
The Biochemical Composition and Antioxidant Properties of Fucus vesiculosus from the Arctic Region

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results and Discussion
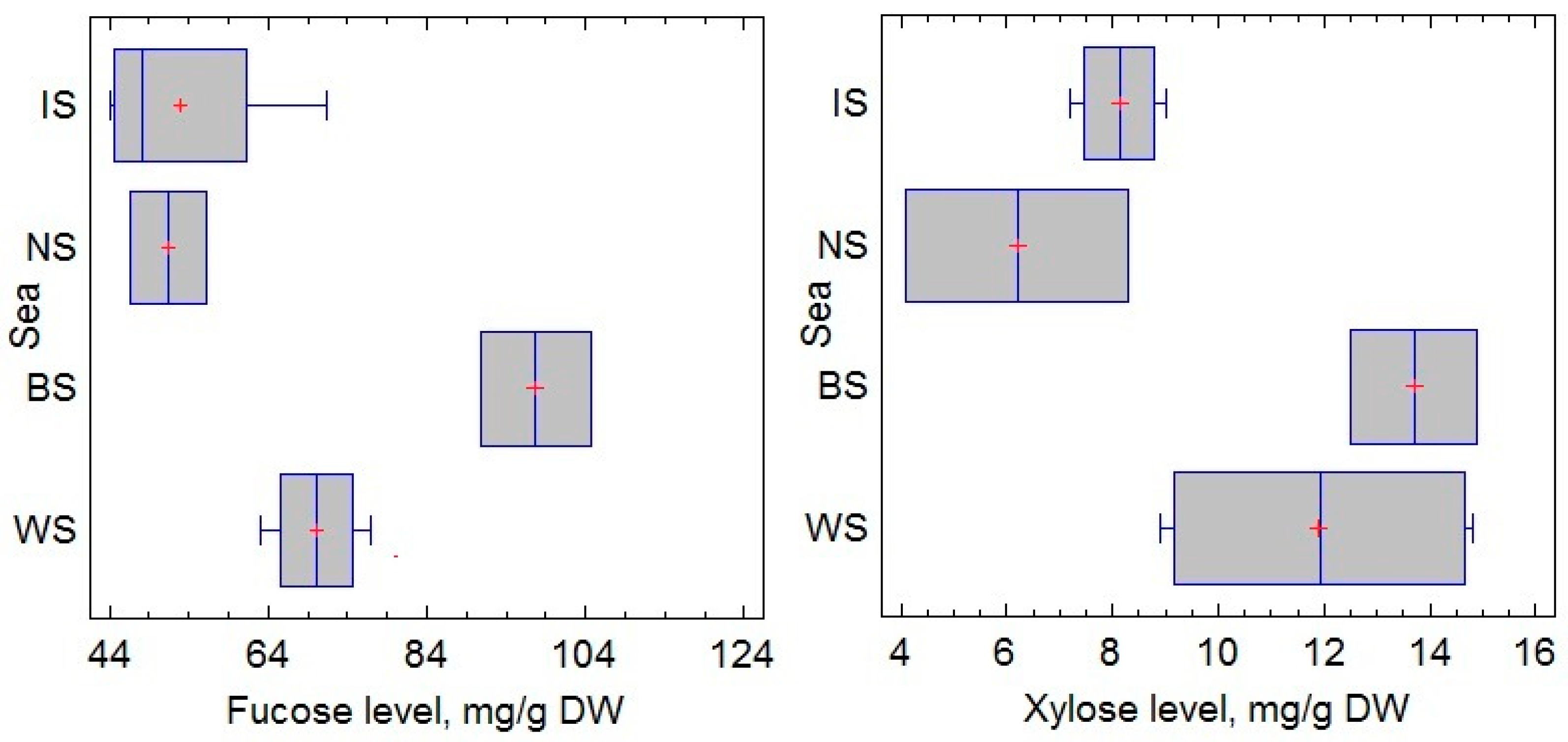
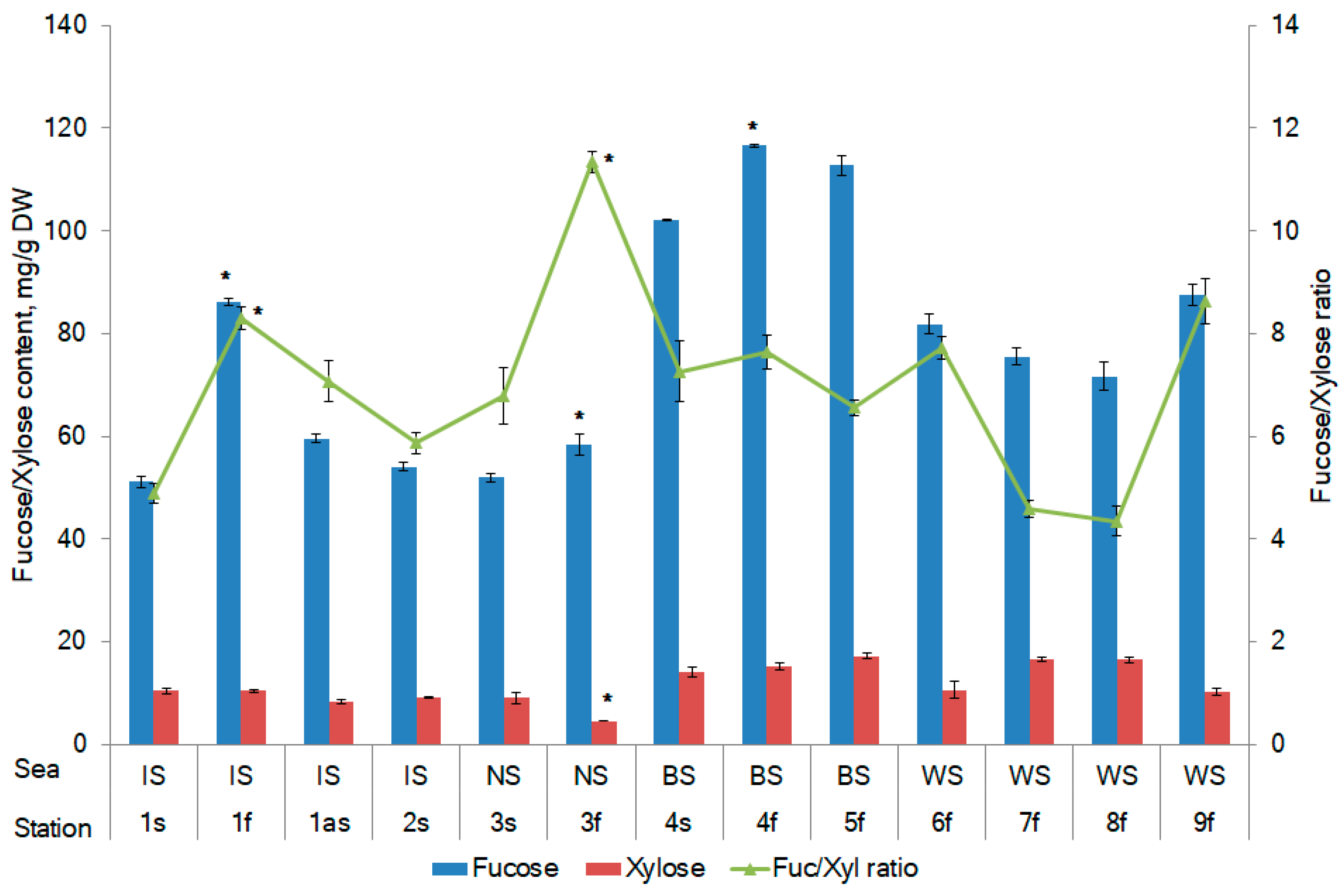
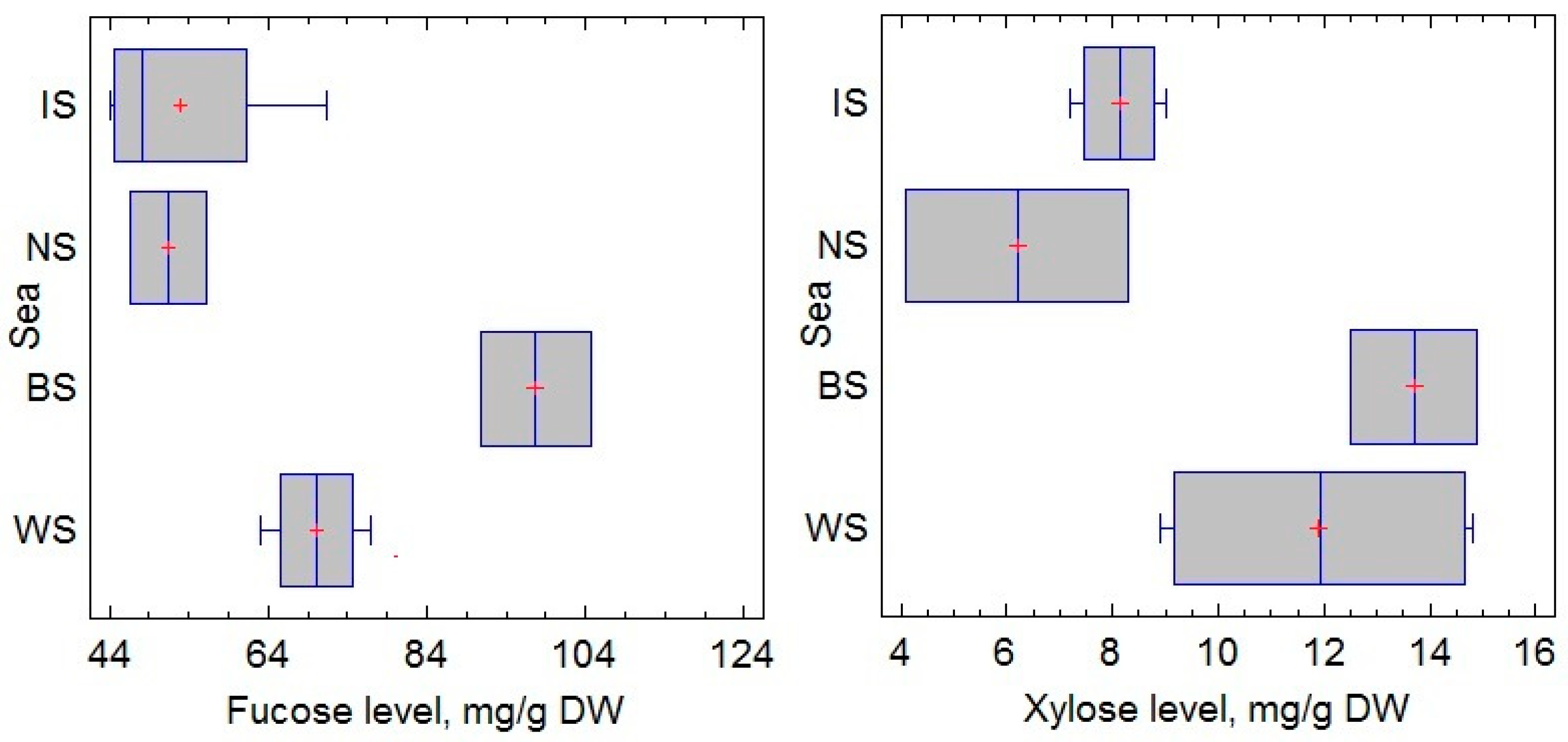
2.1. Fucose and Xylose Content
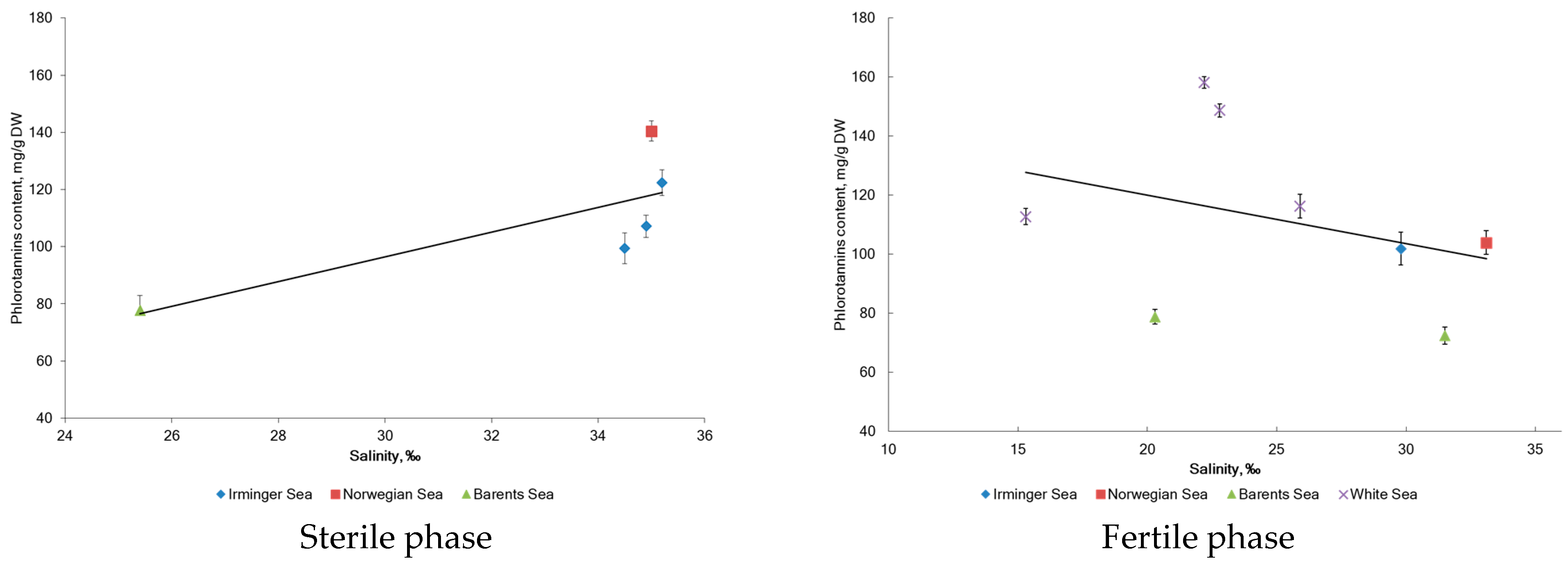
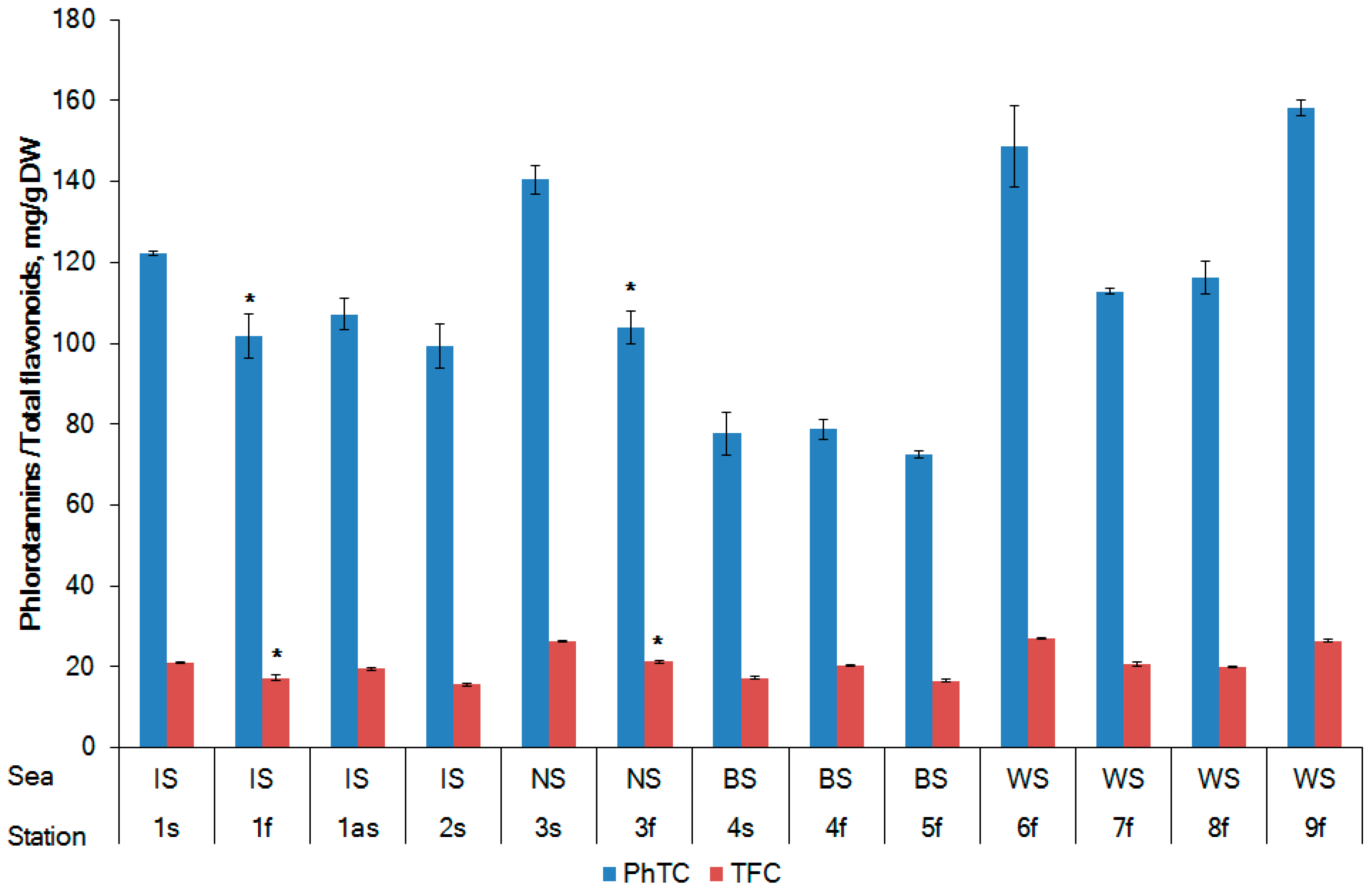
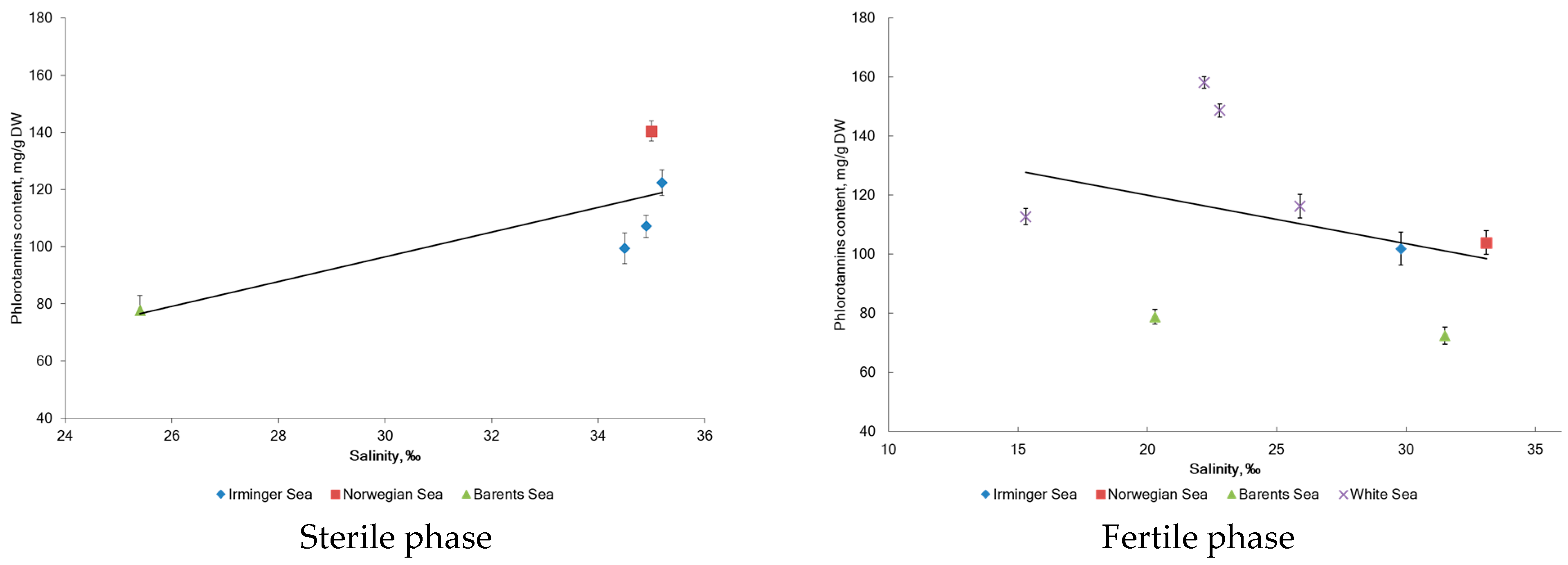
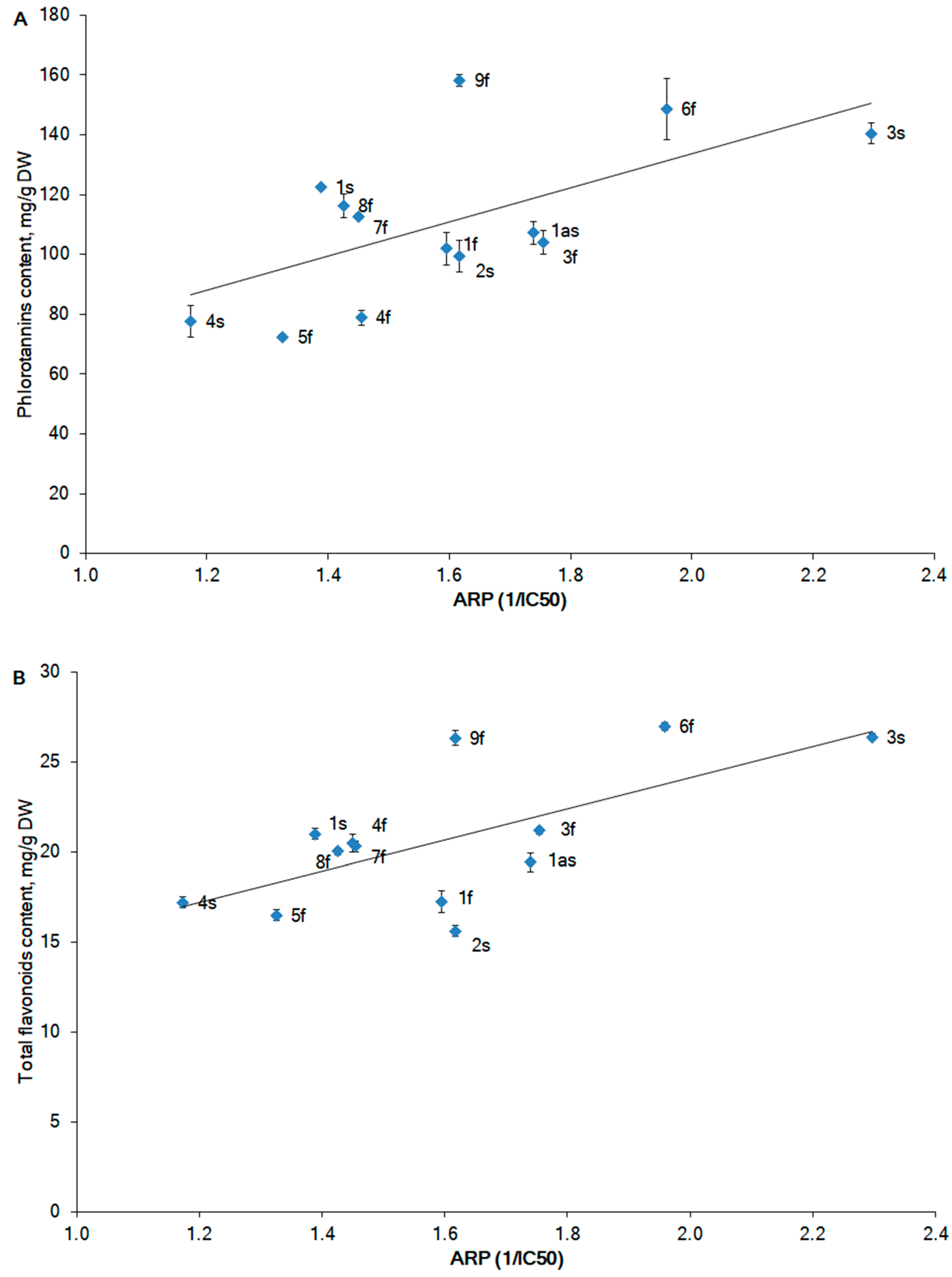
2.2. Phlorotannin Content
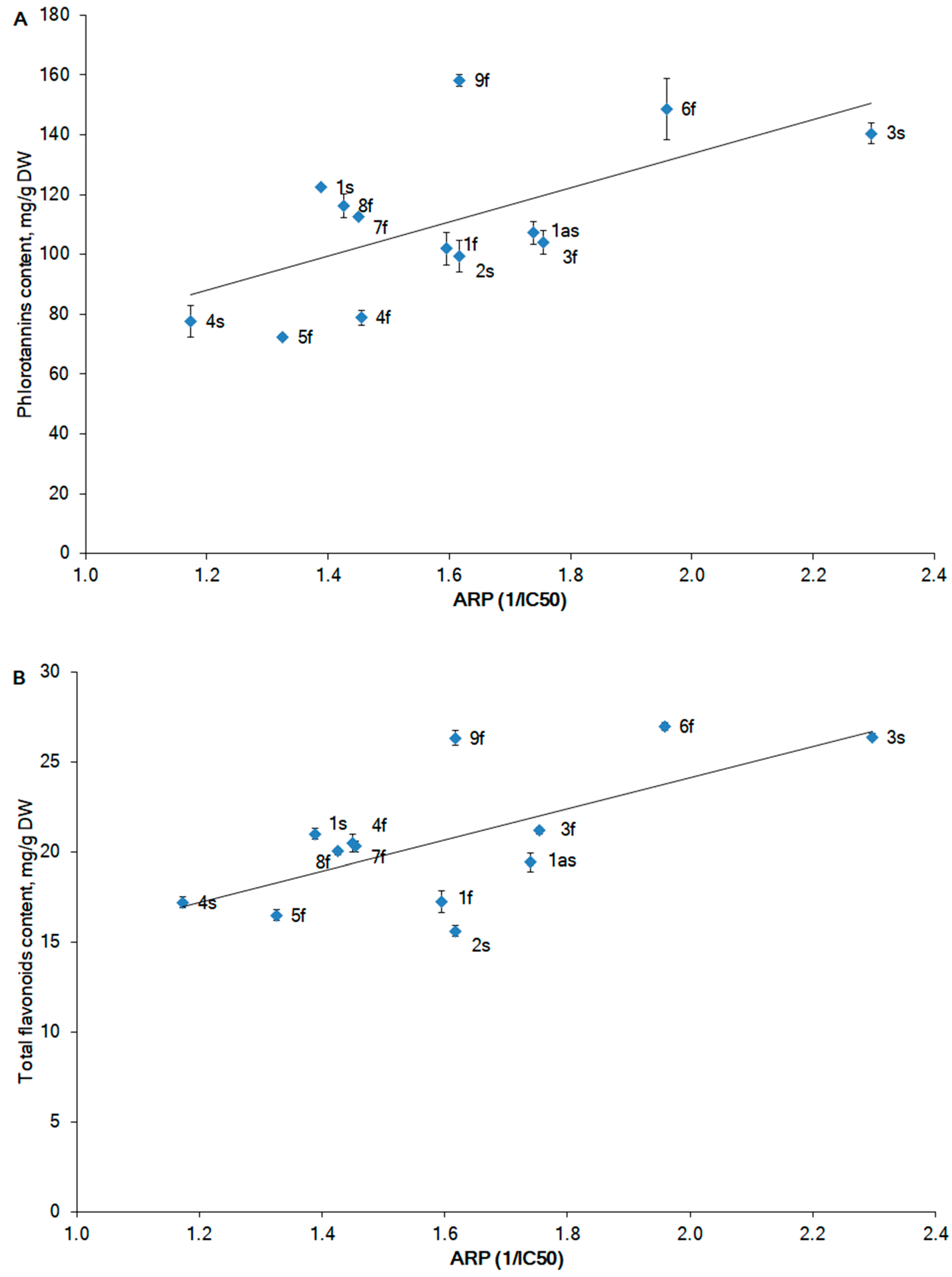
2.3. Antioxidant Activity
2.4. Ash Contennt
2.5. Elemental Concentrations
3. Materials and Methods
3.1. Alga Sample Collection
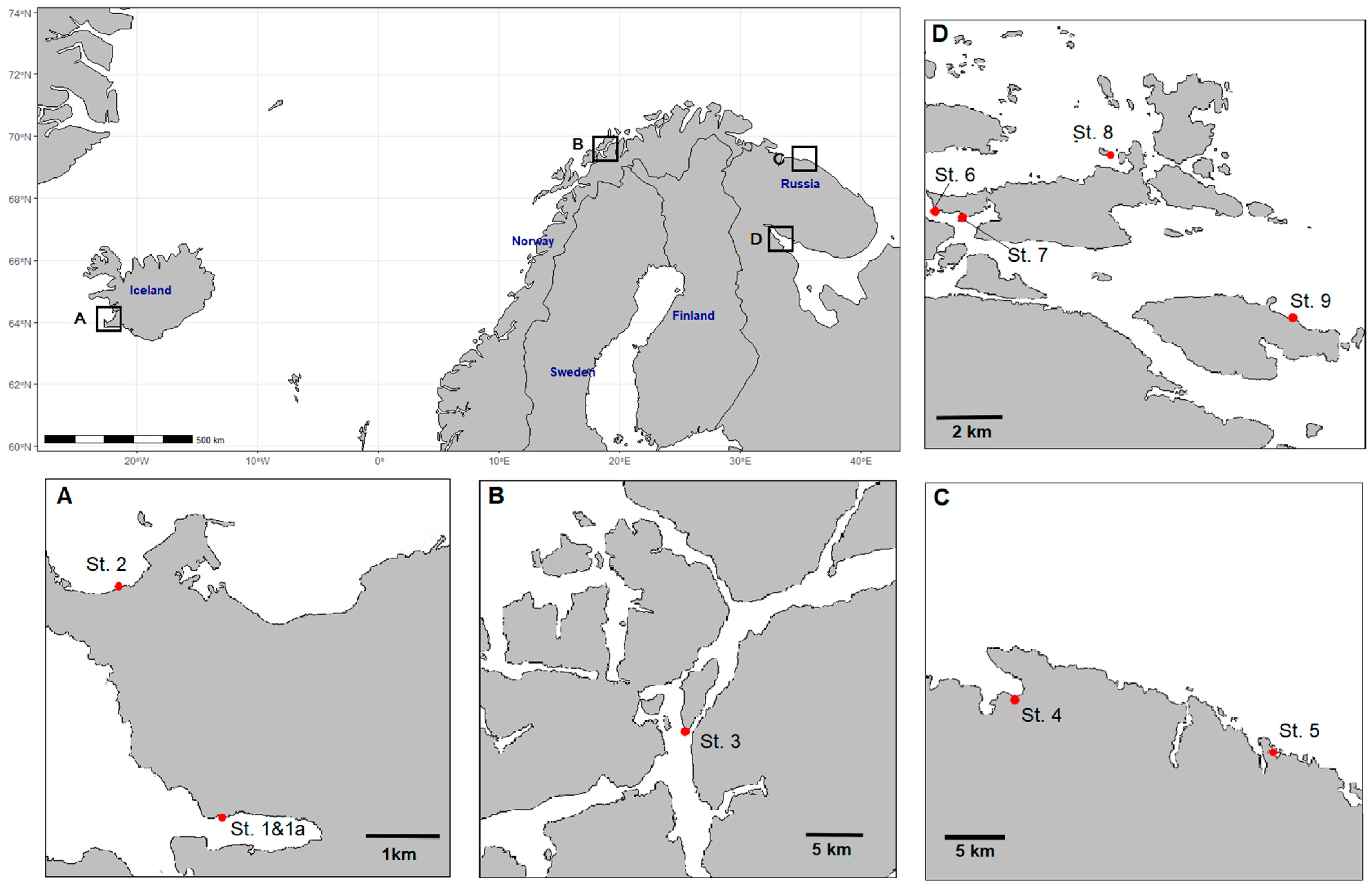
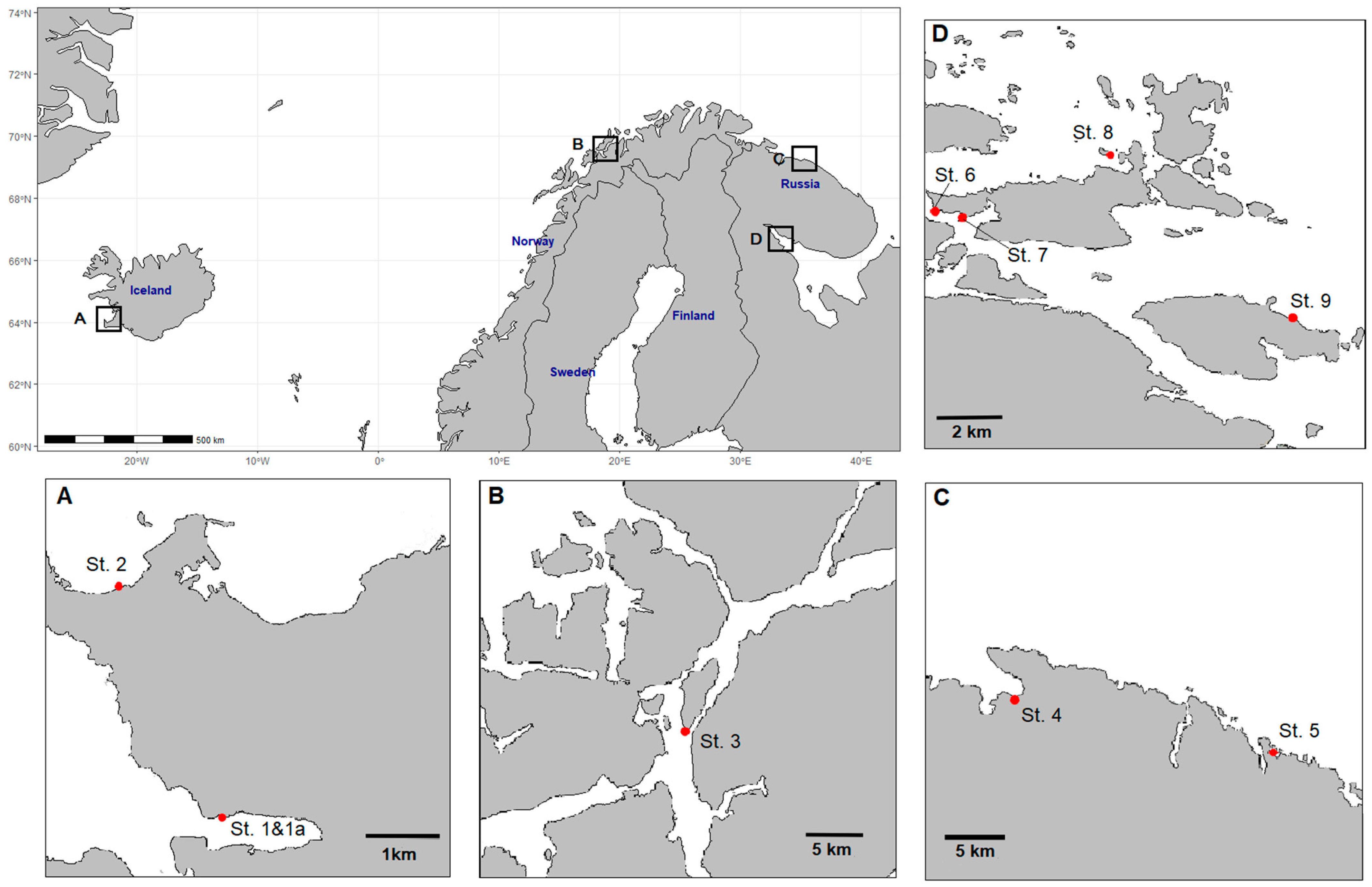
3.2. Sampling Stations
3.3. Chemicals
3.4. Fucose–Xylose Composition
3.5. Determination of the Phlorotannin Content, Total Flavonoids, and Antioxidant Activity
3.6. Ash Content
3.7. Elemental Analysis
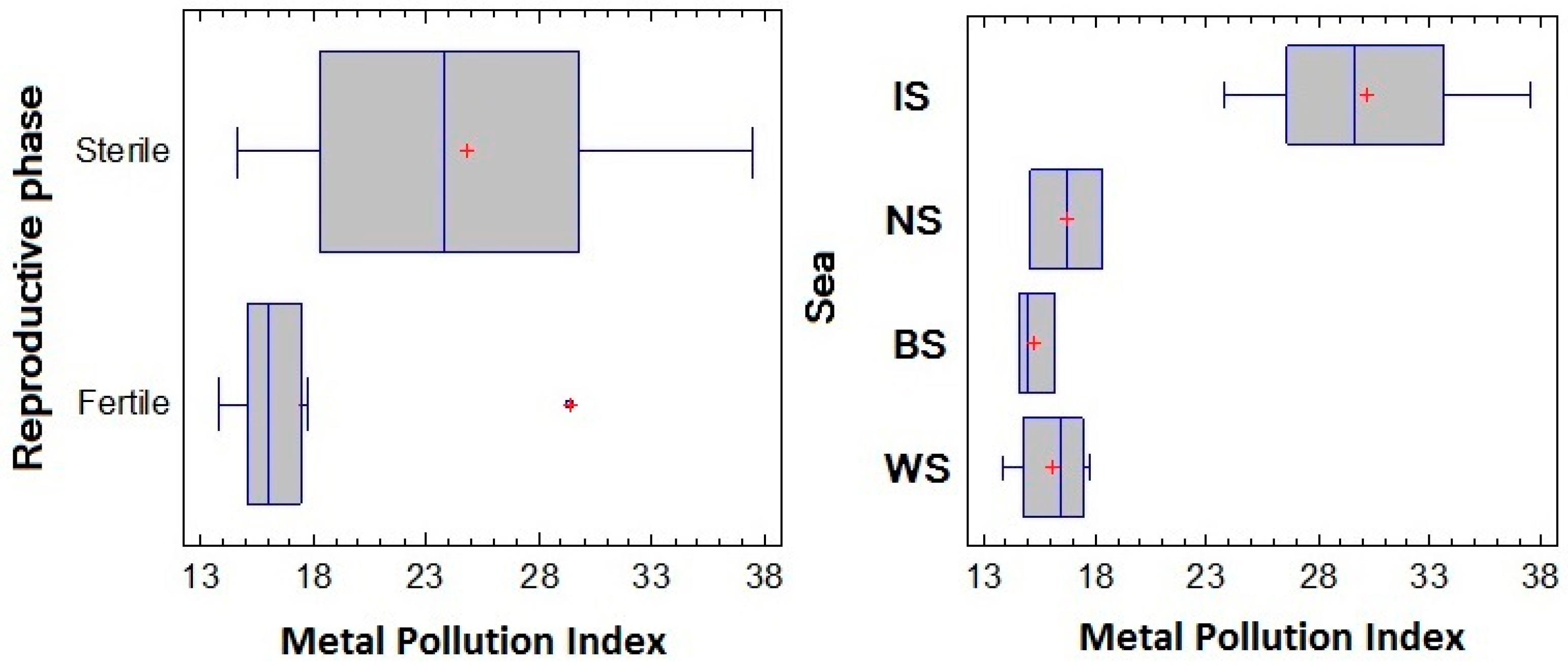
3.8. Metal Pollution Index
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aminina, N.M.; Vishnevskaya, T.I.; Karaulova, E.P.; Epur, N.V.; Yakush, E.V. Prospects for the use of commercial and potentially commercial brown algae of the far eastern seas as a source of polyphenols. Russ. J. Mar. Biol. 2020, 46, 34–41. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, Y.; Zhang, Y.; Yang, Y.; Wang, P.; Imre, B.; Wong, A.C.Y.; Hsieh, Y.S.Y.; Wang, D. Brown algae carbohydrates: Structures, pharmaceutical properties, and research challenges. Mar. Drugs 2021, 19, 620. [Google Scholar] [CrossRef] [PubMed]
- Lomartire, S.; Gonçalves, A.M.M. An Overview of potential seaweed-derived bioactive compounds for pharmaceutical applications. Mar. Drugs 2022, 20, 141. [Google Scholar] [CrossRef]
- Torn, K.; Krause-Jensen, D.; Martin, G. Present and past depth distribution of bladderwrack (Fucus vesiculosus) in the Baltic Sea. Aquat. Bot. 2006, 84, 53–62. [Google Scholar] [CrossRef]
- Pereira, L. Edible Seaweeds of the World. CRC Press: Coimbra, Portugal, 2016; ISBN 9781498730471. [Google Scholar]
- Fitton, J.H.; Dell’Acqua, G.; Gardiner, V.-A.; Karpiniec, S.S.; Stringer, D.N.; Davis, E. Topical benefits of two fucoidan-rich extracts from marine macroalgae. Cosmetics 2015, 2, 66–81. [Google Scholar] [CrossRef] [Green Version]
- Blunden, G.; Challen, S.B.; Woods, D.L. Seaweed extracts as fertilisers. J. Sci. Food Agric. 1968, 19, 289–292. [Google Scholar] [CrossRef]
