
Deep-Sea Coral Garden Invertebrates and Their Associated Fungi Are Genetic Resources for Chronic Disease Drug Discovery

 , ,
, ,  , ,
, ,
Abstract
1. Introduction
2. Results
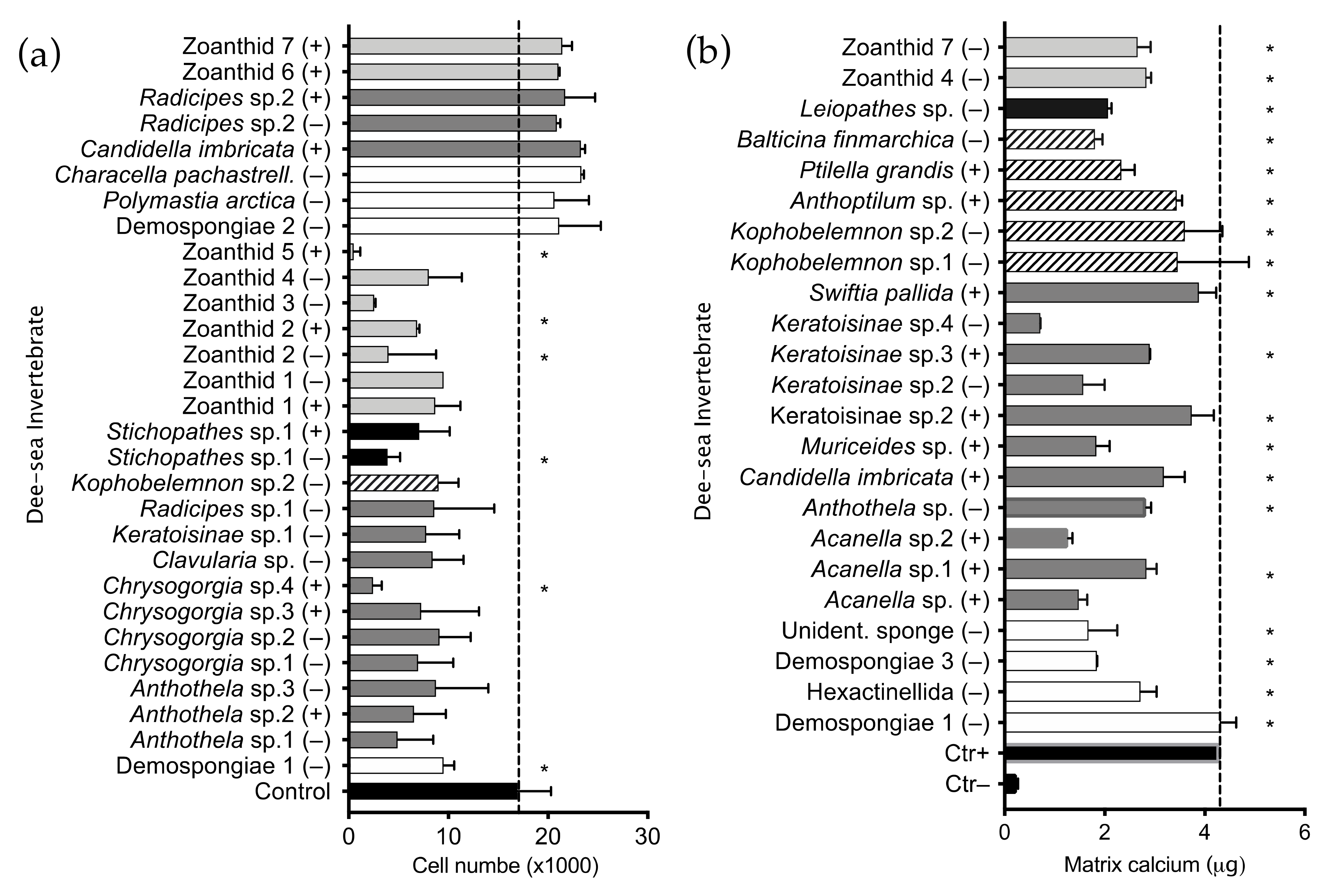
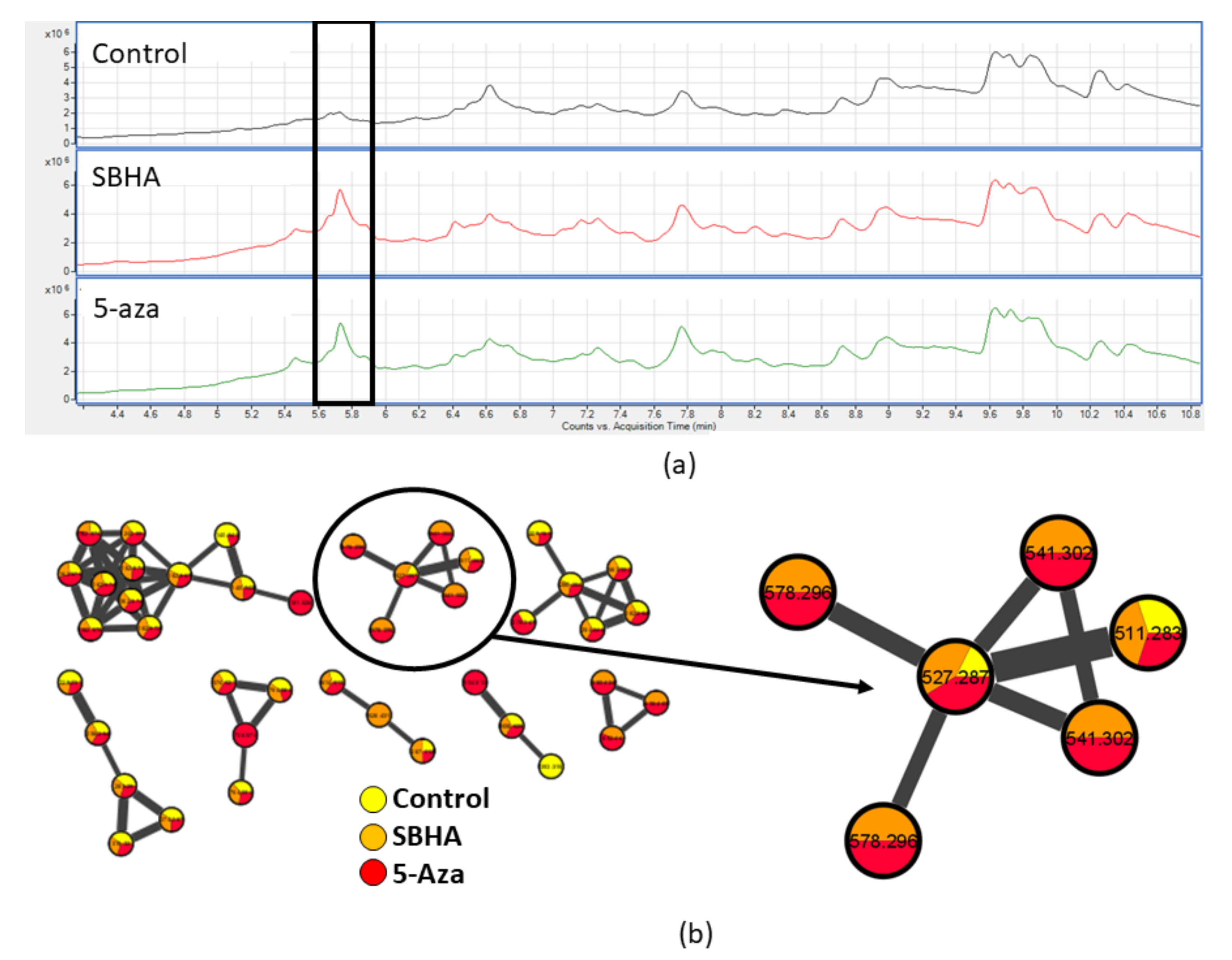
2.1. Invertebrate Extract Bioactivity toward hMSCs
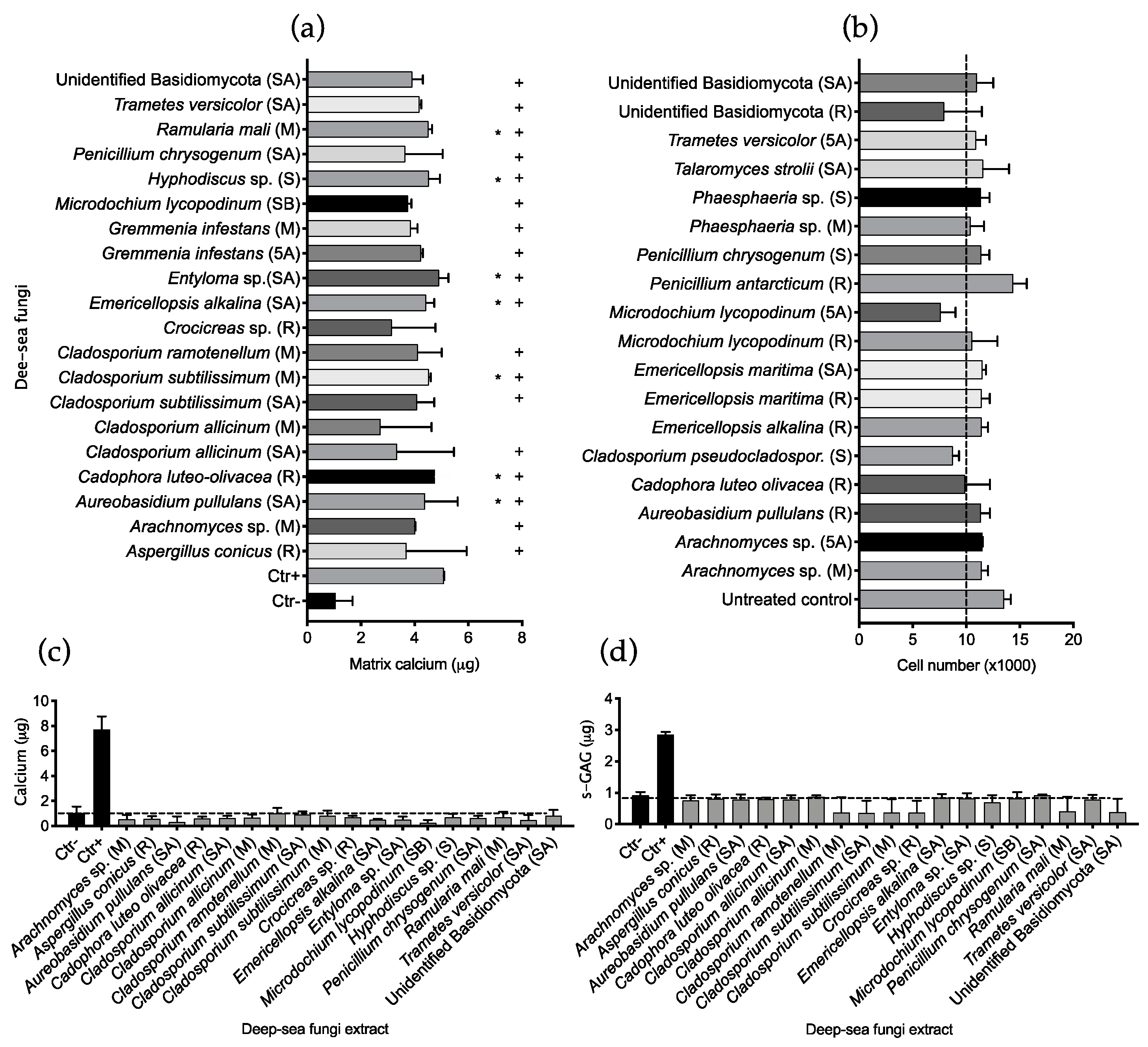
2.2. Fungal Extract Bioactivity toward hMSCs
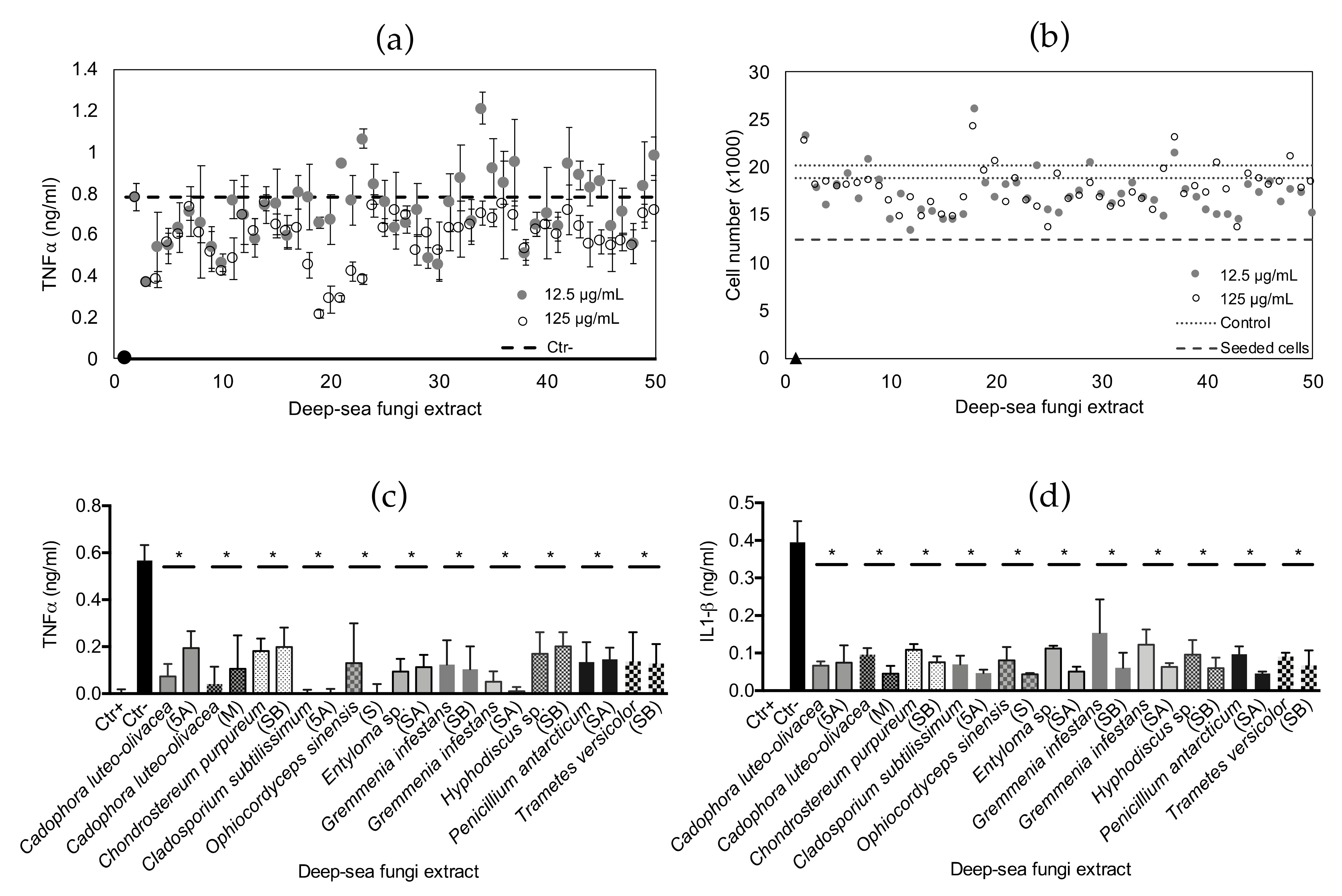
2.3. Anti-Inflammatory Bioactivity
3. Discussion
4. Materials and Methods
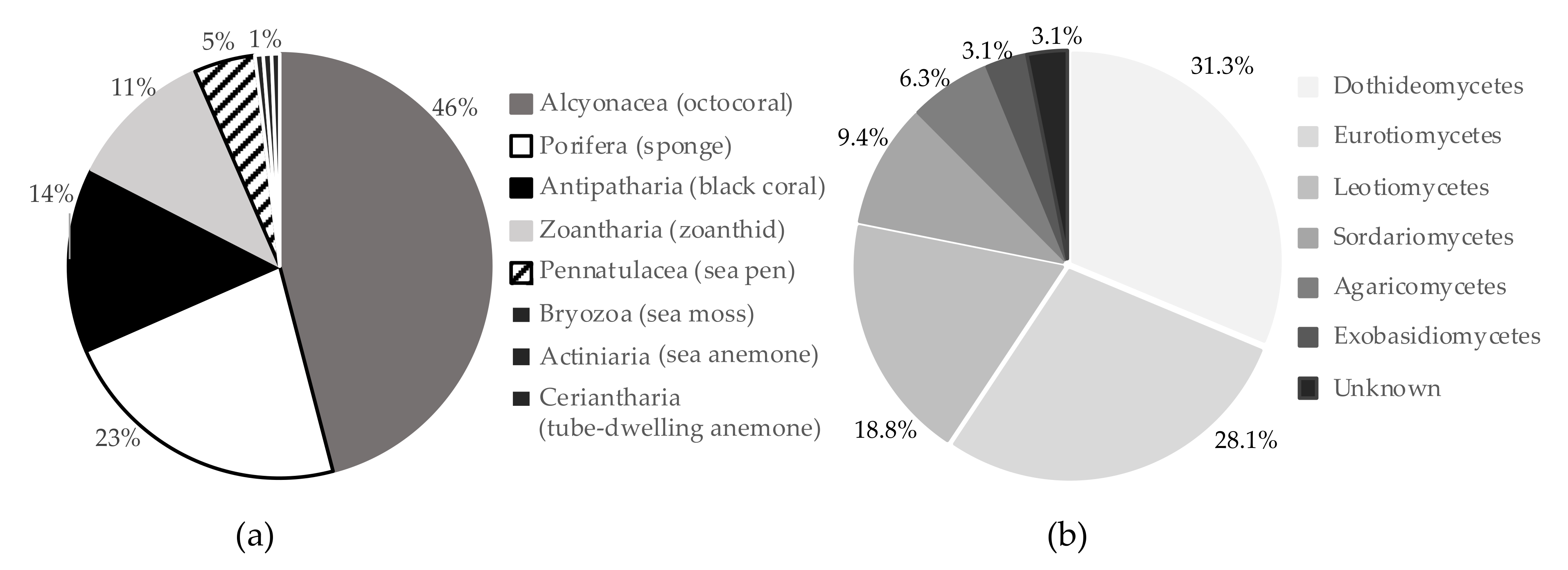
4.1. Deep-Sea Organism Collection
4.2. Deep-Sea Benthic Invertebrate Metabolite Extraction
4.3. Deep-Sea Fungal Culture and Metabolite Extraction
4.4. Culture of Human Mesenchymal Stem Cells and THP1 Macrophages
4.5. Marine Extract Library Screening
4.6. High-Throughput Osteogenic Assay
4.7. Chondrogenic Assay
4.8. High-Throughput Anti-Inflammatory Assay
4.9. LC-MS Analysis of Extracts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Barry, F.; Boynton, R.E.; Liu, B.; Murphy, M. Chondrogenic Differentiation of Mesenchymal Stem Cells from Bone Marrow: Differentiation-Dependent Gene Expression of Matrix Components. Exp. Cell Res. 2001, 268, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Sacchetti, B.; Funari, A.; Michienzi, S.; Di Cesare, S.; Piersanti, S.; Saggio, I.; Tagliafico, E.; Ferrari, S.; Robey, P.; Riminucci, M.; et al. Self-Renewing Osteoprogenitors in Bone Marrow Sinusoids Can Organize a Hematopoietic Microenvironment. Cell 2007, 131, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Friendenstein, A.J.; Chailakhyan, R.K.; Gerasimov, U.V. Bone marrow osteogenic stem cells. In Calcium Regulation and Bone Metabolism. Basic and Clinical Aspects; Cohn, D.V., Glorieux, F.H., Martin, T.J., Eds.; Excerpta Medica: Amsterdam, The Netherlands, 1990; Volume 10, pp. 19–29. [Google Scholar]
