
Chemistry and Bioactivity of the Deep-Water Antarctic Octocoral Alcyonium sp.


 and
and 
Abstract
:1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Biological Material
3.3. Extraction and Isolation of Coral Metabolites
3.4. Leishmania donovani Infected Macrophage Assay
3.5. Clostridium difficile Susceptibility Screening
3.6. Determination of the Half Maximal Inhibitory Concentration (IC50) toward Human Liver Cells and Kidney Cells
3.7. X-ray Diffraction of Alcyosterone (5)
4. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tsao, F.; Morgan, L.E. Corals that live on mountaintops. J. Mar. Ed. 2005, 21, 9–11. [Google Scholar]
- Morgan, L.E. What are deep-sea corals? J. Mar. Ed. 2005, 21, 2–4. [Google Scholar]
- Moore, J.M.; Carvajal, J.I.; Rouse, G.W.; Wilson, N.G. The Antarctic Circumpolar Current isolates and connects: Structured circumpolarity in the sea star Glabraster antarctica. Ecol. Evol. 2018, 8, 10621–10633. [Google Scholar] [CrossRef] [PubMed]
- Dueñas, L.F.; Tracey, D.M.; Crawford, A.J.; Wilke, T.; Alderslade, P.; Sánchez, J.A. The Antarctic Circumpolar Current as a diversification trigger for deep-sea octocorals. BMC Evol. Biol. 2016, 16, 2. [Google Scholar] [CrossRef] [PubMed]
- von Salm, J.L.; Wilson, N.G.; Vesely, B.A.; Kyle, D.E.; Cuce, J.; Baker, B.J. Shagenes A and B, new tricyclic sesquiterpenes produced by an undescribed Antarctic octocoral. Org. Lett. 2014, 16, 2630–2633. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.A.L.; von Salm, J.L.; Clark, S.; Nemani, P.; Ferlita, S.; Wilson, N.G.; Baker, B.J. Keikipukalides, furanocembranoid aldehydes from the deep sea Antarctic coral Plumerella delicatissima. J. Nat. Prod. 2018, 81, 117–123. [Google Scholar] [CrossRef]
- Núñez-Ponz, L.; Shilling, A.J.; Verde, C.; Baker, B.J.; Giordano, D. Marine terpenoids from polar latitudes and their potential applications in biotechnology. Mar. Drugs 2020, 18, 401. [Google Scholar] [CrossRef]
