1. Introduction
Cardiovascular diseases (CVDs) are the leading cause of death worldwide [
1]. Several hypotheses have been implicated in the development and progression of CVDs, including lipid infiltration, thrombosis, damage response, oxidative stress, and inflammation. Among these, the lipid infiltration hypothesis is currently the most accepted, and this hypothesis argues that dyslipidemia plays a significant role in atherogenesis and progression [
2]. In response to this hypothesis, the first statin was developed to lower plasma lipid levels, marking a new era in the prevention and treatment of CVDs. Statins competitively inhibit the rate-limiting enzyme of cholesterol synthesis, the 3-hydroxy-3-methyl-glutaryl-CoA reductase (HMGCR), in vivo, thereby reducing endogenous cholesterol synthesis [
3]. As a result, the liver compensatorily increases the uptake of low-density lipoprotein cholesterol (LDL-C), effectively clearing LDL-C from the blood. However, statins can lead to severe adverse events, and some patients may experience various degrees of muscle damage, such as myositis, myonecrosis, rhabdomyolysis, and occasionally diabetes and possible hemorrhagic stroke [
4,
5]. Therefore, it is of great significance to develop novel hypolipidemic drugs.
The concept of Medicine Food Homology (MFH) refers to foods that exhibit medicinal properties and can also act as drugs [
6,
7]. Novel natural products from MFH with lipid-lowering activity have garnered significant attention, and their constituents can be classified into five main categories: polysaccharides, flavonoids, steroidal saponins, quinones, and alkaloids [
8]. Laminarin (
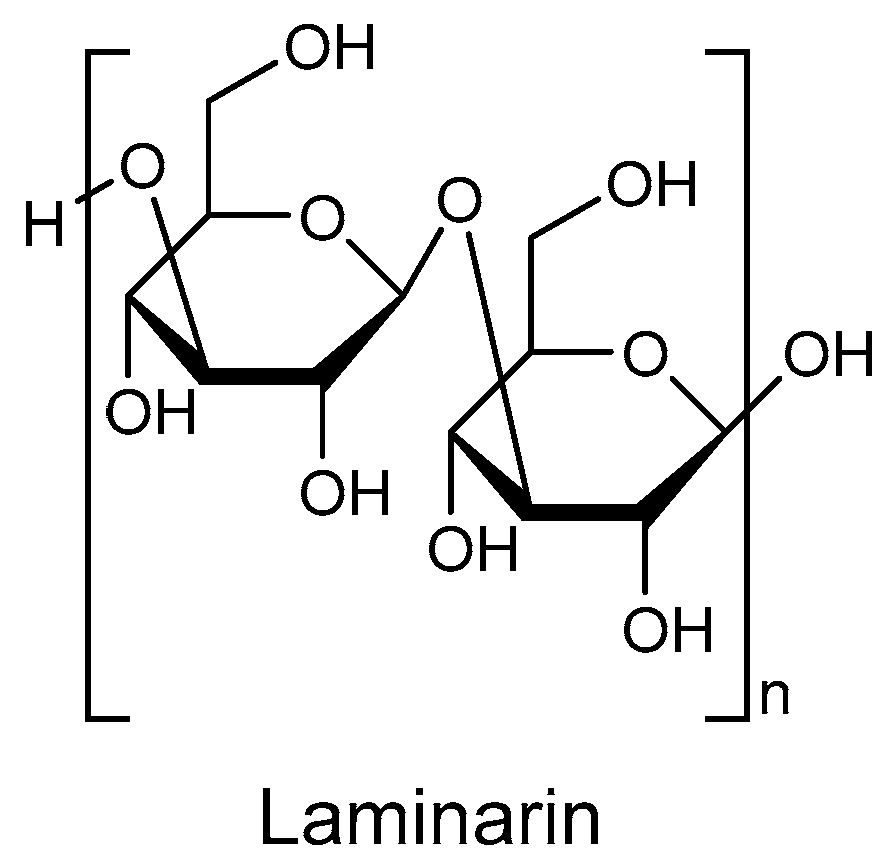
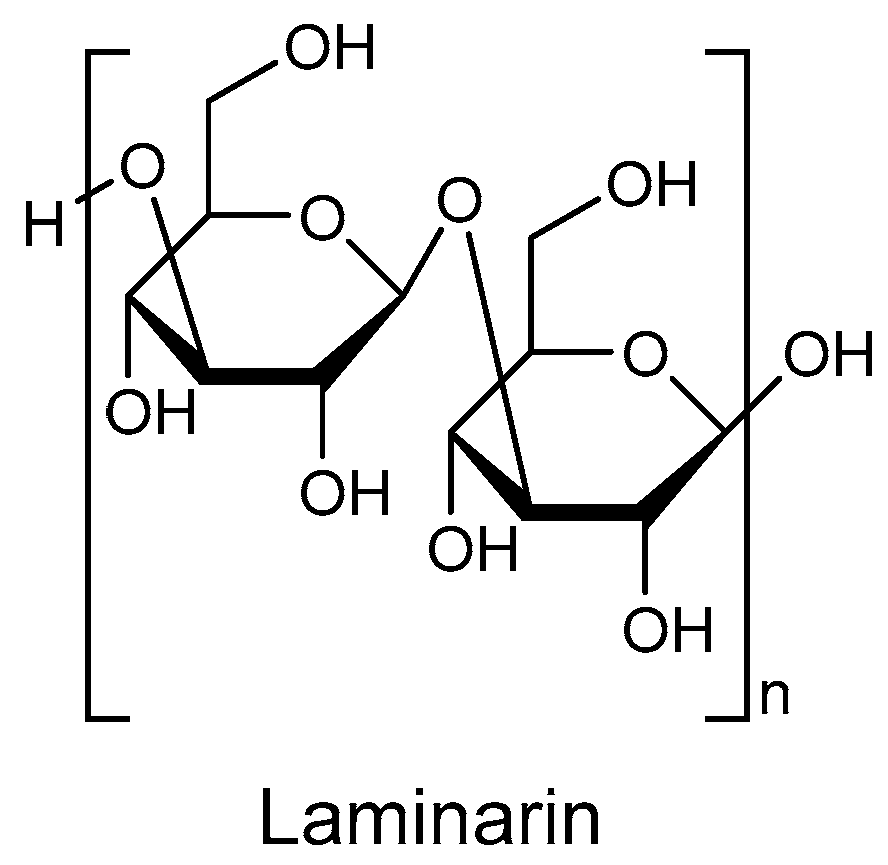
Figure 1), also known as kelch starch or brown algal starch, is a polysaccharide found in brown algae [
9]. Extensive literature has established that laminarin possesses biological activities, including immune modulation, blood glucose, and blood pressure reduction, as well as anti-tumor, anti-oxidant, anti-inflammatory, and skin barrier repair properties [
10,
11,
12,
13]. Furthermore, studies have shown that laminarin can reverse metabolic disorders such as dyslipidemia and atherosclerosis in mice induced by a high-fat diet (HFD) [
14,
15,
16]. Some reports also suggest that laminarin improves the lipid profile and ameliorates atherosclerosis in vivo through decreasing levels of total triglycerides (TG), total cholesterol (TC), LDL-C, while increasing levels of high-density lipoprotein cholesterol (HDL-C) [
17]. However, the specific mechanism of action of laminarin is still inconclusive.
Niemann–Pick C1-like 1 protein (NPC1L1) is a crucial transporter for cholesterol uptake and absorption at the villus brush border of the small intestine [
18,
19]. Its primary function is to facilitate the uptake of cholesterol from the small intestinal lumen into epithelial cells. Additionally, NPC1L1 promotes hepatic cholesterol storage and elevates lipid levels by facilitating cholesterol transport through the small intestine to the liver [
20,
21]. Ezetimibe is currently the first and only marketed inhibitor of NPC1L1 [
22]. By blocking this transporter, ezetimibe effectively inhibits cholesterol uptake in the intestine, reduces hepatic cholesterol storage, and, consequently, lowers plasma cholesterol levels [
23]. It has been reported that extracts from brown algae, particularly polysaccharides, have the ability to increase cholesterol excretion in feces [
17]. However, whether the cholesterol-lowering effect of brown algae polysaccharides is associated with NPC1L1 has not yet been clarified. Therefore, the purpose of this study is twofold: (i) to investigate whether laminarin, like ezetimibe, can inhibit cholesterol uptake and lower blood lipids by targeting NPC1L1; and (ii) to examine the mechanism underlying the blood lipid-lowering effect of laminarin by utilizing a HFD model of mice.
3. Discussion
This study aimed to investigate the lipid-lowering potential of laminarin, a natural bioactive molecule derived from edible brown algae. Our findings show that laminarin can counteract diet-induced hyperlipidemia by reducing intestinal cholesterol uptake, possibly through the down-regulation of NPC1L1 expression, a key transporter for cholesterol uptake.
