
Mouse N2a Neuroblastoma Assay: Uncertainties and Comparison with Alternative Cell-Based Assays for Ciguatoxin Detection
 , , , , , and
, , , , , and 
Abstract
:
1. Introduction
2. Results
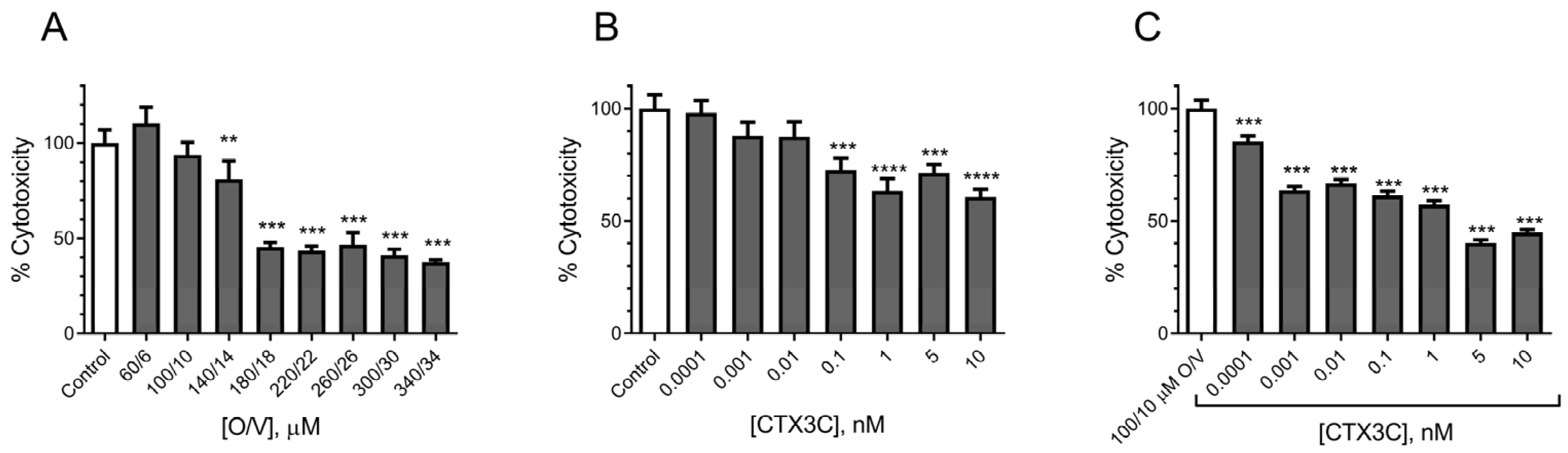
2.1. Action of CTXs in the N2a Cell Line
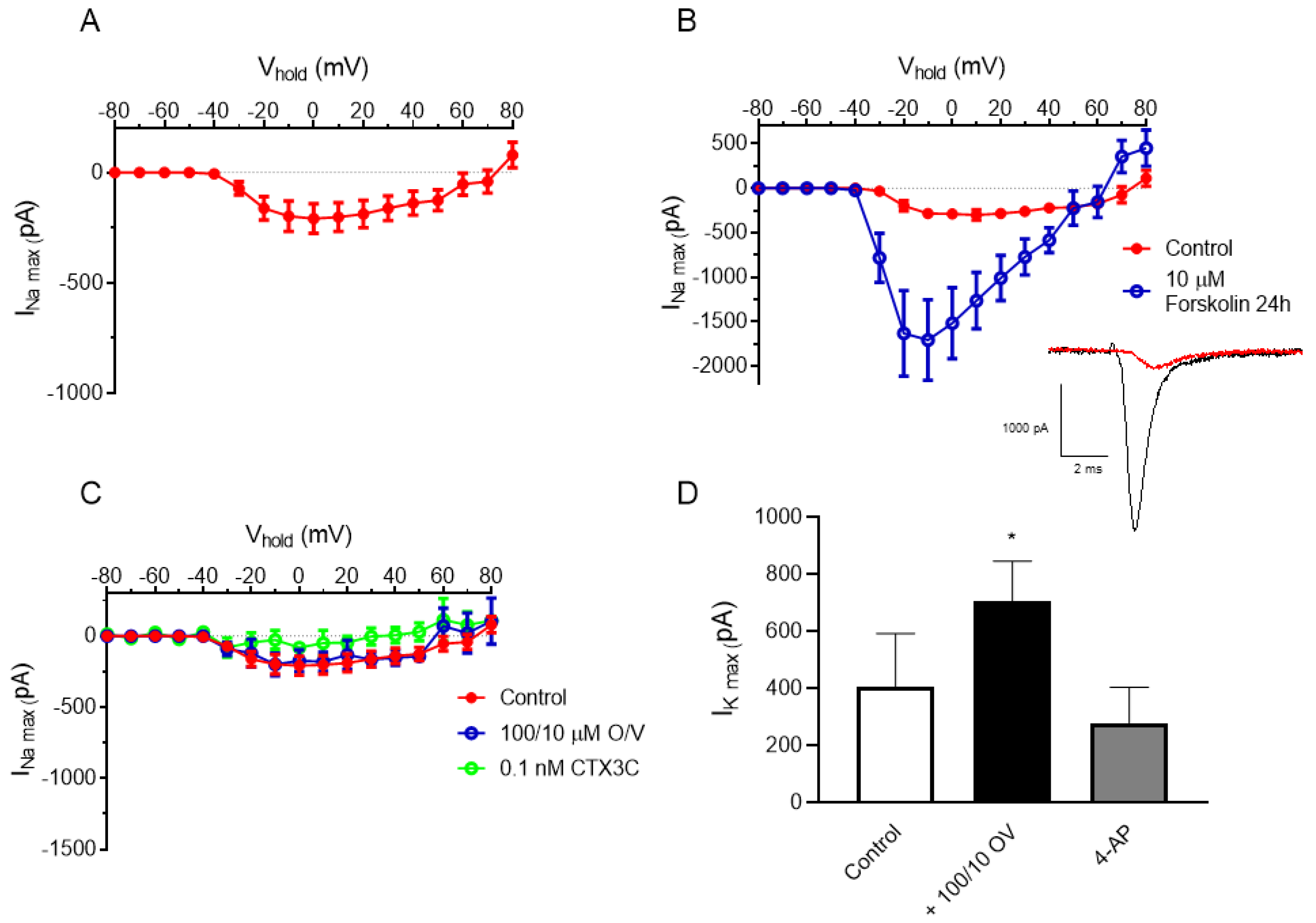
2.1.1. Electrophysiological Recordings of Sodium Currents in N2a Cells
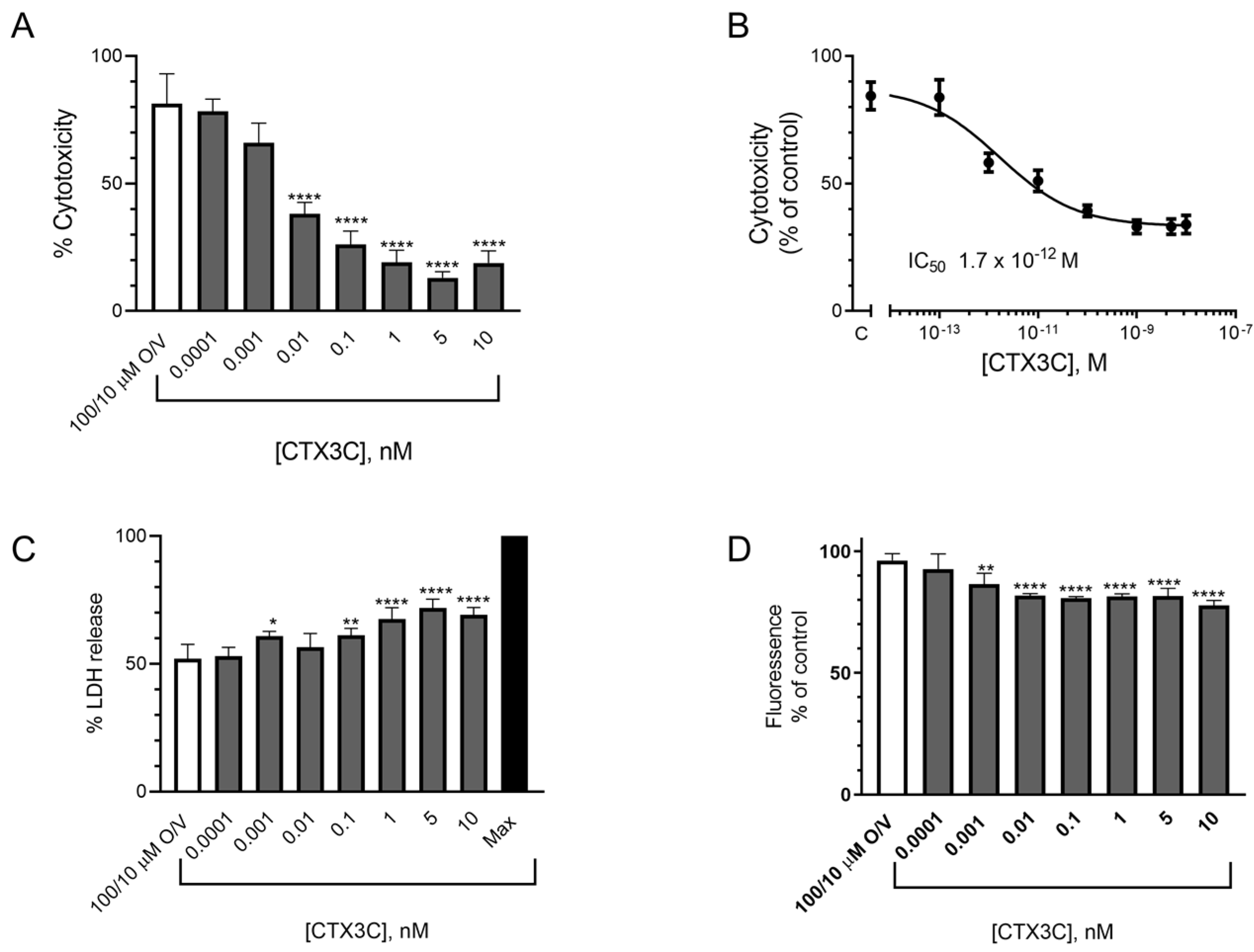
2.1.2. Evaluation of the Effect of CTX3C Using the MTT Assay on the N2a Cell Line
2.1.3. Evaluation of the Effect of CTX3C Using the LDH Test in the N2a Cell Line
2.1.4. Alamar Blue
2.1.5. Direct Effects of CTX3C and Ouabain/Veratridine in the N2a Actin Cytoskeleton
2.2. Increasing N2a Sensitivity to Ciguatoxins
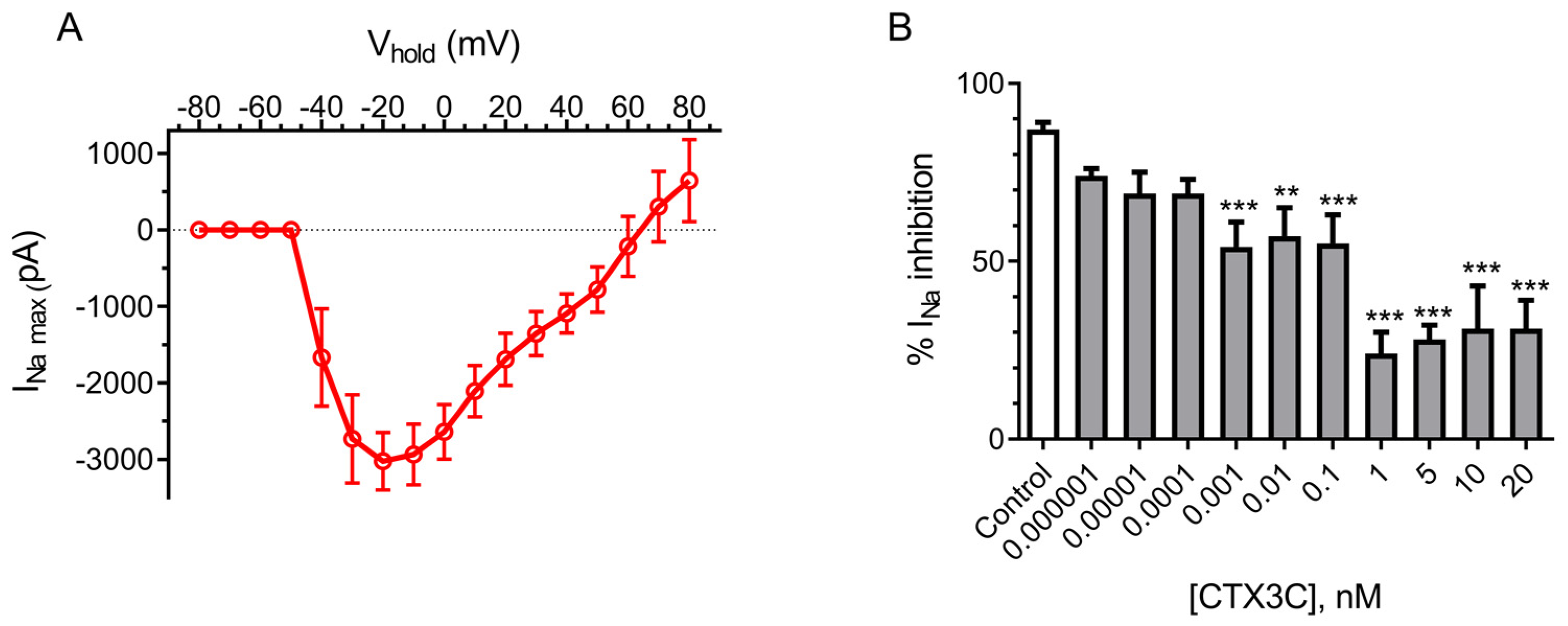
2.3. Effect of CTX3C in the Human SH-SY5Y Neuroblastoma Cell Line
2.3.1. Voltage-Gated Sodium Channel Functionality
2.3.2. Evaluation of CT3C Cytotoxicity by MTT in Human Neuroblastoma Cells
2.4. Evaluation of the Effect of CTX3C in HEK293 Cells Expressing the Human Nav1.6 Sodium Channel Alpha Subunit
MTT in HEK Cells Expressing the Human Nav1.6 Sodium Channel Subunit
3. Discussion
4. Materials and Methods
4.1. Toxins and Drugs
4.2. Human and Mouse Cell Lines
4.2.1. N2a Mouse Neuroblastoma Cell Line
4.2.2. Human Neuroblastoma Cell Line SH-SY5Y
4.2.3. Human Embryonic Kidney Cell Line (HEK293) Cell Culture
4.3. Determination of Cell Viability
4.3.1. MTT Assay
4.3.2. Alamar Blue Assay
4.3.3. Lactate Dehydrogenase Assay
4.4. Immunocytochemistry
4.5. Electrophysiology
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nicholson, G.M.; Lewis, R.J. Ciguatoxins: Cyclic polyether modulators of voltage-gated Iion channel function. Mar. Drugs 2006, 4, 82–118. [Google Scholar] [CrossRef]
- Martin, V.; Vale, C.; Hirama, M.; Yamashita, S.; Rubiolo, J.A.; Vieytes, M.R.; Botana, L.M. Synthetic ciguatoxin CTX 3C induces a rapid imbalance in neuronal excitability. Chem. Res. Toxicol. 2015, 28, 1095–1108. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.; Vale, C.; Rubiolo, J.A.; Roel, M.; Hirama, M.; Yamashita, S.; Vieytes, M.R.; Botana, L.M. Chronic ciguatoxin treatment induces synaptic scaling through voltage gated sodium channels in cortical neurons. Chem. Res. Toxicol. 2015, 28, 1109–1119. [Google Scholar] [CrossRef] [PubMed]
