The Effects of Marine Algal Polyphenols, Phlorotannins, on Skeletal Muscle Growth in C2C12 Muscle Cells via Smad and IGF-1 Signaling Pathways





Abstract
1. Introduction
2. Results
2.1. Cytotoxicity of Phlorotannins
2.2. Cell Proliferation Activity of Phlorotannins
2.3. Creatine Kinase (CK) Activity
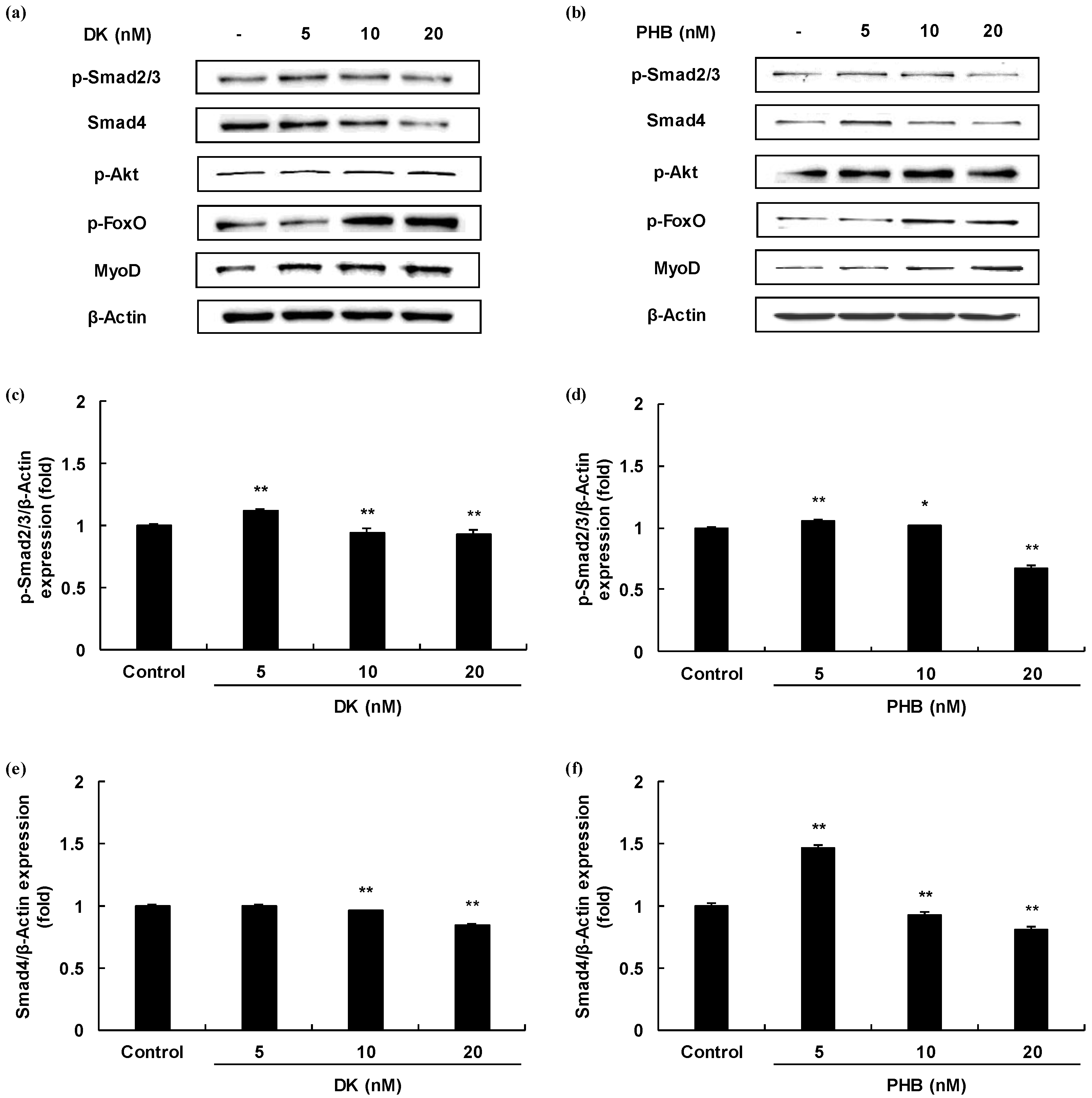
2.4. Effects of Muscle Growth Regulation of DK and/or PHB
2.5. Anaylsis of In Silico Molecular Docking of Phlorotannins with TGF-β (Myostatin)
2.6. Anaylsis of In Silico Molecular Docking of Phlorotannins with IGF-1 Receptor (IGF-1R)
3. Discussion
4. Materials and Methods
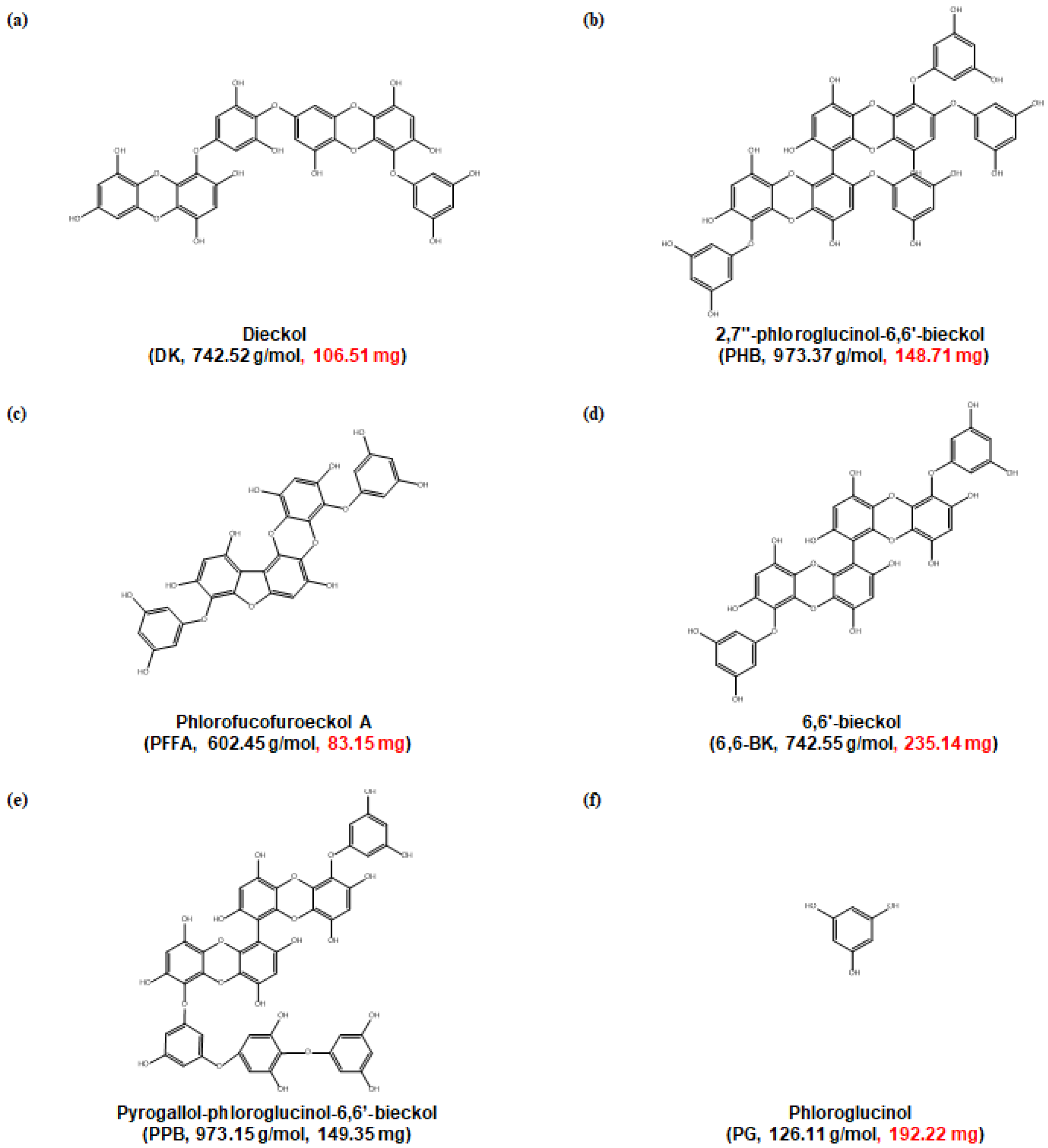
4.1. Preparation of Crude Extract and Phlorotannins from E. cava
4.2. Skeletal Muscle Cell Grwoth Activities of Phlorotannins
4.2.1. Myoblasts Cell Culture and Differentiation to Myocytes
4.2.2. Cytotoxicty of Phlorotannins
4.2.3. Cell Proliferation Activity of Phlorotannins
4.2.4. CK Activity of Phlorotannins
4.3. Effects of Phlorotanins on Skeletal Muscel Cells Differentiation
4.3.1. Western Blot Analysis
4.3.2. In Silico Molecular Docking Simulation
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Elkina, Y.; Heahling, S.; Anker, S.D.; Springer, J. The role of myostatin in muscle wasting: An overview. J. Cachexia Sarcopenia Muscle 2011, 2, 143–151. [Google Scholar] [CrossRef]
- Glass, D.J. Skeletal muscle hypertrophy and atrophy signaling pathways. Int. J. Biochem. Cell Biol. 2005, 37, 1974–1984. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; McPherron, A.C. Regulation of myostatin activity and muscle growth. Proc. Natl. Acad. Sci. USA 2001, 98, 9306–9311. [Google Scholar] [CrossRef]
- Kolias, H.D.; McDermott, J.C. Transforming growth factor-beta and myostatin signaling in skeletal muscle. J. Appl. Physiol. 2008, 104, 579–587. [Google Scholar] [CrossRef]
- Dominique, J.E.; Gérard, C. The myostatin gene: Physiology and pharmacological relevance. Curr. Opin. Pharmacol. 2007, 7, 310–315. [Google Scholar]