- Soldatou, S.; Baker, B.J. Cold-water marine natural products, 2006 to 2016. Nat. Prod. Rep. 2017, 34, 585–626. [Google Scholar] [CrossRef]
- von Salm, J.L.; Schoenrock, K.M.; McClintock, J.B.; Amsler, C.D.; Baker, B.J. The status of marine chemical ecology in Antarctica: Form and function of unique high-latitude chemistry. In Chemical Ecology: The Ecological Impacts of Marine Natural Products; Puglisi, M.P., Becerr, M.A., Eds.; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Rocha, J.; Peixe, L.; Gomes, N.C.M.; Calado, R. Cnidarians as a source of new marine bioactive compounds—An overview of the last decade and future steps for bioprospecting. Mar. Drugs 2011, 9, 1860–1886. [Google Scholar] [CrossRef]
- Leal, M.C.; Puga, J.; Serodio, J.; Gomes, N.C.; Calado, R. Trends in the discovery of new marine natural products from invertebrates over the last two decades--where and what are we bioprospecting? PLoS ONE 2012, 7, e30580. [Google Scholar] [CrossRef]
- Paul, V.J.; Puglisi, M.P. Chemical mediation of interactions among marine organisms. Nat. Prod. Rep. 2004, 21, 189–209. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez Brasco, M.F.; Seldes, A.M.; Palermo, J.A. Paesslerins A and B: Novel tricyclic sesquiterpenoids from the soft coral Alcyonium paessleri. Org. Lett. 2001, 3, 1415–1417. [Google Scholar] [CrossRef] [PubMed]
- Iken, K.B.; Baker, B.J. Ainigmaptilones, sesquiterpenes from the Antarctic gorgonian coral Ainigmaptilon antarcticus. J. Nat. Prod. 2003, 66, 888–890. [Google Scholar] [CrossRef] [PubMed]
- McMorris, T.C.; Anchel, M. The structures of the basidiomycete metabolites Illudin S and Illudin M. J. Am. Chem. Soc. 1963, 85, 831–832. [Google Scholar] [CrossRef]
- Palermo, J.A.; Rodriguez Brasco, M.F.; Spagnuolo, C.; Seldes, A.M. Illudalane sesquiterpenoids from the soft coral Alcyonium paessleri: The first natural nitrate esters. J. Org. Chem. 2000, 65, 4482–4486. [Google Scholar] [CrossRef]
- Carbone, M.; Núñez-Pons, L.; Castelluccio, F.; Avila, C.; Gavagnin, M. Illudalane sesquiterpenoids of the alcyopterosin series from the Antarctic marine soft coral Alcyonium grandis. J. Nat. Prod. 2009, 72, 1357–1360. [Google Scholar] [CrossRef]