- Campbell, M.; Ortuño, J.; Ford, L.; Davies, D.R.; Koidis, A.; Walsh, P.J.; Theodoridou, K. The Effect of ensiling on the nutritional composition and fermentation characteristics of brown seaweeds as a ruminant feed ingredient. Animals 2020, 10, 1019. [Google Scholar] [CrossRef] [PubMed]
- Catarino, M.D.; Silva, A.M.S.; Mateus, N.; Cardoso, S.M. Optimization of phlorotannins extraction from Fucus vesiculosus and evaluation of their potential to prevent metabolic disorders. Mar. Drugs 2019, 17, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, C.; Paíga, P.; Marques, M.; Neto, T.; Carvalho, A.P.; Paiva, A.; Simões, P.; Costa, L.; Bernardo, A.; Fernández, N.; et al. Multi-Step subcritical water extracts of Fucus vesiculosus L. and Codium tomentosum stackhouse: Composition, health-benefits and safety. Processes 2021, 9, 893. [Google Scholar] [CrossRef]
- Obluchinskaya, E.D.; Pozharitskaya, O.N.; Flisyuk, E.V.; Shikov, A.N. Formulation, optimization and in vivo evaluation of fucoidan-based cream with anti-inflammatory properties. Mar. Drugs 2021, 19, 643. [Google Scholar] [CrossRef]
- Ayrapetyan, O.N.; Obluchinskaya, E.D.; Zhurishkina, E.V.; Skorik, Y.A.; Lebedev, D.V.; Kulminskaya, A.A.; Lapina, I.M. Antibacterial properties of fucoidans from the brown algae Fucus vesiculosus L. of the Barents Sea. Biology 2021, 10, 67. [Google Scholar] [CrossRef] [PubMed]
- Fitton, J.H.; Stringer, D.N.; Park, A.Y.; Karpiniec, S.S. Therapies from fucoidan: New developments. Mar. Drugs 2019, 17, 571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daurtseva, A.V.; Obluchinskay, E.D. The stability of pigments in the thalli and extracts of the Barents Sea fucus algae. Vestn. MSTU 2019, 22, 314–321. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.; Cardoso, S.M. Phycochemical constituents and biological activities of Fucus spp. Mar. Drugs 2018, 16, 249. [Google Scholar] [CrossRef] [Green Version]
- Obluchinskaya, E.; Daurtseva, A. Effects of air drying and freezing and long-term storage on phytochemical composition of brown seaweeds. J. Appl. Phycol. 2020, 32, 4235–4249. [Google Scholar] [CrossRef]
- Obluchinskaya, E.D.; Zakharova, L.V. Comparative study of polyphenols of brown algae of the Barents sea and the White sea, as well as the waters of the North Atlantic. Chem. Plant Raw Mater. 2020, 4, 129–137. [Google Scholar] [CrossRef]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [Green Version]