Previous studies have shown that another polysaccharide from brown algae, fucoidan, inhibits atherosclerosis by regulating the expression of genes involved in cholesterol reverse transport (RCT), such as ABCA1, SR-A1, and PPAR [
24]. Fucoidan has also been found to reduce NPC1L1 expression, inhibit cholesterol uptake in the intestine, and prevent hyperlipidemia-induced atherosclerosis in apoE
−/− mice [
25]. To explore the effects of laminarin, also derived from brown algae, on RCT, we monitored cholesterol efflux in J774A.1 murine macrophage. However, laminarin did not show a significant effect on cholesterol efflux (
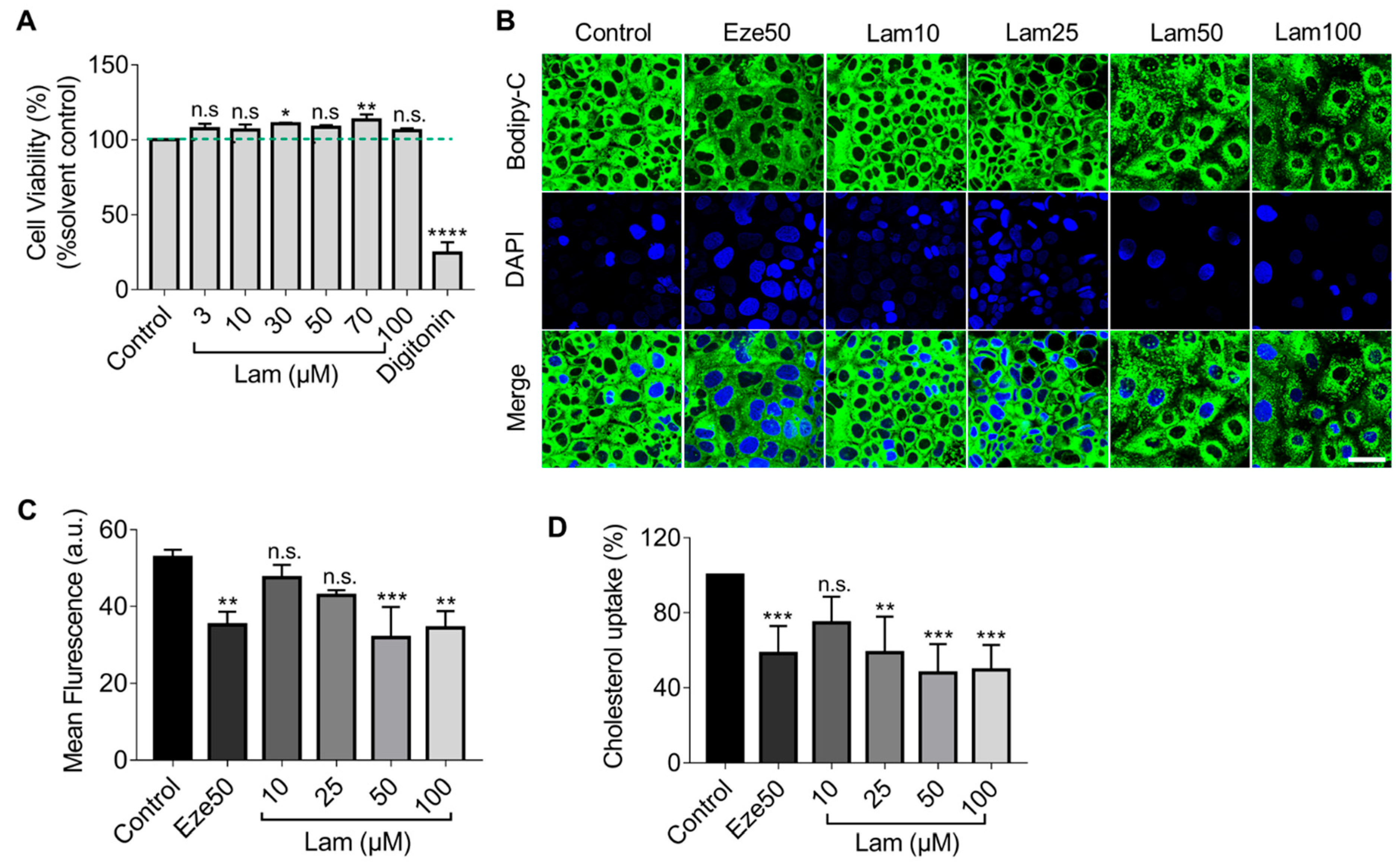
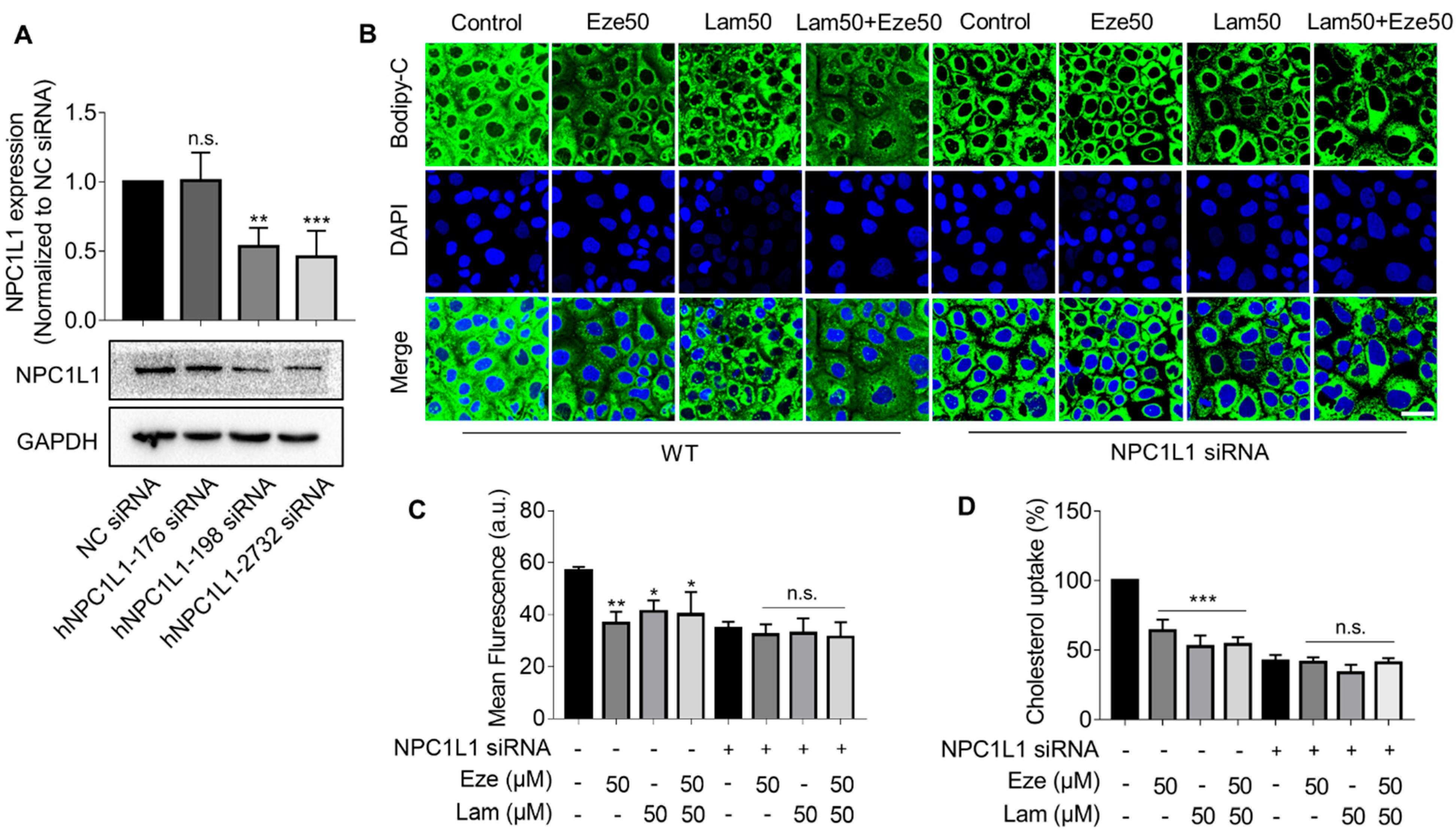
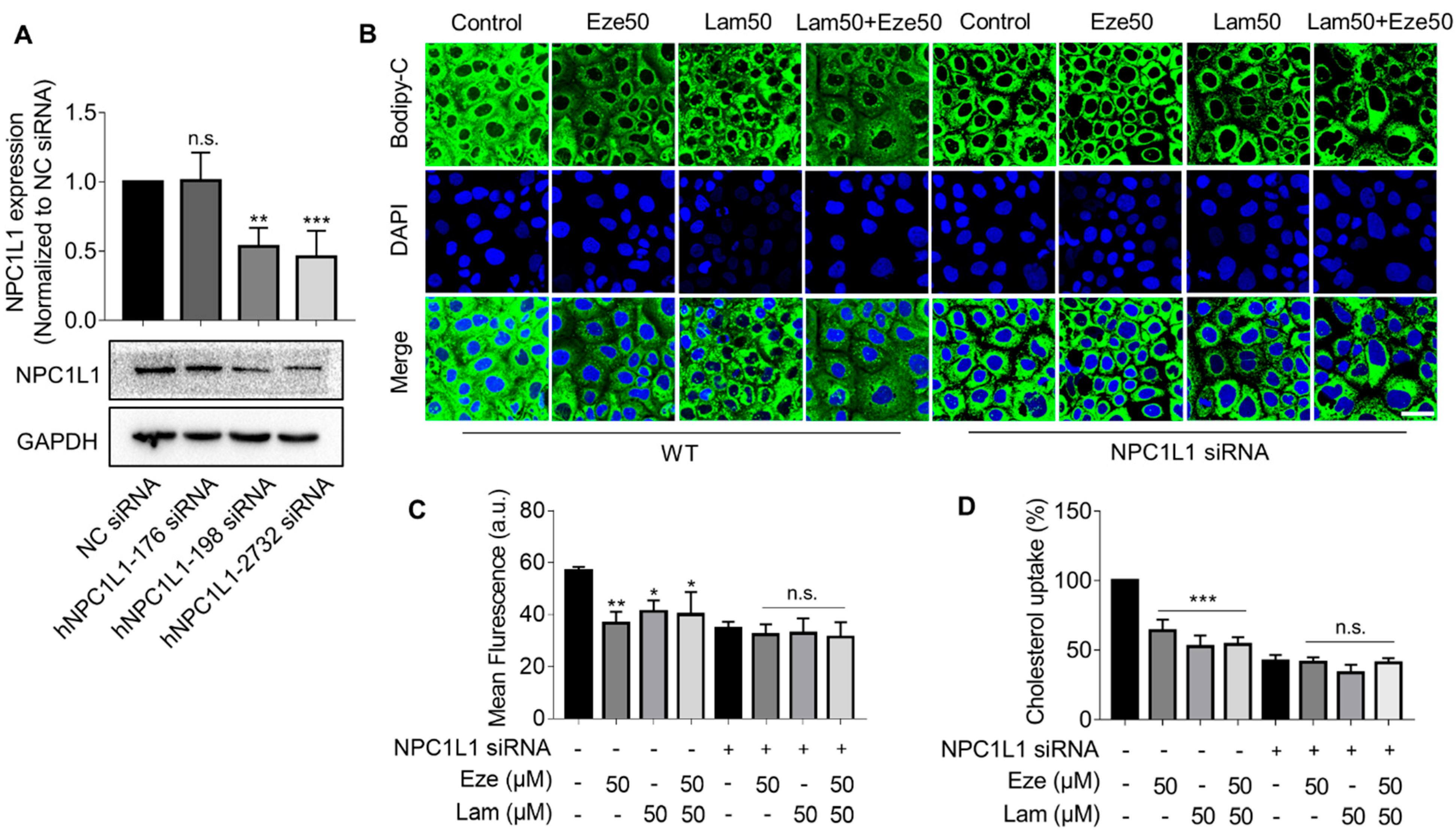
Figure S4). Nevertheless, when using Caco-2 cells to simulate cholesterol transport in the small intestinal epithelium, we found that laminarin effectively inhibited cholesterol uptake in a concentration-dependent manner. The results also showed that the effect of laminarin at 50 μM has reached a plateau, and even if the concentration continues to increase, the effect could not be further improved. That might be because there was a plateau in the dose-effect relationship of laminarin. When the compound reaches a certain threshold of concentration, further increasing its concentration will not improve its efficacy and may even produce toxic effects. Additionally, 50 μM laminarin and 50 μM ezetimibe exhibited competitive effects on cholesterol uptake in Caco-2 cells. These results led us to speculate that laminarin may act by inhibiting NPC1L1 in the small intestine.
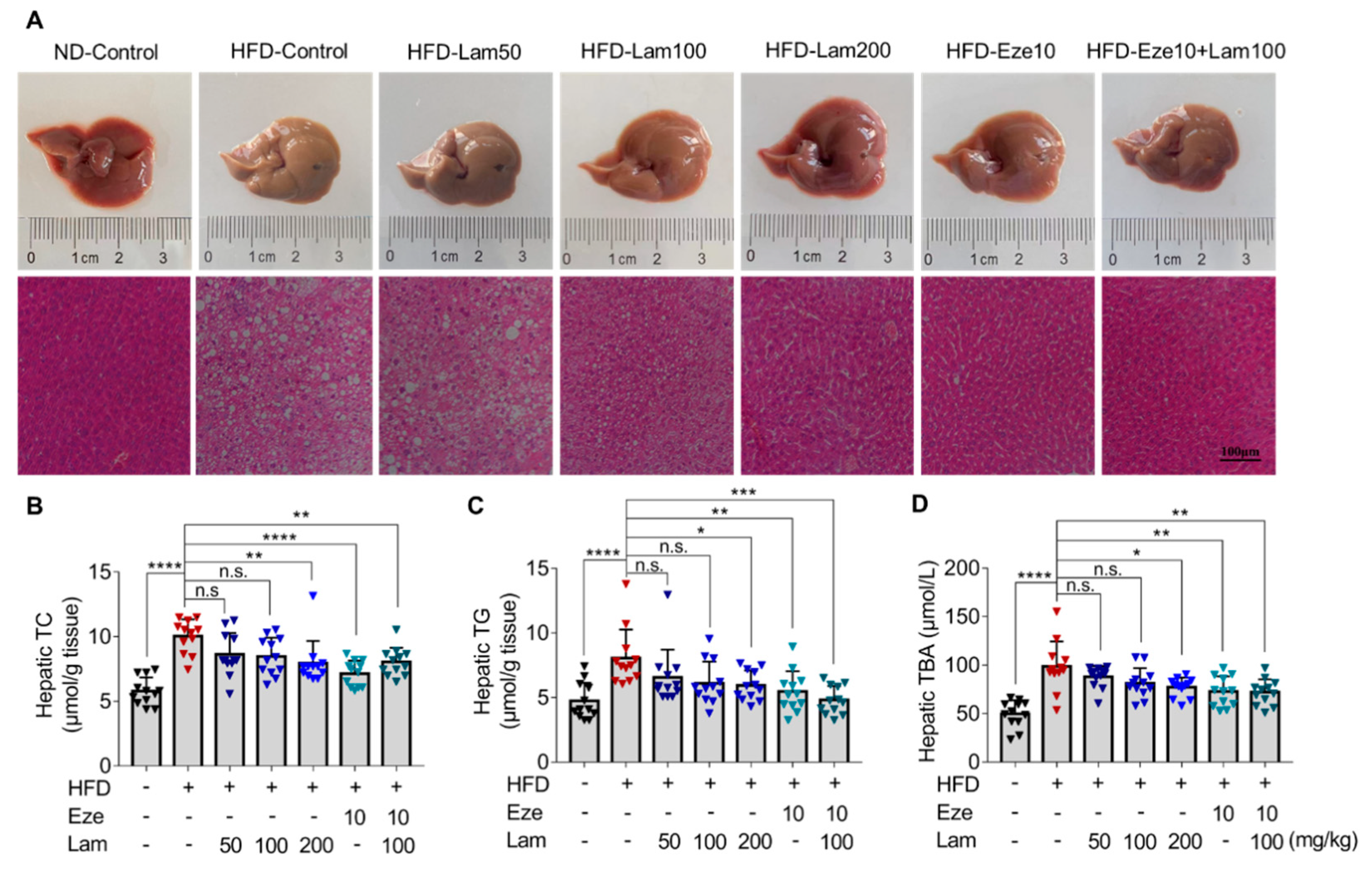
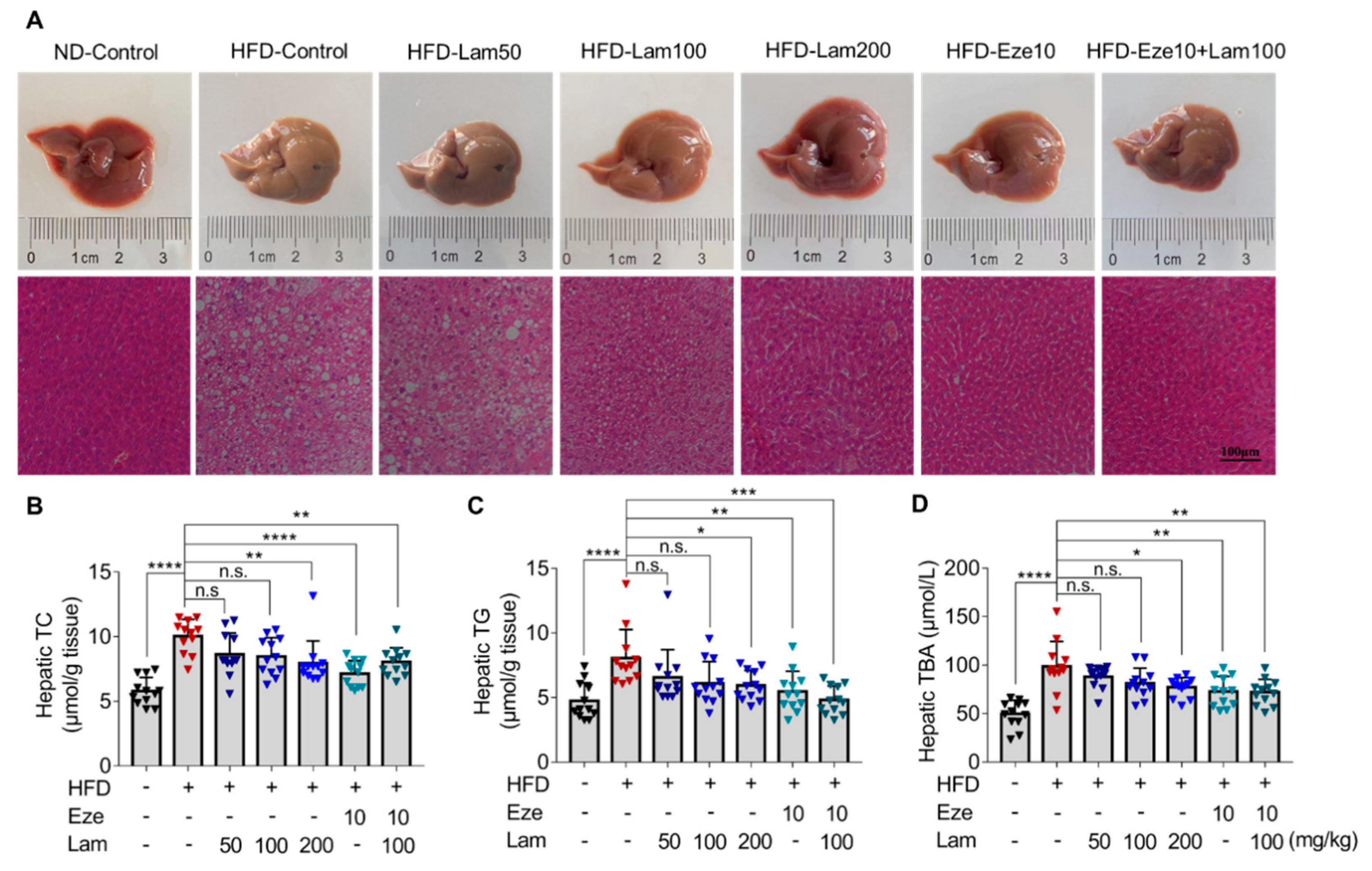
