

Anti-Pollutant Activity of Porphyra yezoensis Water Extract and Its Active Compound, Porphyra 334, against Urban Particulate Matter-Induced Keratinocyte Cell Damage
 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Porphyra yezoensis Extract Reduces ROS Production Induced by UPM through Inhibiting XRE Promoter Activity
2.2. Preparation of P. yezoensis Extract and Porphyra 334 from Laver
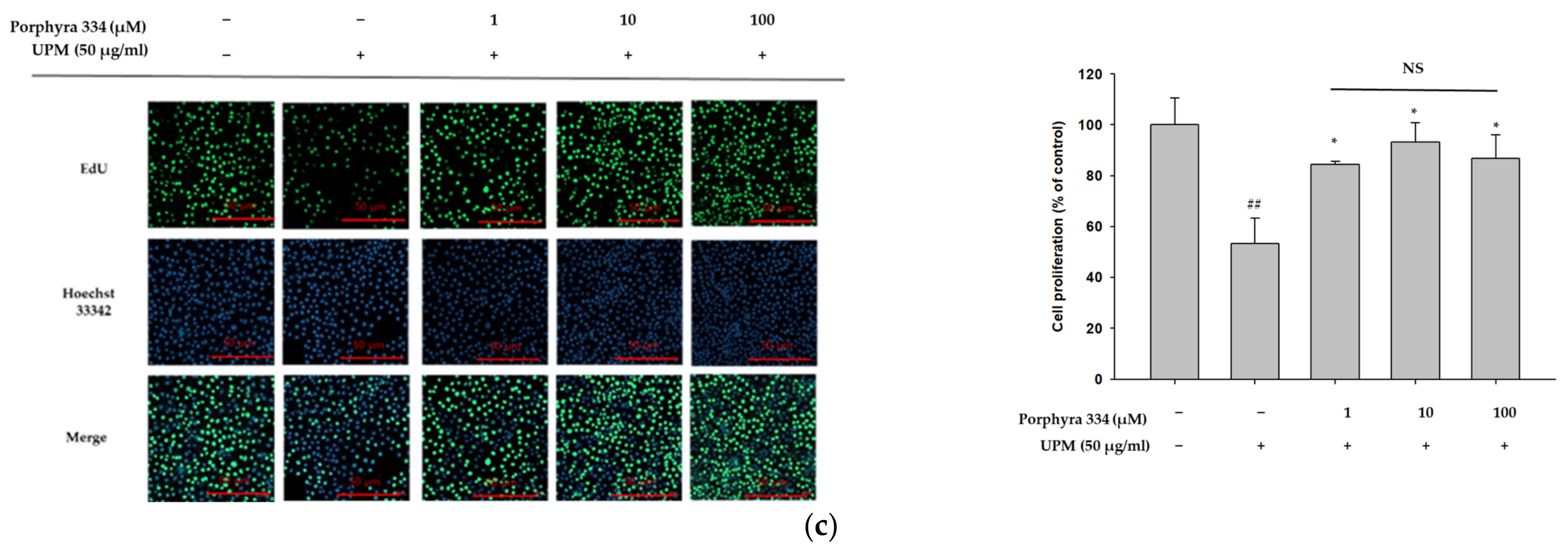
2.3. Porphyra 334 Induces Cell Proliferation by Inhibiting ROS Production Induced by UPM in Human Keratinocytes
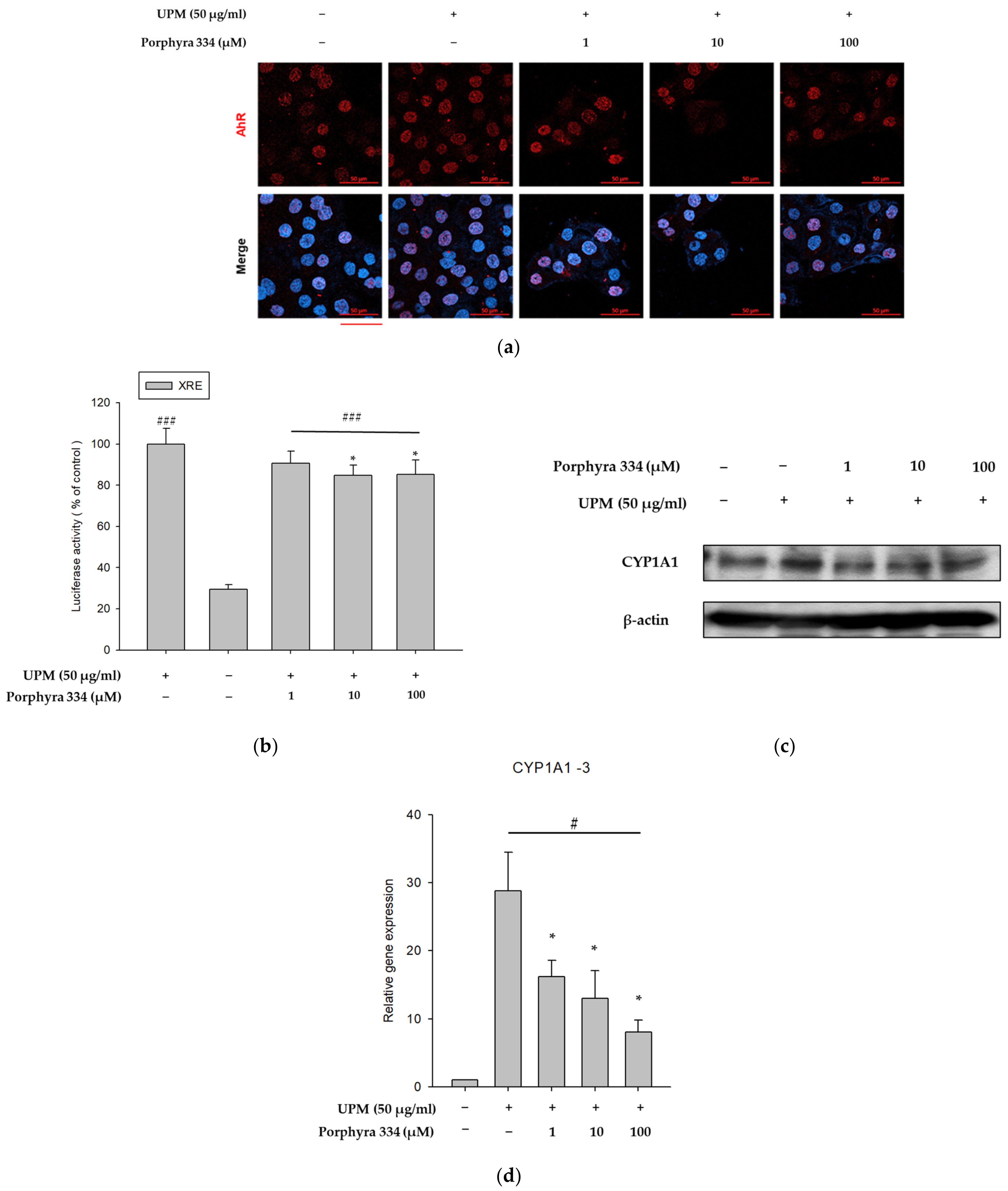
2.4. Porphyra 334 Exerts Anti-Pollutant Effects by Reducing XRE Promoter Activity
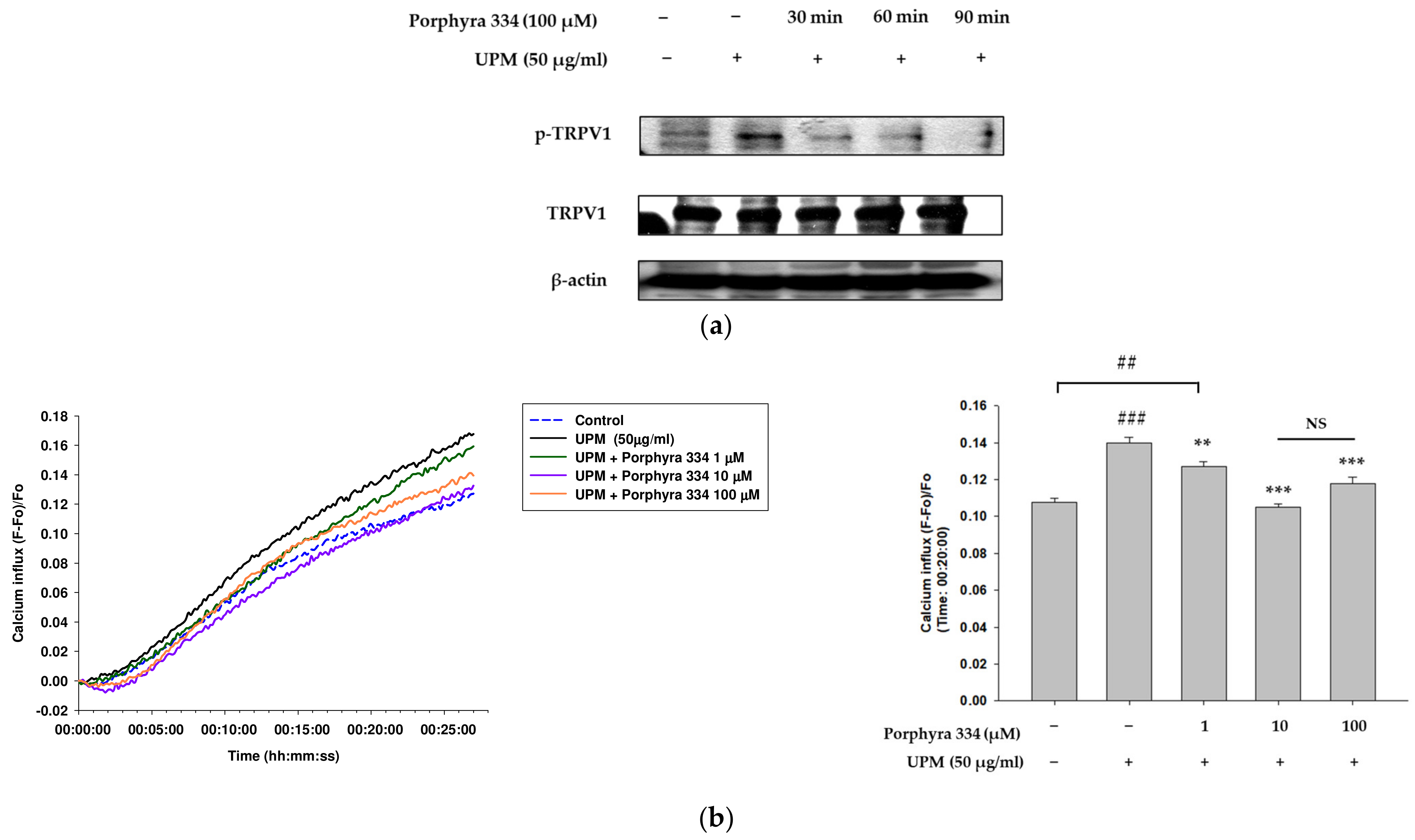
2.5. Porphyra 334 Inhibits the Activation of TRPV1 and Calcium Influx Induced by UPM
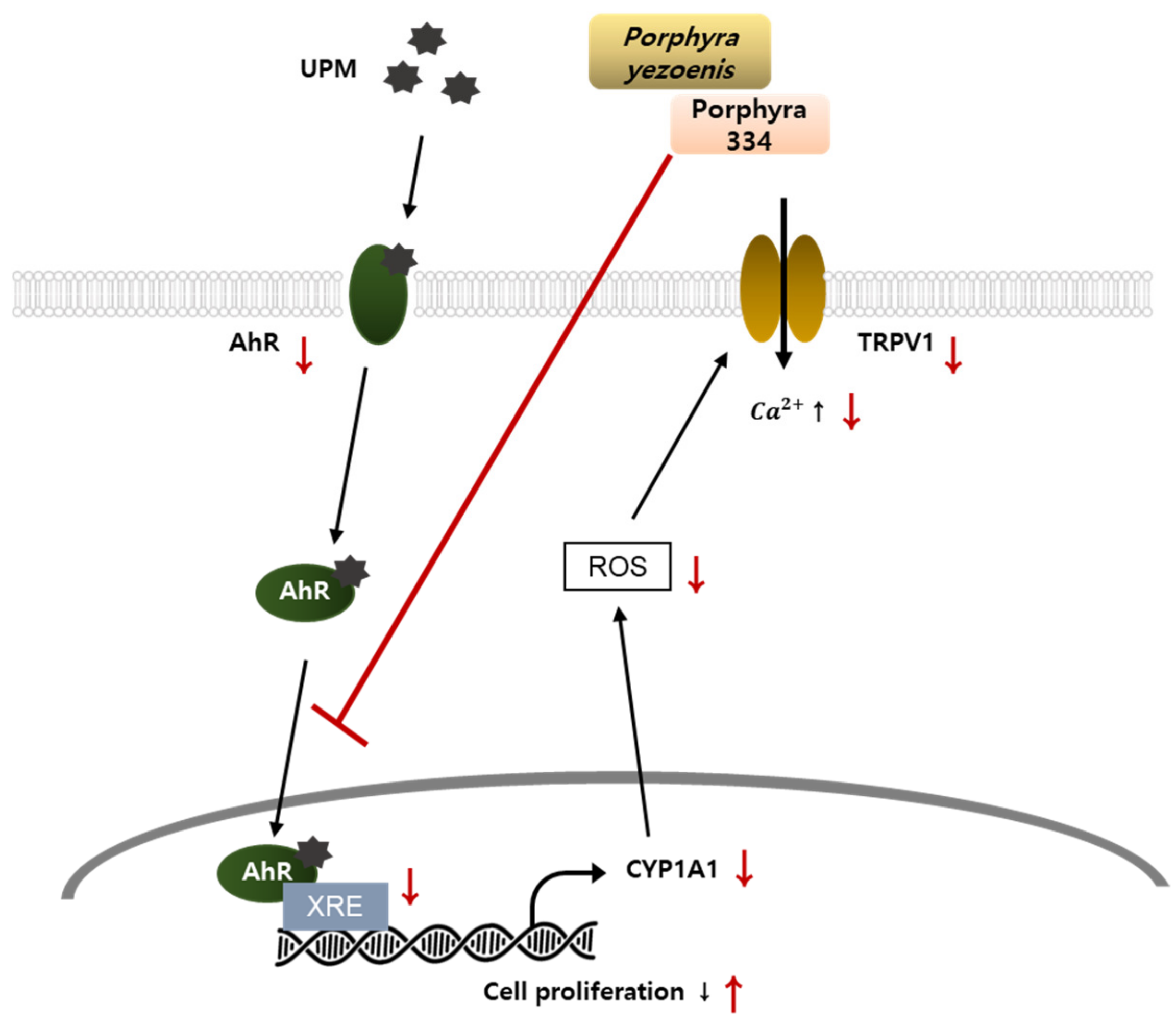
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Materials
4.2. Preparation of Porphyra yezoensis Extract and Porphyra 334 from Laver
4.3. Porphyra 334 and UPM Treatment
4.4. ABTS Radical Scavenging Activity
4.5. EdU Incorporation Assay
4.6. Fluo-4 Ca2+ Influx Assay
4.7. DCF-DA Cellular ROS Detection Assay
4.8. Western Blot Analysis
4.9. Luciferase Reporter Assay and β-Galactosidase Activity Assay
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Dagouassat, M.; Lanone, S.; Boczkowski, J. Interaction of matrix metalloproteinases with pulmonary pollutants. Eur. Respir. J. 2012, 39, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Kampa, M.; Castanas, E. Human health effects of air pollution. Environ. Pollut. 