- Somoza, R.; Welter, J.F.; Correa, D.; Caplan, A.I. Chondrogenic Differentiation of Mesenchymal Stem Cells: Challenges and Unfulfilled Expectations. Tissue Eng. Part B Rev. 2014, 20, 596–608. [Google Scholar] [CrossRef] [PubMed]
- Clarke, S.; Walsh, P.; Maggs, C.; Buchanan, F. Designs from the deep: Marine organisms for bone tissue engineering. Biotechnol. Adv. 2011, 29, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, N.; Haynesworth, S.E.; Caplan, A.I.; Bruder, S.P. Osteogenic differentiation of purified, culture-expanded human mesenchymal stem cells in vitro. J. Cell. Biochem. 1997, 64, 295–312. [Google Scholar] [CrossRef]
- Friedman, M.S.; Long, M.W.; Hankenson, K.D. Osteogenic differentiation of human mesenchymal stem cells is regulated by bone morphogenetic protein-6. J. Cell. Biochem. 2006, 98, 538–554. [Google Scholar] [CrossRef] [PubMed]
- Fiorentini, E.; Granchi, D.; Leonardi, E.; Baldini, N.; Ciapetti, G. Effects of osteogenic differentiation inducers on in vitro ex-panded adult mesenchymal stromal cells. Int. J. Artif. Organs 2011, 34, 998–1011. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, B.; Hering, T.M.; Caplan, A.; Goldberg, V.M.; Yoo, J.U. In Vitro Chondrogenesis of Bone Marrow-Derived Mesenchymal Progenitor Cells. Exp. Cell Res. 1998, 238, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Crecente-Campo, J.; Borrajo, E.; Vidal, A.; Garcia-Fuentes, M. New scaffolds encapsulating TGF-beta3/BMP-7 combinations driving strong chondrogenic differentiation. Eur. J. Pharm. Biopharm. 2017, 114, 69–78. [Google Scholar] [CrossRef]
- Friedlaender, G.E.; Perry, C.R.; Cole, J.D.; Cook, S.D.; Cierny, G.; Muschler, G.F.; Zych, G.A.; Calhoun, J.H.; LaForte, A.J.; Yin, S. Osteogenic Protein-1 (Bone Morphogenetic Protein-7) in the Treatment of Tibial Nonunions: A Prospective, Random-ized Clinical Trial Comparing rhOP-1 with Fresh Bone Autograft. J. Bone Jt. Surg. 2001, 83-A, 151–158. [Google Scholar] [CrossRef]
