
Emerging Trends in Genetic Engineering of Microalgae for Commercial Applications


Abstract
1. Introduction
1.1. Phylogenetical and Biochemical Diversity of Microalgae
1.2. Various Applications of Microalgae
1.3. Recent Development of Microalgal Biotechnology
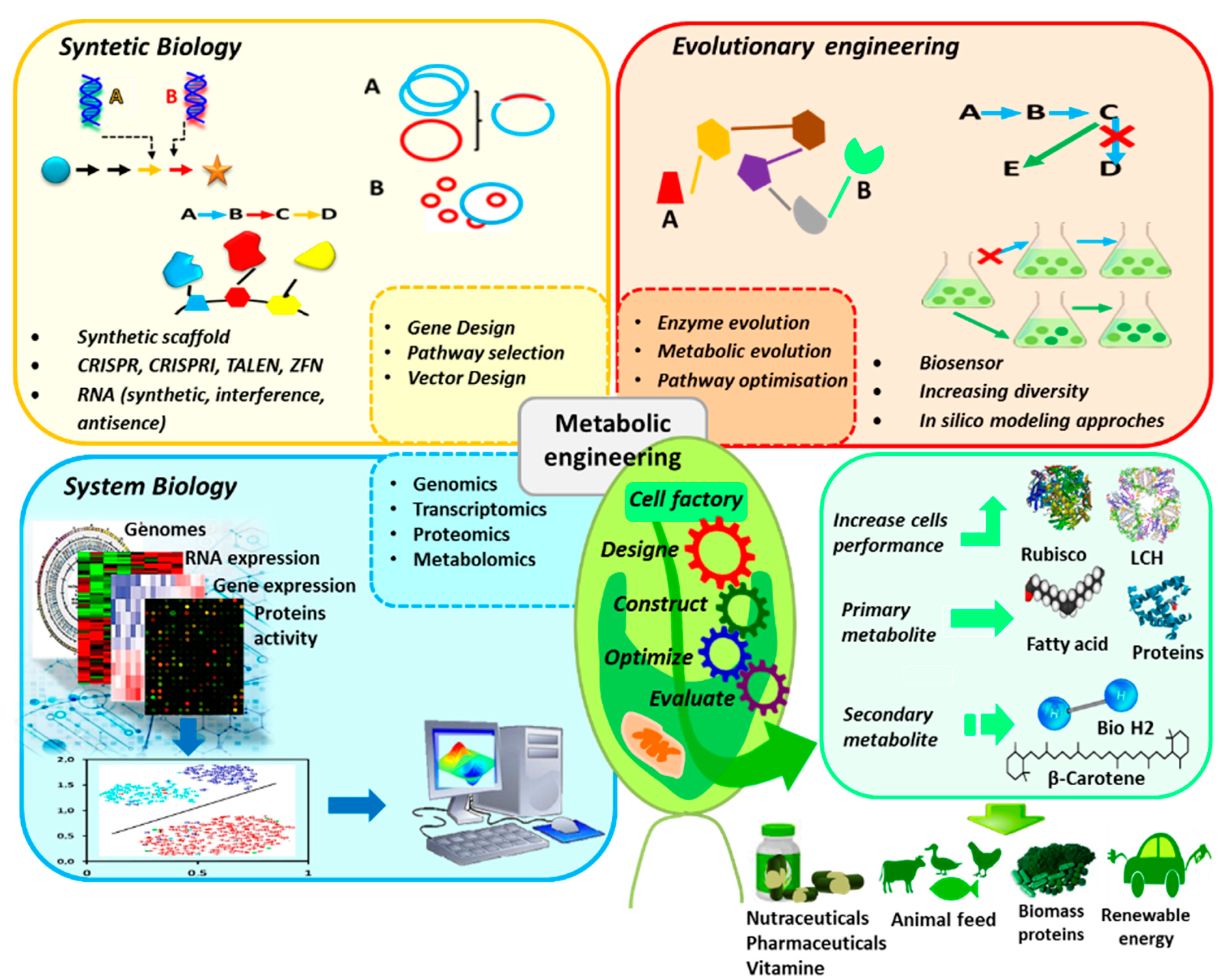
2. Emerging Trends in Genetic Engineering of Microalgae for Commercial Applications
2.1. Genetic Engineering of Microalgae for Pharmaceutical Protein Production

{kind=link}
{kind=link}
| Microalgae Strain | Gene/Target Site | Approach | Results | References |
|---|---|---|---|---|
| Chlamydomonas reinhardtii | Endolysins Cpl-1 and Pal | Foreign gene expression | Total recombinant protein yield was ~1.3 mg/g algal dry weight | Stoffels et al. [23] |
| Chlamydomonas reinhardtii | Birch pollen allergen Bet v 1 | Codon-optimized gene and stably integrated | Allergen expression with yields between 0.01 and 0.04% of TSP | Hirschl et al. [27] |
| Thalassiosira pseudonana | Antigen IbpA DR2 | Nuclear-based expression | Increased recombinant protein by 1.2% | Davis et al. [21] |
| Chlamydomonas reinhardtii | HIV antigen P24 | Codon-optimized | The yield of the recombinant protein increased up to 0.25% of the total cellular protein | Barahimipour et al. [28] |
| Chlamydomonas reinhardtii and Chlorella vulgaris | SARS-CoV-2 receptor binding domain (RBD) and basic fibroblast growth factor (bFGF) | Nuclear transformation | Up to 1.14 mg/g RBD and 1.61 ng/g FGF in C. vulgaris and 1.61 mg/g RBD and 1.025 ng/g FGF in C. reinhardtii | Malla et al. [33] |
| Schizochytrium sp. | Epitopes from tumor associated antigens | Cloning and ex-pression | BCB protein was expressed at levels up to 637 μg/g fresh weight | Hernández-Ramírez et al. [34] |
| Haematococcus pluvialis | antimicrobial peptide piscidin-4 | Expression of codon-optimized | Confirmed that the antimicrobial peptide could be expressed from H. pluvialis | Wang et al. [35] |
| Tetraselmis subcordiformis | rt-PA | nuclear transformation | rt-PA was integrated, and the expression product was bioactive | Wu et al. [36] |
| Fistulifera solaris | cox gene | Cloning and ex-pression | The total content of Prostaglandins (PGs) was 1290.4 ng/g of dry cell weight | Maeda et al. [37] |
| Schizochytrium sp. | LTB:RAGE vaccine | Algevir system (inducible geminiviral vector) | Led to yields of up to 380 μg LTB:RAGE/g fresh weight | Ortega-Berlanga et al. [38] |
| Schizochytrium sp. | vaccine against Zika virus (ZIKV) | Algevir technology to express an antigenic protein | Antigen yields of up to 365 μg g−1 microalgae fresh weight | Márquez-Escobar et al. [39] |
| Chlamydomonas reinhardtii | Human interferon (IFN) | Cloning and ex-pression | IFN-α2a is expressed and it is functionally active as anticancer and antiviral agent | El-Ayouty et al. [40] |
| Chlamydomonas reinhardtii | PfCelTOS Antigen | Chloroplast expressed | Expressed recombinant PfCelTOS accumulates as a soluble, properly folded and functional protein | Shamriz et al. [41] |
| Chlamydomonas reinhardtii | Human growth hormone (hGH) | Codon-optimized and new vectors | 0.5 mg hGH per liter of culture | Wannathong et al. [22] |
2.2. Genetic Engineering of Microalgae for Lipid Production
| Microalgae Strain | Gene/Target Site | Approach | Results | References |
|---|---|---|---|---|
| Synechocystis sp. PCC 6803 | Acyl-ACP synthetase (aas) | Overexpression of aas | Increased lipid production by 5.4% | Eungrasamee et al. [60] |
| Chlamydomonas reinhartdii | Phospholipase A2 (PLA2) | Knock-out/CRISPR/Cas9 | Improves the lipids’ production up to 64.25% | Shin et al. [62] |
| Chlamydomonas reinhardtii PTS42 | Malic enzyme isoform 2 (ME2) | Overexpression | Increasing lipid rate up to 23.4% | Kim et al. [46] |
| Chlamydomonas reinhardtii CC400 | PEPC1 | Down regulation by CRISPRi/Cas9 | Lipid (content and productivity of 28.5% DCW and 34.9 mg/L/day) | Kao and Ng. [63] |
| Chlamydomonas reinhardtii | HpDGAT2D | Heterologous expression | Increasing TAG content by ~1.4-fold | Cui et al. [64] |
| Chlamydomonas reinhardtii CC-4349 | ZEP and AGP genes | CRISPR-Cas9 RNP-mediated knock-out method | Increased oil productivity by 81% | Song et al. [65] |
| Nannochloropsis oceanica | AtDXS gene | Engineering a control-knob gene | Lipid production increased by ~68.6% in nitrogen depletion and ~110.6% in high light | Han et al. [66] |
| Phaeodactylum tricornutum. | GPAT and DGAT2 genes | Overexpression | Total lipid content increased by 2.6-fold and reached up to 57.5% DCW | Zou et al. [67] |
| Nannochloropsis salina | bZIP | Overexpressed a bZIP TF, NsbZIP1 | Lipid production increased by 50% | Kwon et al. [68] |
| Scenedesmus obliqnus | Differential expression genes (DEGs) | up-regulated genes | Lipid yield increased by 2.4 fold | Xi et al. [69] |
| Phaeodactylum tricornutum | ptTES1 | Transcription activator-like effector nucleases (TALENs) | 1.7-fold increase in TAG content | Hao et al. [70] |
| Nannochloropsis oceanica | Transposome | Insertion of a Transposome complex (mutagenesis) | Increased PUFA by 180% and EPA by 40% | Osorio et al. [71] |
| Phaeodactylum tricornutum | PhyA | Overexpression | Increased DHA by 12% and EPA by 18% | Pudney et al. [72] |
| Synechocystis sp. | Acetyl-CoA carboxylase (ACC) | Overexpression | Increased its lipid content by 3.6-fold | Fathy et al. [73] |