2008, 151, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Newby, D.E.; Mannucci, P.M.; Tell, G.S.; Baccarelli, A.A.; Brook, R.D.; Donaldson, K.; Forastiere, F.; Franchini, M.; Franco, O.H.; Graham, I.; et al. Expert position paper on air pollution and cardiovascular disease. Eur. Heart J. 2015, 36, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Franchini, M.; Mannucci, P.M. Short-term effects of air pollution on cardiovascular diseases: Outcomes and mechanisms. J. Thromb. Haemost. 2007, 5, 2169–2174. [Google Scholar] [CrossRef]
- Langrish, J.P.; Bosson, J.; Unosson, J.; Muala, A.; Newby, D.E.; Mills, N.L.; Blomberg, A.; Sandström, T. Cardiovascular effects of particulate air pollution exposure: Time course and underlying mechanisms. J. Intern. Med. 2012, 272, 224–239. [Google Scholar] [CrossRef]
- Kim, K.H.; Kabir, E.; Kabir, S. A review on the human health impact of airborne particulate matter. Environ. Int. 2015, 74, 136–143. [Google Scholar] [CrossRef]
- Kim, J.; Kim, E.H.; Oh, I.; Jung, K.; Han, Y.; Cheong, H.K.; Ahn, K. Symptoms of atopic dermatitis are influenced by outdoor air pollution. J. Allergy Clin. Immunol. 2013, 132, 495–498.e1. [Google Scholar] [CrossRef]
- Song, S.; Lee, K.; Lee, Y.M.; Lee, J.H.; Lee, S.I.; Yu, S.D.; Paek, D. Acute health effects of urban fine and ultrafine particles on children with atopic dermatitis. Environ. Res. 2011, 111, 394–399. [Google Scholar] [CrossRef]