- Govender, S.; Csimma, C.; Genant, H.K.; Valentin-Opran, A.; Amit, Y.; Arbel, R.; Aro, H.; Atar, D.; Bishay, M.; Börner, M.G.; et al. Recombinant human bone morphogenetic protein-2 for treatment of open tibial fractures: A prospective, controlled, randomized study of four hundred and fifty patients. J. Bone Jt. Surg. 2002, 84, 2123–2134. [Google Scholar] [CrossRef]
- Blaney Davidson, E.N.; Vitters, E.L.; van Beuningen, H.M.; van de Loo, F.A.; van den Berg, W.B.; van der Kraan, P.M. Re-semblance of osteophytes in experimental osteoarthritis to transforming growth factor beta-induced osteophytes: Limited role of bone morphogenetic protein in early osteoarthritic osteophyte formation. Arthritis Rheum. 2007, 56, 4065–4073. [Google Scholar] [CrossRef]
- Walsh, S.; Jordan, G.R.; Jefferiss, C.; Stewart, K.; Beresford, J.N. High concentrations of dexamethasone suppress the prolifer-ation but not the differentiation or further maturation of human osteoblast precursors in vitro: Relevance to glucocorti-coid-induced osteoporosis. Rheumatology 2001, 40, 70–83. [Google Scholar] [CrossRef]
- Goldring, S.R.; Goldring, M. The Role of Cytokines in Cartilage Matrix Degeneration in Osteoarthritis. Clin. Orthop. Relat. Res. 2004, 427, S27–S36. [Google Scholar] [CrossRef]
- Blom, A.B.; Van Der Kraan, P.M.; Van Den Berg, W.B. Cytokine Targeting in Osteoarthritis. Curr. Drug Targets 2007, 8, 283–292. [Google Scholar] [CrossRef]
- Doherty, T.M.; Asotra, K.; Fitzpatrick, L.A.; Qiao, J.-H.; Wilkin, D.J.; Detrano, R.C.; Dunstan, C.; Shah, P.K.; Rajavashisth, T.B. Calcification in atherosclerosis: Bone biology and chronic inflammation at the arterial crossroads. Proc. Natl. Acad. Sci. USA 2003, 100, 11201–11206. [Google Scholar] [CrossRef]
- Leszczynska, A.; O’Doherty, A.; Farrell, E.; Pindjakova, J.; O’Brien, F.J.; O’Brien, T.; Barry, F.; Murphy, J.M. Differentiation of Vascular Stem Cells Contributes to Ectopic Calcification of Atherosclerotic Plaque. Stem Cells 2016, 34, 913–923. [Google Scholar] [CrossRef]