| Chlamydomonas reinhardtii | Diacylglycerol acyltransferase 2 (DGAT) | Heterologous expression | α-linolenic acid, an important omega-3 fatty acid, was improved by more than 12% | Ahmad et al. [51] |
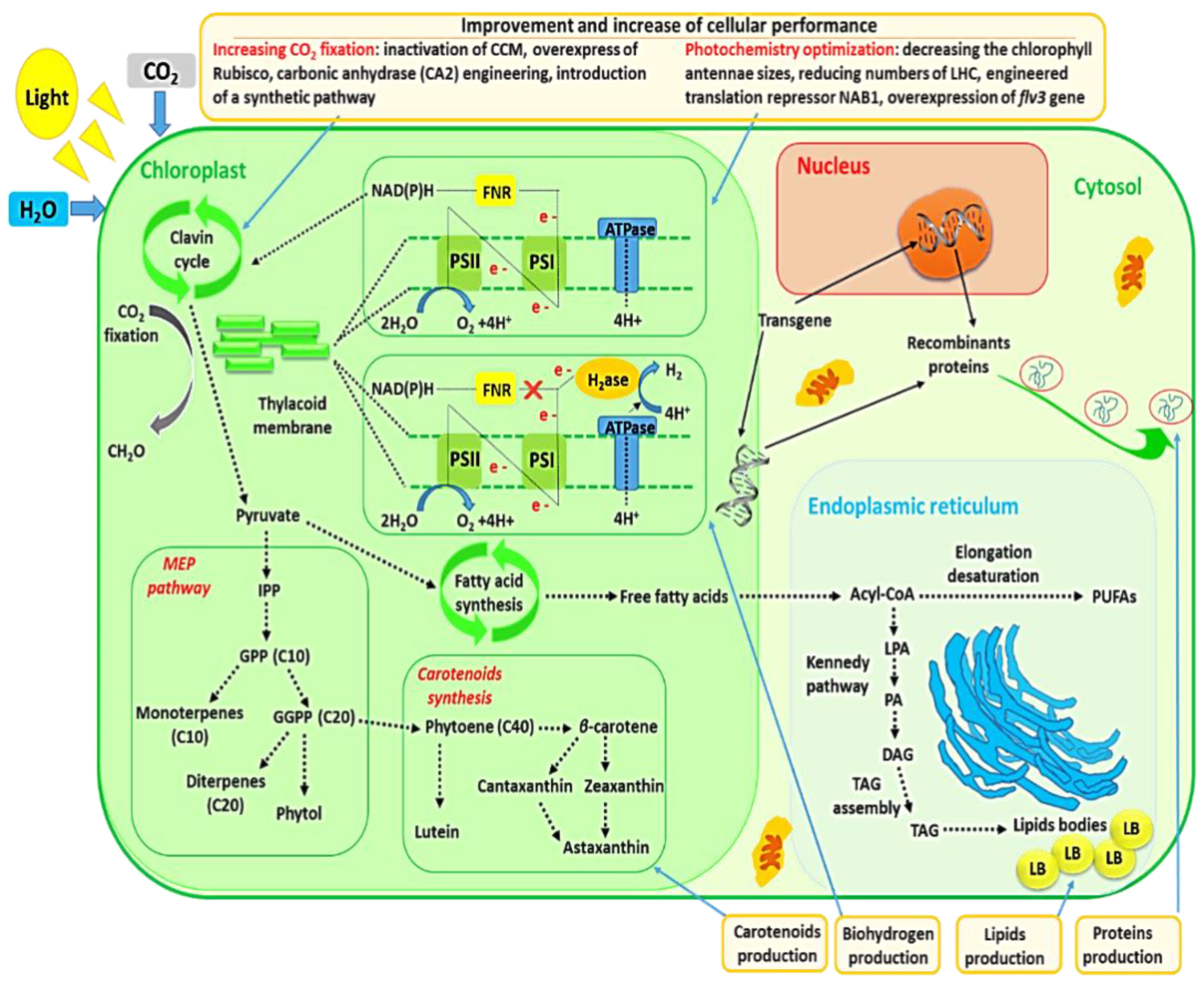
2.3. Genetic Engineering of Microalgae for Carotenoid Production
2.4. Genetic Engineering of Microalgae for Biohydrogen Production
2.5. Genetic Engineering of Microalgae for CO2 Sequestration
2.6. Genetic Engineering of Microalgae for Photochemistry Optimization
3. Opportunities, Challenges and Prospects
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef]
- Metting, F. Biodiversity and application of microalgae. J. Ind. Microbiol. 1996, 17, 477–489. [Google Scholar] [CrossRef]
- Cadoret, J.-P.; Garnier, M.; Saint-Jean, B. Microalgae, functional genomics and biotechnology. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2012; Volume 64, pp. 285–341. [Google Scholar]
- Mobin, S.M.A.; Chowdhury, H.; Alam, F. Commercially important bioproducts from microalgae and their current applications–A review. Energy Procedia 2019, 160, 752–760. [Google Scholar] [CrossRef]
- Camacho, F.; Macedo, A.; Malcata, F. Potential industrial applications and commercialization of microalgae in the functional food and feed industries: A Short Review. Mar. Drugs 2019, 17, 312. [Google Scholar] [CrossRef]
- Li, X.; Wang, X.; Duan, C.; Yi, S.; Gao, Z.; Xiao, C.; Agathos, S.N.; Wang, G.; Li, J. Biotechnological production of astaxanthin from the microalga Haematococcus pluvialis. Biotechnol. Adv. 2020, 43, 107602. [Google Scholar] [CrossRef]
- Guiry, M.D. How many species of microalgae are there? J. Phycol. 2012, 48, 1057–1063. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef]
- Olaizola, M. Commercial development of microalgal biotechnology: From the test tube to the marketplace. Biomol. Eng. 2003, 20, 459–466. [Google Scholar] [CrossRef]
- Harun, R.; Singh, M.; Forde, G.M.; Danquah, M.K. Bioprocess engineering of microalgae to produce a variety of consumer products. Renew. Sustain. Energy Rev. 2010, 14, 1037–1047. [Google Scholar] [CrossRef]
- Skjånes, K.; Rebours, C.; Lindblad, P. Potential for green microalgae to produce hydrogen, pharmaceuticals and other high value products in a combined process. Crit. Rev. Biotechnol. 2013, 33, 172–215. [Google Scholar] [CrossRef]
- Barsanti, L.; Gualtieri, P. Algae: Anatomy, Biochemistry, and Biotechnology; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Person, J.; Lando, D.; Mathieu, D. Livre Turquoise: Algues, Filières du Futur; Édition Adebiotech: Romainville, France, 2011. [Google Scholar]
- Borowitzka, M.A. High-value products from microalgae—their development and commercialisation. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- Choi, K.R.; Jang, W.D.; Yang, D.; Cho, J.S.; Park, D.; Lee, S.Y. Systems metabolic engineering strategies: Integrating systems and synthetic biology with metabolic engineering. Trends Biotechnol. 2019, 37, 817–837. [Google Scholar] [CrossRef] [PubMed]
- Sproles, A.E.; Fields, F.J.; Smalley, T.N.; Le, C.H.; Badary, A.; Mayfield, S.P. Recent advancements in the genetic engineering of microalgae. Algal Res. 2021, 53, 102158. [Google Scholar] [CrossRef]
- Fajardo, C.; De Donato, M.; Carrasco, R.; Martínez-Rodríguez, G.; Mancera, J.M.; Fernández-Acero, F.J. Advances and challenges in genetic engineering of microalgae. Rev. Aquac. 2020, 12, 365–381. [Google Scholar] [CrossRef]
- Rasala, B.A.; Mayfield, S.P. Photosynthetic biomanufacturing in green algae; production of recombinant proteins for industrial, nutritional, and medical uses. Photosynth. Res. 2015, 123, 227–239. [Google Scholar] [CrossRef]
- Vanier, G.; Hempel, F.; Chan, P.; Rodamer, M.; Vaudry, D.; Maier, U.G.; Lerouge, P.; Bardor, M. Biochemical characterization of human anti-hepatitis B monoclonal antibody produced in the microalgae Phaeodactylum tricornutum. PLoS ONE 2015, 10, e0139282. [Google Scholar] [CrossRef]
- Ramos-Martinez, E.M.; Fimognari, L.; Sakuragi, Y. High-yield secretion of recombinant proteins from the microalga Chlamydomonas reinhardtii. Plant Biotechnol. J. 2017, 15, 1214–1224. [Google Scholar] [CrossRef]
- Davis, A.; Crum, L.T.; Corbeil, L.B.; Hildebrand, M. Expression of Histophilus somni IbpA DR2 protective antigen in the diatom Thalassiosira pseudonana. Appl. Microbiol. Biotechnol. 2017, 101, 5313–5324. [Google Scholar] [CrossRef]
- Wannathong, T.; Waterhouse, J.C.; Young, R.E.; Economou, C.K.; Purton, S. New tools for chloroplast genetic engineering allow the synthesis of human growth hormone in the green alga Chlamydomonas reinhardtii. Appl. Microbiol. Biotechnol. 2016, 100, 5467–5477. [Google Scholar] [CrossRef]
- Stoffels, L.; Taunt, H.N.; Charalambous, B.; Purton, S. Synthesis of bacteriophage lytic proteins against Streptococcus pneumoniae in the chloroplast of Chlamydomonas reinhardtii. Plant Biotechnol. J. 2017, 15, 1130–1140. [Google Scholar] [CrossRef]
- Corchero, J.L.; Gasser, B.; Resina, D.; Smith, W.; Parrilli, E.; Vázquez, F.; Abasolo, I.; Giuliani, M.; Jäntti, J.; Ferrer, P. Unconventional microbial systems for the cost-efficient production of high-quality protein therapeutics. Biotechnol. Adv. 2013, 31, 140–153. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Fan, C.; Chen, Y.; Hu, Z. The potential for microalgae as bioreactors to produce pharmaceuticals. Int. J. Mol. Sci. 2016, 17, 962. [Google Scholar] [CrossRef] [PubMed]