- Zayed, A.; Muffler, K.; Hahn, T.; Rupp, S.; Finkelmeier, D.; Burger-Kentischer, A.; Ulber, R. Physicochemical and biological characterization of fucoidan from Fucus vesiculosus purified by dye affinity chromatography. Mar. Drugs 2016, 14, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imbs, T.I.; Skriptsova, A.V.; Zvyagintseva, T.N. Antioxidant activity of fucose-containing sulfated polysaccharides obtained from Fucus evanescens by different extraction methods. J. Appl. Phycol. 2014, 27, 545–553. [Google Scholar] [CrossRef]
- Obluchinskaya, E.D. Antioxidant complex extracts from fucus algae of the Barents Sea. Vestn. MSTU 2018, 21, 395–401. [Google Scholar] [CrossRef]
- Park, M.K.; Jung, U.; Roh, C. Fucoidan from marine brown algae inhibits lipid accumulation. Mar. Drugs 2011, 9, 1359–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozharitskaya, O.N.; Obluchinskaya, E.D.; Shikov, A.N. Mechanisms of bioactivities of fucoidan from the brown seaweed Fucus vesiculosus L. of the Barents Sea. Mar. Drugs 2020, 18, 275. [Google Scholar] [CrossRef] [PubMed]
- Wijesinghe, W.A.J.P.; Jeon, Y.J. Biological activities and potential industrial applications of fucose rich sulfated polysaccharides and fucoidans isolated from brown seaweeds: A review. Carbohyd. Polym. 2012, 88, 13–20. [Google Scholar] [CrossRef]
- Imbs, T.I.; Zvyagintseva, T.N. Phlorotannins are polyphenolic metabolites of brown algae. Russ. J. Mar. Biol. 2018, 44, 263–273. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Fucaceae: A source of bioactive phlorotannins. Int. J. Mol. Sci. 2017, 18, 1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, R.M.; Ramalho Ribeiro, A.; Patinha, C.; Silva, A.; Cardoso, S.M.; Costa, R. Water extraction kinetics of bioactive compounds of Fucus vesiculosus. Molecules 2019, 24, 3408. [Google Scholar] [CrossRef] [Green Version]
- Singh, I.P.; Sidana, J. Phlorotannins. In Functional Ingredients from Algae for Foods and Nutraceuticals; Elsevier: Amsterdam, The Netherlands, 2013; pp. 181–204. [Google Scholar]
- Circuncisão, A.R.; Catarino, M.D.; Cardoso, S.M.; Silva, A.M.S. Minerals from macroalgae origin: Health benefits and risks for consumers. Mar. Drugs 2018, 16, 400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Pan, X.D.; Huang, B.F.; Han, J.L. Distribution of metals and metalloids in dried seaweeds and health risk to population in southeastern China. Sci. Rep. 2018, 8, 3578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skriptsova, A.V.; Shevchenko, N.M.; Tarbeeva, D.V.; Zvyagintseva, T.N. Comparative study of polysaccharides from reproductive and sterile tissues of five brown seaweeds. Mar. Biotechnol. 2012, 14, 304–311. [Google Scholar] [CrossRef]