- Goldring, M.B. Anticytokine therapy for osteoarthritis. Expert Opin. Biol. Ther. 2001, 1, 817–829. [Google Scholar] [CrossRef]
- Rothschild, L.J.; Mancinelli, R.L. Life in extreme environments. Nat. Cell Biol. 2001, 409, 1092–1101. [Google Scholar] [CrossRef]
- Wright, P.C.; Westacott, R.E.; Burja, A.M. Piezotolerance as a metabolic engineering tool for the biosynthesis of natural products. Biomol. Eng. 2003, 20, 325–331. [Google Scholar] [CrossRef]
- Skropeta, D. Deep-sea natural products. Nat. Prod. Rep. 2008, 25, 1131–1166. [Google Scholar] [CrossRef]
- Skropeta, D.; Wei, L. Recent advances in deep-sea natural products. Nat. Prod. Rep. 2014, 31, 999–1025. [Google Scholar] [CrossRef]
- Schupp, P.J.; Kohlert-Schupp, C.; Whitefield, S.; Engemann, A.; Rohde, S.; Hemscheidt, T.; Pezzuto, J.M.; Kondratyuk, T.P.; Park, E.J.; Marler, L.; et al. Cancer Chemopreventive and Anticancer Evaluation of Extracts and Fractions from Marine Macro-and Micro-organisms Collected from Twilight Zone Waters Around Guam. Nat. Prod. Commun. 2009, 4, 1717. [Google Scholar]
- Dumdei, E.J.; Blunt, J.W.; Munro, M.H.G.; Pannell, L.K. Isolation of Calyculins, Calyculinamides, and Swinholide H from the New Zealand Deep-Water Marine Sponge Lamellomorpha strongylata. J. Org. Chem. 1997, 62, 2636–2639. [Google Scholar] [CrossRef]
- Pilkington, L.I. A Chemometric Analysis of Deep-Sea Natural Products. Molecules 2019, 24, 3942. [Google Scholar] [CrossRef]
- Thao, N.P.; Luyen, B.T.T.; Lee, S.H.; Jang, H.D.; Kiem, P.V.; Minh, C.V.; Kim, Y.H. Antiosteoporotic and antioxidant activi-ties of diterpenoids from the Vietnamese soft corals Sinularia maxima and Lobophytum crassum. Med. Chem. Res. 2015, 24, 3551–3560. [Google Scholar] [CrossRef]