We then evaluated the lipid-lowering effect and mechanism of laminarin in vivo using HFD mice. Laminarin effectively reduced serum TC, TG, and LDL-C levels in mice. In this study, laminarin at 100 mg/kg with ezetimibe at 10 mg/kg rather than laminarin at 200 mg/kg with ezetimibe at 10 mg/kg was used to ascertain the full therapeutic potential of these compounds. One would expect that the combination of 200 mg/kg laminarin with 10 mg/kg ezetimibe may produce a stronger effect than 100 mg/kg laminarin with ezetimibe at 10 mg/kg. However, this only occurs when the binding sites of NPC1L1 are not fully saturated. Since laminarin and ezetimibe were speculated to act on the same target, they compete with each other for binding sites on NPC1L1. Therefore, there is a competitive inhibitory effect between them rather than a synergistic or additive effect. Unlike fucoidan [
26], laminarin did not affect HDL-C levels. This finding aligns with our hypothesis that laminarin shares mechanistic similarities with ezetimibe, as ezetimibe has also been shown to reduce intestinal cholesterol uptake without affecting serum HDL-C levels [
27]. The competitive inhibitory effect of laminarin and ezetimibe on TC and TG levels in vivo mirrored the results obtained in the in vitro experiments, except for LDL-C, where an additive inhibitory effect was observed. Polysaccharides have the ability to create a water layer barrier in the intestine and facilitate the excretion of bile acids. This process promotes the conversion of hepatic cholesterol into bile acids at an increased rate, thereby reducing the cholesterol content in the liver and lowering serum LDL-C concentration [
28,
29]. These findings suggest that the mechanisms of action of laminarin and ezetimibe are not identical, and that multiple effects may contribute to the observed in vivo results.
