández-Castro, B.; Gilcoto, M.; Nogueira, E.; Souto, C.; Mouriño-Carballido, B. Thin layers of phytoplankton and harmful algae events in a coastal upwelling system. Prog. Oceanogr. 2020, 189, 102449. [Google Scholar] [CrossRef]
- Velo-Suárez, L.; González-Gil, S.; Gentien, P.; Lunven, M.; Bechemin, B.; Fernand, L.; Raine, R.; Reguera, B. Thin layers of Pseudo-nitzschia spp. and the fate of Dinophysis acuminata during an upwelling-downwelling cycle in a Galician Ría. Limnol. Oceanogr. 2008, 53, 1816–1834. [Google Scholar] [CrossRef]
- Villarino, M.L.; Figueiras, F.; Jones, K.; Alvarez-Salgado, X.A.; Richard, J.; Edwards, A. Evidence of in situ diel vertical migration of a red-tide microplancton species in Ría de Vigo (NW Spain). Mar. Biol. 1995, 123, 607–617. [Google Scholar] [CrossRef]
- MacKenzie, L. Does Dinophysis (Dinophyceae) have a sexual life cycle? J. Phycol. 1992, 28, 399–406. [Google Scholar] [CrossRef]
- Lassus, P.; Proniewski, F.; Pigeon, C.; Veret, L.; Le Déan, L.; Bardoul, M.; Truquet, P. The diurnal vertical migrations of Dinophysis acuminata in an outdoor tank at Antifer (Normandy, France). Aquat. Living Res. 1990, 3, 143–145. [Google Scholar] [CrossRef]
- Escalera, L.; Pazos, Y.; Doval, M.D.; Reguera, B. A comparison of integrated and discrete depth sampling for monitoring toxic species of Dinophysis. Mar. Pollut. Bull. 2012, 64, 106–113. [Google Scholar] [CrossRef] [PubMed]
- GEOHAB. Global Ecology and Oceanography of Harmful Algal Blooms, GEOHAB Core Research Project: HABs in Stratified Systems; Gentien, P., Reguera, B., Yamazaki, H., Fernand, L., Berdalet, E., Raine, R., Eds.; IOC and SCOR: Paris, France; Newark, DE, USA, 2008; p. 59. [Google Scholar]
- Dominguez, H.J.; Paz, B.; Daranas, A.H.; Norte, M.; Franco, J.M.; Fernández, J.J. Dinoflagellate polyether within the yessotoxin, pectenotoxin and okadaic acid toxin groups: Characterization, analysis and human health implications. Toxicon 2010, 15, 191–217. [Google Scholar] [CrossRef] [PubMed]
- Miles, C.O.; Wilkins, A.L.; Munday, R.; Dines, M.H.; Hawkes, A.D.; Briggs, L.R.; Sandvik, M.; Jensen, D.J.; Cooney, J.M.; Holland, P.T.; et al. Isolation of pectenotoxin-2 from Dinophysis acuta and its conversion to pectenotoxin-2 seco acid, and preliminary assessment of their acute toxicities. Toxicon 2004, 43, 1–9. [Google Scholar] [CrossRef]
- Gaillard, S.; Le Goïc, N.; Malo, F.; Boulais, M.; Fabioux, C.; Zaccagnini, L.; Carpentier, L.; Sibat, M.; Réveillon, D.; Séchet, V.; et al. Cultures of Dinophysis sacculus, D. acuminata and pectenotoxin 2 affect gametes and fertilization success of the Pacific oyster, Crassostrea gigas. Environ. Pollut. 2020, 265, 114840. [Google Scholar] [CrossRef]
- Rountos, K.; Kim, J.; Hattenrath-Lehmann, T.; Gobler, C.J. Effects of the harmful algae, Alexandrium catenella and Dinophysis acuminata, on the survival, growth, and swimming activity of early life stages of forage fish. Mar. Environ. Res. 2019, 148, 46–56. [Google Scholar] [CrossRef]
- Pease, S.K.D.; Brosnahan, M.L.; Sanderson, M.P.; Smith, J.L. Effects of two toxin-884 producing harmful algae, Alexandrium catenella and Dinophysis acuminata (Dinophyceae), on activity and mortality of larval shellfish. Toxins 2022, 14, 335. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission Implementing Regulation (EU) 2021/1709 of 23 September 2021 amending Implementing Regulation (EU) 2019/627 as regards uniform practical arrangements for the performance of official controls on products of animal origin. Off. J. Eur. Union 2021, 339, 84–87. [Google Scholar]