- Allemand, D.; Tambutté, É.; Zoccola, D.; Tambutté, S. Coral Calcification, Cells to Reefs. In Coral Reefs: An Ecosystem in Transition; Springer: Berlin/Heidelberg, Germany, 2011; pp. 119–150. [Google Scholar]
- Gentili, C.; Cancedda, R. Cartilage and Bone Extracellular Matrix. Curr. Pharm. Design 2009, 15, 1334–1348. [Google Scholar] [CrossRef]
- Zoccola, D.; Moya, A.; Beranger, G.E.; Tambutte, E.; Allemand, D.; Carle, G.F.; Tambutte, S. Specific expression of BMP2/4 ortholog in biomineralizing tissues of corals and action on mouse BMP receptor. Mar. Biotechnol. 2009, 11, 260–269. [Google Scholar] [CrossRef]
- Kim, H.K.; Cho, S.G.; Kim, J.H.; Doan, T.K.P.; Hu, Q.S.; Ulhaq, R.; Song, E.K.; Yoon, T.R. Mevinolin enhances osteogenic genes (ALP, type I collagen and osteocalcin), CD44, CD47 and CD51 expression during osteogenic differentiation. Life Sci. 2009, 84, 290–295. [Google Scholar] [CrossRef]
- Xu, J.; Yi, M.; Ding, L.; He, S. A Review of Anti-Inflammatory Compounds from Marine Fungi, 2000–2018. Mar. Drugs 2019, 17, 636. [Google Scholar] [CrossRef]
- Marchese, P.; Mahajan, N.; O’Connell, E.; Fearnhead, H.; Tuohy, M.; Krawczyk, J.; Thomas, O.P.; Barry, F.; Murphy, M.J. A Novel High-Throughput Screening Platform Identifies Itaconate Derivatives from Marine Penicillium antarcticum as Inhibi-tors of Mesenchymal Stem Cell Differentiation. Mar. Drugs 2020, 18, 192. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Llodra, E.; Brandt, A.; Danovaro, R.; De Mol, B.; Escobar, E.; German, C.R.; Levin, L.A.; Martinez Arbizu, P.; Me-not, L.; Buhl-Mortensen, P.; et al. Deep, diverse and definitely different: Unique attributes of the world’s largest ecosystem. Biogeosciences 2010, 7, 2851–2899. [Google Scholar] [CrossRef]
- Harden-Davies, H. Deep-sea genetic resources: New frontiers for science and stewardship in areas beyond national jurisdic-tion. Deep Sea Res. Part II Top. Stud. Oceanogr. 2017, 137, 504–513. [Google Scholar] [CrossRef]