- Roleda, M.Y.; Marfaing, H.; Desnica, N.; Jónsdóttir, R.; Skjermo, J.; Rebours, C.; Nitschke, U. Variations in polyphenol and heavy metal contents of wild-harvested and cultivated seaweed bulk biomass: Health risk assessment and implication for food applications. Food Control 2019, 95, 121–134. [Google Scholar] [CrossRef]
- Lobban, C.S.; Harrison, P.J. (Eds.) Seaweed Ecology and Physiology; Cambridge University Press: Cambridge, UK, 1994; 366p. [Google Scholar]
- Birkemeyer, C.; Osmolovskaya, N.; Kuchaeva, L.; Tarakhovskaya, E. Distribution of natural ingredients suggests a complex network of metabolic transport between source and sink tissues in the brown alga Fucus vesiculosus. Planta 2019, 249, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I.; et al. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imbs, T.I.; Shevchenko, N.M.; Sukhoverkhov, S.V.; Semenova, T.L.; Skriptsova, A.V.; Zvyagintseva, T.N. Seasonal variations of the composition and structural characteristics of polysaccharides from the brown alga Costaria costata. Chem. Nat. Compd. 2009, 45, 786–791. [Google Scholar] [CrossRef]
- Skriptsova, A.V.; Shevchenko, N.M.; Zvyagintseva, T.N.; Imbs, T.I. Monthly changes in the content and monosaccharide composition of fucoidan from Undaria pinnatifida (Laminariales, Phaeophyta). J. Appl. Phycol. 2010, 22, 79–86. [Google Scholar] [CrossRef]
- Honya, M.; Mori, H.; Anzai, M.; Araki, Y.; Nisizawa, K. Monthly changes in the content of fucans, their constituent sugars and sulphate in cultured Laminaria japonica. Hydrobiologia 1999, 398, 411–416. [Google Scholar] [CrossRef]
- Pedersen, A. Studies on phenol content and heavy metal uptake in fucoids. In Eleventh International Seaweed Symposium; Bird, C.J., Ragan, M.A., Eds.; Springer: Dordrecht, The Netherlands, 1984; pp. 498–504. [Google Scholar]
- Chernane, H.; Mansori, M.; Latique, S.; El Kaoua, M. Evaluation of antioxidant capacity of methanol extract and its solvent fractions obtained from four Moroccan macro algae species. Eur. Sci. J. 2014, 10, 35–48. [Google Scholar]
- Kim, S.K.; Himaya, S.W.A. Medicinal effects of phlorotannins from marine brown algae. In Advances in Food and Nutrition Research; Kim, S., Ed.; Academic Press: San Diego, CA, USA, 2011; Volume 64, pp. 97–109. [Google Scholar]
- Pavia, H.; Brock, E. Extrinsic factors influencing phlorotannin production in the brown alga. Mar. Ecol. Prog. Ser. 2000, 193, 285–294. [Google Scholar] [CrossRef]
- Pavia, H.; Toth, G.B. Influence of light and nitrogen on the phlorotannin content of the brown seaweeds Ascophyllum nodosum and Fucus vesiculosus. Hydrobiology 2000, 440, 299–305. [Google Scholar] [CrossRef]