- Amthor, H.; Nicholas, G.; Mckinnell, I.; Kemp, C.F.; Sharma, M.; Kambadur, R.; Patel, K. Follistatin complexes Myostatin and antagonizes Myostatin-mediated inhibition of myogenesis. Dev. Biol. 2004, 270, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.J.; Qiu, Y.; Hewick, R.M.; Wolfman, N.M. Regulation of myostatin in vivo by growth and differentiation factor-associated serum protein-1: A novel protein with protease inhibitor and follistatin domains. Mol. Endocrinol. 2003, 17, 1144–1154. [Google Scholar] [CrossRef]
- Cash, J.N.; Rejon, C.A.; McPherron, A.C.; Bernard, D.J.; Thompson, T.B. The structure of myostatin:follistatin 288: Insights intro receptor utilization and heparin binding. EMBO J. 2009, 28, 2662–2676. [Google Scholar] [CrossRef]
- Yoon, M.S.; Chen, J. PLD regulates myoblast differentiation through the mTOR-IGF2 pathway. J. Cell Sci. 2008, 121, 143–151. [Google Scholar] [CrossRef]
- McFarlane, C.; Plummer, E.; Thomas, M.; Hennebry, A.; Ashby, M.; Ling, N. Myostatin induces cachexia by activating the ubiquitin proteolytic system through an NF-kappaB-independent, FoxO-1dependent mechanism. J. Cell. Physiol. 2006, 209, 501–514. [Google Scholar] [CrossRef]
- Kim, K.N.; Kang, M.C.; Kang, N.; Kim, S.Y.; Hyun, C.G.; Roh, S.W.; Ko, E.Y.; Cho, K.; Jung, W.K.; Ahn, G.; et al. 2,4,6-Trihydroxybenzaldehyde, a potential anti-obesity treatment, suppressed adipocyte differentiation in 3T3-L1 cells and fat accumulation induced by high-fat diet in C57B6/6 mice. Environ. Toxicol. Pharmacol. 2015, 39, 962–968. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Jin, D.H.; Kim, Y.D.; Jin, H.J. Effects of Carnosic Acid on Muscle Growth in Zebrafish (Danio rerio). Korean J. Ichthyol. 2014, 26, 171–178. [Google Scholar]
- Shin, H.C.; Kim, S.H.; Park, Y.; Lee, B.H.; Hwang, H.J. Effects of 12-week Oral Supplementation of Ecklonia cava Polyphenols on Anthropometric and Blood Lipid Parameters in Overweight Korean Individuals: A Double-blind Randomized Clinical Trial. Phytother. Res. 2012, 26, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.G.; Kang, O.H.; Brice, O.O.; Lee, Y.S.; Chae, H.S.; Oh, Y.C.; Sohn, D.H.; Park, H.; Choi, H.G.; Kim, S.G.; et al. Antibacterial Activity of Ecklonia cava Against Methicillin-Resistant Staphylococcus aureus and Salmonella spp. Foodborne Pathog. Dis. 2010, 7, 435–441. [Google Scholar] [CrossRef]
- Li, Y.; Qian, Z.J.; Ryu, B.M.; Lee, S.H.; Kim, M.M.; Kim, S.K. Chemical components and its antioxidant properties in vitro: An edible marine brown alga, Ecklonia cava. Bioorg. Med. Chem. 2009, 17, 1963–1973. [Google Scholar] [CrossRef]