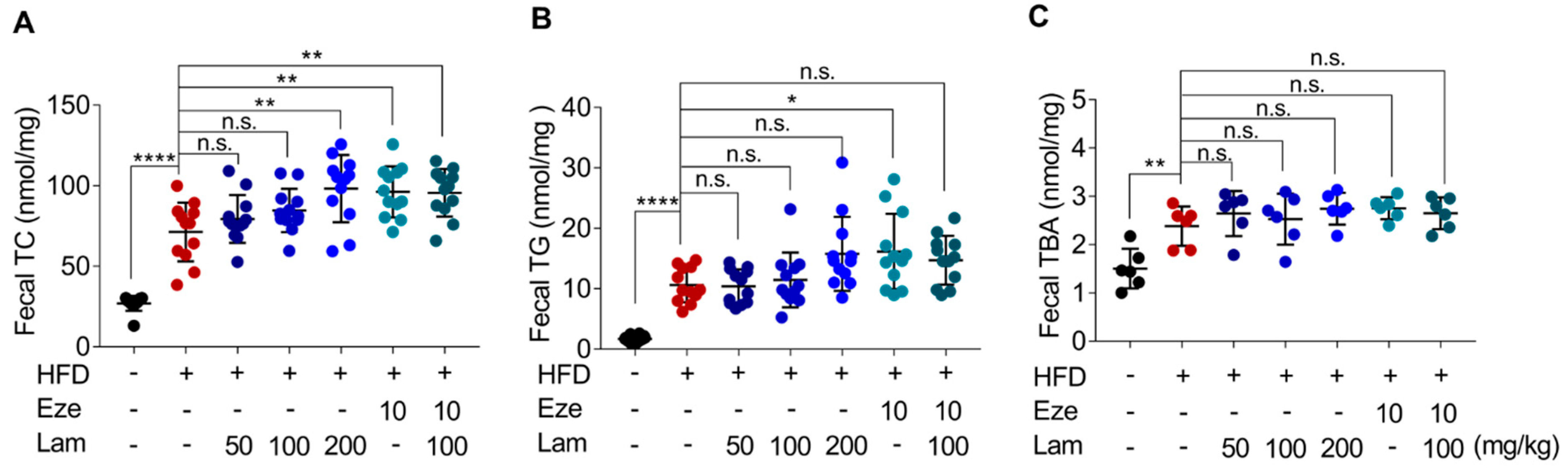
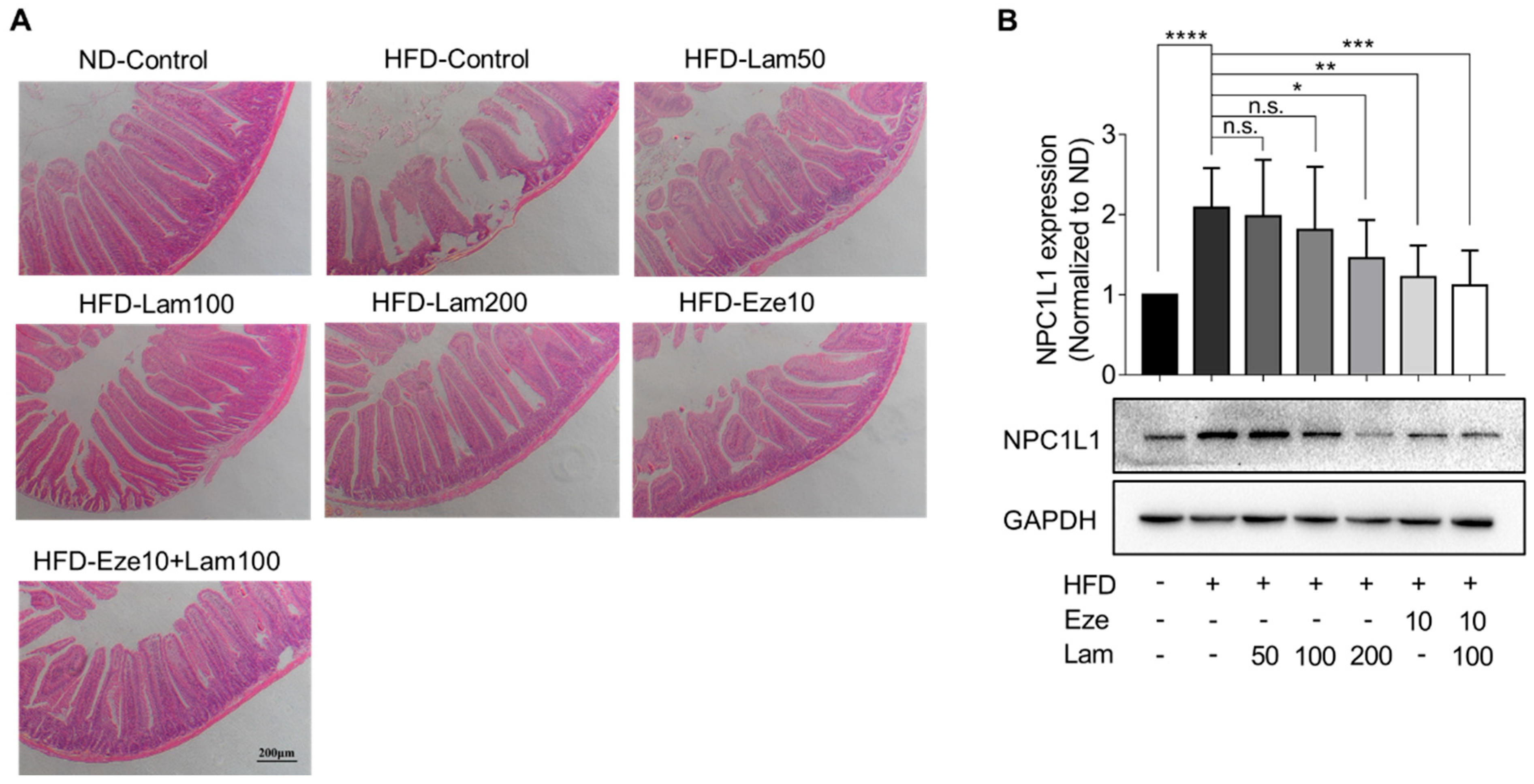
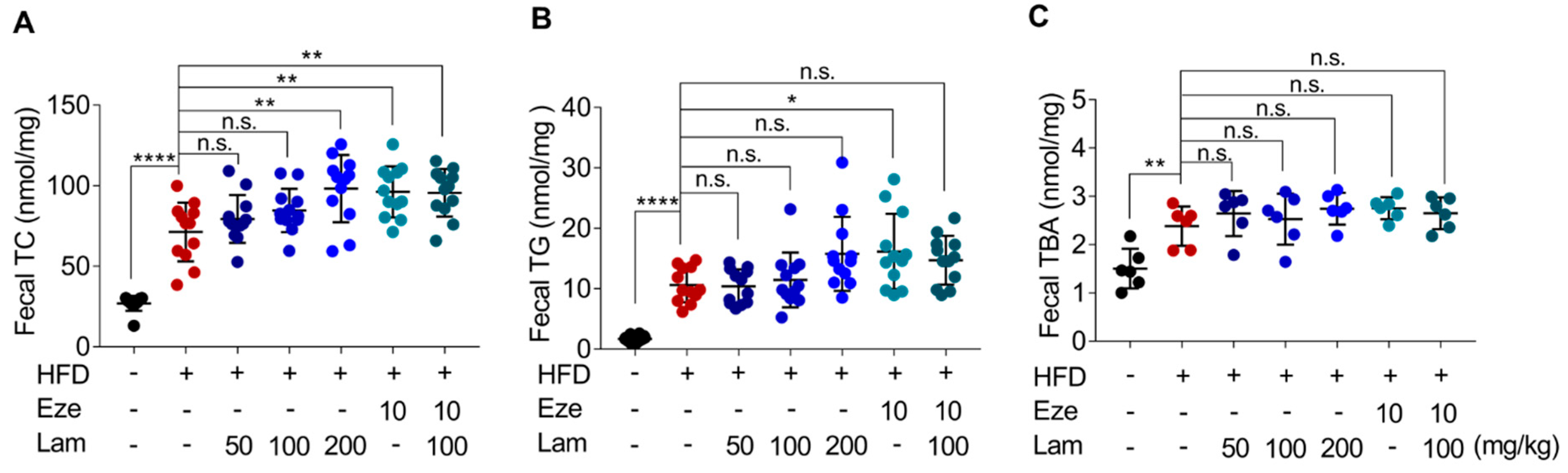
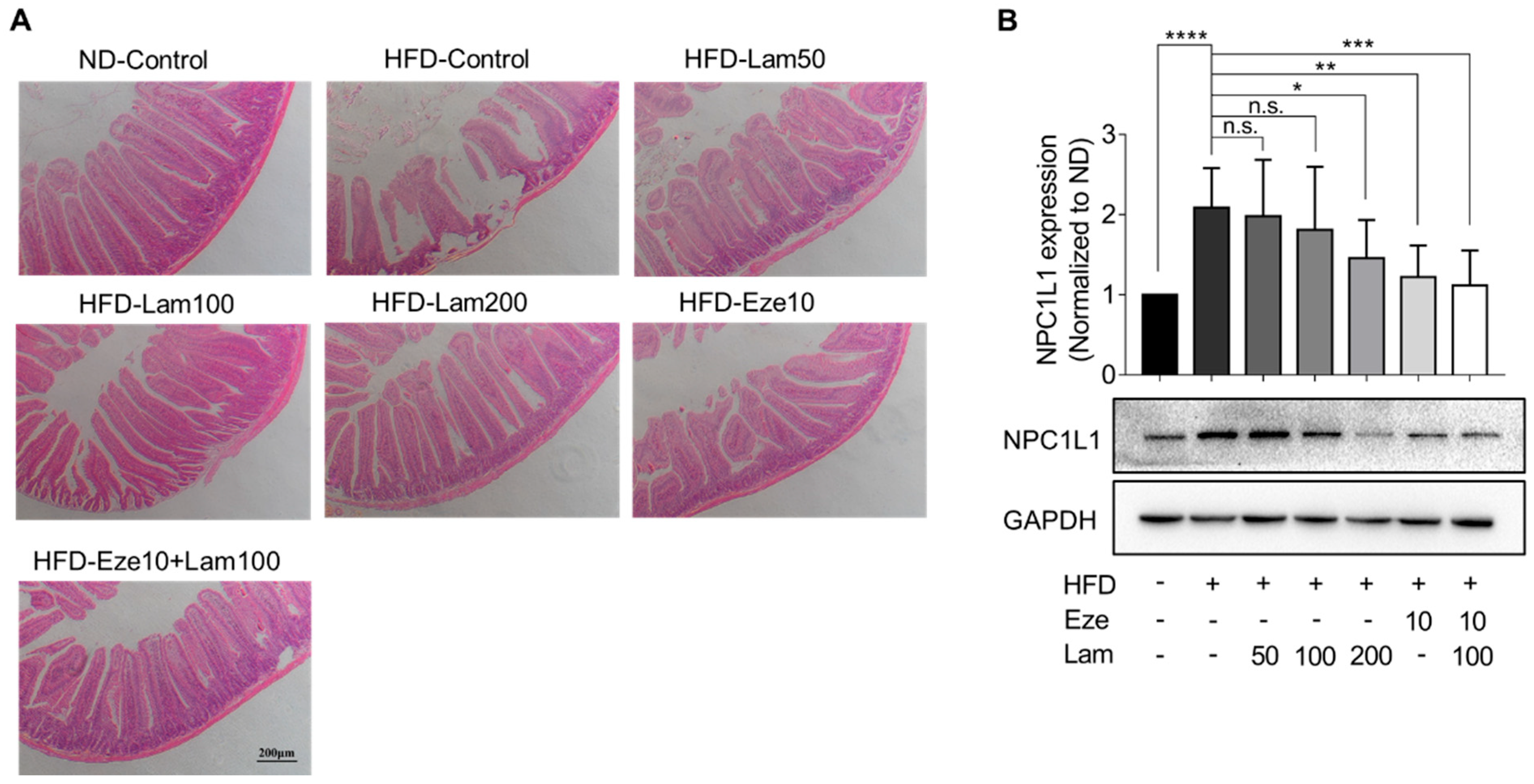
It is widely recognized that increased excretion in fecal lipids is associated with lower levels of serum lipids, which helps mitigate the accumulation of fat in the liver [
30]. Our results demonstrated that laminarin significantly increases fecal excretion of TC in mice, while not significantly affecting fecal levels of TG or TBA levels. In the mouse liver, laminarin effectively reduced the levels of TC, TG, and TBA and improved the diffuse vesicular steatosis of hepatocytes in HFD-mice. We also observed that laminarin significantly ameliorated jejunal villus breakage and intestinal structural damage. It also helped reduce the accumulation of subcutaneous and visceral fat in HFD-mice, leading to partial restoration of pathological changes. Hyperlipidemia has been associated with increased levels of TNF-α, which promotes a chronic inflammatory state and can impair intestinal barrier function [
31]. Therefore, it would be interesting to explore the potential effect of laminarin on intestinal inflammation in future research.
NPC1L1 is the most abundant protein in the epithelium of the proximal segment of the jejunum, which explains the active uptake of cholesterol by this part of the intestine, particularly the proximal jejunum, compared with other regions of the gut [
18,
32]. The lipid-lowering effect of ezetimibe is largely attributed to its inhibition of NPC1L1 transporter activity. In addition, ezetimibe has been shown to downregulate NPC1L1 protein expression [
20,