- Raposo-Garcia, S.; Louzao, M.C.; Fuwa, H.; Sasaki, M.; Vale, C.; Botana, L.M. Determination of the toxicity equivalency factors for ciguatoxins using human sodium channels. Food Chem. Toxicol. 2022, 160, 112812. [Google Scholar] [CrossRef]
- Loeffler, C.R.; Spielmeyer, A.; Friedemann, M.; Kapp, K.; Schwank, U.; Kappenstein, O.; Bodi, D. Food Safety Risk in Germany From Mislabeled Imported Fish: Ciguatera Outbreak Trace-Back, Toxin Elucidation, and Public Health Implications. Front. Mar. Sci. 2022, 9, 849857. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on marine biotoxins in shellfish–Emerging toxins: Ciguatoxin group. EFSA J. 2010, 8, 1627. [Google Scholar]
- WHO. Report of the Expert Meeting on Ciguatera Poisoning: Rome, 19–23 November 2018; Food and Agriculture Organization: Rome, Italy, 2020; Volume 9. [Google Scholar]
- FDA. Appendix 5—FDA and EPA safety levels in regulations and guidance. In Fish and Fishery Products Hazards and Control Guidance, 4th ed.; University of Florida: Gainesville, FL, USA, 2020; p. A5-11. [Google Scholar]
- Raposo-García, S.; Castro, D.; Lence, E.; Estévez, P.; Leão, J.M.; González-Bello, C.; Gago-Martínez, A.; Louzao, M.C.; Vale, C.; Botana, L.M. In silico simulations and functional cell studies evidence similar potency and distinct binding of Pacific and Caribbean ciguatoxins. Expo. Health 2022, 15, 641–660. [Google Scholar] [CrossRef]
- Caillaud, A.; Eixarch, H.; de la Iglesia, P.; Rodriguez, M.; Dominguez, L.; Andree, K.B.; Diogene, J. Towards the standardisation of the neuroblastoma (neuro-2a) cell-based assay for ciguatoxin-like toxicity detection in fish: Application to fish caught in the Canary Islands. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2012, 29, 1000–1010. [Google Scholar] [CrossRef]
- Van Meerloo, J.; Kaspers, G.J.; Cloos, J. Cell sensitivity assays: The MTT assay. Cancer Cell Cult. 2011, 731, 237–245. [Google Scholar]
- Viallon, J.; Chinain, M.; Darius, H.T. Revisiting the Neuroblastoma Cell-Based Assay (CBA-N2a) for the Improved Detection of Marine Toxins Active on Voltage Gated Sodium Channels (VGSCs). Toxins 2020, 12, 281. [Google Scholar] [CrossRef]
- Pasinszki, T.; Lako, J.; Dennis, T.E. Advances in Detecting Ciguatoxins in Fish. Toxins 2020, 12, 494. [Google Scholar] [CrossRef]
- Nagae, M.; Igarashi, T.; Mizukoshi, K.; Kuniyoshi, K.; Oshiro, N.; Yasumoto, T. Development and Validation of an LC-MS/MS Method for the Ultra-Trace Analysis of Pacific Ciguatoxins in Fish. J. AOAC Int. 2021, 104, 1272–1281. [Google Scholar] [CrossRef] [PubMed]