- Reguera, B.; Riobó, P.; Rodríguez, F.; Díaz, P.A.; Pizarro, G.; Paz, B.; Franco, J.M.; Blanco, J. Dinophysis toxins: Causative organisms, distribution and fate in shellfish. Mar. Drugs 2014, 12, 394–461. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.; Arévalo, F.; Correa, J.; Moroño, Á. Lipophilic toxins in Galicia (NW Spain) between 2014 and 2017: Incidence on the main molluscan species and analysis of the monitoring efficiency. Toxins 2019, 11, 612. [Google Scholar] [CrossRef]
- Escalera, L.; Reguera, B.; Pazos, Y.; Moroño, A.; Cabanas, J.M. Are different species of Dinophysis selected by climatological conditions? Afr. J. Mar. Sci. 2006, 28, 283–288. [Google Scholar] [CrossRef]
- Reguera, B.; Mariño, J.; Campos, J.; Bravo, I.; Fraga, S. Trends in the occurrence of Dinophysis spp. in Galician waters. In Toxic Phytoplankton Blooms in the Sea; Smayda, T., Shimizu, Y., Eds.; Elsevier Science Publishers B.V.: Amsterdam, The Netherland, 1993; pp. 559–564. [Google Scholar]
- Palma, A.S.; Vilarinho, M.G.; Moita, M.T. Interannual trends in the longshore distribution of Dinophysis off the Portuguese coast. In Harmful Algae; Reguera, B., Blanco, J., Fernández, M.L., Wyatt, T., Eds.; Xunta de Galicia and IOC of UNESCO: Vigo, Spain, 1998; Volume 124–127. [Google Scholar]
- Lindahl, O.; Lundve, B.; Johansen, M. Toxicity of Dinophysis spp. in relation to population density and environmental conditions on the Swedish west coast. Harmful Algae 2007, 6, 218–231. [Google Scholar] [CrossRef]
- Díaz, P.A.; Reguera, B.; Moita, T.; Bravo, I.; Ruíz-Villarreal, M.; Fraga, S. Mesoscale dynamics and niche segregation of two Dinophysis species in Galician-Portuguese coastal waters. Toxins 2019, 11, 37. [Google Scholar] [CrossRef]
- Díaz, P.A.; Pérez-Santos, I.; Álvarez, G.; Garreaud, R.; Pinilla, E.; Díaz, M.; Sandoval, A.; Araya, M.; Álvarez, F.; Rengel, J.; et al. Multiscale physical background to an exceptional harmful algal bloom of Dinophysis acuta in a fjord system. Sci. Total Environ. 2021, 773, 145621. [Google Scholar] [CrossRef]
- Alves de Souza, C.; Iriarte, J.L.; Mardones, J.I. Interannual variability of Dinophysis acuminata and Protoceratium reticulatum in a Chilean fjord: Insights from the realized niche analysis. Toxins 2019, 11, 19. [Google Scholar] [CrossRef]
- Baldrich, Á.M.; Molinet, C.; Reguera, B.; Espinoza-González, O.; Pizarro, G.; Rodríguez-Villegas, C.; Opazo, D.; Mejías, P.; Díaz, P.A. Interannual variability in mesoscale distribution of Dinophysis acuminata and D. acuta in Northwestern Patagonian fjords. Harmful Algae 2022, 115, 102228. [Google Scholar] [CrossRef]
- Díaz, P.A.; Álvarez, G.; Pizarro, G.; Blanco, J.; Reguera, B. Lipophilic toxins in Chile: History, producers and impacts. Mar. Drugs 2022, 20, 122. [Google Scholar] [CrossRef] [PubMed]
- Díaz, P.A.; Álvarez, G.; Seguel, M.; Marín, A.; Krock, B. First detection of pectenotoxin- 2 in shellfish associated with an intense spring bloom of Dinophysis acuminata on the central Chilean coast. Mar. Pollut. Bull. 2020, 158, 11414. [Google Scholar] [CrossRef]