- Hempel, F.; Maier, U.G. Microalgae as Solar-Powered Protein Factories. In Advanced Technologies for Protein Complex Production and Characterization; Vega, M.C., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 241–262. [Google Scholar]
- Hirschl, S.; Ralser, C.; Asam, C.; Gangitano, A.; Huber, S.; Ebner, C.; Bohle, B.; Wolf, M.; Briza, P.; Ferreira, F. Expression and characterization of functional recombinant Bet v 1.0101 in the chloroplast of Chlamydomonas reinhardtii. Int. Arch. Allergy Immunol. 2017, 173, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Barahimipour, R.; Neupert, J.; Bock, R. Efficient expression of nuclear transgenes in the green alga Chlamydomonas: Synthesis of an HIV antigen and development of a new selectable marker. Plant Mol. Biol. 2016, 90, 403–418. [Google Scholar] [CrossRef] [PubMed]
- Scranton, M.A.; Ostrand, J.T.; Georgianna, D.R.; Lofgren, S.M.; Li, D.; Ellis, R.C.; Carruthers, D.N.; Dräger, A.; Masica, D.L.; Mayfield, S.P. Synthetic promoters capable of driving robust nuclear gene expression in the green alga Chlamydomonas reinhardtii. Algal Res. 2016, 15, 135–142. [Google Scholar] [CrossRef]
- Zedler, J.A.; Mullineaux, C.W.; Robinson, C. Efficient targeting of recombinant proteins to the thylakoid lumen in Chlamydomonas reinhardtii using a bacterial Tat signal peptide. Algal Res. 2016, 19, 57–62. [Google Scholar] [CrossRef]
- Zedler, J.A.; Gangl, D.; Guerra, T.; Santos, E.; Verdelho, V.V.; Robinson, C. Pilot-scale cultivation of wall-deficient transgenic Chlamydomonas reinhardtii strains expressing recombinant proteins in the chloroplast. Appl. Microbiol. Biotechnol. 2016, 100, 7061–7070. [Google Scholar] [CrossRef]
- Mishra, A.; Medhi, K.; Malaviya, P.; Thakur, I.S. Omics approaches for microalgal applications: Prospects and challenges. Bioresour. Technol. 2019, 291, 121890. [Google Scholar] [CrossRef]
- Malla, A.; Rosales-Mendoza, S.; Phoolcharoen, W.; Vimolmangkang, S. Efficient Transient Expression of Recombinant Proteins Using DNA Viral Vectors in Freshwater Microalgal Species. Front. Plant Sci. 2021, 12, 513. [Google Scholar] [CrossRef]
- Hernández-Ramírez, J.; Wong-Arce, A.; González-Ortega, O.; Rosales-Mendoza, S. Expression in algae of a chimeric protein carrying several epitopes from tumor associated antigens. Int. J. Biol. Macromol. 2020, 147, 46–52. [Google Scholar] [CrossRef]
- Wang, K.; Cui, Y.; Wang, Y.; Gao, Z.; Liu, T.; Meng, C.; Qin, S. Chloroplast Genetic Engineering of a Unicellular Green Alga Haematococcus pluvialis with Expression of an Antimicrobial Peptide. Mar. Biotechnol. 2020, 22, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zheng, C.; Wang, J.; Jiang, P. Recombinant Expression of Thrombolytic Agent Reteplase in Marine Microalga Tetraselmis subcordiformis (Chlorodendrales, Chlorophyta). Mar. Drugs 2021, 19, 315. [Google Scholar] [CrossRef]
- Maeda, Y.; Tsuru, Y.; Matsumoto, N.; Nonoyama, T.; Yoshino, T.; Matsumoto, M.; Tanaka, T. Prostaglandin production by the microalga with heterologous expression of cyclooxygenase. Biotechnol. Bioeng. 2021, 118, 2734–2743. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Berlanga, B.; Bañuelos-Hernández, B.; Rosales-Mendoza, S. Efficient Expression of an Alzheimer’s Disease Vaccine Candidate in the Microalga Schizochytrium sp. Using the Algevir System. Mol. Biotechnol. 2018, 60, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Márquez-Escobar, V.A.; Bañuelos-Hernández, B.; Rosales-Mendoza, S. Expression of a Zika virus antigen in microalgae: Towards mucosal vaccine development. J. Biotechnol. 2018, 282, 86–91. [Google Scholar] [CrossRef] [PubMed]
- El-Ayouty, Y.; El-Manawy, I.; Nasih, S.; Hamdy, E.; Kebeish, R. Engineering Chlamydomonas reinhardtii for Expression of Functionally Active Human Interferon-α. Mol. Biotechnol. 2019, 61, 134–144. [Google Scholar] [CrossRef]
- Shamriz, S.; Ofoghi, H. Expression of Recombinant PfCelTOS Antigen in the Chloroplast of Chlamydomonas reinhardtii and its Potential Use in Detection of Malaria. Mol. Biotechnol. 2019, 61, 102–110. [Google Scholar] [CrossRef]
- Park, S.; Nguyen, T.H.T.; Jin, E. Improving lipid production by strain development in microalgae: Strategies, challenges and perspectives. Bioresour. Technol. 2019, 292, 121953. [Google Scholar] [CrossRef]
- Yan, J.; Cheng, R.; Lin, X.; You, S.; Li, K.; Rong, H.; Ma, Y. Overexpression of acetyl-CoA synthetase increased the biomass and fatty acid proportion in microalga Schizochytrium. Appl. Microbiol. Biotechnol. 2013, 97, 1933–1939. [Google Scholar] [CrossRef]
- Ma, Y.-H.; Wang, X.; Niu, Y.-F.; Yang, Z.-K.; Zhang, M.-H.; Wang, Z.-M.; Yang, W.-D.; Liu, J.-S.; Li, H.-Y. Antisense knockdown of pyruvate dehydrogenase kinase promotes the neutral lipid accumulation in the diatom Phaeodactylum tricornutum. Microb. Cell Factories 2014, 13, 100. [Google Scholar] [CrossRef]
- Xue, J.; Niu, Y.-F.; Huang, T.; Yang, W.-D.; Liu, J.-S.; Li, H.-Y. Genetic improvement of the microalga Phaeodactylum tricornutum for boosting neutral lipid accumulation. Metab. Eng. 2015, 27, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kwak, H.S.; Sim, S.J.; Jin, E. Overexpression of malic enzyme isoform 2 in Chlamydomonas reinhardtii PTS42 increases lipid production. Bioresour. Technol. Rep. 2019, 7, 100239. [Google Scholar] [CrossRef]