33], although this downregulation is not as direct and efficient as inhibiting the transporter activity itself. Similar to ezetimibe, our study showed that laminarin can downregulate NPC1L1 protein levels. This effect on the NPC1L1 transporter allows laminarin to rapidly and effectively reduce serum lipid levels induced by a HFD in mice. Considering the significant improvements observed in blood lipids, fecal lipids, and hepatic lipids due to laminarin treatment but the lack of inhibition in cholesterol efflux from macrophages (
Figure S4), which is an initial step in RCT, it is plausible to speculate that laminarin may also interact with other regulatory players involved in RCT, such as ABCG5/G8, to maintain cholesterol homeostasis. However, further experimental evidence is required to confirm this speculation.
4. Materials and Methods
4.1. Chemicals and Reagents
A small interfering RNA oligonucleotide duplex targeting NPC1L1 was designed and synthesized by Sangon Biotech Co., Ltd. (Shanghai, China). Laminarin (purity: ≥96%) and ezetimibe (purity: ≥99%) were acquired from Macklin Biochemical Technology Co., Ltd. (Shanghai, China). Fetal bovine serum (FBS) was supplied by PAN-SERATECH (Adenbach, Germany). Minimum essential medium (MEM), sodium pyruvate, MEM non-essential amino acids solution (NEAA), benzylpenicillin–streptomycin, lipofectamine 3000, minus serum medium (Opti-MEM), and PBS were obtained from Gibco (Waltham, MA, USA). Resazurin sodium salt, dimethyl sulfoxide (DMSO), and sodium carboxymethyl cellulose (CMC-Na) were purchased from Sigma-Aldrich (St. Louis, MO, USA), and digitonin was obtained from TOPSCIENCE Biotechnology Co., Ltd. (Shanghai, China). Modified Harris hematoxylin solution, eosin Y solution, poly-D-lysine and 4′,6-diamidino-2-phenylindole (DAPI) were bought from Solarbio Science & Technology Co., Ltd. (Beijing, China).