- Blanco, J.; Álvarez, G.; Uribe, E. Identification of pectenotoxins in plankton, filter feeders, and isolated cells of a Dinophysis acuminata with an atypical toxin profile, from Chile. Toxicon 2007, 49, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Uribe, J.C.; García, C.; Rivas, M.; Lagos, N. First report of diarrhetic shellfish toxins in Magellanic fjord, Southern Chile. J. Shellfish Res. 2001, 20, 69–74. [Google Scholar]
- Alves de Souza, C.; Varela, D.; Contreras, C.; de la Iglesia, P.; Fernández, P.; Hipp, B.; Hernández, C.; Riobó, P.; Reguera, B.; Franco, J.M.; et al. Seasonal variability of Dinophysis spp. and Protoceratium reticulatum associated to lipophilic shellfish toxins in a strongly stratified Chilean fjord. Deep Sea Res. II 2014, 101, 152–162. [Google Scholar] [CrossRef]
- Suárez-Isla, B.A.; Barrera, F.; Carrasco, D.; Cigarra, L.; López-Rivera, A.M.; Rubilar, I.; Alcayaga, C.; Contreras, V.; Seguel, M. Comprehensive study of the occurrence and distribution of lipophilic marine toxins in shellfish from production areas in Chile. In Harmful Algae 2018—From ecosystems to socioecosystems. In Proceedings of the 18th International Conference on Harmful Algae, Nantes, France, 21–26 October 2018; Hess, P., Ed.; International Society for the Study of Harmful Algae: Oostende, Belgium, 2020; pp. 163–166, ISBN 978-87-990827-7-3. [Google Scholar]
- Paredes-Mella, J.; Mardones, J.; Norambuena, L.; Fuenzalida, G.; Labra, G.; Nagai, K. Toxic Dinophysis acuminata in southern Chile: A comparative approach based on its first local in vitro culture and offshore-estuarine bloom dynamics. Prog. Oceanogr. 2022, 209, 102918. [Google Scholar] [CrossRef]
- Hutchinson, G.E. Population studies-animal ecology and demography-concluding remarks. In Cold Spring Harbor Symposia on Quantitative Biology 22: 415–427; Bungtown, R.D., Ed.; Cold Spring Harbor Lab Press: New York, NY, USA, 1957. [Google Scholar]
- Colwell, R.K.; Rangel, T.F. Hutchinson’s duality: The once and future niche. Proc. Natl. Acad. Sci. USA 2009, 106, 19651–19658. [Google Scholar] [CrossRef]
- Holt, R.D. Bringing the Hutchinsonian niche into the 21st century: Ecological and evolutionary perspectives. Proc. Natl. Acad. Sci. USA 2009, 106, 19659–19665. [Google Scholar] [CrossRef]
- Pagel, J.; Schurr, F.M. Forecasting species ranges by statistical estimation of ecological niches and spatial population dynamics. Glob. Ecol. Biogeogr. 2012, 21, 293–304. [Google Scholar] [CrossRef]
- Dolédec, S.; Chessel, D.; Gimaret-Carpentier, C. Niche separation in community analysis: A new method. Ecology 2000, 81, 2914–2927. [Google Scholar] [CrossRef]
- Grüner, N.; Gebühr, C.; Boersma, M.; Feudel, U.; Wiltshire, K.H.; Freund, J.A. Reconstructing the realized niche of phytoplankton species from environmental data: Fitness versus abundance approach. Limnol. Oceanogr. Methods 2011, 9, 432–442. [Google Scholar] [CrossRef]
- Hernández-Fariñas, T.; Bacher, C.; Soudant, D.; Belin, C.; Barillé, L. Assessing phytoplankton realized niches using a French national phytoplankton monitoring network. Estuar. Coast. Shelf Sci. 2015, 159, 15–27. [Google Scholar] [CrossRef]