- Yang, Y.I.; Shin, H.C.; Kim, S.H.; Park, W.Y.; Lee, K.Y.; Choi, J.H. 6,6′-Bieckol, isolated from marine alga Ecklonia cava, suppressed LPS-induced nitric oxide and PGE2 production and inflammatory cytokine expression in macrophages: The inhibition of NFκB. Int. Immunopharmacol. 2012, 12, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Wijesinghe, W.A.J.P.; Ko, S.C.; Jeon, Y.J. Effect of phlorotannins isolated from Ecklonia cava on angiotensin I-converting enzyme (ACE) inhibitory activity. Nutr. Res. Pract. 2011, 5, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Wijesinghe, W.A.J.P.; Jeon, Y.J. Exploiting biological activities of brown seaweed Ecklonia cava for potential industrial applications: A review. Int. J. Food Sci. Nutr. 2012, 63, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Sanjeewa, K.K.A.; Kim, E.A.; Son, K.T.; Jeon, Y.J. Bioactive properties and potentials cosmeceutical applications of phlorotannins isolated from brown seaweeds: A review. J. Photochem. Photobiol. B-Biol. 2016, 162, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Diel, P.; Baanders, D.; Schlüpmann, K.; Velders, M.; Schwarz, J.P. C2C12 myoblastoma cell differentiation and proliferation stimulated by androgens and associated with a modulation of myostatin and Pax7 expression. J. Mol. Endocrinol. 2008, 40, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Braun, T.; Gautel, M. Transcriptional mechanisms regulating skeletal muscle differentiation, growth and homeostasis. Nat. Rev. Mol. Cell Biol. 2011, 12, 349–361. [Google Scholar] [CrossRef] [PubMed]
- McPherron, A.C.; Lawler, A.M.; Lee, S.J. Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member. Nature 1997, 387, 83–90. [Google Scholar] [CrossRef]
- Bodine, S.C.; Stitt, T.N.; Gonzalez, M.; Kline, W.O.; Stover, G.L.; Bauerlein, R. Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nat. Cell Biol. 2001, 3, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.M.V.; Gonzalez, M.; Poueymirou, W.T.; Kline, W.O.; Na, E.; Zlotchenko, E.; Stitt, T.N.; Economides, A.N.; Yancopoulos, G.D.; Glass, D.J. Condotional Activation of Akt in Adult Skeletal Muscle Induces Rapid Hypertrophy. Mol. Cell. Biol. 2004, 24, 9295–9304. [Google Scholar] [CrossRef]