- Oguri, H.; Hirama, M.; Tsumuraya, T.; Fujii, I.; Maruyama, M.; Uehara, H.; Nagumo, Y. Synthesis-based approach toward direct sandwich immunoassay for ciguatoxin CTX3C. J. Am. Chem. Soc. 2003, 125, 7608–7612. [Google Scholar] [CrossRef] [PubMed]
- Sibat, M.; Herrenknecht, C.; Darius, H.T.; Roué, M.; Chinain, M.; Hess, P. Detection of pacific ciguatoxins using liquid chromatography coupled to either low or high resolution mass spectrometry (LC-MS/MS). J. Chromatogr. A 2018, 1571, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, C.; Luan, W.; Li, X.; Liu, Y.; Luo, X. Ultrasensitive and accelerated detection of ciguatoxin by capillary electrophoresis via on-line sandwich immunoassay with rotating magnetic field and nanoparticles signal enhancement. Anal. Chim. Acta 2015, 888, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Bottein Dechraoui, M.Y.; Wang, Z.; Turquet, J.; Chinain, M.; Darius, T.; Cruchet, P.; Radwan, F.F.; Dickey, R.W.; Ramsdell, J.S. Biomonitoring of ciguatoxin exposure in mice using blood collection cards. Toxicon 2005, 46, 243–251. [Google Scholar] [CrossRef]
- Vernoux, J. The mouse ciguatoxin bioassay: Directions for use to control fish for consumption. Mem. Qld. Museum. Brisb. 1994, 34, 625–629. [Google Scholar]
- Vernoux, J.-P.; Lewis, R.J. Isolation and characterisation of Caribbean ciguatoxins from the horse-eye jack (Caranx latus). Toxicon 1997, 35, 889–900. [Google Scholar] [CrossRef]
- Raposo-Garcia, S.; Boente-Juncal, A.; Rodriguez-Vieytes, M.; Camina, M.; Costas, C.; Cao, A.; Louzao, M.C.; Cifuentes, M.; Vale, C.; Botana, L.M. In vivo subchronic effects of ciguatoxin-related compounds, reevaluation of their toxicity. Arch. Toxicol. 2022, 96, 2621–2638. [Google Scholar] [CrossRef]
- Kogure, K.; Tamplin, M.L.; Simidu, U.; Colwell, R.R. A tissue culture assay for tetrodotoxin, saxitoxin and related toxins. Toxicon 1988, 26, 191–197. [Google Scholar] [CrossRef]
- Coccini, T.; Caloni, F.; De Simone, U. Human neuronal cell based assay: A new in vitro model for toxicity evaluation of ciguatoxin. Environ. Toxicol. Pharmacol. 2017, 52, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, C.R.; Bodi, D.; Tartaglione, L.; Dell’Aversano, C.; Preiss-Weigert, A. Improving in vitro ciguatoxin and brevetoxin detection: Selecting neuroblastoma (Neuro-2a) cells with lower sensitivity to ouabain and veratridine (OV-LS). Harmful Algae 2021, 103, 101994. [Google Scholar] [CrossRef]