- Connan, S.; Goulard, F.; Stiger, V.; Deslandes, E.; Gall, E.A. Interspecific and temporal variation in phlorotannin levels in an assemblage of brown algae. Bot. Mar. 2004, 47, 410–416. [Google Scholar] [CrossRef]
- Ragan, M.A.; Jensen, A. Quantitative studies on brown algal phenols. II. Seasonal variation in polyphenol content of Ascophyllum nodosum (L.) Le Jol. and Fucus vesiculosus (L.). J. Exp. Mar. Biol. Ecol. 1978, 34, 245–258. [Google Scholar] [CrossRef]
- Kamiya, M.; Nishio, T.; Yokoyama, A.; Yatsuya, K.; Nishigaki, T.; Yoshikawa, S.; Ohki, K. Seasonal variation of phlorotannin in sargassacean species from the coast of the Sea of Japan. Phycol. Res. 2010, 58, 53–61. [Google Scholar] [CrossRef]
- Roleda, M.Y.; Wiencke, C.; Lüder, U.H. Impact of ultraviolet radiation on cell structure, UV-absorbing compounds, photosynthesis, DNA damage, and germination in zoospores of Arctic Saccorhiza dermatodea. J. Exp. Bot. 2006, 57, 3847–3856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, S.; Abu-Ghannam, N.; Gupta, S. An assessment of the antioxidant and antimicrobial activity of six species of edible Irish seaweeds. Int. Food Res. J. 2010, 17, 205–220. [Google Scholar]
- Marinho, G.S.; Sørensen, A.D.M.; Safafar, H.; Pedersen, A.H.; Holdt, S.L. Antioxidant content and activity of the seaweed Saccharina latissima: A seasonal perspective. J. Appl. Phycol. 2019, 31, 1343–1354. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Jónsdóttir, R.; Ólafsdóttir, G. Total phenolic compounds, radical scavenging and metal chelation of extracts from Icelandic seaweeds. Food Chem. 2009, 116, 240–248. [Google Scholar] [CrossRef]
- Tierney, M.S.; Smyth, T.J.; Rai, D.K.; Soler-Vila, A.; Croft, A.K.; Brunton, N. Enrichment of polyphenol contents and antioxidant activities of Irish brown macroalgae using food-friendly techniques based on polarity and molecular size. Food Chem. 2013, 139, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.J.; Aida, W.M.W.; Maskat, M.Y.; Mamot, S.; Ropien, J.; Mohd, D.M. Isolation and antioxidant capacity of fucoidan from selected Malaysian seaweeds. Food Hydrocoll. 2014, 42, 280–288. [Google Scholar] [CrossRef]
- Palanisamy, S.; Vinosha, M.; Manikandakrishnan, M.; Anjali, R.; Rajasekar, P.; Marudhupandi, T.; Manikandan, R.; Vaseeharan, B.; Prabhu, N.M. Investigation of antioxidant and anticancer potential of fucoidan from Sargassum polycystum. Int. J. Biol. Macromol. 2018, 116, 151–161. [Google Scholar] [CrossRef]