- Hoyne, P.A.; Elleman, T.C.; Adams, T.E.; Richards, K.M.; Ward, C.W. Properties of an insulin receptor with an IGF-1 receptor loop exchange in the cysteine-rich region. FEBS Lett. 2000, 469, 57–60. [Google Scholar] [CrossRef]
- Jacquemin, V.; Bulter-Browne, G.S.; Furling, D.; Mouly, V. IL-13 mediates the recruitment of reserve cells for fusion during IGF-1-induced hypertrophy of human myotubes. J. Cell Sci. 2007, 120, 670–681. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Park, M.Y.; Kim, H.K.; Park, Y.; Whang, K.Y. Cortisone and dexamethasone inhibit myogenesis by modulating the AKT/mTOR signaling pathway in C2C12. Biosci. Biotechnol. Biochem. 2016, 80, 2093–2099. [Google Scholar] [CrossRef]
- Garrett, T.; McKern, N.M.; Lou, M.; Frenkel, M.J.; Bentley, J.D.; Lovrecz, G.O.; Ellemang, T.C.; Cosgrove, L.J.; Ward, C.W. Crystal structure of the first three domains of the type-1 insulin-like growth factor receptor. Nature 1998, 394, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, Y.; Shi, X.; Kim, S.K. Inhibition of the expression on MMP-2, 9 and morphological changes via human fibrosarcoma cell line by 6,6′-bieckol from marine alga Ecklonia cava. BMB Rep. 2010, 43, 62–68. [Google Scholar] [CrossRef]
- Kang, S.M.; Lee, S.H.; Heo, S.J.; Kim, K.N.; Jeon, Y.J. Evaluation of antioxidant properties of a new compound, pyrogallol-phloroglucinol-6,6′-bieckol isolated from brown algae, Ecklonia cava. Nutr. Res. Pract. 2011, 5, 495–502. [Google Scholar] [CrossRef]
- Lee, J.H.; Ko, J.Y.; Oh, J.Y.; Kim, C.Y.; Lee, H.J.; Kim, J.; Jeon, Y.J. Preparative isolation and purification of phlorotannins from Ecklonia cava using centrifugal partition chromatography by one-step. Food Chem. 2014, 158, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Le, Q.T.; Li, Y.; Qian, Z.J.; Kim, M.M.; Kim, S.K. Inhibitory effects of polyphenols isolated from marine alga Ecklonia cava on histamine release. Process Biochem. 2009, 44, 168–176. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, H.S.; Cho, M.; Jeon, Y.J. Enzymatic Hydrolysates of Hippocampus abdominalis Regulates the Skeletal Muscle Growth in C2C12 Cells and Zebrafish Model. J. Aquat. Food Prod. Technol. 2019, 28, 264–274. [Google Scholar] [CrossRef]