- Louzao, M.C.; Vieytes, M.R.; Yasumoto, T.; Botana, L.M. Detection of sodium channel activators by a rapid fluorimetric microplate assay. Chem. Res. Toxicol. 2004, 17, 572–578. [Google Scholar] [CrossRef]
- Tudo, A.; Rambla-Alegre, M.; Flores, C.; Sagrista, N.; Aguayo, P.; Reverte, L.; Campas, M.; Gouveia, N.; Santos, C.; Andree, K.B.; et al. Identification of New CTX Analogues in Fish from the Madeira and Selvagens Archipelagos by Neuro-2a CBA and LC-HRMS. Mar. Drugs 2022, 20, 236. [Google Scholar] [CrossRef] [PubMed]
- Hardison, D.R.; Holland, W.C.; Darius, H.T.; Chinain, M.; Tester, P.A.; Shea, D.; Bogdanoff, A.K.; Morris, J.A., Jr.; Flores Quintana, H.A.; Loeffler, C.R.; et al. Investigation of ciguatoxins in invasive lionfish from the greater caribbean region: Implications for fishery development. PLoS ONE 2018, 13, e0198358. [Google Scholar] [CrossRef] [PubMed]
- Hardison, D.R.; Holland, W.C.; McCall, J.R.; Bourdelais, A.J.; Baden, D.G.; Darius, H.T.; Chinain, M.; Tester, P.A.; Shea, D.; Quintana, H.A.; et al. Fluorescent Receptor Binding Assay for Detecting Ciguatoxins in Fish. PLoS ONE 2016, 11, e0153348. [Google Scholar] [CrossRef] [PubMed]
- Pisapia, F.; Holland, W.C.; Hardison, D.R.; Litaker, R.W.; Fraga, S.; Nishimura, T.; Adachi, M.; Nguyen-Ngoc, L.; Sechet, V.; Amzil, Z.; et al. Toxicity screening of 13 Gambierdiscus strains using neuro-2a and erythrocyte lysis bioassays. Harmful Algae 2017, 63, 173–183. [Google Scholar] [CrossRef]
- Kerbrat, A.S.; Darius, H.T.; Pauillac, S.; Chinain, M.; Laurent, D. Detection of ciguatoxin-like and paralysing toxins in Trichodesmium spp. from New Caledonia lagoon. Mar. Pollut. Bull. 2010, 61, 360–366. [Google Scholar] [CrossRef]
- Cañete, E.; Diogène, J. Comparative study of the use of neuroblastoma cells (Neuro-2a) and neuroblastoma× glioma hybrid cells (NG108-15) for the toxic effect quantification of marine toxins. Toxicon 2008, 52, 541–550. [Google Scholar] [CrossRef]
- Castro, D.; Manger, R.; Vilarino, O.; Gago-Martinez, A. Evaluation of Matrix Issues in the Applicability of the Neuro-2a Cell Based Assay on the Detection of CTX in Fish Samples. Toxins 2020, 12, 308. [Google Scholar] [CrossRef]
- Abdullaeva, O.S. Activation of Voltage-Gated Ion Channels in Neuroblastoma Cells by an Organic Artificial Photoreceptor. Ph.D. Thesis, Universität Oldenburg, Oldenburg, Germany, 2019. [Google Scholar]