- Macreadie, P.I.; McLean, D.; Thomson, P.G.; Partridge, J.C.; Jones, D.O.; Gates, A.; Benfield, M.C.; Collin, S.P.; Booth, D.; Smith, L.L.; et al. Eyes in the sea: Unlocking the mysteries of the ocean using industrial, remotely operated vehicles (ROVs). Sci. Total. Environ. 2018, 634, 1077–1091. [Google Scholar] [CrossRef]
- Bullimore, R.D.; Foster, N.L.; Howell, K.L. Coral-characterized benthic assemblages of the deep Northeast Atlantic: Defining “Coral Gardens” to support future habitat mapping efforts. ICES J. Mar. Sci. 2013, 70, 511–522. [Google Scholar] [CrossRef]
- Marchese, P.; Garzoli, L.; Young, R.; Allcock, L.; Barry, F.; Tuohy, M.; Murphy, M. Fungi populate deep-sea coral gardens as well as marine sediments in the Irish Atlantic Ocean. Environ. Microbiol. 2021. [Google Scholar] [CrossRef]
- Mancuso, P.; Raman, S.; Glynn, A.; Barry, F.; Murphy, J.M. Mesenchymal Stem Cell Therapy for Osteoarthritis: The Critical Role of the Cell Secretome. Front. Bioeng. Biotechnol. 2019, 7, 9. [Google Scholar] [CrossRef]
- Demers, D.H.; Knestrick, M.A.; Fleeman, R.; Tawfik, R.; Azhari, A.; Souza, A.; Vesely, B.; Netherton, M.; Gupta, R.; Colon, B.; et al. Exploitation of Mangrove Endophytic Fungi for Infectious Disease Drug Discovery. Mar. Drugs 2018, 16, 376. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzat-to-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecu-lar Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. ChemBioChem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Brey, D.M.; Motlekar, N.A.; Diamond, S.L.; Mauck, R.L.; Garino, J.P.; Burdick, J.A. High-throughput screening of a small molecule library for promoters and inhibitors of mesenchymal stem cell osteogenic differentiation. Biotechnol. Bioeng. 2011, 108, 163–174. [Google Scholar] [CrossRef]
- Alves, H.; Dechering, K.; Van Blitterswijk, C.; De Boer, J. High-throughput assay for the identification of compounds regu-lating osteogenic differentiation of human mesenchymal stromal cells. PLoS ONE 2011, 6, e26678. [Google Scholar] [CrossRef]