- Song, S.; Paek, D.; Park, C.; Lee, C.; Lee, J.H.; Yu, S.D. Exposure to ambient ultrafine particles and urinary 8-hydroxyl-2-deoxyguanosine in children with and without eczema. Sci. Total Environ. 2013, 458–460, 408–413. [Google Scholar] [CrossRef]
- Jin, S.P.; Li, Z.; Choi, E.K.; Lee, S.; Kim, Y.K.; Seo, E.Y.; Chung, J.H.; Cho, S. Urban particulate matter in air pollution penetrates into the barrier-disrupted skin and produces ROS-dependent cutaneous inflammatory response in vivo. J. Dermatol. Sci. 2018, 91, 175–183. [Google Scholar] [CrossRef]
- Ngoc, L.T.N.; Park, D.; Lee, Y.; Lee, Y.C. Systematic Review and Meta-Analysis of Human Skin Diseases Due to Particulate Matter. Int. J. Environ. Res. Public Health 2017, 14, 1458. [Google Scholar] [CrossRef]
- Jang, H.S.; Lee, J.E.; Myung, C.H.; Park, J.I.; Jo, C.S.; Hwang, J.S. Particulate Matter-Induced Aryl Hydrocarbon Receptor Regulates Autophagy in Keratinocytes. Biomol. Ther. 2019, 27, 570–576. [Google Scholar] [CrossRef]
- Lawal, A.O. Air particulate matter induced oxidative stress and inflammation in cardiovascular disease and atherosclerosis: The role of Nrf2 and AhR-mediated pathways. Toxicol. Lett. 2017, 270, 88–95. [Google Scholar] [CrossRef]
- Bodó, E.; Kovács, I.; Telek, A.; Varga, A.; Paus, R.; Kovács, L.; Bíró, T. Vanilloid receptor-1 (VR1) is widely expressed on various epithelial and mesenchymal cell types of human skin. J. Investig. Dermatol. 2004, 123, 410–413. [Google Scholar] [CrossRef]