- Badisa, R.B.; Wi, S.; Jones, Z.; Mazzio, E.; Zhou, Y.; Rosenberg, J.T.; Latinwo, L.M.; Grant, S.C.; Goodman, C.B. Cellular and molecular responses to acute cocaine treatment in neuronal-like N2a cells: Potential mechanism for its resistance in cell death. Cell Death Discov. 2018, 4, 13. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Saify, Z.S.; Jamali, K.S.; Naz, S.; Hassan, S.; Siddiqui, S. Vitex negundo induces an anticonvulsant effect by inhibiting voltage gated sodium channels in murine Neuro 2A cell line. Pak. J. Pharm. Sci. 2018, 31, 297–303. [Google Scholar] [PubMed]
- Ammer, H.; Schulz, R. Regulation of stimulatory adenylyl cyclase signaling during forskolin-induced differentiation of mouse neuroblastoma x rat glioma (NG108-15) cells. Neurosci. Lett. 1997, 230, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Bonvallet, R.; Lemtiri-Chlieh, F.; Rougier, O. Forskolin but not isoprenaline increases sodium current from frog cardiac myocytes. Exp. Physiol. 1990, 75, 843–846. [Google Scholar] [CrossRef]
- Deng, W.; Obrocka, M.; Fischer, I.; Prockop, D.J. In vitro differentiation of human marrow stromal cells into early progenitors of neural cells by conditions that increase intracellular cyclic AMP. Biochem. Biophys. Res. Commun. 2001, 282, 148–152. [Google Scholar] [CrossRef]
- England, S.; Bevan, S.; Docherty, R.J. PGE2 modulates the tetrodotoxin-resistant sodium current in neonatal rat dorsal root ganglion neurones via the cyclic AMP-protein kinase A cascade. J. Physiol. 1996, 495 Pt 2, 429–440. [Google Scholar] [CrossRef]
- Fitzgerald, E.M.; Okuse, K.; Wood, J.N.; Dolphin, A.C.; Moss, S.J. cAMP-dependent phosphorylation of the tetrodotoxin-resistant voltage-dependent sodium channel SNS. J. Physiol. 1999, 516 Pt 2, 433–446. [Google Scholar] [CrossRef]
- Thompson, R.; Casali, C.; Chan, C. Forskolin and IBMX Induce Neural Transdifferentiation of MSCs Through Downregulation of the NRSF. Sci. Rep. 2019, 9, 2969. [Google Scholar] [CrossRef]
- Yusta, B.; Ortiz-Caro, J.; Pascual, A.; Aranda, A. Comparison of the effects of forskolin and dibutyryl cyclic AMP in neuroblastoma cells: Evidence that some of the actions of dibutyryl cyclic AMP are mediated by butyrate. J. Neurochem. 1988, 51, 1808–1818. [Google Scholar] [CrossRef]