The normal diet and the customized high-fat diet were purchased from Keao Xieli Feed Co., Ltd. (Beijing, China). RIPA lysis and extraction buffer, halt protease and phosphatase inhibitor cocktail, protein ladder, NuPAGE LDS sample buffer, SuperSignal west pico plus chemiluminescent substrate, BCA protein assay kit, and histoplast paraffin were obtained from Thermo Scientific (Waltham, MA, USA). Polyvinylidene fluoride (PVDF) membrane was provided by GE Healthcare Life Sciences (Pittsburgh, PA, USA). Nonfat dry milk (NFDM) was purchased from Cell Signaling Technology (Beverly, MA, USA). The primary antibody against NPC1L1 was obtained from Novus Biologicals (Littleton, CO, USA). The anti-GAPDH antibody and goat anti-rabbit IgG H&L (HRP) were products of Abcam (Cambridge, UK). Assay kits for measuring total cholesterol (T-CHO), triglyceride (TG), low-density lipoprotein cholesterol (LDL-C), high-density lipoprotein cholesterol (HDL-C), and total bile acid (TBA) were purchased from JianCheng Bioengineering Institute (Nanjing, China).
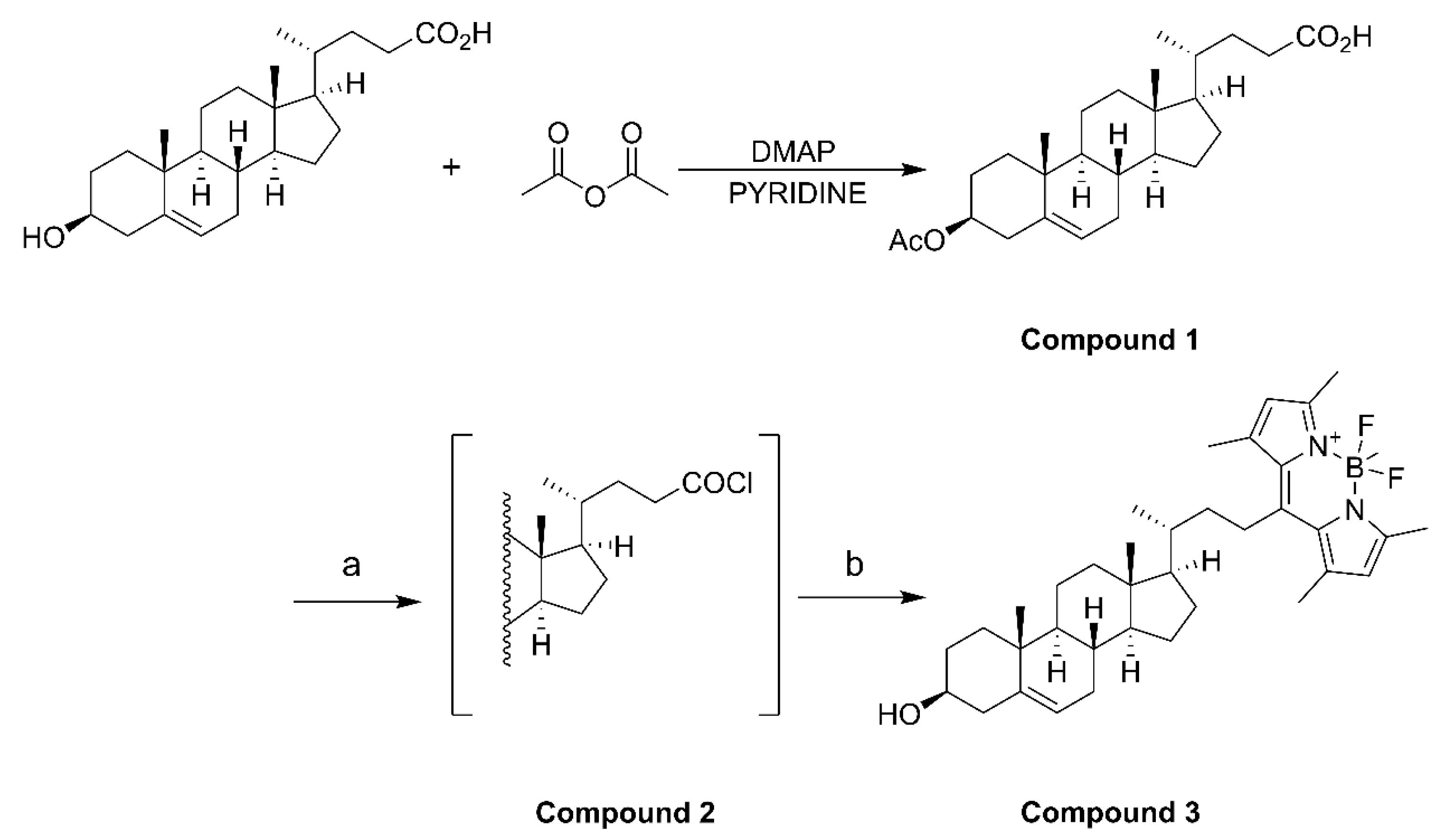
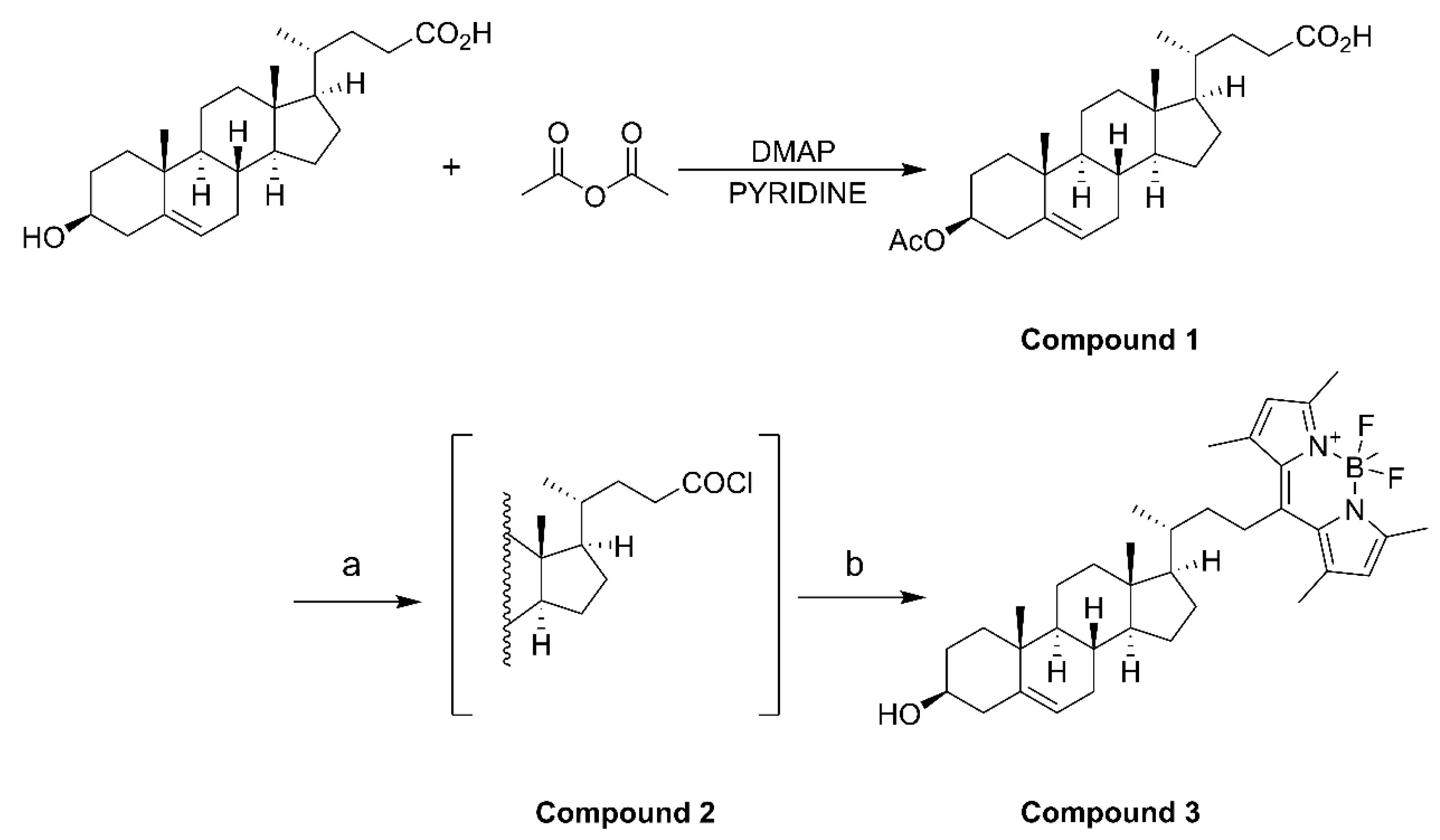
The synthesis of BODIPY-cholesterol involved the use of the following reagents: 3β-hydroxy-Δ5-cholenoic acid and 2,4-dimethylpyrrole were acquired from Macklin Biochemical Technology Co., Ltd. (Shanghai, China). 4-dimethylaminopyridine was purchased from Bide Pharmaceutical Technology Co., Ltd. (Shanghai, China). Boron trifluoride diethyl ether, pyridine, oxalyl chloride, and anhydrous Na2SO4 were acquired from Aladdin Bio-Chem Technology Co., Ltd. (Shanghai, China). Muriatic acid, ethyl acetate, dichloromethane, methyl alcohol, petroleum ether, and acetic anhydride were purchased from China National Pharmaceutical Group Co., Ltd. (Beijing, China). Triethylamine and dimethyl sulfoxide-d6 were purchased from Titan Scientific Co., Ltd. (Shanghai, China).