- Ständer, S.; Moormann, C.; Schumacher, M.; Buddenkotte, J.; Artuc, M.; Shpacovitch, V.; Brzoska, T.; Lippert, U.; Henz, B.M.; Luger, T.A.; et al. Expression of vanilloid receptor subtype 1 in cutaneous sensory nerve fibers, mast cells, and epithelial cells of appendage structures. Exp. Dermatol. 2004, 13, 129–139. [Google Scholar] [CrossRef]
- Wang, Y. The functional regulation of TRPV1 and its role in pain sensitization. Neurochem. Res. 2008, 33, 2008–2012. [Google Scholar] [CrossRef]
- Yoshida, T.; Inoue, R.; Morii, T.; Takahashi, N.; Yamamoto, S.; Hara, Y.; Tominaga, M.; Shimizu, S.; Sato, Y.; Mori, Y. Nitric oxide activates TRP channels by cysteine S-nitrosylation. Nat. Chem. Biol. 2006, 2, 596–607. [Google Scholar] [CrossRef]
- Ibi, M.; Matsuno, K.; Shiba, D.; Katsuyama, M.; Iwata, K.; Kakehi, T.; Nakagawa, T.; Sango, K.; Shirai, Y.; Yokoyama, T.; et al. Reactive oxygen species derived from NOX1/NADPH oxidase enhance inflammatory pain. J. Neurosci. 2008, 28, 9486–9494. [Google Scholar] [CrossRef]
- Lafarga, T.; Acién-Fernández, F.G.; Garcia-Vaquero, M. Bioactive peptides and carbohydrates from seaweed for food applications: Natural occurrence, isolation, purification, and identification. Algal Res. 2020, 48, 101909. [Google Scholar] [CrossRef]