- Orfali, R.; Perveen, S.; Khan, M.F.; Ahmed, A.F.; Wadaan, M.A.; Al-Taweel, A.M.; Alqahtani, A.S.; Nasr, F.A.; Tabassum, S.; Luciano, P.; et al. Antiproliferative illudalane sesquiterpenes from the marine sediment ascomycete Aspergillus oryzae. Mar. Drugs 2021, 19, 333. [Google Scholar] [CrossRef]
- Finkielsztein, L.M.; Bruno, A.M.; Renou, S.G.; Moltrasio Iglesias, G.Y. Design, synthesis, and biological evaluation of alcyopterosin A and illudalane derivatives as anticancer agents. Bioorg. Med. Chem. 2006, 14, 1863–1870. [Google Scholar] [CrossRef]
- Leffler, D.A.; Lamont, J.T. Clostridium difficile infection. N. Eng. J. Med. 2015, 373, 287–288. [Google Scholar] [CrossRef]
- Baker, B.J.; Wilson, N.G.; Kyle, D.E.; Limon, A.-C. Leishmania inhibitors. U.S. Patent 10898460-B1, 26 January 2021. [Google Scholar]
- Salari, S.; Bamorovat, M.; Sharifi, I.; Almani, P.G.N. Global distribution of treatment resistance gene markers for leishmaniasis. J. Clin. Lab. Anal. 2022, 36, e24599. [Google Scholar] [CrossRef]
- France, S.C.; Hoover, L.L. Analysis of variation in mitochondrial DNA sequences (ND3, ND4L, MSH) among Octocorallia (= Alcyonaria)(Cnidaria: Anthozoa). Bull. Biol. Soc. Wash. 2001, 10, 110–118. [Google Scholar]
- France, S.C.; Hoover, L.L. DNA sequences of the mitochondrial COI gene have low levels of divergence among deep-sea octocorals (Cnidaria: Anthozoa). Hydrobiologia 2002, 471, 149–155. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Shilling, A.J.; Witowski, C.G.; Maschek, J.A.; Azhari, A.; Vesely, B.; Kyle, D.E.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Spongian diterpenoids derived from the Antarctic sponge Dendrilla antarctica are potent inhibitors of the Leishmania parasite. J. Nat. Prod. 2020, 83, 1553–1562. [Google Scholar] [CrossRef]