- Paiva, L.; Lima, E.; Neto, A.I.; Baptista, J. Seasonal variability of the biochemical composition and antioxidant properties of Fucus spiralis at two Azorean Islands. Mar. Drugs 2018, 16, 248. [Google Scholar] [CrossRef] [Green Version]
- Marinho-Soriano, E.; Fonseca, P.C.; Carneiro, M.A.A.; Moreira, W.S.C. Seasonal variation in the chemical composition of two tropical seaweeds. Bioresour. Technol. 2006, 97, 2402–2406. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.B.; Jones, J.M.; Kubacki, M.L.; Bridgeman, T. Classification of macroalgae as fuel and its thermochemical behaviour. Bioresour. Technol. 2008, 99, 6494–6504. [Google Scholar] [CrossRef]
- Yuan, Y.V. Marine algal constituens. In Marine Nutraceuticals and Functional Foods; Barrow, C., Shahidi, F., Eds.; CRC Press & Taylor and Francis Group: Boca Raton, FL, USA, 2008; pp. 259–296. [Google Scholar]
- Rupérez, P. Mineral content of edible marine seaweeds. Food Chem. 2002, 79, 23–26. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Agregán, R.; Munekata, P.E.; Franco, D.; Carballo, J.; Şahin, S.; Lacomba, R.; Barba, F.J. Proximate composition and nutritional value of three macroalgae: Ascophyllum nodosum, Fucus vesiculosus and Bifurcaria bifurcata. Mar. Drugs 2017, 15, 360. [Google Scholar] [CrossRef] [Green Version]
- Truus, K.; Vaher, M.; Taure, I. Algal biomass from Fucus vesiculosus (Phaeophyta): Investigation of the mineral and alginate components. Proc. Estonian Acad. Sci. Chem. 2001, 50, 95–103. [Google Scholar]
- Renaud, S.M.; Luong-Van, J.T. Seasonal variation in the chemical composition of tropical Australian marine macroalgae. J. Appl. Phycol. 2006, 18, 381–387. [Google Scholar] [CrossRef]
- Bogolitsyn, K.G.; Kaplitsin, P.A.; Kashina, E.M.; Ivanchenko, N.L.; Kokryatskaya, N.M.; Ovchinnikov, D.V. Features of the mineral composition of brown algae in the White and Barents Seas. Chem. Plant Raw Mater. 2014, 1, 243–250. [Google Scholar] [CrossRef]
- Titlyanov, E.A.; Titlyanova, T.V.; Belous, O.S. Useful marine plants and their uses. Izv. TINRO 2011, 164, 140–156. [Google Scholar]
- Burtin, P. Nutritional value of seaweeds. Electron. J. Environ. Agric. Food chem. 2003, 2, 498–503. [Google Scholar]
- Food Raw Materials and Food Products Hygiene Requirements for Safety and Nutritional Food. Sanitary-Epidemiological Rules and Standards; SanPiN 2.3.2.1078-01; Federal Center for Hygiene and Epidemiology of Rospotrebnadzor: Moscow, Russia, 2002. [Google Scholar]
- Joint FAO/WHO Expert Committee on Food Additives. In Proceedings of the Summary and Conclusions of the Sixty-Seventh Meeting of the Joint FAO/WHO Expert Committee on Food Additives, Rome, Italy, 20–29 June 2006.