- Corsetto, P.A.; Montorfano, G.; Zava, S.; Colombo, I.; Ingadottir, B.; Jonsdottir, R.; Sveinsdottir, K.; Rizzo, A.M. Characterization of Antioxidant Potential of Seaweed Extracts for Enrichment of Convenience Food. Antioxidants 2020, 9, 249. [Google Scholar] [CrossRef]
- Lin, R.; Stekoll, M.S. Responses of chlorophyll a content for conchocelis phase of alaskan porphyra (bangiales, rhodophyta) species to environmental factors. Arch. Biochem. Biophys. 2013, 1, 28–39. [Google Scholar] [CrossRef]
- Subramaniyan, S.A.; Begum, N.; Kim, S.J.; Choi, Y.H.; Nam, T.J. Biopeptides of Pyropia yezoensis and their potential health benefits: A review. Asian Pac. J. Trop. Biomed. 2021, 11, 375–384. [Google Scholar] [CrossRef]
- Ryu, J.; Park, S.J.; Kim, I.H.; Choi, Y.H.; Nam, T.J. Protective effect of porphyra-334 on UVA-induced photoaging in human skin fibroblasts. Int. J. Mol. Med. 2014, 34, 796–803. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.C.; Wu, W.T.; Lin, Y.J.; Weng, C.Y.; Tsai, M.H.; Tsai, H.T.; Chen, Y.C.; Yet, S.F.; Lin, P. Aryl hydrocarbon receptor activation-mediated vascular toxicity of ambient fine particulate matter: Contribution of polycyclic aromatic hydrocarbons and osteopontin as a biomarker. Part. Fibre Toxicol. 2022, 19, 43. [Google Scholar] [CrossRef]
- Yang, S.; Park, S.H.; Oh, S.W.; Kwon, K.; Yu, E.; Lee, C.W.; Son, Y.K.; Kim, C.; Lee, B.H.; Cho, J.Y.; et al. Antioxidant Activities and Mechanisms of Tomentosin in Human Keratinocytes. Antioxidants 2022, 11, 990. [Google Scholar] [CrossRef]