- Holman, G.D.; Kasuga, M. From receptor to transporter: Insulin signalling to glucose transport. Diabetologia 1997, 40, 991–1003. [Google Scholar] [CrossRef]
- Levin, G.; Keren, T.; Peretz, T.; Chikvashvili, D.; Thornhill, W.B.; Lotan, I. Regulation of RCK1 currents with a cAMP analog via enhanced protein synthesis and direct channel phosphorylation. J. Biol. Chem. 1995, 270, 14611–14618. [Google Scholar] [CrossRef] [PubMed]
- Tartakoff, A.M. Perturbation of the structure and function of the Golgi complex by monovalent carboxylic ionophores. Methods Enzymol. 1983, 98, 47–59. [Google Scholar] [PubMed]
- Zhou, J.; Yi, J.; Hu, N.; George, A.L., Jr.; Murray, K.T. Activation of protein kinase A modulates trafficking of the human cardiac sodium channel in Xenopus oocytes. Circ. Res. 2000, 87, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Silva, E.; Soares-da-Silva, P. New insights into the regulation of Na+, K+-ATPase by ouabain. Int. Rev. Cell Mol. Biol. 2012, 294, 99–132. [Google Scholar] [PubMed]
- Castro, D.; Estévez, P.; Leao-Martins, J.M.; Dickey, R.W.; García-Álvarez, N.; Real, F.; Costa, P.R.; Gago-Martínez, A. Preparation of ciguatoxin reference materials from Canary Islands (Spain) and Madeira Archipelago (Portugal) fish. J. Mar. Sci. Eng. 2022, 10, 835. [Google Scholar] [CrossRef]
- Yokozeki, T.; Hama, Y.; Fujita, K.; Igarashi, T.; Hirama, M.; Tsumuraya, T. Evaluation of relative potency of calibrated ciguatoxin congeners by near-infrared fluorescent receptor binding and neuroblastoma cell-based assays. Toxicon 2023, 230, 107161. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.-W.; Li, X.; Lam, P.K.S.; Cheng, S.H.; Schlenk, D.; de Mitcheson, Y.S.; Li, Y.; Gu, J.-D.; Chan, L.L. Proteomic analysis of hepatic tissue of ciguatoxin (CTX) contaminated coral reef fish Cephalopholis argus and moray eel Gymnothorax undulatus. Harmful Algae 2012, 13, 65–71. [Google Scholar] [CrossRef]
- Sauviat, M.P.; Boydron-Le Garrec, R.; Masson, J.B.; Lewis, R.L.; Vernoux, J.P.; Molgo, J.; Laurent, D.; Benoit, E. Mechanisms involved in the swelling of erythrocytes caused by Pacific and Caribbean ciguatoxins. Blood Cells Mol. Dis. 2006, 36, 1–9. [Google Scholar] [CrossRef]
- Contreras, R.G.; Shoshani, L.; Flores-Maldonado, C.; Lazaro, A.; Cereijido, M. Relationship between Na(+),K(+)-ATPase and cell attachment. J. Cell Sci. 1999, 112 Pt 23, 4223–4232. [Google Scholar] [CrossRef]