- Radakovits, R.; Jinkerson, R.E.; Darzins, A.; Posewitz, M.C. Genetic engineering of algae for enhanced biofuel production. Eukaryot. Cell 2010, 9, 486–501. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.-F.; Zhang, M.-H.; Li, D.-W.; Yang, W.-D.; Liu, J.-S.; Bai, W.-B.; Li, H.-Y. Improvement of neutral lipid and polyunsaturated fatty acid biosynthesis by overexpressing a type 2 diacylglycerol acyltransferase in marine diatom Phaeodactylum tricornutum. Mar. Drugs 2013, 11, 4558–4569. [Google Scholar] [CrossRef]
- Hamilton, M.L.; Haslam, R.P.; Napier, J.A.; Sayanova, O. Metabolic engineering of Phaeodactylum tricornutum for the enhanced accumulation of omega-3 long chain polyunsaturated fatty acids. Metab. Eng. 2014, 22, 3–9. [Google Scholar] [CrossRef]
- Ahmad, I.; Sharma, A.K.; Daniell, H.; Kumar, S. Altered lipid composition and enhanced lipid production in green microalga by introduction of brassica diacylglycerol acyltransferase 2. Plant Biotechnol. J. 2015, 13, 540–550. [Google Scholar] [CrossRef]
- Klaitong, P.; Fa-Aroonsawat, S.; Chungjatupornchai, W. Accelerated triacylglycerol production and altered fatty acid composition in oleaginous microalga Neochloris oleoabundans by overexpression of diacylglycerol acyltransferase 2. Microb. Cell Factories 2017, 16, 61. [Google Scholar] [CrossRef]
- Rismani-Yazdi, H.; Haznedaroglu, B.Z.; Hsin, C.; Peccia, J. Transcriptomic analysis of the oleaginous microalga Neochloris oleoabundans reveals metabolic insights into triacylglyceride accumulation. Biotechnol. Biofuels 2012, 5, 74. [Google Scholar] [CrossRef]
- Tan, K.W.M.; Lee, Y.K. The dilemma for lipid productivity in green microalgae: Importance of substrate provision in improving oil yield without sacrificing growth. Biotechnol. Biofuels 2016, 9, 255. [Google Scholar] [CrossRef]
- Guo, X.; Fan, C.; Chen, Y.; Wang, J.; Yin, W.; Wang, R.R.; Hu, Z. Identification and characterization of an efficient acyl-CoA: Diacylglycerol acyltransferase 1 (DGAT1) gene from the microalga Chlorella ellipsoidea. BMC Plant Biol. 2017, 17, 48. [Google Scholar] [CrossRef]
- Rismani-Yazdi, H.; Haznedaroglu, B.Z.; Bibby, K.; Peccia, J. Transcriptome sequencing and annotation of the microalgae Dunaliella tertiolecta: Pathway description and gene discovery for production of next-generation biofuels. BMC Genom. 2011, 12, 148. [Google Scholar] [CrossRef] [PubMed]
- Courchesne, N.M.D.; Parisien, A.; Wang, B.; Lan, C.Q. Enhancement of lipid production using biochemical, genetic and transcription factor engineering approaches. J. Biotechnol. 2009, 141, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Bajhaiya, A.K.; Moreira, J.Z.; Pittman, J.K. Transcriptional engineering of microalgae: Prospects for high-value chemicals. Trends Biotechnol. 2017, 35, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hao, Q.; Bai, L.; Xu, J.; Yin, W.; Song, L.; Xu, L.; Guo, X.; Fan, C.; Chen, Y. Overexpression of the soybean transcription factor GmDof4 significantly enhances the lipid content of Chlorella ellipsoidea. Biotechnol. Biofuels 2014, 7, 128. [Google Scholar] [CrossRef] [PubMed]
- Ngan, C.Y.; Wong, C.-H.; Choi, C.; Yoshinaga, Y.; Louie, K.; Jia, J.; Chen, C.; Bowen, B.; Cheng, H.; Leonelli, L. Lineage-specific chromatin signatures reveal a regulator of lipid metabolism in microalgae. Nat. Plants 2015, 1, 15107. [Google Scholar] [CrossRef] [PubMed]
- Eungrasamee, K.; Miao, R.; Incharoensakdi, A.; Lindblad, P.; Jantaro, S. Improved lipid production via fatty acid biosynthesis and free fatty acid recycling in engineered Synechocystis sp. PCC 6803. Biotechnol. Biofuels 2019, 12, 8. [Google Scholar] [CrossRef]
- Athanasakoglou, A.; Kampranis, S.C. Diatom isoprenoids: Advances and biotechnological potential. Biotechnol. Adv. 2019, 37, 107417. [Google Scholar] [CrossRef]
- Shin, Y.S.; Jeong, J.; Nguyen, T.H.T.; Kim, J.Y.H.; Jin, E.; Sim, S.J. Targeted knockout of phospholipase A2 to increase lipid productivity in Chlamydomonas reinhardtii for biodiesel production. Bioresour. Technol. 2019, 271, 368–374. [Google Scholar] [CrossRef]
- Kao, P.-H.; Ng, I.-S. CRISPRi mediated phosphoenolpyruvate carboxylase regulation to enhance the production of lipid in Chlamydomonas reinhardtii. Bioresour. Technol. 2017, 245, 1527–1537. [Google Scholar] [CrossRef]
- Cui, H.; Zhao, C.; Xu, W.; Zhang, H.; Hang, W.; Zhu, X.; Ji, C.; Xue, J.; Zhang, C.; Li, R. Characterization of type-2 diacylglycerol acyltransferases in Haematococcus lacustris reveals their functions and engineering potential in triacylglycerol biosynthesis. BMC Plant Biol. 2021, 21, 20. [Google Scholar] [CrossRef]
- Song, I.; Kim, S.; Kim, J.; Oh, H.; Jang, J.; Jeong, S.J.; Baek, K.; Shin, W.-S.; Sim, S.J.; Jin, E. Macular pigment-enriched oil production from genome-edited microalgae. Microb. Cell Factories 2022, 21, 27. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Song, X.; Li, F.; Lu, Y. Improving lipid productivity by engineering a control-knob gene in the oleaginous microalga Nannochloropsis oceanica. Metab. Eng. Commun. 2020, 11, e00142. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.-G.; Chen, J.-W.; Zheng, D.-L.; Balamurugan, S.; Li, D.-W.; Yang, W.-D.; Liu, J.-S.; Li, H.-Y. High-efficiency promoter-driven coordinated regulation of multiple metabolic nodes elevates lipid accumulation in the model microalga Phaeodactylum tricornutum. Microb. Cell Factories 2018, 17, 54. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.; Kang, N.K.; Koh, H.G.; Shin, S.-E.; Lee, B.; Jeong, B.-r.; Chang, Y.K. Enhancement of biomass and lipid productivity by overexpression of a bZIP transcription factor in Nannochloropsis salina. Biotechnol. Bioeng. 2018, 115, 331–340. [Google Scholar] [CrossRef]