4.2. Fluorescent BODIPY-Cholesterol Synthesis
Acetic anhydride (1.66 mmol), 3β-hydroxy-Δ5-cholenoic acid (1.33 mmol), and 4-dimethylaminopyridine (0.133 mmol) were dissolved in pyridine (10 mL) and stirred for 4 h at room temperature. After the reaction, iced water (10 mL) was added to quench the reaction. Then, 3 N HCl (3 mmol) was added dropwise with stirring to adjust the pH of the solution from 5 to 2. The mixture was extracted with ethyl acetate (3 × 20 mL), and the organic phases were combined before being washed with saturated aqueous NaCl (2 × 20 mL) and filtered and dried over anhydrous Na
2SO
4. After concentration, compound
1 was isolated as white solid powder by column chromatography using CH
2Cl
2/MeOH (60:1) as the eluent. Compound
1 (0.82 mmol) was dissolved in CH
2Cl
2 (30 mL) in an ice bath under nitrogen, and oxalyl chloride (3.28 mmol) was added. The mixture was stirred overnight at room temperature, and compound
2 was obtained as a white solid after concentration. Compound
2 was dissolved in CH
2Cl
2 (10 mL), and 2,4-dimethylpyrrole (1.804 mmol) was added. The reaction mixture was heated under reflux under nitrogen for 4 h. After cooling to room temperature, boron trifluoride diethyl ether (4.92 mmol) and triethylamine (3.28 mmol) were added, and the mixture was stirred under nitrogen overnight. After completion of the reaction, 5 mL of water was added, and then the mixture was extracted before the organic phase was collected and dried. The filtrate was concentrated and purified by column chromatography using EtOAc/petroleum ether (1:10) as the eluent. A total of three synthesis cycles resulted in a yield of 608.8 mg of compound
3 (BODIPY-cholesterol) with fluorescence activity (
Figure S1), yielding approximately 65% to 79% (
Scheme 1).
3β-Acetoxy-23-(4,4-difluoro-1,3,5,7-tetramethyl-4-bora-3a,4a-diaza-sindacene-8-yl)-24-norchol-5-ene (compound 3, BODIPY-cholesterol): 1H NMR (500 MHz, DMSO-d6) δ 6.06 (s, 2H), 5.38 (d, J = 5.0 Hz, 1H), 4.62 (dd, J = 11.4, 6.1 Hz, 1H), 3.16 (td, J = 12.9, 5.5 Hz, 1H), 2.72 (td, J = 12.8, 3.1 Hz, 1H), 2.53 (s, 5H), 2.44 (s, 5H), 2.34 (s, 1H), and 2.06 (s, 1H).
4.3. Cell Culture and Resazurin Conversion Assay
Caco-2 cells were obtained from the Cell Bank of the Chinese Academy of Sciences and cultured in 25 cm2 cell culture flasks with minimum essential medium supplemented with 1 mM NEAA, 1 mM sodium pyruvate, 100 U/mL benzylpenicillin, 100 µg/mL streptomycin, and 20% FBS. Cells were maintained at 37 °C with 5% CO2 in a humidified atmosphere. For the experiments, Caco-2 cells were seeded at a density of 0.2 × 106 per mL in 6-well-plates or 96-well-plates for experiments.
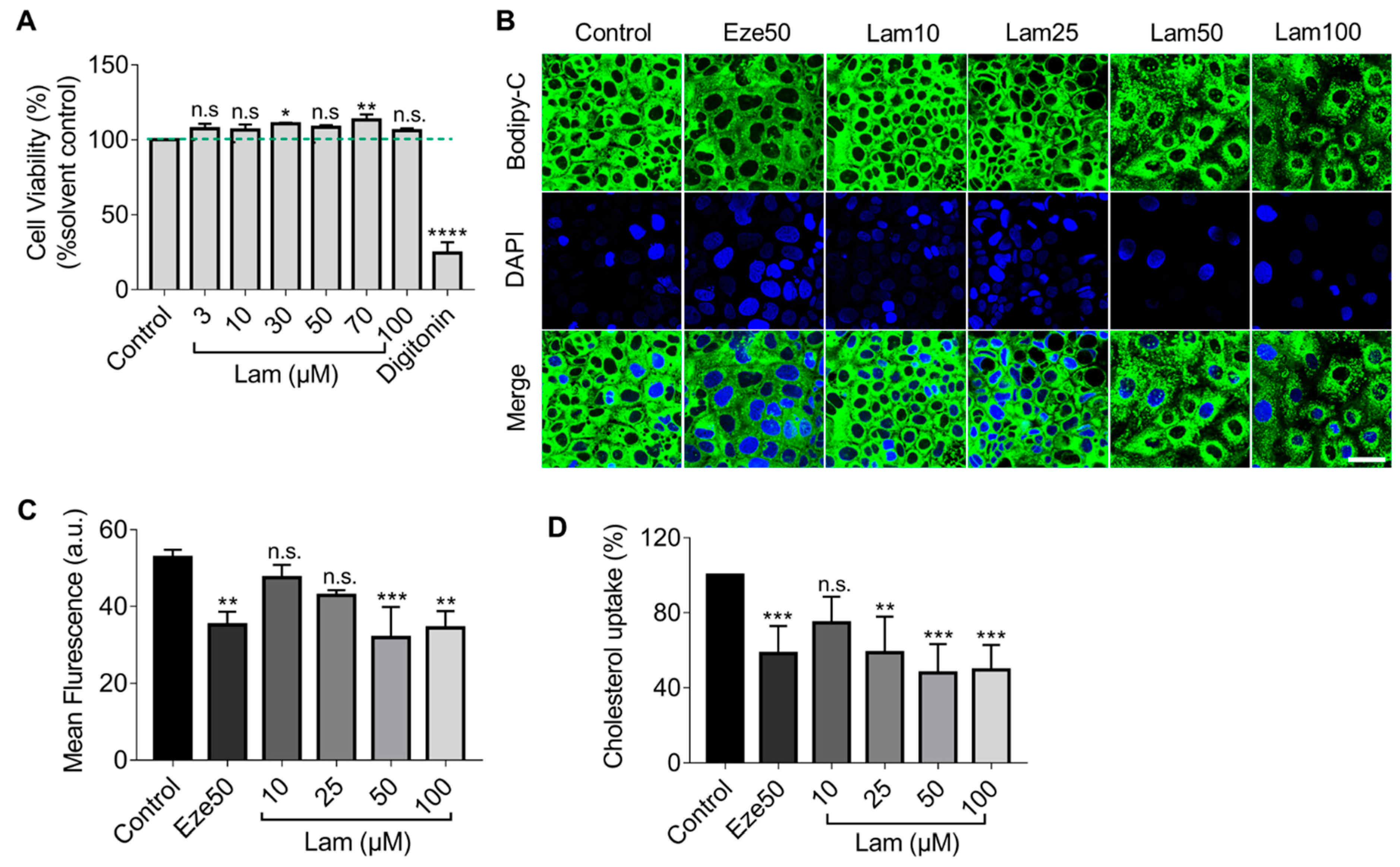
Caco-2 cells were treated with digitonin (50 µg/mL) and laminarin (3, 10, 30, 50, 70, and 100 µM) for 24 h. After incubation, cells were washed with PBS and incubated with resazurin for another 4 h. Fluorescence emitted by the generated resorufin was quantified using excitation/emission wavelengths of 535/580 nm as a measurement of cell viability.
4.4. Cholesterol Uptake Assay
First, Caco-2 cells in the logarithmic growth phase were harvested and seeded at a density of 0.2 × 106 cells/mL in a 96-well-plate in a total volume of 100 μL medium/well using a Vi-Cell XR cell counter (Beckman Coulter, Indianapolis, IN, USA). After 24 h incubation, Caco-2 cells were treated with ezetimibe (50 µM), various concentrations of laminarin (5, 10, 25, 50, 100, and 150 µM) and a solvent vehicle control (DMSO) for 24 h. Then, the medium was discarded and washed once with PBS to remove non-adherent or dead cells before being replaced with the MEM medium containing 0.2% BSA. Caco-2 cells were labeled with BODIPY-cholesterol (0.0625 mM) and treated with ezetimibe (50 µM), various concentrations of laminarin (5, 10, 25, 50, 100, and 150 µM) and a solvent vehicle control (DMSO) for another 24 h. After the incubation, the extracellular fluid was discarded, and 100 µL of 10% SDS were added to each well before being shaken for 6 min, and 80 µL of cell lysates were collected into a black 96-well plate. The fluorescence value (Ex/Em = 482/515 nm) was detected using the Flexstation3 (Molecular Devices, San Jose, CA, USA), and the fluorescence values were normalized to the cell number in each well.