- Sutani, D.; Utsumi, M.; Kato, Y.; Sugiura, N. Estimation of the changes in phytoplankton community composition in a volcanic acidotrophic Lake, Inawashiro, Japan. Jpn. J. Water Treat. Biol. 2014, 50, 53–69. [Google Scholar] [CrossRef]
- Karasiewicz, S.; Chapelle, A.; Bacher, C.; Soudant, D. Harmful algae niche responses to environmental and community variation along the French coast. Harmful Algae 2020, 93, 101785. [Google Scholar] [CrossRef] [PubMed]
- Karasiewicz, S.; Breton, E.; Lefebvre, A.; Hernández-Fariñas, T.; Lefebvre, S. Realized niche analysis of phytoplankton communities involving HAB: Phaeocystis spp. as a case study. Harmful Algae 2018, 73, 1–13. [Google Scholar] [CrossRef]
- Pérez-Santos, I.; Garcés-Vargas, J.; Schneider, W.; Ross, L.; Parra, S.; Valle-Levinson, A. Double-diffusive layering and mixing in Patagonian fjords. Prog. Oceanogr. 2014, 129, 35–49. [Google Scholar] [CrossRef]
- Fernandes-Salvador, J.A.; Davidson, K.; Sourisseau, M.; Revilla, M.; Schmidt, W.; Clarke, D.; Miller, P.I.; Arce, P.; Fernández, R.; Maman, L.; et al. Current status of forecasting toxic harmful algae for the North-East Atlantic shellfish aquaculture Industry. Front. Mar. Sci. 2021, 8, 666583. [Google Scholar] [CrossRef]
- MacKenzie, L.; Truman, P.; Satake, M.; Yasumoto, T.; Adamson, J.; Mountfort, D.; White, D. Dinoflagellate blooms and associated DSP-toxicity in shellfish in New Zealand. In Harmful Algae; Reguera, B., Blanco, J., Fernández, M.L., Wyatt, T., Eds.; Xunta de Galicia and Intergovermental Oceanographic Commission of UNESCO: Vigo, Spain, 1998; pp. 74–77. [Google Scholar]
- Swan, S.C.; Turner, A.D.; Bresnan, E.; Whyte, C.; Paterson, R.F.; McNeill, S.; Mitchell, E.; Davidson, K. Dinophysis acuta in Scottish coastal waters and its influence on Diarrhetic Shellfish Toxin profiles. Toxins 2018, 10, 399. [Google Scholar] [CrossRef]
- Raine, R.; Cosgrove, S.; Fennell, S.; Gregory, C.; Barnett, M.; Purdie, D.; Cave, R. Origins of Dinophysis blooms which impact Irish aquaculture. In Marine and Fresh-Water Harmful Algae, Proceedings of the 17th International Conference on Harmful Algae, Florianópolis, Brazil, 9–14 October 2016; Proença, L.A.O., Hallegraeff, G., Eds.; International Society for the Study of Harmful Algae and Intergovernmental Oceanographic Commission of UNESCO: Oostende, Belgium, 2017; pp. 46–49. [Google Scholar]
- Díaz, P.A.; Reguera, B.; Ruiz-Villarreal, M.; Pazos, Y.; Velo-Suárez, L.; Berger, H.; Sourisseau, M. Climate variability and oceanographic settings associated with interannual variability in the initiation of Dinophysis acuminata blooms. Mar. Drugs 2013, 11, 2964–2981. [Google Scholar] [CrossRef]
- Díaz, P.A.; Ruiz-Villarreal, M.; Pazos, Y.; Moita, M.T.; Reguera, B. Climate variability and Dinophysis acuta blooms in an upwelling system. Harmful Algae 2016, 53, 145–159. [Google Scholar] [CrossRef]
- Schneider, W.; Pérez-Santos, I.; Ross, L.; Bravo, L.; Seguel, R.; Hernández, F. On the hydrography of Puyuhuapi Channel, Chilean Patagonia. Prog. Oceanogr. 2014, 128, 8–18. [Google Scholar] [CrossRef]
- Díaz, P.; Molinet, C.; Cáceres, M.; Valle-Levinson, A. Seasonal and intratidal distribution of Dinophysis spp. in a Chilean fjord. Harmful Algae 2011, 10, 155–164. [Google Scholar] [CrossRef]