- Xi, Y.; Yin, L.; Chi, Z.Y.; Luo, G. Characterization and RNA-seq transcriptomic analysis of a Scenedesmus obliqnus mutant with enhanced photosynthesis efficiency and lipid productivity. Sci. Rep. 2021, 11, 11795. [Google Scholar] [CrossRef]
- Hao, X.; Luo, L.; Jouhet, J.; Rébeillé, F.; Maréchal, E.; Hu, H.; Pan, Y.; Tan, X.; Chen, Z.; You, L.; et al. Enhanced triacylglycerol production in the diatom Phaeodactylum tricornutum by inactivation of a Hotdog-fold thioesterase gene using TALEN-based targeted mutagenesis. Biotechnol. Biofuels 2018, 11, 312. [Google Scholar] [CrossRef]
- Osorio, H.; Jara, C.; Fuenzalida, K.; Rey-Jurado, E.; Vásquez, M. High-efficiency nuclear transformation of the microalgae Nannochloropsis oceanica using Tn5 Transposome for the generation of altered lipid accumulation phenotypes. Biotechnol. Biofuels 2019, 12, 134. [Google Scholar] [CrossRef]
- Pudney, A.; Gandini, C.; Economou, C.K.; Smith, R.; Goddard, P.; Napier, J.A.; Spicer, A.; Sayanova, O. Multifunctionalizing the marine diatom Phaeodactylum tricornutum for sustainable co-production of omega-3 long chain polyunsaturated fatty acids and recombinant phytase. Sci. Rep. 2019, 9, 11444. [Google Scholar] [CrossRef]
- Fathy, W.; Essawy, E.; Tawfik, E.; Khedr, M.; Abdelhameed, M.S.; Hammouda, O.; Elsayed, K. Recombinant overexpression of the Escherichia coli acetyl-CoA carboxylase gene in Synechocystis sp. boosts lipid production. J. Basic Microbiol. 2021, 61, 330–338. [Google Scholar] [CrossRef]
- Grama, B.S.; Chader, S.; Khelifi, D.; Agathos, S.N.; Jeffryes, C. Induction of canthaxanthin production in a Dactylococcus microalga isolated from the Algerian Sahara. Bioresour. Technol. 2014, 151, 297–305. [Google Scholar] [CrossRef]
- Grama, B.S.; Delhaye, A.; Agathos, S.N.; Jeffryes, C. β-Carotene and Other Carotenoids and Pigments from Microalgae. In Industrial Biotechnology of Vitamins, Biopigments, and Antioxidants; Wiley: Hoboken, NJ, USA, 2016. [Google Scholar]
- Ma, R.; Zhao, X.; Xie, Y.; Ho, S.-H.; Chen, J. Enhancing lutein productivity of Chlamydomonas sp. via high-intensity light exposure with corresponding carotenogenic genes expression profiles. Bioresour. Technol. 2019, 275, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sun, Z.; Gerken, H.; Huang, J.; Jiang, Y.; Chen, F. Genetic engineering of the green alga Chlorella zofingiensis: A modified norflurazon-resistant phytoene desaturase gene as a dominant selectable marker. Appl. Microbiol. Biotechnol. 2014, 98, 5069–5079. [Google Scholar] [CrossRef] [PubMed]
- Anila, N.; Simon, D.P.; Chandrashekar, A.; Ravishankar, G.; Sarada, R. Metabolic engineering of Dunaliella salina for production of ketocarotenoids. Photosynth. Res. 2016, 127, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Kathiresan, S.; Chandrashekar, A.; Ravishankar, G.; Sarada, R. Regulation of astaxanthin and its intermediates through cloning and genetic transformation of β-carotene ketolase in Haematococcus pluvialis. J. Biotechnol. 2015, 196, 33–41. [Google Scholar] [CrossRef]
- Huang, W.; Ye, J.; Zhang, J.; Lin, Y.; He, M.; Huang, J. Transcriptome analysis of Chlorella zofingiensis to identify genes and their expressions involved in astaxanthin and triacylglycerol biosynthesis. Algal Res. 2016, 17, 236–243. [Google Scholar] [CrossRef]
- Baek, K.; Yu, J.; Jeong, J.; Sim, S.J.; Bae, S.; Jin, E. Photoautotrophic production of macular pigment in a Chlamydomonas reinhardtii strain generated by using DNA-free CRISPR-Cas9 RNP-mediated mutagenesis. Biotechnol. Bioeng. 2018, 115, 719–728. [Google Scholar] [CrossRef]
- Kim, M.; Ahn, J.; Jeon, H.; Jin, E. Development of a Dunaliella tertiolecta strain with increased zeaxanthin content using random mutagenesis. Mar. Drugs 2017, 15, 189. [Google Scholar] [CrossRef]
- Galarza, J.I.; Gimpel, J.A.; Rojas, V.; Arredondo-Vega, B.O.; Henríquez, V. Over-accumulation of astaxanthin in Haematococcus pluvialis through chloroplast genetic engineering. Algal Res. 2018, 31, 291–297. [Google Scholar] [CrossRef]
- Cui, H.; Xu, W.; Zhu, X.; Zhao, C.; Cui, Y.; Ji, C.; Zhang, C.; Xue, J.; Qin, S.; Jia, X.; et al. Characterization of a Haematococcus pluvialis Diacylglycerol Acyltransferase 1 and Its Potential in Unsaturated Fatty Acid-Rich Triacylglycerol Production. Front. Plant Sci. 2021, 12, 771300. [Google Scholar] [CrossRef]
- Chen, Y.; Bi, C.; Zhang, J.; Hou, H.; Gong, Z. Astaxanthin biosynthesis in transgenic Dunaliella salina (Chlorophyceae) enhanced tolerance to high irradiation stress. S. Afr. J. Bot. 2020, 133, 132–138. [Google Scholar] [CrossRef]
- Perozeni, F.; Cazzaniga, S.; Baier, T.; Zanoni, F.; Zoccatelli, G.; Lauersen, K.J.; Wobbe, L.; Ballottari, M. Turning a green alga red: Engineering astaxanthin biosynthesis by intragenic pseudogene revival in Chlamydomonas reinhardtii. Plant Biotechnol. J. 2020, 18, 2053–2067. [Google Scholar] [CrossRef] [PubMed]
- Eilers, U.; Bikoulis, A.; Breitenbach, J.; Büchel, C.; Sandmann, G. Limitations in the biosynthesis of fucoxanthin as targets for genetic engineering in Phaeodactylum tricornutum. J. Appl. Phycol. 2016, 28, 123–129. [Google Scholar] [CrossRef]