- Kokkaliari, S.; Pham, K.; Shahbazi, N.; Calcul, L.; Wojtas, L.; Wilson, N.G.; Crawford, A.D.; Baker, B.J. Australindolones, new aminopyrimidine substituted indolone alkaloids from an Antarctic tunicate Synoicum sp. Mar. Drugs 2022, 20, 196. [Google Scholar] [CrossRef]
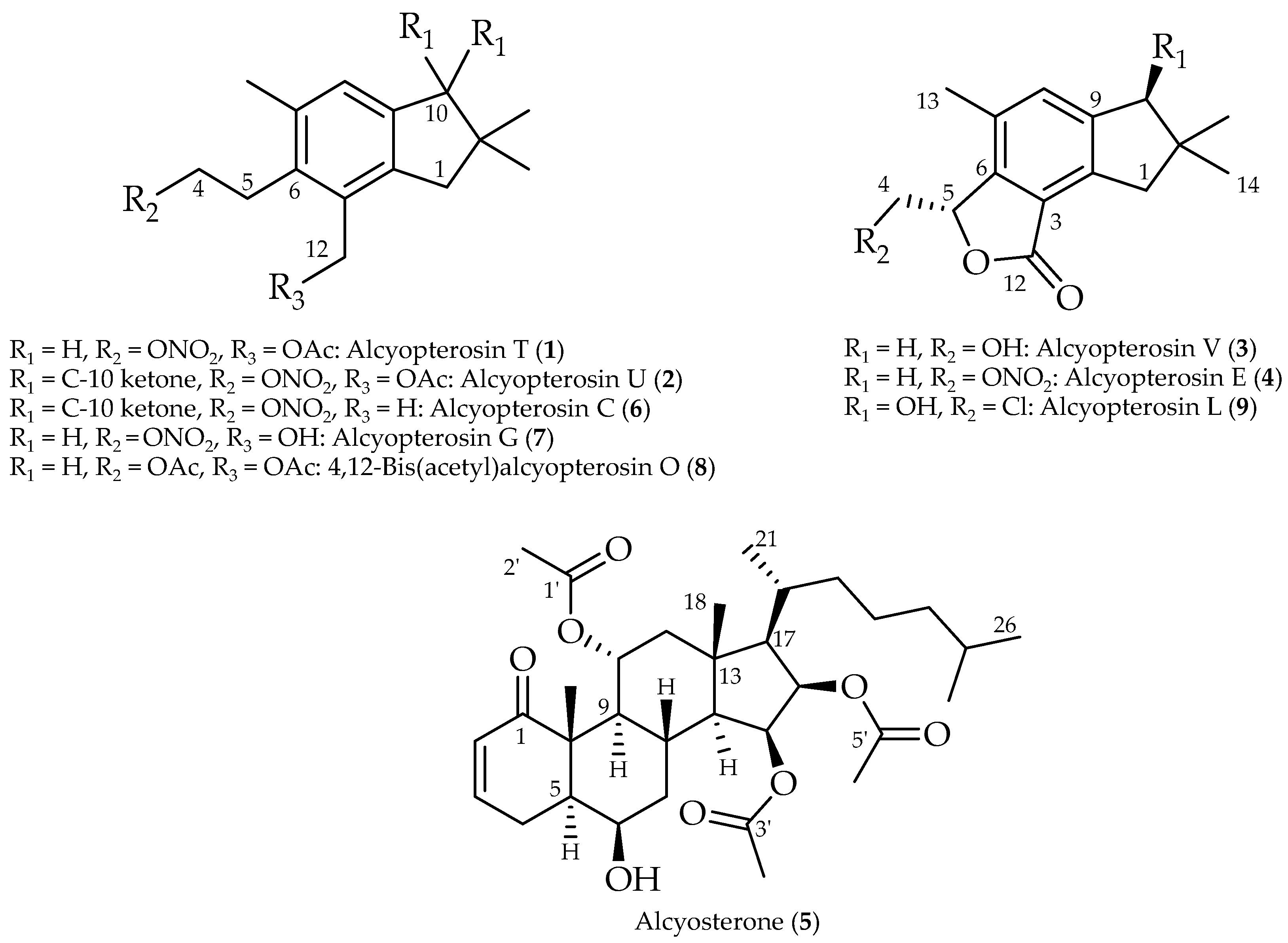





{kind=link}
{kind=link}
{kind=link}
{kind=link}
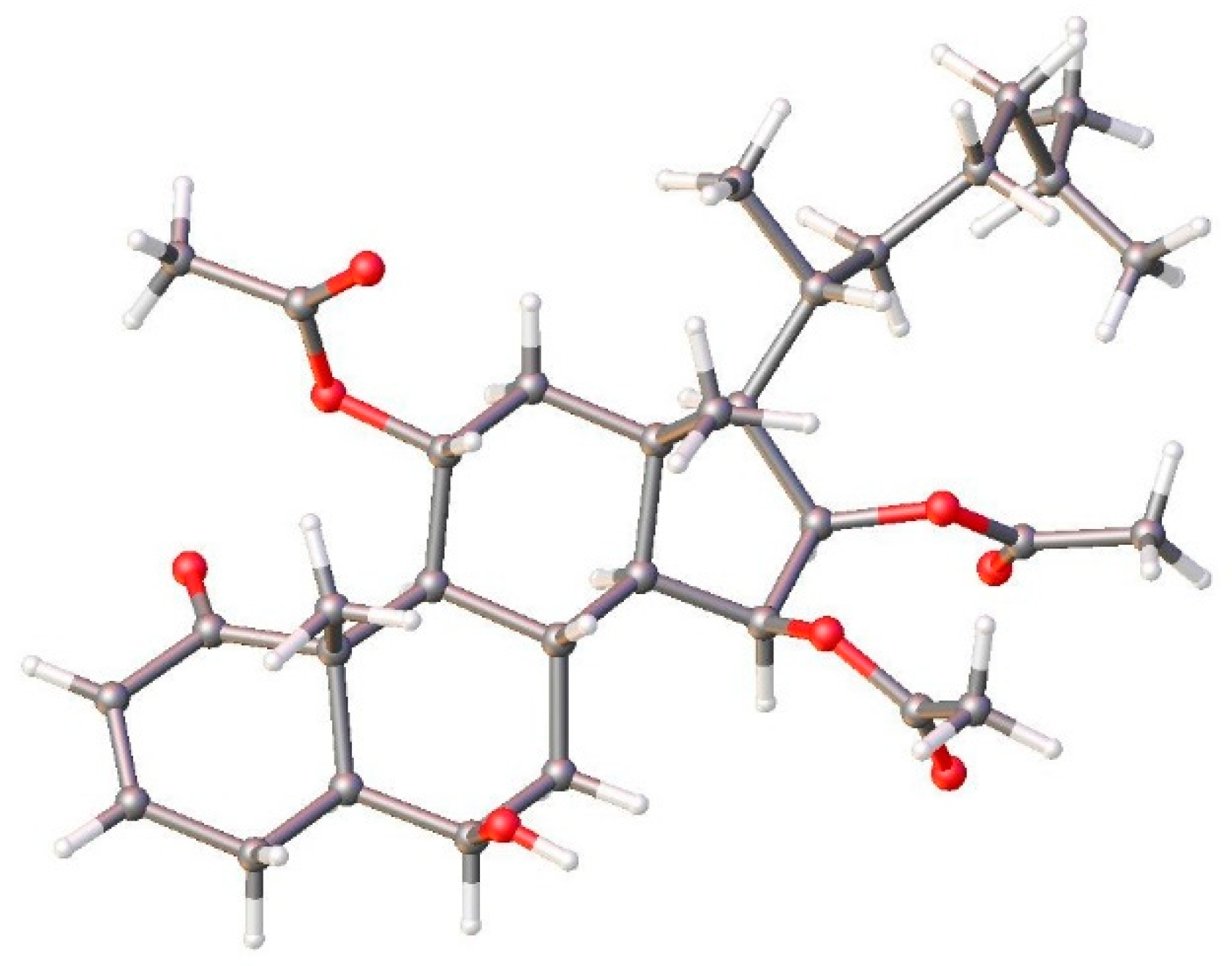{kind=link}
| Position | Alcyopterosin T (1) | Alcyopterosin U (2) | Alcyopterosin V (3) | |||
|---|---|---|---|---|---|---|
| δC, 1 Type | δH, 2 Mult. (J) | δC, 1 Type | δH, 2 Mult. (J) | δC, 1 Type | δH, 2 Mult. (J) | |
| 1 | 47.1, CH2 | 2.79, s | 42.2, CH2 | 3.03, s | 44.8, CH2 | 3.04, s |
| 2 | 143.2, C | 151.5, C | 146.7, C | |||
| 3 | 131.1, C | 133.0, C | 122.5, C | |||
| 4 | 72.5, CH2 | 4.57, t (7.9) | 71.9, CH2 | 4.60, t (7.6) | 63.2, CH2 | 4.25, dd (2.5, 12.6) |
| 3.81, dd (6.1, 12.6) | ||||||
| 5 | 27.7, CH2 | 3.15, t (7.9) | 28.6, CH2 | 3.28, t (7.7) | 82.2, CH | 5.55, br dd (2.0, 5.9) |
| 6 | 131.4, C | 142.2, C | 142.4, C | |||
| 7 | 135.9, C | 138.2, C | 130.0, C | |||
| 8 | 128.0, CH | 7.06, s | 127.4, CH | 7.64, s | 131.8, CH | 7.24, s |
| 9 | 143.4, C | 135.4, C | 141.1, C | |||
| 10 | 48.5, CH2 | 2.73, s | 211.4, C | 47.0, CH2 | 2.74, s | |
| 11 | 40.4, C | 46.3, C | 40.9, C | |||
| 12 | 62.5, CH2 | 5.16, s | 61.0, CH2 | 5.25, s | 170.8, C | |
| 13 | 20.7, CH3 | 2.38, s | 21.0, CH3 | 2.47, s | 18.0, CH3 | 2.37, s |
| 14 | 29.7, CH3 | 1.17, s | 26.2, CH3 | 1.24, s | 28.8, CH3 | 1.16, s |
| 15 | 29.7, CH3 | 1.17, s | 26.2, CH3 | 1.24, s | 28.8, CH3 | 1.19, s |
| 1′ | 171.1, C | 171.2, C | ||||
| 2′ | 21.7, CH3 | 2.09, s | 21.8, CH3 | 2.10, s | ||
| Position | δC1, Type | δH, 2 Integ., Mult., J |
|---|---|---|
| 1 | 203.9, C | |
| 2 | 128.4, CH | 5.83, 1H, dd, 2.2, 9.9 |
| 3 | 142.5, CH | 6.58, 1H, ddd, 2.1, 4.8, 9.6 |
| 4 | 28.4, CH2 | 2.79, 1H, dddd, 0.7, 2.4, 11.4, 19.8 |
| 2.11, 1H, ddd, 0.8, 4.8, 19.5 | ||
| 5 | 46.6, CH | 1.86, 1H, ddd, 0.7, 2.9, 10.8 |
| 6 | 69.7, CH | 3.87, 1H, q, 2.4 |
| 7 | 36.8, CH2 | 1.74, 1H, ov 3 |
| 1.21, 1H, ov | ||