- Lopina, O.D.; Tverskoi, A.M.; Klimanova, E.A.; Sidorenko, S.V.; Orlov, S.N. Ouabain-induced cell death and survival. Role of α1-Na, K-ATPase-mediated signaling and [Na+]i/[K+]i-dependent gene expression. Front. Physiol. 2020, 11, 1060. [Google Scholar] [CrossRef]
- Qiu, J.; Gao, H.-Q.; Zhou, R.-H.; Liang, Y.; Zhang, X.-H.; Wang, X.-P.; You, B.-A.; Cheng, M. Proteomics analysis of the proliferative effect of low-dose ouabain on human endothelial cells. Biol. Pharm. Bull. 2007, 30, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.H.; Gao, H.Q.; Ji, X.; Wang, Y.; Liu, X.J.; You, B.A.; Cui, X.P.; Qiu, J. Effect of ouabain on myocardial ultrastructure and cytoskeleton during the development of ventricular hypertrophy. Heart Vessel. 2013, 28, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Toselli, M.; Tosetti, P.; Taglietti, V. Functional changes in sodium conductances in the human neuroblastoma cell line SH-SY5Y during in vitro differentiation. J. Neurophysiol. 1996, 76, 3920–3927. [Google Scholar] [CrossRef]
- Inserra, M.C.; Israel, M.R.; Caldwell, A.; Castro, J.; Deuis, J.R.; Harrington, A.M.; Keramidas, A.; Garcia-Caraballo, S.; Maddern, J.; Erickson, A.; et al. Multiple sodium channel isoforms mediate the pathological effects of Pacific ciguatoxin-1. Sci. Rep. 2017, 7, 42810. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, K.; Inoue, M.; Miyazaki, K.; Hirama, M.; Kondo, C.; Kinoshita, E.; Miyoshi, H.; Seyama, I. Synthetic ciguatoxins selectively activate Nav1.8-derived chimeric sodium channels expressed in HEK293 cells. J. Biol. Chem. 2009, 284, 7597–7605. [Google Scholar] [CrossRef] [PubMed]
- Botana, L.M. Toxicological Perspective on Climate Change: Aquatic Toxins. Chem. Res. Toxicol. 2016, 29, 619–625. [Google Scholar] [CrossRef]
- LePage, K.T.; Dickey, R.W.; Gerwick, W.H.; Jester, E.L.; Murray, T.F. On the use of neuro-2a neuroblastoma cells versus intact neurons in primary culture for neurotoxicity studies. Crit. Rev. Neurobiol. 2005, 17, 27–50. [Google Scholar] [CrossRef]
- Dechraoui, M.Y.; Tiedeken, J.A.; Persad, R.; Wang, Z.; Granade, H.R.; Dickey, R.W.; Ramsdell, J.S. Use of two detection methods to discriminate ciguatoxins from brevetoxins: Application to great barracuda from Florida Keys. Toxicon 2005, 46, 261–270. [Google Scholar] [CrossRef]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Wekell, M.M. Cell Bioassay of Neurotoxins. US5858687A, 12 January 1999. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Diaz-Asencio, L.; Clausing, R.J.; Ranada, M.L.; Alonso-Hernandez, C.M.; Dechraoui Bottein, M.Y. A radioligand receptor binding assay for ciguatoxin monitoring in environmental samples: Method development and determination of quality control criteria. J. Environ. Radioact. 2018, 192, 289–294. [Google Scholar] [CrossRef]