- Kadono, T.; Kira, N.; Suzuki, K.; Iwata, O.; Ohama, T.; Okada, S.; Nishimura, T.; Akakabe, M.; Tsuda, M.; Adachi, M. Effect of an Introduced Phytoene Synthase Gene Expression on Carotenoid Biosynthesis in the Marine Diatom Phaeodactylum tricornutum. Mar. Drugs 2015, 13, 5334–5357. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Y.; Jeon, H.; Sim, S.J.; Lee, Y.; Jin, E. Homologous sense and antisense expression of a gene in Dunaliella tertiolecta. Planta 2015, 242, 1051–1058. [Google Scholar] [CrossRef] [PubMed]
- Rathod, J.P.; Vira, C.; Lali, A.M.; Prakash, G. Metabolic Engineering of Chlamydomonas reinhardtii for Enhanced β-Carotene and Lutein Production. Appl. Biochem. Biotechnol. 2020, 190, 1457–1469. [Google Scholar] [CrossRef]
- Morikawa, T.; Uraguchi, Y.; Sanda, S.; Nakagawa, S.; Sawayama, S. Overexpression of DnaJ-Like Chaperone Enhances Carotenoid Synthesis in Chlamydomonas reinhardtii. Appl. Biochem. Biotechnol. 2018, 184, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Esquível, M.G.; Amaro, H.M.; Pinto, T.S.; Fevereiro, P.S.; Malcata, F.X. Efficient H2 production via Chlamydomonas reinhardtii. Trends Biotechnol. 2011, 29, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Chien, L.-F.; Kuo, T.-T.; Liu, B.-H.; Lin, H.-D.; Feng, T.-Y.; Huang, C.-C. Solar-to-bioH2 production enhanced by homologous overexpression of hydrogenase in green alga Chlorella sp. DT. Int. J. Hydrogen Energy 2012, 37, 17738–17748. [Google Scholar] [CrossRef]
- Quintana, N.; Van der Kooy, F.; Van de Rhee, M.D.; Voshol, G.P.; Verpoorte, R. Renewable energy from Cyanobacteria: Energy production optimization by metabolic pathway engineering. Appl. Microbiol. Biotechnol. 2011, 91, 471–490. [Google Scholar] [CrossRef]
- Melis, A. Photosynthetic H2 metabolism in Chlamydomonas reinhardtii (unicellular green algae). Planta 2007, 226, 1075–1086. [Google Scholar] [CrossRef]
- Beer, L.L.; Boyd, E.S.; Peters, J.W.; Posewitz, M.C. Engineering algae for biohydrogen and biofuel production. Curr. Opin. Biotechnol. 2009, 20, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-C.; Newton, A.J.; Melis, A. Role of SulP, a nuclear-encoded chloroplast sulfate permease, in sulfate transport and H2 evolution in Chlamydomonas reinhardtii. Photosynth. Res. 2005, 84, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Chen, M.; Yang, H.; Zhang, J.; Kuang, T.; Huang, F. Enhanced H2 photoproduction by down-regulation of ferredoxin-NADP+ reductase (FNR) in the green alga Chlamydomonas reinhardtii. Int. J. Hydrogen Energy 2013, 38, 16029–16037. [Google Scholar] [CrossRef]
- McNeely, K.; Xu, Y.; Bennette, N.; Bryant, D.A.; Dismukes, G.C. Redirecting reductant flux into hydrogen production via metabolic engineering of fermentative carbon metabolism in a cyanobacterium. Appl. Environ. Microbiol. 2010, 76, 5032–5038. [Google Scholar] [CrossRef]
- Kosourov, S.N.; Ghirardi, M.L.; Seibert, M. A truncated antenna mutant of Chlamydomonas reinhardtii can produce more hydrogen than the parental strain. Int. J. Hydrogen Energy 2011, 36, 2044–2048. [Google Scholar] [CrossRef]
- Lin, H.-D.; Liu, B.-H.; Kuo, T.-T.; Tsai, H.-C.; Feng, T.-Y.; Huang, C.-C.; Chien, L.-F. Knockdown of PsbO leads to induction of HydA and production of photobiological H2 in the green alga Chlorella sp. DT. Bioresour. Technol. 2013, 143, 154–162. [Google Scholar] [CrossRef]
- Wu, S.; Xu, L.; Huang, R.; Wang, Q. Improved biohydrogen production with an expression of codon-optimized hemH and lba genes in the chloroplast of Chlamydomonas reinhardtii. Bioresour. Technol. 2011, 102, 2610–2616. [Google Scholar] [CrossRef]
- Toepel, J.; Illmer-Kephalides, M.; Jaenicke, S.; Straube, J.; May, P.; Goesmann, A.; Kruse, O. New insights into C hlamydomonas reinhardtii hydrogen production processes by combined microarray/RNA-seq transcriptomics. Plant Biotechnol. J. 2013, 11, 717–733. [Google Scholar] [CrossRef]
- Yang, S.; Guarnieri, M.T.; Smolinski, S.; Ghirardi, M.; Pienkos, P.T. De novo transcriptomic analysis of hydrogen production in the green alga Chlamydomonas moewusii through RNA-Seq. Biotechnol. Biofuels 2013, 6, 118. [Google Scholar] [CrossRef]
- Terashima, M.; Specht, M.; Naumann, B.; Hippler, M. Characterizing the anaerobic response of Chlamydomonas reinhardtii by quantitative proteomics. Mol. Cell. Proteom. 2010, 9, 1514–1532. [Google Scholar] [CrossRef]
- Chader, S.; Mahmah, B.; Chetehouna, K.; Amrouche, F.; Abdeladim, K. Biohydrogen production using green microalgae as an approach to operate a small proton exchange membrane fuel cell. Int. J. Hydrogen Energy 2011, 36, 4089–4093. [Google Scholar] [CrossRef]
- Eroglu, E.; Melis, A. Microalgal hydrogen production research. Int. J. Hydrogen Energy 2016, 41, 12772–12798. [Google Scholar] [CrossRef]
- Khanna, N.; Esmieu, C.; Mészáros, L.S.; Lindblad, P.; Berggren, G. In vivo activation of an [FeFe] hydrogenase using synthetic cofactors. Energy Environ. Sci. 2017, 10, 1563–1567. [Google Scholar] [CrossRef]
- Ban, S.; Lin, W.; Luo, Z.; Luo, J. Improving hydrogen production of Chlamydomonas reinhardtii by reducing chlorophyll content via atmospheric and room temperature plasma. Bioresour. Technol. 2019, 275, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Durall, C.; Lindblad, P. Mechanisms of carbon fixation and engineering for increased carbon fixation in cyanobacteria. Algal Res. 2015, 11, 263–270. [Google Scholar] [CrossRef]