| 8 | 24.9, CH | 2.23, 1H, ov |
| 9 | 47.8, CH | 2.07, 1H, ov |
| 10 | 47.7, C | |
| 11 | 70.4 *, C | 5.02, 1H, dt, 3.9, 11 |
| 12 | 46.7, CH2 | 2.20, 1H, ov |
| 1.48, 1H, ov | ||
| 13 | 43.7, CH | |
| 14 | 56.6, CH | 1.31, 1H, dd, 5.8, 11.2 |
| 15 | 70.5 *, CH | 5.34, 1H, dd, 6.3, 6.6 |
| 16 | 73.0, CH | 5.51, 1H, dd, 6.9, 7.0 |
| 17 | 59.9, CH | 1.34, 1H, ov |
| 18 | 15.8, CH3 | 1.22, 3H, s |
| 19 | 13.2, CH3 | 1.28, 3H, s |
| 20 | 30.0, CH | 1.76, 1H, ov |
| 21 | 18.2, CH3 | 0.95, 3H, d, 6.6 |
| 22 | 35.6, CH2 | 1.20, 1H, ov |
| 0.90, 1H, ov | ||
| 23 | 24.4, CH2 | 1.36, 1H, ov |
| 1.11, 1H, ov | ||
| 24 | 39.1, CH2 | 1.09, 1H, ov |
| 1.05, 1H, d, 6.6 | ||
| 25 | 27.9, CH | 1.48, 1H, ov |
| 26 | 22.6, CH3 | 0.85, 3H, d, 6.5 |
| 27 | 22.4, CH3 | 0.85, 3H, d, 6.5 |
| 1′ | 170.4, C | |
| 2′ | 21.5, CH3 | 1.93, 3H, s |
| 3′ | 169.9, C | |
| 4′ | 20.7, CH3 | 2.06, 3H, s |
| 5′ | 169.4, C | |
| 6′ | 20.5, CH3 | 2.02, 3H, s |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Limon, A.-C.D.; Patabendige, H.M.L.W.; Azhari, A.; Sun, X.; Kyle, D.E.; Wilson, N.G.; Baker, B.J. Chemistry and Bioactivity of the Deep-Water Antarctic Octocoral Alcyonium sp. Mar. Drugs 2022, 20, 576. https://doi.org/10.3390/md20090576
Limon A-CD, Patabendige HMLW, Azhari A, Sun X, Kyle DE, Wilson NG, Baker BJ. Chemistry and Bioactivity of the Deep-Water Antarctic Octocoral Alcyonium sp. Marine Drugs. 2022; 20(9):576. https://doi.org/10.3390/md20090576
Chicago/Turabian StyleLimon, Anne-Claire D., Hiran M. L. W. Patabendige, Ala Azhari, Xingmin Sun, Dennis E. Kyle, Nerida G. Wilson, and Bill J. Baker. 2022. "Chemistry and Bioactivity of the Deep-Water Antarctic Octocoral Alcyonium sp." Marine Drugs 20, no. 9: 576. https://doi.org/10.3390/md20090576
APA StyleLimon, A.-C. D., Patabendige, H. M. L. W., Azhari, A., Sun, X., Kyle, D. E., Wilson, N. G., & Baker, B. J. (2022). Chemistry and Bioactivity of the Deep-Water Antarctic Octocoral Alcyonium sp. Marine Drugs, 20(9), 576. https://doi.org/10.3390/md20090576