- Burbidge, S.A.; Dale, T.J.; Powell, A.J.; Whitaker, W.R.; Xie, X.M.; Romanos, M.A.; Clare, J.J. Molecular cloning, distribution and functional analysis of the NA(V)1.6. Voltage-gated sodium channel from human brain. Brain Res. Mol. Brain Res. 2002, 103, 80–90. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| METHOD | LOQ (M) |
|---|---|
| MTT in N2a sensitized with O/V | 1 × 10−12 |
| LDH in N2a sensitized with O/V | No cytotoxic effect |
| Alamar blue in N2a sensitized with O/V | No cytotoxic effect |
| MTT in N2a sensitized with O/V and pretreated with 10 µM forskolin | 1 × 10−11 |
| LDH in N2a sensitized with O/V and pretreated with 10 µM forskolin | 1 × 10−12 |
| Alamar blue in N2a sensitized with O/V and pretreated with 10 µM forskolin | 1 × 10−12 |
| MTT in HEK293 Nav | 1 × 10−10 |
| MTT in HEK293 Nav sensitized with O/V | 1 × 10−13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raposo-Garcia, S.; Cao, A.; Costas, C.; Louzao, M.C.; Vilariño, N.; Vale, C.; Botana, L.M. Mouse N2a Neuroblastoma Assay: Uncertainties and Comparison with Alternative Cell-Based Assays for Ciguatoxin Detection. Mar. Drugs 2023, 21, 590. https://doi.org/10.3390/md21110590
Raposo-Garcia S, Cao A, Costas C, Louzao MC, Vilariño N, Vale C, Botana LM. Mouse N2a Neuroblastoma Assay: Uncertainties and Comparison with Alternative Cell-Based Assays for Ciguatoxin Detection. Marine Drugs. 2023; 21(11):590. https://doi.org/10.3390/md21110590
Chicago/Turabian StyleRaposo-Garcia, Sandra, Alejandro Cao, Celia Costas, M. Carmen Louzao, Natalia Vilariño, Carmen Vale, and Luis M. Botana. 2023. "Mouse N2a Neuroblastoma Assay: Uncertainties and Comparison with Alternative Cell-Based Assays for Ciguatoxin Detection" Marine Drugs 21, no. 11: 590. https://doi.org/10.3390/md21110590
APA StyleRaposo-Garcia, S., Cao, A., Costas, C., Louzao, M. C., Vilariño, N., Vale, C., & Botana, L. M. (2023). Mouse N2a Neuroblastoma Assay: Uncertainties and Comparison with Alternative Cell-Based Assays for Ciguatoxin Detection. Marine Drugs, 21(11), 590. https://doi.org/10.3390/md21110590