- González-Fernández, C.; Ballesteros, M. Linking microalgae and cyanobacteria culture conditions and key-enzymes for carbohydrate accumulation. Biotechnol. Adv. 2012, 30, 1655–1661. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Shen, C.; El Hajjami, M.; You, W.; Wang, Q.; Zhang, P.; Ji, Y.; Hu, H.; Hu, Q.; Poetsch, A. Knockdown of carbonate anhydrase elevates Nannochloropsis productivity at high CO2 level. Metab. Eng. 2019, 54, 96–108. [Google Scholar] [CrossRef]
- Atsumi, S.; Higashide, W.; Liao, J.C. Direct photosynthetic recycling of carbon dioxide to isobutyraldehyde. Nat. Biotechnol. 2009, 27, 1177–1180. [Google Scholar] [CrossRef]
- Shih, P.M.; Zarzycki, J.; Niyogi, K.K.; Kerfeld, C.A. Introduction of a synthetic CO2-fixing photorespiratory bypass into a cyanobacterium. J. Biol. Chem. 2014, 289, 9493–9500. [Google Scholar] [CrossRef]
- Hu, J.; Li, T.; Xu, W.; Zhan, J.; Chen, H.; He, C.; Wang, Q. Small antisense RNA RblR positively regulates RuBisCo in Synechocystis sp. PCC 6803. Front. Microbiol. 2017, 8, 231. [Google Scholar] [CrossRef][Green Version]
- Gimpel, J.A.; Henríquez, V.; Mayfield, S.P. In metabolic engineering of eukaryotic microalgae: Potential and challenges come with great diversity. Front. Microbiol. 2015, 6, 1376. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Nelson, D.R.; Mystikou, A.; Daakour, S.; Salehi-Ashtiani, K. Advances in microalgal research and engineering development. Curr. Opin. Biotechnol. 2019, 59, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Melis, A. Solar energy conversion efficiencies in photosynthesis: Minimizing the chlorophyll antennae to maximize efficiency. Plant Sci. 2009, 177, 272–280. [Google Scholar] [CrossRef]
- Polle, J.E.; Kanakagiri, S.-D.; Melis, A. tla1, a DNA insertional transformant of the green alga Chlamydomonas reinhardtii with a truncated light-harvesting chlorophyll antenna size. Planta 2003, 217, 49–59. [Google Scholar] [CrossRef]
- Mussgnug, J.H.; Thomas-Hall, S.; Rupprecht, J.; Foo, A.; Klassen, V.; McDowall, A.; Schenk, P.M.; Kruse, O.; Hankamer, B. Engineering photosynthetic light capture: Impacts on improved solar energy to biomass conversion. Plant Biotechnol. J. 2007, 5, 802–814. [Google Scholar] [CrossRef]
- Beckmann, J.; Lehr, F.; Finazzi, G.; Hankamer, B.; Posten, C.; Wobbe, L.; Kruse, O. Improvement of light to biomass conversion by de-regulation of light-harvesting protein translation in Chlamydomonas reinhardtii. J. Biotechnol. 2009, 142, 70–77. [Google Scholar] [CrossRef]
- Kirst, H.; Formighieri, C.; Melis, A. Maximizing photosynthetic efficiency and culture productivity in cyanobacteria upon minimizing the phycobilisome light-harvesting antenna size. Biochim. Biophys. Acta (BBA)-Bioenerg. 2014, 1837, 1653–1664. [Google Scholar] [CrossRef]
- Jeong, J.; Baek, K.; Kirst, H.; Melis, A.; Jin, E. Loss of CpSRP54 function leads to a truncated light-harvesting antenna size in Chlamydomonas reinhardtii. Biochim. Biophys. Acta (BBA)-Bioenerg. 2017, 1858, 45–55. [Google Scholar] [CrossRef]
- Mussgnug, J.H.; Wobbe, L.; Elles, I.; Claus, C.; Hamilton, M.; Fink, A.; Kahmann, U.; Kapazoglou, A.; Mullineaux, C.W.; Hippler, M.; et al. NAB1 Is an RNA Binding Protein Involved in the Light-Regulated Differential Expression of the Light-Harvesting Antenna of Chlamydomonas reinhardtii. Plant Cell 2005, 17, 3409–3421. [Google Scholar] [CrossRef]
- Berger, H.; Blifernez-Klassen, O.; Ballottari, M.; Bassi, R.; Wobbe, L.; Kruse, O. Integration of carbon assimilation modes with photosynthetic light capture in the green alga Chlamydomonas reinhardtii. Mol. Plant 2014, 7, 1545–1559. [Google Scholar] [CrossRef]
- Hasunuma, T.; Matsuda, M.; Senga, Y.; Aikawa, S.; Toyoshima, M.; Shimakawa, G.; Miyake, C.; Kondo, A. Overexpression of flv3 improves photosynthesis in the cyanobacterium Synechocystis sp. PCC6803 by enhancement of alternative electron flow. Biotechnol. Biofuels 2014, 7, 493. [Google Scholar] [CrossRef] [PubMed]
- Rea, G.; Lambreva, M.; Polticelli, F.; Bertalan, I.; Antonacci, A.; Pastorelli, S.; Damasso, M.; Johanningmeier, U.; Giardi, M.T. Directed evolution and in silico analysis of reaction centre proteins reveal molecular signatures of photosynthesis adaptation to radiation pressure. PLoS ONE 2011, 6, e16216. [Google Scholar] [CrossRef] [PubMed]
- Morozova, O.; Marra, M.A. Applications of next-generation sequencing technologies in functional genomics. Genomics 2008, 92, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Mardis, E.R. The impact of next-generation sequencing technology on genetics. Trends Genet. 2008, 24, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Horner, D.S.; Pavesi, G.; Castrignano, T.; De Meo, P.D.O.; Liuni, S.; Sammeth, M.; Picardi, E.; Pesole, G. Bioinformatics approaches for genomics and post genomics applications of next-generation sequencing. Brief. Bioinform. 2010, 11, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Guarnieri, M.T.; Pienkos, P.T. Algal omics: Unlocking bioproduct diversity in algae cell factories. Photosynth. Res. 2015, 123, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Vavricka, C.J.; Hasunuma, T.; Kondo, A. Dynamic metabolomics for engineering biology: Accelerating learning cycles for bioproduction. Trends Biotechnol. 2020, 38, 68–82. [Google Scholar] [CrossRef]