- Hattenrath-Lehmann, T.; Gobler, C.J. The contribution of inorganic and organic nutrients to the growth of a North American isolate of the mixotrophic dinoflagellate, Dinophysis acuminata. Limnol. Oceanogr. 2015, 60, 1588–1603. [Google Scholar] [CrossRef]
- García-Portela, M.; Reguera, B.; Gago, J.; Le Gac, M.; Rodríguez, F. Uptake of inorganic and organic nitrogen sources by Dinophysis acuminata and D. acuta. Microorganisms 2020, 8, 187. [Google Scholar] [CrossRef] [PubMed]
- Hansen, P.J.; Nielsen, L.T.; Johnson, M.; Berge, T.; Flynn, K.R. Acquired phototrophy in Mesodinium and Dinophysis—A review of cellular organization, prey selectivity, nutrient uptake and bioenergetics. Harmful Algae 2013, 28, 126–139. [Google Scholar] [CrossRef]
- Velo-Suárez, L.; González-Gil, S.; Pazos, Y.; Reguera, B. The growth season of Dinophysis acuminata in an upwelling system embayment: A conceptual model based on in situ measurements. Deep Sea Res. II 2014, 101, 141–151. [Google Scholar] [CrossRef]
- Tong, M.; Smith, J.L.; Kulis, D.M.; Andersen, D.M. Role of dissolved nitrate and phosphate in isolates of Mesodinium rubrum and toxin-producing Dinophysis acuminata. Aquat. Microb. Ecol. 2015, 72, 169–185. [Google Scholar] [CrossRef]
- Pizarro, G.; Alarcón, C.; Franco, J.M.; Palma, M.; Escalera, L.; Reguera, B.; Vidal, G.; Guzmán, L. Distribución espacial de Dinophysis spp. y detección de toxinas DSP en el agua mediante resinas DIAION (verano 2006, Región de Los Lagos, Chile). Cienc. Tecnol. Mar 2011, 34, 31–48. [Google Scholar]
- Park, M.; Kim, S.; Kim, H.; Myung, G.; Kang, Y.; Yih, W. First successful culture of the marine dinoflagellate Dinophysis acuminata. Aquat. Microb. Ecol. 2006, 45, 101–106. [Google Scholar] [CrossRef]
- González-Gil, S.; Velo-Suárez, L.; Gentien, P.; Ramilo, I.; Reguera, R. Phytoplankton assemblages and characterization of a Dinophysis acuminata population during an upwelling-downwelling cycle. Aquat. Microb. Ecol. 2010, 58, 273–286. [Google Scholar] [CrossRef]
- Sjöqvist, C.O.; Lindholm, T.J. Natural co-occurrence of Dinophysis acuminata (Dinoflagellata) and Mesodinium rubrum (Ciliophora) in thin layers in a coastal inlet. J. Eukaryot. Microbiol. 2011, 58, 365–372. [Google Scholar] [CrossRef]
- Reguera, B.; Velo-Suárez, L.; Raine, R.; Park, M. Harmful Dinophysis species: A review. Harmful Algae 2012, 14, 87–106. [Google Scholar] [CrossRef]
- Moita, M.T.; Pazos, Y.; Rocha, C.; Nolasco, R.; Oliveira, P.B. Towards predicting Dinophysis blooms off NW Iberia: A decade of events. Harmful Algae 2016, 52, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Pickard, G.L. Some physical oceanographic features of inlets of Chile. J. Fish. Res. Board Can. 1971, 28, 1077–1106. [Google Scholar] [CrossRef]
- Sauter, T. Revisiting extreme precipitation amounts over southern South America and implications for the Patagonian Icefields. Hydrol. Earth Syst. Sci. 2020, 24, 203–2016. [Google Scholar] [CrossRef]
- Silva, N.; Calvete, C. Características oceanográficas físicas y químicas de canales australes chilenos entre el golfo de Penas y el estrecho de Magallanes (crucero Cimar Fiordo-2) Cienc. Tecnol. Mar 2002, 25, 23–88. [Google Scholar]
- Valle-Levinson, A.; Sarkar, N.; Sanay, R.; Soto, D.; León, J. Spatial structure of hydrography and flow in a Chilean fjord, Estuario Reloncaví. Estuaries Coasts 2007, 30, 113–126. [Google Scholar] [CrossRef]