- Roberts, J.M.; Murray, F.; Anagnostou, E.; Hennige, S.; Gori, A.; Henry, L.-A.; Fox, A.; Kamenos, N.; Foster, G.L. Cold-Water Corals in an Era of Rapid Global Change: Are These the Deep Ocean’s Most Vulnerable Ecosystems? In The Cnidaria, Past, Present and Future; Springer: Berlin/Heidelberg, Germany, 2016; pp. 593–606. [Google Scholar]
- Fernandez-Arcaya, U.; Ramirez-Llodra, E.; Aguzzi, J.; Allcock, A.L.; Davies, J.S.; Dissanayake, A.; Harris, P.; Howell, K.; Huvenne, V.A.I.; Macmillan-Lawler, M.; et al. Ecological Role of Submarine Canyons and Need for Canyon Conservation: A Review. Front. Mar. Sci. 2017, 4, 4. [Google Scholar] [CrossRef]
- Daly, E.; Johnson, M.P.; Wilson, A.M.; Gerritsen, H.D.; Kiriakoulakis, K.; Allcock, A.L.; White, M. Bottom trawling at Whit-tard Canyon: Evidence for seabed modification, trawl plumes and food source heterogeneity. Prog. Oceanogr. 2018, 169, 227–240. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Polarity | Bioactivity | ||||
|---|---|---|---|---|---|---|
| Pro-Osteogenic | Pro-Chondrogenic | Proliferative | Anti-prolif. | Cytotoxic | ||
| Acanella sp.1 | + | x | ||||
| Anthoptilum sp. | + | x | ||||
| Anthothela sp. | − | x | ||||
| Anthothela sp.1 | − | x | ||||
| Anthothela sp.2 | + | x | ||||
| Anthothela sp.3 | − | x | ||||
| Balticina finmarchica | − | x | ||||
| Candidella imbricata | + | x | ||||
| Characella pachastrell. | − | x | ||||
| Chrysogorgia sp.1 | − | x | ||||
| Chrysogorgia sp.2 | − | x | ||||
| Chrysogorgia sp.3 | + | x | ||||
| Chrysogorgia sp.4 | + | x | ||||
| Clavularia sp. | − | x | ||||
| Demospongiae 1 | − | x | ||||
| Demospongiae 2 | − | x | ||||
| Hexactinellida | − | x | ||||
| Keratoisinae sp.1 | − | x | ||||
| Keratoisinae sp.2 | + | x | ||||
| Keratoisinae sp.3 | + | x | ||||
| Kophobelemnon sp.1 | − | x | ||||