- Lin, W.-R.; Tan, S.-I.; Hsiang, C.-C.; Sung, P.-K.; Ng, I.-S. Challenges and opportunity of recent genome editing and multi-omics in cyanobacteria and microalgae for biorefinery. Bioresour. Technol. 2019, 291, 121932. [Google Scholar] [CrossRef]
- Hegde, K.; Chandra, N.; Sarma, S.J.; Brar, S.K.; Veeranki, V.D. Genetic engineering strategies for enhanced biodiesel production. Mol. Biotechnol. 2015, 57, 606–624. [Google Scholar] [CrossRef]
- Daboussi, F.; Leduc, S.; Marechal, A.; Dubois, G.; Guyot, V.; Perez-Michaut, C.; Amato, A.; Falciatore, A.; Juillerat, A.; Beurdeley, M. Genome engineering empowers the diatom Phaeodactylum tricornutum for biotechnology. Nat. Commun. 2014, 5, 3831. [Google Scholar] [CrossRef]
- Jinkerson, R.E.; Jonikas, M.C. Molecular techniques to interrogate and edit the Chlamydomonas nuclear genome. Plant J. 2015, 82, 393–412. [Google Scholar] [CrossRef] [PubMed]
- Valverde, F.; Romero-Campero, F.J.; León, R.; Guerrero, M.G.; Serrano, A. New challenges in microalgae biotechnology. Eur. J. Protistol. 2016, 55, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Hachicha, R.; Elleuch, F.; Ben Hlima, H.; Dubessay, P.; de Baynast, H.; Delattre, C.; Pierre, G.; Hachicha, R.; Abdelkafi, S.; Michaud, P.; et al. Biomolecules from Microalgae and Cyanobacteria: Applications and Market Survey. Appl. Sci. 2022, 12, 1924. [Google Scholar] [CrossRef]
- Jareonsin, S.; Pumas, C. Advantages of Heterotrophic Microalgae as a Host for Phytochemicals Production. Front. Bioeng. Biotechnol. 2021, 9, 58. [Google Scholar] [CrossRef]
- Mehariya, S.; Goswami, R.K.; Karthikeysan, O.P.; Verma, P. Microalgae for high-value products: A way towards green nutraceutical and pharmaceutical compounds. Chemosphere 2021, 280, 130553. [Google Scholar] [CrossRef]


| Microalgae Strain | Gene/Target Site | Approach | Results | References |
|---|---|---|---|---|
| Chlamydomonas sp. JSC4 | lut1 and zep | High-intensity light induced repression of lut1 and zep | High lutein productivity (5.08 mg/L/d) | Ma et al. [76] |
| Haematococcus pluvialis | Endogenous phytoenedesaturase (PDS) | Codon optimized/ overexpressed | Accumulation of astaxanthin up to 67% higher | Galarza et al. [83] |
| Chlamydomonas reinhardtii | Zeaxanthin epoxidase (ZEP) | DNA-free CRISPR-Cas9, knock-out mutant | Increase in both zeaxanthin content and productivity by 56- and 47-fold, respectively | Baek et al. [81] |
| Haematococcus pluvialis | β-carotene ketolase (bkt) | Cloning and overexpressed | Increase in total carotenoids and astaxanthin content by 2–3-fold higher | Kathiresan et al. [79] |
| Haematococcus pluvialis | HpDGAT1 | Upregulated expression | Increase in esterified astaxanthin (EAST) | Cui et al. [84] |
| Haematococcus pluvialis | β-carotene ketolase and b-carotene hydroxylase | Cloning and expression plasmids’ construction | Genes PSY, PDS, ZDS, LCYB expressed 2~4 fold higher, with amount of astaxanthin of 5.56 mg/g dry weight | Chen et al. [85] |
| Chlamydomonas reinhardtii | β-carotene ketolase (CrBKT) | Overexpression of the optimized CrBKT | Up to 50% of native carotenoids could be converted into astaxanthin | Perozeni et al. [86] |
| Phaeodactylum tricornutum | dxs and psy | Transcriptional upregulation | 2.4-fold and a 1.8-fold higher fucoxanthin content, respectively | Eilers et al. [87] |
| Phaeodactylum tricornutum | Phytoene synthase gene (psy) | Transformation and gene expression | Increased the fucoxanthin content by approximately 1.45-fold | Kadono et al. [88] |
| Dunaliella tertiolecta | Carotenoid biosynthesis-related (CBR) | Antisense expression and overexpression | Zeaxanthin increased with the increasing irradiation time by 2.22-fold | Zhang et al. [89] |
| Chlamydomonas reinhardtii | Bifunctional PBS gene | Heterologous expression | 38% enhancement in β-carotene along with 60% increase in the lutein | Rathod et al. [90] |
| Chlamydomonas reinhardtii | DXS and DXR | Overexpressed via nuclear transformation | Increased lutein and β-carotene by 1.9-fold and 1.7-fold per cell, respectively | Morikawa et al. [91] |
| Dunaliella salina | Introduction of a bkt gene | Transformation procedure | Astaxanthin and canthaxanthin with maximum content of 3.5 and 1.9 lg/g DW, respectively | Anila et al. [78] |
| Dunaliella tertiolecta | mp3 | Random mutagenesis | 10–15% higher cellular zeaxanthin content | Kim et al. [82] |
| Chlorella zofingiensis | Phytoene desaturase (PDS) | Overexpression | Increase total carotenoid and astaxanthin production by 32.1% and 54.1% respectively. | Liu et al. [77] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grama, S.B.; Liu, Z.; Li, J. Emerging Trends in Genetic Engineering of Microalgae for Commercial Applications. Mar. Drugs 2022, 20, 285. https://doi.org/10.3390/md20050285
Grama SB, Liu Z, Li J. Emerging Trends in Genetic Engineering of Microalgae for Commercial Applications. Marine Drugs. 2022; 20(5):285. https://doi.org/10.3390/md20050285
Chicago/Turabian StyleGrama, Samir B., Zhiyuan Liu, and Jian Li. 2022. "Emerging Trends in Genetic Engineering of Microalgae for Commercial Applications" Marine Drugs 20, no. 5: 285. https://doi.org/10.3390/md20050285
APA StyleGrama, S. B., Liu, Z., & Li, J. (2022). Emerging Trends in Genetic Engineering of Microalgae for Commercial Applications. Marine Drugs, 20(5), 285. https://doi.org/10.3390/md20050285