4.5. Rapid High-Resolution Confocal Microscope Imaging
Caco-2 cells were seeded at a density of 1 × 105 cells/mL in a 35 mm laser confocal culture dish coated with 0.05 mg/mL poly-D-lysine. After incubation for 24 h, Caco-2 cells were treated with ezetimibe (50 µM), various concentrations of laminarin (5, 10, 25, 50, 100, and 150 µM) and a solvent vehicle control (DMSO) for 24 h. Then, the medium was discarded and washed once with PBS to remove non-adherent cells or dead cells before replacement of the MEM medium containing 0.2% BSA. Caco-2 cells were labeled with BODIPY-cholesterol (0.0625 mM) and treated with ezetimibe (50 µM), various concentrations of laminarin (5, 10, 25, 50, 100, and 150 µM) and a solvent vehicle control (DMSO) for another 24 h. Then the medium was discarded, and cells were fixed with paraformaldehyde cell fixative for 20 min. After fixation, 10 µg/mL of DAPI was added to cells for nuclear staining. After 10 min, the supernatant was discarded, and cells were washed with PBS and imaged with a confocal microscope (Leica STELLARIS 5, Leica, Wetzlar, Germany).
4.6. Design and Synthesis of siRNA
A small interfering RNA oligonucleotide duplex targeting NPC1L1
Table 1 was designed and synthesized by Sangon Biotech Co., Ltd. (Shanghai, China).
Table 1.
Genetic sequence of NPC1L1 siRNA oligonucleotide duplex.
Table 1.
Genetic sequence of NPC1L1 siRNA oligonucleotide duplex.
| Gene ID | Genetic Sequence |
|---|
| Sense (5′-3′) | Antisense (5′-3′) |
|---|
| hNPC1L1-176 | CUAUGACGAAUGUGGGAAGAATT | UUCUUCCCACAUUCGUCAUAGTT |
| hNPC1L1-2732 | CCUCUUUCUGAACCGCUACUUTT | AAGUAGCGGUUCAGAAAGAGGTT |
| hNPC1L1-1980 | GCCACCAGCUACAUUGUCAUATT | UAUGACAAUGUAGCUGGUGGCTT |
| NC | UUC UCC GAA CGU GUC ACG UTT | ACG UGA CAC GUU CGG AGA ATT |
| FAM-NC | UUC UCC GAA CGU GUC ACG UTT | ACG UGA CAC GUU CGG AGA ATT |
4.7. Transfection of Caco-2 Cells
Caco-2 cells (0.2 × 106 cells/well) were plated on 6 well-plates and incubated for 24 h. Cells were transfected with small interfering RNA (siRNA) against NPC1L1 (200 nM) using Lipofectamine 3000 and minus serum medium (Opti-MEM) according to the manufacturer’s recommendations. Cells were harvested 48 h after transfection for further experiments.
4.8. Animal Experiments
Eighty-four male C57BL/6J mice (pathogen-free; 6 weeks of age) were purchased from Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China). Throughout the study, the animals were housed in plastic mouse cages at a controlled temperature of 23 ± 2 °C with 12/12 h light-dark cycle and 50 ± 10% humidity. Mice had free access to water and food. The in vivo study was conducted in accordance with the Animal Center of Medical College of Qingdao University according to the guidelines and regulations of the Laboratory Animal Care and Use Committee of Qingdao University (protocol code: No.20220504C579120221220051, 21 April 2022). The mice were acclimated for 1 week to stabilize their metabolic conditions.
Mice were monitored for body weight and food intake. Mice were fed either a normal diet (ND) or a high-fat diet (HFD). HFD consists of 10% lard, 0.5% bile salt (pig), and 3% cholesterol mixed with ND, as formerly described [
34]. After feeding the mice with ND or HFD for 14 weeks, blood samples were collected from the tail vein, and blood lipid biochemical indexes were determined to confirm the establishment of a hyperlipidemia model. Then, the mice were randomly divided into seven groups, 12 for each group, and the treatment dose of compounds was determined according to previous studies [
16,
35]. The seven treatment groups were as follows: ND, control group; HFD, control group; HFD-ezetimibe group (10 mg/kg/day); and HFD high-dose laminarin (200 mg/kg/day); HFD medium-dose laminarin (100 mg/kg/day); HFD low-dose laminarin (50 mg/kg/day); and HFD drug combination group (Ezetimibe: 10 mg/kg/day and laminarin: 100 mg/kg/day). The mice in the vehicle control group were given the same volume of 0.3% CMC-Na by gavage, and all animals in all groups were gavaged once a day for 8 weeks with their respective treatments. During the treatment period, food intake was recorded daily, and the mouse weight was measured weekly. At the end of the experiment, fresh feces were collected and immediately stored at −80 °C for subsequent analysis. Mice were fasted for 12 h, blood samples were collected, centrifuged at 1342×
g for 15 min at 4 °C, and serum was stored at −80 °C for further analysis. After the mice were euthanized by isoflurane anesthesia, samples such as liver and jejunum were dissected for subsequent analysis.
4.9. Analysis of Serum, Hepatic and Fecal Lipids
After a 12 h fasting period, mice were anesthetized, and blood samples were collected through eye bleeding, and kept motionless for 30 min. The whole blood was centrifuged at 1342× g for 15 min to separate the serum. The serum levels of TG, TC, TBA, HDL-C, and LDL-C were determined in a microplate reader (Tecan SUNRISE, Grödig, Austria) using corresponding commercial assay kits. Liver and feces samples were accurately weighed and mechanically homogenized under ice-water bath conditions. Then, the homogenized samples were centrifuged at 685× g for 10 min, and the resulting supernatant was used to measure the same biochemical markers mentioned above using the corresponding assay kits.
4.10. Hematoxylin and Eosin (H&E) Staining
After the mice were sacrificed, a 0.5 cm section of jejunal tissue and a lobular section of the liver were fixed in 4% paraformaldehyde for at least 24 h. For dehydration of specimens, graded alcohols (70~100%, 5% a grade) were applied before clearing and infiltration in xylene. Tissue sections were embedded in paraffin wax and cut into 6 μm thick using a rotary microtome (Leica RM2255, Leica, Wetzlar, Germany). Tissue sections were attached to poly-L-lysine-treated glass slides. After deparaffinization in xylene and rehydration in graded alcohols (100~70%), tissue samples were stained with modified Harris hematoxylin solution for 6 min and counterstained with Eosin Y solution for 3 min. Excessive eosin of tissue sections was removed with alcohol and subsequent clearing in xylene. Finally, images of the tissue sections were captured using a microscope (Nikon Eclipse TS100, Nikon, Tokyo, Japan).
4.11. Protein Extraction and Western Blotting
Jejunal tissue samples were homogenized in RIPA lysis and extraction buffer containing 1% protease and phosphatase inhibitor cocktail (Thermo Scientific, Waltham, MA, USA) using a high flux tissue grinder (SCIENTZ-48, SCIENTZ, Zhejiang, China). After full lysis for 30 min at 4 °C using a rotary mixer (MIULAB, MIULAB, Hangzhou China), the tissue lysate was centrifuged at 15,777× g for 15 min to collect the supernatant. Total protein concentrations in the supernatant were detected using a BCA protein analysis kit. The protein samples were separated in 10% SDS-PAGE gels in tris-glycine buffer and then transferred to PVDF membranes. The PVDF membrane was blocked in TBS-T (24.9 mM Tris, 137 mM NaCl, 2.7 mM KCl, 0.1% Tween-20, pH 7.4) with 5% nonfat dry milk for 1 h at room temperature, followed by incubated with primary antibody at 4 °C overnight. The primary antibodies against NPC1L1 (#NB400-127), and GAPDH (#ab181602) were used. Membranes were incubated with a goat anti-rabbit IgG H&L (HRP) at room temperature for 1 h. Protein bands were visualized by using a chemiluminescent substrate kit. Finally, Image Lab software (Bio-Rad Laboratories, version 5.2.1, Hercules, CA, USA) was used to quantify the relative protein expression levels. The target protein abundance was normalized to the reference protein GAPDH, and then the values of all groups were normalized to the ND group.
4.12. Living Small Animal CT Imaging
After eight weeks of laminarin treatment, a subgroup of three mice from each group was randomly selected and anesthetized with inhaled isoflurane gas using a small animal anesthesia machine (RWD R500, Shenzhen, China). After the animals were completely anesthetized, a whole-body scan was conducted using a Living small animal CT instrument (PerkinElmer Quantum GX2, Waltham, MA, USA), and images of the subcutaneous and visceral fat were captured.
4.13. Statistical Analysis
All data were presented as the mean ± SD, and statistical analysis was performed using GraphPad Prism 7.00 software. Student’s t-tests and one-way ANOVA corrected by the Bonferroni test were used to assess statistical differences between groups, and a value of p < 0.05 was considered statistically significant.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}