| Kophobelemnon sp.2 | − | x | x | |||
| Leiopathes sp. | − | x | ||||
| Muriceides sp. | + | x | ||||
| Polymastia arctica | − | x | ||||
| Radicipes sp.1 | − | x | ||||
| Radicipes sp.2 | − | x | ||||
| Radicipes sp.2 | + | x | ||||
| Stichopathes sp.1 | − | x | ||||
| Stichopathes sp.1 | + | x | ||||
| Swiftia pallida | + | x | ||||
| Unidentif. sponge | − | x | ||||
| Zoanthid 1 | − | x | ||||
| Zoanthid 1 | + | x | ||||
| Zoanthid 2 | − | x | ||||
| Zoanthid 2 | + | x | ||||
| Zoanthid 3 | − | x | x | |||
| Zoanthid 4 | − | x | x | |||
| Zoanthid 5 | + | x | ||||
| Zoanthid 6 | + | x | ||||
| Zoanthid 7 | − | x | x | |||
| Zoanthid 7 | + | x | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marchese, P.; Young, R.; O’Connell, E.; Afoullouss, S.; Baker, B.J.; Allcock, A.L.; Barry, F.; Murphy, J.M. Deep-Sea Coral Garden Invertebrates and Their Associated Fungi Are Genetic Resources for Chronic Disease Drug Discovery. Mar. Drugs 2021, 19, 390. https://doi.org/10.3390/md19070390
Marchese P, Young R, O’Connell E, Afoullouss S, Baker BJ, Allcock AL, Barry F, Murphy JM. Deep-Sea Coral Garden Invertebrates and Their Associated Fungi Are Genetic Resources for Chronic Disease Drug Discovery. Marine Drugs. 2021; 19(7):390. https://doi.org/10.3390/md19070390
Chicago/Turabian StyleMarchese, Pietro, Ryan Young, Enda O’Connell, Sam Afoullouss, Bill J. Baker, A. Louise Allcock, Frank Barry, and J. Mary Murphy. 2021. "Deep-Sea Coral Garden Invertebrates and Their Associated Fungi Are Genetic Resources for Chronic Disease Drug Discovery" Marine Drugs 19, no. 7: 390. https://doi.org/10.3390/md19070390
APA StyleMarchese, P., Young, R., O’Connell, E., Afoullouss, S., Baker, B. J., Allcock, A. L., Barry, F., & Murphy, J. M. (2021). Deep-Sea Coral Garden Invertebrates and Their Associated Fungi Are Genetic Resources for Chronic Disease Drug Discovery. Marine Drugs, 19(7), 390. https://doi.org/10.3390/md19070390