- World Health Organization. Evaluation of Certain Food Additives and Contaminants. Seventy-Second Report of the Joint FAO/WHO Expert Committee on Food Additives; WHO Technical Report Series No. 959; WHO: Geneva, Switzerland, 2011.
- World Health Organization. Evaluation of Certain Food Additives and Contaminants. Seventy-Third Report of the Joint FAO/WHO Expert Committee on Food Additives, Geneva, 2010; WHO Technical Report Series No. 960; WHO: Geneva, Switzerland, 2011.
- European Food Safety Authority. Tolerable Upper Intake Levels for Vitamins and Minerals. February 2006. Available online: www.efsa.europa.eu/sites/default/files/efsa_rep/blobserver_assets/ndatolerableuil.pdf (accessed on 4 February 2022).
- Norms of Physiological Needs Energy and Nutrients for Various Groups of the Population of the Russian Federation; Methodical Recommendations MR 2.3.1.2432−08; Federal Center for Hygiene and Epidemiology of Rospotrebnadzor: Moscow, Russia, 2009.
- Phaneuf, D.; Côté, I.; Dumas, P.; Ferron, L.A.; LeBlanc, A. Evaluation of the contamination of marine algae (seaweed) from the St. Lawrence River and likely to be consumed by humans. Environ. Res. 1999, 80, S175–S182. [Google Scholar] [CrossRef]
- Almela, C.; Algora, S.; Benito, V.; Clemente, M.J.; Devesa, V.; Suner, M.A.; Velez, D.; Montoro, R. Heavy metal, total arsenic, and inorganic arsenic contents of algae food products. J. Agric. Food Chem. 2002, 50, 918–923. [Google Scholar] [CrossRef] [PubMed]
- Besada, V.; Andrade, J.M.; Schultze, F.; Gonzalez, J.J. Heavy metals in edible seaweeds commercialised for human consumption. J. Mar. Syst. 2009, 75, 305–313. [Google Scholar] [CrossRef]
- Desideri, D.; Cantaluppi, C.; Ceccotto, F.; Meli, M.A.; Roselli, C.; Feduzi, L. Essential and toxic elements in seaweeds for human consumption. J. Toxicol. Environ. Health Part A 2016, 79, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Sartal, C.G.; Alonso, M.C.B.; Barrera, P.B. Arsenic in seaweed: Presence, bioavailability and speciation. In Seafood Science: Advances in Chemistry Technology and Applications; CRC Press: Boca Raton, FL, USA; Taylor & Francis Group: Abingdon, UK, 2014; pp. 276–351. [Google Scholar]
- IARC Monographs on the Evaluation of Carcinogenic Risks to Humans Volume 100C. Arsenic, Metals, Fibres, and Dusts. Available online: https://publications.iarc.fr/120 (accessed on 4 February 2022).
- Obluchinskaya, E.; Zakharova, L. Metal concentrations in three species of Fucus L. on the Murmansk coast of the Barents Sea. Polar Sci. 2021, 28, 100646. [Google Scholar] [CrossRef]
- Ruuskanen, A.; Bäck, S. Does environmental stress affect fertility and frond regeneration of Fucus vesiculosus? Finnish Zool. Bot. Publ. Board 1999, 36, 285–290. [Google Scholar]
- Graiff, A.; Dankworth, M.; Wahl, M.; Karsten, U.; Bartsch, I. Seasonal variations of Fucus vesiculosus fertility under ocean acidification and warming in the western Baltic Sea. Bot. Mar. 2017, 60, 239–255. [Google Scholar] [CrossRef]
- Rodríguez-Jasso, R.M.; Mussatto, S.I.; Pastrana, L.; Aguilar, C.N.; Teixeira, J.A. Extraction of sulfated polysaccharides by autohydrolysis of brown seaweed Fucus vesiculosus. J. Appl. Phycol. 2013, 25, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Ummat, V.; Tiwari, B.K.; Jaiswal, A.K.; Condon, K.; Garcia-Vaquero, M.; O’Doherty, J.; O’Donnell, C.; Rajauria, G. Optimisation of ultrasound frequency, extraction time and solvent for the recovery of polyphenols, phlorotannins and associated antioxidant activity from brown seaweeds. Mar. Drugs 2020, 18, 250. [Google Scholar] [CrossRef]
- Uribe, E.; Pardo-Orellana, C.M.; Vega-Gálvez, A.; Ah-Hen, K.S.; Pastén, A.; García, V.; Aubourg, S.P. Effect of drying methods on bioactive compounds, nutritional, antioxidant, and antidiabetic potential of brown alga Durvillaea antarctica. Dry. Technol. 2020, 38, 1915–1928. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Shikov, A.N.; Makarova, M.N.; Kosman, V.M.; Faustova, N.M.; Tesakova, S.V.; Makarov, V.G.; Galambosi, B. Anti-inflammatory activity of a HPLC-fingerprinted aqueous infusion of aerial part of Bidens tripartita L. Phytomedicine 2010, 17, 463–468. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of free radical method to evaluate antioxidant capacity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Charles, B.; Fredeen, K.J. Concepts, instrumentation and techniques in inductively coupled plasma optical emission spectrometry. Perkin Elmer Corp. 1997, 3, 2. [Google Scholar]
- Flores, É.M.D.M.; Barin, J.S.; Paniz, J.N.G.; Medeiros, J.A.; Knapp, G. Microwave-assisted sample combustion: A technique for sample preparation in trace element determination. Anal. Chem. 2004, 76, 3525–3529. [Google Scholar] [CrossRef] [PubMed]
- Usero, J.; Morillo, J.; Gracia, I. Heavy metal concentrations in molluscs from the Atlantic coast of southern Spain. Chemosphere 2005, 59, 1175–1181. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element | LOQ | Range | St. 1 | St. 1a | St. 2 | St. 3 | St. 4 | St. 5 | St. 6 | St. 7 | St. 8 | St. 9 | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| s/f | ∆ | s | s | s/f | ∆ | s/f | ∆ | f | f | f | f | f | |||
| Al | 1.6 | 28–724 | 423 ± 40 | * | 183 ± 36 | 688 ± 124 | 54 ± 8 | * | 45 ± 1 | * | 54 ± 12 | 95 ± 6 | 88 ± 5 | 42 ± 9 | 57 ± 3 |
| 724 ± 27 | ↑ | 28 ± 3 | ↓ | 62 ± 3 | ↑ | ||||||||||
| As | 6.3 | 23–58 | 35 ± 2 | 30 ± 4 | 28 ± 2 | 39 ± 1 | * | 58 ± 2 | * | 40 ± 1 | 30 ± 3 | 31 ± 2 | 36 ± 1 | 22 ± 1 | |
| 32 ± 1 | ↓ | 48 ± 2 | ↑ | 44 ± 1 | ↓ | ||||||||||
| Ba | 0.016 | 10–24 | 13 ± 0.1 | 13.8 ± 0.8 | 12 ± 0.2 | 10.2 ± 0.1 | 11.0 ± 0.2 | 10.6 ± 0.2 | 18 ± 1 | 20 ± 0.3 | 16 ± 0.2 | 24 ± 2 | |||
| 10 ± 0.3 | ↓ | 9.7 ± 0.1 | ↓ | 10.2 ± 0.1 | ↓ | ||||||||||
| Ca | 1.9 | 9756–30,093 | 30,093 ± 100 | * | 15,228 ± 230 | 18,278 ± 210 | 20,832 ± 315 | * | 11,430 ± 188 | * | 9890 ± 20 | 11,922 ± 205 | 11,666 ± 290 | 20,065 ± 260 | 11,592 ± 470 |
| 18,436 ± 265 | ↓ | 9756 ± 113 | ↓ | 12,015 ± 389 | ↑ | ||||||||||
| Co | 0.12 | 0.4–2.0 | 1.73 ± 0.03 | 1.56 ± 0.09 | 1.95 ± 0.38 | 1.38 ± 0.02 | 1.09 ± 0.01 | * | 0.59 ± 0.05 | 0.61 ± 0.03 | 0.92 ± 0.10 | 0.39 ± 0.03 | 2.01 ± 0.20 | ||
| 1.49 ± 0.10 | ↓ | 1.46 ± 0.95 | ↑ | 0.60 ± 0.02 | ↓ | ||||||||||