- Sepúlveda, J.; Pantoja, S.; Hughen, K.; Lange, C.; Gonzalez, F.; Muñoz, P.; Rebolledo, L.; Castro, R.; Contreras, S.; Ávila, A. Fluctuations in export productivity over the last century from sediments of a southern Chilean fjord (44 S). Estuar. Coast. Shelf Sci. 2005, 65, 587–600. [Google Scholar] [CrossRef]
- Pinilla, E.; Soto, G.; Soto-Riquelme, C. Determinación de las escalas de intercambio de agua en fiordos y canales de la Patagonia Sur, Etapa II. Alparaiso Inst. Fom. Pesq. (IFOP) 2019, 10, 1–52. [Google Scholar]
- Pinilla, E. Determinación de las escalas de intercambio de agua en fiordos y canales de la Patagonia norte. IFOP 2018, 10, 1–35. [Google Scholar]
- Schlitzer, R. Data analysis and visualization with Ocean Data View. CMOS Bull. SCMO 2015, 41, 9–13. [Google Scholar]
- Lovegrove, T. An improved form of sedimentation apparatus for use with an inverted microscope. J. Cons. Int. Explor. Mer. 1960, 25, 279–284. [Google Scholar] [CrossRef]
- Grasshoff, K.; Ehrhardt, M.; Kremling, K. Methods of Seawater Analysis, 2nd ed.; Verlag Chemie Weinhein: New York, NY, USA, 1983. [Google Scholar]
- Kattner, G.; Becker, H. Nutrients and organic nitrogenous compounds in the marginal ice zone of the Fram Strait. J. Mar. Syst. 1991, 2, 385–394. [Google Scholar] [CrossRef]
- Utermöhl, H. Zur Vervollkomnung der quantitativen phytoplankton-Methodik. Mitt. Int. Ver. Limnol. 1958, 9, 1–38. [Google Scholar]
- Reguera, B.; Garcés, E.; Pazos, Y.; Bravo, I.; Ramilo, I.; González-Gil, S. Cell cycle patterns and estimates of in situ división rates of dinoflagellates of the genus Dinophysis by a postmitotic index. Mar. Ecol. Prog. Ser. 2003, 249, 117–131. [Google Scholar] [CrossRef]
- Carpenter, E.J.; Chang, J. Species-specific phytoplankton growth rates via diel DNA synthesis cycles. I. Concept of the method. Mar. Ecol. Prog. Ser. 1988, 43, 105–111. [Google Scholar] [CrossRef]
- Regueiro, J.; Rossignoli, A.; Álvarez, G.; Blanco, J. Automated on-line solid-phase extraction coupled to liquid chromatography–tandem mass spectrometry for determination of lipophilic marine toxins in shellfish. Food Chem. 2011, 129, 533–540. [Google Scholar] [CrossRef]
- Karasiewicz, S.; Dolédec, S.; Lefebvre, S. Within outlying mean indexes: Re ning the OMI analysis for the realized niche decomposition. PeerJ 2017, 5, 3364. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational multivariate analysis of variance (PERMANOVA). In Wiley Statsref: Statistics Reference Online; Wiley: Hoboken, NJ, USA, 2014; pp. 1–15. [Google Scholar]
- Dray, S.; Dufour, A.-B. The ade4 package: Implementing the duality diagram for ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; et al. Vegan: Community ecology package. R Package Version 2018, 2, 321–326. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; ISBN 3-900051-07-0. Available online: http://www.r-project.org/ (accessed on 15 December 2022).











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Code | Inertia | OMI | Tol | Rtol | p-Value |
|---|---|---|---|---|---|---|
| Puyuhuapi Fjord | ||||||
| Dinophysis acuminata | Dacuminata | 2.77 | 0.30 | 1.15 | 1.32 | <0.01 |
| Dinophysis acuta | Dacuta | 2.55 | 0.97 | 0.76 | 0.82 | <0.01 |
| Mesodinium spp. | Meso | 4.09 | 0.07 | 2.68 | 1.22 | <0.01 |