- Rawlings, A.V.; Harding, C.R. Moisturization and skin barrier function. Dermatol. Ther. 2004, 17 (Suppl. 1), 43–48. [Google Scholar] [CrossRef]
- Krutmann, J.; Liu, W.; Li, L.; Pan, X.; Crawford, M.; Sore, G.; Seite, S. Pollution and skin: From epidemiological and mechanistic studies to clinical implications. J. Dermatol. Sci. 2014, 76, 163–168. [Google Scholar] [CrossRef]
- Dijkhoff, I.M.; Drasler, B.; Karakocak, B.B.; Petri-Fink, A.; Valacchi, G.; Eeman, M.; Rothen-Rutishauser, B. Impact of airborne particulate matter on skin: A systematic review from epidemiology to in vitro studies. Part. Fibre Toxicol. 2020, 17, 35. [Google Scholar] [CrossRef]
- Piao, M.J.; Hyun, Y.J.; Cho, S.J.; Kang, H.K.; Yoo, E.S.; Koh, Y.S.; Lee, N.H.; Ko, M.H.; Hyun, J.W. An ethanol extract derived from Bonnemaisonia hamifera scavenges ultraviolet B (UVB) radiation-induced reactive oxygen species and attenuates UVB-induced cell damage in human keratinocytes. Mar. Drugs 2012, 10, 2826–2845. [Google Scholar] [CrossRef]
- Moore, C.; Gupta, R.; Jordt, S.E.; Chen, Y.; Liedtke, W.B. Regulation of Pain and Itch by TRP Channels. Neurosci. Bull. 2018, 34, 120–142. [Google Scholar] [CrossRef]
- Iftinca, M.; Defaye, M.; Altier, C. TRPV1-Targeted Drugs in Development for Human Pain Conditions. Drugs 2021, 81, 7–27. [Google Scholar] [CrossRef]
- Bickers, D.R.; Athar, M. Oxidative stress in the pathogenesis of skin disease. J. Investig. Dermatol. 2006, 126, 2565–2575. [Google Scholar] [CrossRef]