| Cu | 0.37 | 0–16.6 | 3.63 ± 0.06 | 2.79 ± 0.15 | 16.60 ± 0.25 | 1.22 ± 0.01 | * | <LOQ | * | 1.67 ± 0.02 | 0 | 0 | 0 | 1.11 ± 0.2 | |
| 4.44 ± 0.30 | ↑ | 0.42 ± 0.10 | ↓ | 0.91 ± 0.10 | ↑ | ||||||||||
| Fe | 0.098 | 52–2217 | 946 ± 10 | 258 ± 17 | 2217 ± 123 | 93 ± 4 | * | 86 ± 1 | * | 273 ± 18 | 325 ± 29 | 385 ± 30 | 216 ± 70 | 202 ± 6 | |
| 1007 ± 43 | ↑ | 52 ± 4 | ↓ | 128 ± 2 | ↑ | ||||||||||
| Mg | 1.7 | 7518–11,571 | 9793 ± 40 | * | 8985 ± 120 | 11,474 ± 338 | 8277 ± 34 | * | 7634 ± 61 | * | 8148 ± 112 | 10,152 ± 25 | 9535 ± 80 | 8442 ± 55 | 9871 ± 58 |
| 11,571 ± 166 | ↑ | 7518 ± 9 | ↓ | 9821 ± 258 | ↑ | ||||||||||
| Mn | 0.058 | 58–176 | 125 ± 2 | * | 118 ± 8 | 88 ± 3 | 84 ± 1 | * | 81 ± 1 | * | 68 ± 2 | 91 ± 3 | 156 ± 4 | 58 ± 3 | 142 ± 9 |
| 142 ± 3 | ↑ | 70 ± 3 | ↓ | 75 ± 2 | ↓ | ||||||||||
| Rb | 0.55 | 5–29 | 24 ± 0.1 | * | 22 ± 1.0 | 20 ± 0.3 | 19 ± 0.2 | * | 4.7 ± 0.4 | * | 20 ± 0.4 | 25 ± 1.0 | 22 ± 0.6 | 29 ± 0.3 | 29 ± 1.0 |
| 29 ± 0.8 | ↑ | 25 ± 0.9 | ↑ | 19 ± 1.3 | ↑ | ||||||||||
| Sr | 0.026 | 802–1365 | 1247 ± 14 | * | 1365 ± 29 | 1048 ± 11 | 1002 ± 18 | * | 909 ± 13 | * | 804 ± 5 | 714 ± 8 | 862 ± 11 | 828 ± 19 | 912 ± 12 |
| 1137 ± 19 | ↓ | 830 ± 3 | ↓ | 802 ± 40 | ↓ | ||||||||||
| Zn | 0.17 | 14–107 | 61 ± 1.0 | * | 36 ± 2.7 | 107 ± 4.3 | 35 ± 1.3 | * | 34.0 ± 1.0 | * | 35.2 ± 1.1 | 14.6 ± 0.6 | 16.6 ± 0.8 | 14.4 ± 0.3 | 15.2 ± 0.4 |
| 48 ± 1.2 | ↓ | 41 ± 1.6 | ↑ | 25.4 ± 2.1 | ↓ | ||||||||||
| Ash, | - | 19–28 | 23.3 ± 0.2 | 20.9 ± 0.3 | 25.1 ± 2.1 | 20.2 ± 5.1 | 21.2 ± 2.5 | 28.2 ± 0.4 | 19.1 ± 0.9 | 20.1 ± 0.3 | 20.1 ± 1.0 | 20.3 ± 0.8 | |||
| % DW | 21.9 ± 0.3 | ↓ | 18.5 ± 0.5 | ↑ | 28.5 ± 0.3 | ↑ | |||||||||
| Elements | Al | As | Ba | Ca | Co | Cu | Fe | Mg | Mn | Rb | Sr | Zn |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Al | 1 | |||||||||||
| As | −0.375 | 1 | ||||||||||
| Ba | −0.242 | −0.650 | 1 | |||||||||
| Ca | 0.508 | −0.222 | −0.183 | 1 | ||||||||
| Co | 0.517 | −0.348 | 0.074 | 0.315 | 1 | |||||||
| Cu | 0.789 | −0.349 | −0.202 | 0.315 | 0.538 | 1 | ||||||
| Fe | 0.889 | −0.420 | −0.114 | 0.431 | 0.482 | 0.947 | 1 | |||||
| Mg | 0.793 | −0.685 | 0.204 | 0.282 | 0.338 | 0.618 | 0.741 | 1 | ||||
| Mn | 0.361 | −0.583 | 0.532 | 0.151 | 0.495 | 0.043 | 0.197 | 0.519 | 1 | |||
| Rb | 0.201 | −0.701 | 0.394 | 0.216 | 0.110 | −0.018 | 0.088 | 0.402 | 0.307 | 1 | ||
| Sr | 0.559 | −0.293 | −0.200 | 0.577 | 0.586 | 0.383 | 0.380 | 0.270 | 0.424 | 0.066 | 1 | |
| Zn | 0.775 | −0.073 | −0.459 | 0.414 | 0.578 | 0.922 | 0.877 | 0.423 | −0.038 | −0.169 | 0.466 | 1 |
| Element | Sampling Site with a Maximum Concentration | Reproductive Phase | Maximum Concentration | Daily Dose for 3.3 g Consumption | Daily Dose for 12.5 g Consumption | Daily Dose from Risk Estimators | Daily Nutritional Requirements |
|---|---|---|---|---|---|---|---|
| Al | IS, St. 1 | Fertile | 724 | 2.39 | 9.03 | 10 1 | |
| As total | BS, St. 4 | Sterile | 58 | 0.19 | 0.72 | 0.15 1 (inorganic) | |
| Ca | IS, St. 1 | Sterile | 30,093 | 99 | 375 | 2500 2 | 1000 3 |
| Co | WS, St. 9 | Fertile | 2.0 | 0.007 | 0.025 | 30 5 | 10 5 |
| Cu | IS, St. 2 | Sterile | 16.6 | 0.05 | 0.21 | 5 2,5 | 0.9 4/1.0 5 |
| Fe | IS, St. 2 | Sterile | 2217 | 7.3 | 27.7 | 45 5 | 10 3,5 |
| Mg | IS, St. 1 | Fertile | 11,571 | 38 | 144 | 800 5 | 400 5 |
| Mn | WS, St. 7 | Fertile | 156 | 0.52 | 1.95 | 11 5 | 2.7 3/2.0 5 |
| Zn | IS, St. 2 | Sterile | 107 | 0.35 | 1.33 | 25 2 | 12 3,5 |
| Sea Area | Sampling Site | Coordinates | Reproductive Phase | Station (No. on the Map; Figure 7) | Mean Water Temperature, °C | Range of Salinity, ‰ |
|---|---|---|---|---|---|---|
| Irminger Sea | Fossvogur Bay | 64.120887 N 21.930663 W | Fertile | 1 | 13.9 | 29.8 ± 0.3 |
| Irminger Sea | Fossvogur Bay | 64.120978 N 21.929122 W | Sterile | 1a | 4.0 | 34.9 ± 0.1 |
| Irminger Sea | Fossvogur Bay | 64.120887 N 21.930663 W | Sterile | 1 | 4.0 | 35.2 ± 0.3 |
| Irminger Sea | Seltjarnarnes Peninsula | 64.15035 N 21.97255 W | Sterile | 2 | 3.9 | 34.5 ± 0.6 |
| Norwegian Sea | Sudspissen Cape | 69.627168 N 18.912621 E | Fertile | 3 | 9.1 | 33.1 ± 0.3 |
| Norwegian Sea | Sudspissen Cape | 69.627168 N 18.912621 E | Sterile | 3 | 6.3 | 35.0 ± 0.2 |
| Barents Sea | Teriberskaya Bay | 69.184068 N 35.259487 E | Fertile | 4 | 9.1 | 20.3 ± 0.4 |
| Barents Sea | Teriberskaya Bay | 69.184068 N 35.259487 E | Sterile | 4 | 4.2 | 25.4 ± 0.6 |
| Barents Sea | Teriberskaya Bay | 69.173088 N 35.168468 E | Fertile | 4 | 11.2 | 15.1 ± 0.4 |
| Barents Sea | Zelenetskaya Bay | 69.117150 N 36.070790 E | Fertile | 5 | 10.3 | 31.5 ± 0.5 |
| White Sea | Bolshoy Gorely island | 66.31376 N 33.612736 E | Fertile | 6 | 12.8 | 22.8 ± 0.7 |
| White Sea | Matrenin Island | 66.30945 N 33.631920 E | Fertile | 7 | 14.1 | 15.3 ± 0.2 |
| White Sea | Malyy Andronin Island | 66.333374 N 33.766743 E | Fertile | 8 | 13.0 | 25.9 ± 0.4 |
| White Sea | Pezhostrov Island | 66.273315 N 33.934406 E | Fertile | 9 | 17.2 | 22.2 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Obluchinskaya, E.D.; Pozharitskaya, O.N.; Zakharov, D.V.; Flisyuk, E.V.; Terninko, I.I.; Generalova, Y.E.; Smekhova, I.E.; Shikov, A.N. The Biochemical Composition and Antioxidant Properties of Fucus vesiculosus from the Arctic Region. Mar. Drugs 2022, 20, 193. https://doi.org/10.3390/md20030193
Obluchinskaya ED, Pozharitskaya ON, Zakharov DV, Flisyuk EV, Terninko II, Generalova YE, Smekhova IE, Shikov AN. The Biochemical Composition and Antioxidant Properties of Fucus vesiculosus from the Arctic Region. Marine Drugs. 2022; 20(3):193. https://doi.org/10.3390/md20030193
Chicago/Turabian StyleObluchinskaya, Ekaterina D., Olga N. Pozharitskaya, Denis V. Zakharov, Elena V. Flisyuk, Inna I. Terninko, Yulia E. Generalova, Irina E. Smekhova, and Alexander N. Shikov. 2022. "The Biochemical Composition and Antioxidant Properties of Fucus vesiculosus from the Arctic Region" Marine Drugs 20, no. 3: 193. https://doi.org/10.3390/md20030193
APA StyleObluchinskaya, E. D., Pozharitskaya, O. N., Zakharov, D. V., Flisyuk, E. V., Terninko, I. I., Generalova, Y. E., Smekhova, I. E., & Shikov, A. N. (2022). The Biochemical Composition and Antioxidant Properties of Fucus vesiculosus from the Arctic Region. Marine Drugs, 20(3), 193. https://doi.org/10.3390/md20030193