| Leegaardiella sp. | Leeg | 2.23 | 0.81 | 0.73 | 0.68 | <0.01 |
| Paratontonia spp. | Para | 2.05 | 0.27 | 0.46 | 1.32 | <0.01 |
| Strombidium spp. | Strom | 2.81 | 0.10 | 0.61 | 2.09 | <0.01 |
| Pseudotontonia sp. | Pse | 2.88 | 0.47 | 0.95 | 1.45 | 0.18 |
| Laboea sp. | Labo | 1.70 | 0.33 | 0.28 | 1.09 | 0.59 |
| Lohmanniella sp. | Loh | 3.61 | 0.02 | 1.52 | 2.06 | 0.81 |
| Pitipalena Fjord | ||||||
| Dinophysis acuminata | Dacuminata | 6.39 | 2.43 | 2.78 | 1.18 | <0.01 |
| Dinophysis acuta | Dacuta | 9.27 | 5.59 | 1.23 | 0.46 | 0.01 |
| Mesodinium spp. | Meso | 4.57 | 0.07 | 3.83 | 0.67 | <0.01 |
| Leegaardiella sp. | Leeg | 2.23 | 0.81 | 0.73 | 0.68 | 0.02 |
| Paratontonia spp. | Para | 4.38 | 0.39 | 3.18 | 0.80 | 0.09 |
| Strombidium spp. | Strom | 5.76 | 1.04 | 3.78 | 0.95 | <0.01 |
| Pseudotontonia sp. | Pse | 6.09 | 1.41 | 3.51 | 1.17 | <0.01 |
| Laboea sp. | Labo | 3.49 | 0.67 | 1.50 | 1.32 | 0.50 |
| Lohmanniella sp. | Loh | 6.69 | 1.31 | 4.42 | 0.95 | <0.01 |
| Predictive Variables | Df | SS | R2 | Pseudo-F | Pr > F |
|---|---|---|---|---|---|
| Puyuhuapi Fjord | |||||
| Depth | 1 | 0.31 | 0.0056 | 0.0684 | 0.92 |
| Temperature | 1 | 15.80 | 0.0289 | 3.5404 | 0.03 |
| Salinity | 1 | 0.20 | 0.0004 | 0.0453 | 0.95 |
| Brunt–Väisälä frequency | 1 | 0.60 | 0.0011 | 0.1334 | 0.85 |
| Residuals | 99 | 441.68 | 0.8098 | ||
| Total | 103 | 545.41 | 1.0000 | ||
| Pitipalena Fjord | |||||
| Depth | 1 | 1.05 | 0.0070 | 0.3531 | 0.67 |
| Temperature | 1 | 17.64 | 0.1172 | 5.9359 | 0.01 |
| Salinity | 1 | 11.26 | 0.0748 | 3.7880 | 0.03 |
| Brunt–Väisälä frequency | 1 | 1.89 | 0.0126 | 0.6385 | 0.51 |
| Residuals | 43 | 127.81 | 0.8488 | ||
| Total | 47 | 150.58 | 1.0000 |
| Toxin | LOQ (ng/mL) | LOD (ng/mL) |
|---|---|---|
| OA | 0.50 ± 0.03 | 0.32 ± 0.03 |
| DTX1 | 0.39 ± 0.02 | 0.24 ± 0.02 |
| PTX2 | 4.33 ± 0.25 | 2.61 ± 0.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldrich, Á.M.; Díaz, P.A.; Álvarez, G.; Pérez-Santos, I.; Schwerter, C.; Díaz, M.; Araya, M.; Nieves, M.G.; Rodríguez-Villegas, C.; Barrera, F.; et al. Dinophysis acuminata or Dinophysis acuta: What Makes the Difference in Highly Stratified Fjords? Mar. Drugs 2023, 21, 64. https://doi.org/10.3390/md21020064
Baldrich ÁM, Díaz PA, Álvarez G, Pérez-Santos I, Schwerter C, Díaz M, Araya M, Nieves MG, Rodríguez-Villegas C, Barrera F, et al. Dinophysis acuminata or Dinophysis acuta: What Makes the Difference in Highly Stratified Fjords? Marine Drugs. 2023; 21(2):64. https://doi.org/10.3390/md21020064
Chicago/Turabian StyleBaldrich, Ángela M., Patricio A. Díaz, Gonzalo Álvarez, Iván Pérez-Santos, Camila Schwerter, Manuel Díaz, Michael Araya, María Gabriela Nieves, Camilo Rodríguez-Villegas, Facundo Barrera, and et al. 2023. "Dinophysis acuminata or Dinophysis acuta: What Makes the Difference in Highly Stratified Fjords?" Marine Drugs 21, no. 2: 64. https://doi.org/10.3390/md21020064
APA StyleBaldrich, Á. M., Díaz, P. A., Álvarez, G., Pérez-Santos, I., Schwerter, C., Díaz, M., Araya, M., Nieves, M. G., Rodríguez-Villegas, C., Barrera, F., Fernández-Pena, C., Arenas-Uribe, S., Navarro, P., & Reguera, B. (2023). Dinophysis acuminata or Dinophysis acuta: What Makes the Difference in Highly Stratified Fjords? Marine Drugs, 21(2), 64. https://doi.org/10.3390/md21020064