- Jenner, P. Oxidative stress in Parkinson’s disease. Ann. Neurol. 2003, 53 (Suppl. 3), S26–S36. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, S.; Lee, J.H.; Oh, S.W.; Yu, E.; Kwon, K.; Jang, S.J.; Shin, D.S.; Moh, S.H.; Lee, J. Anti-Pollutant Activity of Porphyra yezoensis Water Extract and Its Active Compound, Porphyra 334, against Urban Particulate Matter-Induced Keratinocyte Cell Damage. Mar. Drugs 2023, 21, 121. https://doi.org/10.3390/md21020121
Choi S, Lee JH, Oh SW, Yu E, Kwon K, Jang SJ, Shin DS, Moh SH, Lee J. Anti-Pollutant Activity of Porphyra yezoensis Water Extract and Its Active Compound, Porphyra 334, against Urban Particulate Matter-Induced Keratinocyte Cell Damage. Marine Drugs. 2023; 21(2):121. https://doi.org/10.3390/md21020121
Chicago/Turabian StyleChoi, Seoyoung, Jeong Hun Lee, Sae Woong Oh, Eunbi Yu, Kitae Kwon, Sung Joo Jang, Dong Sun Shin, Sang Hyun Moh, and Jongsung Lee. 2023. "Anti-Pollutant Activity of Porphyra yezoensis Water Extract and Its Active Compound, Porphyra 334, against Urban Particulate Matter-Induced Keratinocyte Cell Damage" Marine Drugs 21, no. 2: 121. https://doi.org/10.3390/md21020121
APA StyleChoi, S., Lee, J. H., Oh, S. W., Yu, E., Kwon, K., Jang, S. J., Shin, D. S., Moh, S. H., & Lee, J. (2023). Anti-Pollutant Activity of Porphyra yezoensis Water Extract and Its Active Compound, Porphyra 334, against Urban Particulate Matter-Induced Keratinocyte Cell Damage. Marine Drugs, 21(2), 121. https://doi.org/10.3390/md21020121