- Ko, J.Y.; Lee, J.H.; Kim, H.S.; Kim, H.H.; Jeon, Y.J. Cell proliferation effect of brown marine algae extracts on Mouse Fibroblast. J. Mar. Biosci. Biotechnol. 2015, 7, 28–34. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Receptor (Binding Site) | Ligand | Binding Energy (kcal/mol) | -CDOCK Interaction Energy (kcal/mol) |
|---|---|---|---|
| Myostatin (3HH2-Active site 1) | DK | −146.06 | 50.77 |
| PHB | −109.90 | 42.44 | |
| DHT | −50.00 | 24.07 | |
| Myostatin (3HH2-Active site 2) | DK | −118.99 | 40.01 |
| PHB | −102.71 | 49.42 | |
| DHT | −48.16 | 18.28 | |
| Myostatin (3HH2-Active site 2) | DK | −126.49 | 47.71 |
| PHB | −143.74 | 57.70 | |
| DHT | −51.51 | 32.07 | |
| Myostatin (3HH2-Active site 2) | DK | −108.64 | 38.27 |
| PHB | −125.90 | 54.60 | |
| DHT | −40.08 | 18.76 |
| Receptor (Binding Site) | Ligand | Binding Energy (kcal/mol) | -CDOCK Interaction Energy (kcal/mol) |
|---|---|---|---|
| IGF-1R (1IGR-Active site) | DK | −118.71 | 50.66 |
| PHB | −98.11 | 60.74 | |
| DHT | −60.25 | 20.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.-Y.; Lee, J.-H.; Kang, N.; Kim, K.-N.; Jeon, Y.-J. The Effects of Marine Algal Polyphenols, Phlorotannins, on Skeletal Muscle Growth in C2C12 Muscle Cells via Smad and IGF-1 Signaling Pathways. Mar. Drugs 2021, 19, 266. https://doi.org/10.3390/md19050266
Kim S-Y, Lee J-H, Kang N, Kim K-N, Jeon Y-J. The Effects of Marine Algal Polyphenols, Phlorotannins, on Skeletal Muscle Growth in C2C12 Muscle Cells via Smad and IGF-1 Signaling Pathways. Marine Drugs. 2021; 19(5):266. https://doi.org/10.3390/md19050266
Chicago/Turabian StyleKim, Seo-Young, Ji-Hyeok Lee, Nalae Kang, Kil-Nam Kim, and You-Jin Jeon. 2021. "The Effects of Marine Algal Polyphenols, Phlorotannins, on Skeletal Muscle Growth in C2C12 Muscle Cells via Smad and IGF-1 Signaling Pathways" Marine Drugs 19, no. 5: 266. https://doi.org/10.3390/md19050266
APA StyleKim, S.-Y., Lee, J.-H., Kang, N., Kim, K.-N., & Jeon, Y.-J. (2021). The Effects of Marine Algal Polyphenols, Phlorotannins, on Skeletal Muscle Growth in C2C12 Muscle Cells via Smad and IGF-1 Signaling Pathways. Marine Drugs, 19(5), 266. https://doi.org/10.3390/md19050266