
Apoptotic Activity of New Oxisterigmatocystin Derivatives from the Marine-Derived Fungus Aspergillus nomius NC06

 ,
,  , ,
, ,  and
and 
Abstract
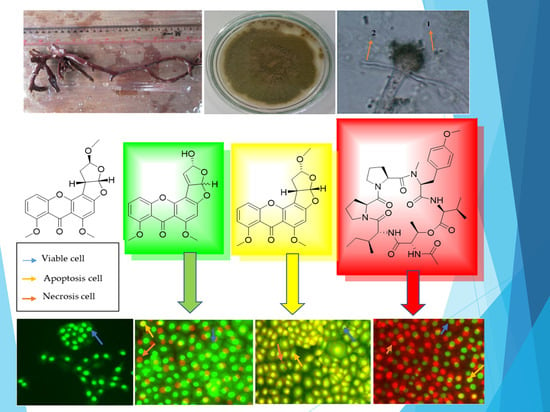
1. Introduction
2. Results and Discussion
3. Conclusions
4. Materials and Methods
4.1. General Procedures
4.2. Fungal Isolation, Identification, and Cultivation
4.3. Extraction and Isolation
4.4. Mosher Ester Analysis of 1
4.5. Cytotoxic Assay
4.6. Antibacterial Activity
4.7. Apoptotic Cells Using AO–PI Double Staining
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, S.; Wang, H.; Su, M.; Hwang, G.J.; Hong, J.; Jung, J.H. New metabolites from the sponge-derived fungus Aspergillus sydowii J05B-7F-4. Nat. Prod. Res. 2017, 31, 1682–1686. [Google Scholar] [CrossRef]
- Pang, X.; Lin, X.; Tian, Y.; Liang, R.; Wang, J.; Yang, B.; Zhou, X.; Kaliyaperumal, K.; Luo, X.; Tu, Z.; et al. Three new polyketides from the marine sponge-derived fungus Trichoderma sp. SCSIO41004. Nat. Prod. Res. 2018, 32, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Du, Y.; Wang, C.; Wang, Y.; Zhu, T.; Zhu, W. Azaphilones from the Marine Sponge-Derived Fungus Penicillium sclerotiorum OUCMDZ-3839. Mar. Drugs 2019, 17, 260. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, J.; Li, L.; Gong, C.; Wang, S.; Yang, F.; Hua, H.; Lin, H. New butenolide derivatives from the marine sponge-derived fungus Aspergillus terreus. Bioorg. Med. Chem. Lett. 2018, 28, 315–318. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Fujii, A.; Kikuchi, T. New Diterpenes with a Fused 6-5-6-6 Ring System Isolated from the Marine Sponge-Derived Fungus Trichoderma harzianum. Mar. Drugs 2019, 17, 480. [Google Scholar] [CrossRef]
- Pang, X.; Cai, G.; Lin, X.; Salendra, L.; Zhou, X.; Yang, B.; Wang, J.; Wang, J.; Xu, S.; Liu, Y. New Alkaloids and Polyketides from the Marine Sponge-Derived Fungus Penicillium sp. SCSIO41015. Mar. Drugs 2019, 17, 398. [Google Scholar] [CrossRef]
- Elissawy, A.M.; Ebada, S.S.; Ashour, M.L.; Özkaya, F.C.; Ebrahim, W.; Singab, A.B.; Proksch, P. Spiroarthrinols a and B, two novel meroterpenoids isolated from the sponge- derived fungus Arthrinium sp. Phytochem. Lett. 2017, 20, 246–251. [Google Scholar] [CrossRef]
- Ma, H.G.; Liu, Q.; Zhu, G.L.; Liu, H.S.; Zhu, W.M. Marine natural products sourced from marine-derived Penicillium fungi. J. Asian Nat. Prod. Res. 2016, 18, 92–115. [Google Scholar] [CrossRef]
- Lei, H.; Lei, J.; Zhou, X.; Hu, M.; Niu, H.; Song, C.; Chen, S.; Liu, Y.; Zhang, D. Cytotoxic Polyketides from the Marine Sponge-Derived Fungus Pestalotiopsis heterocornis XWS03F09. Molecules 2019, 24, 2655. [Google Scholar] [CrossRef]
- Li, Y.; Liu, D.; Cheng, Z.; Proksch, P.; Lin, W. Cytotoxic trichothecene-type sesquiterpenes from the sponge-derived fungus Stachybotrys chartarum with tyrosine kinase inhibition. RSC Adv. 2017, 7, 7259–7267. [Google Scholar] [CrossRef]
- Shah, M.; Sun, C.; Sun, Z.; Zhang, G.; Che, Q.; Gu, Q.; Zhu, T.; Li, D. Antibacterial Polyketides from Antarctica Sponge-Derived Fungus Penicillium sp. HDN151272. Mar. Drugs 2020, 18, 71. [Google Scholar] [CrossRef]
- Zhang, L.; Qiu, P.; Ding, L.; Li, Q.; Song, J.; Han, Z.; He, S. A New Antibacterial Chlorinated Amino Acid Derivative from the Sponge-Derived Fungus Aspergillus sp. LS53. Chem. Nat. Compd. 2020, 56, 109–111. [Google Scholar] [CrossRef]
- Liu, Y.; Ding, L.; Fang, F.; He, S. Penicillilactone A, a novel antibacterial 7-membered lactone derivative from the sponge-associated fungus Penicillium sp. LS54. Nat. Prod. Res. 2019, 33, 2466–2470. [Google Scholar] [CrossRef]
- Artasasta, M.A.; Taher, M.; Djamaan, A.; Handayani, D. Cytotoxic and Antibacterial Activities of Marine Sponge-Derived Fungus Aspergillus Nomius NC06. Rasayan J. Chem. 2019, 12, 1463–1469. [Google Scholar] [CrossRef]
- Artasasta, M.A.; Djamaan, A.; Handayani, D. Cytotoxic activity screening of ethyl acetate fungal extracts derived from the marine sponge Neopetrosia chaliniformis AR-01. J. Appl. Pharm. Sci. 2017, 7, 174–178. [Google Scholar] [CrossRef]
- Lee, Y.M.; Li, H.; Hong, J.; Cho, H.Y.; Bae, K.S.; Kim, M.A.; Kim, D.K.; Jung, J.H. Bioactive metabolites from the sponge-derived fungus Aspergillus versicolor. Arch. Pharm. Res. 2010, 33, 231–235. [Google Scholar] [CrossRef]
- Tian, Y.Q.; Lin, X.P.; Wang, Z.; Zhou, X.F.; Qin, X.C.; Kaliyaperumal, K.; Zhang, T.Y.; Tu, Z.C.; Liu, Y. Asteltoxins with Antiviral Activities from the Marine Sponge-Derived Fungus Aspergillus sp. SCSIO XWS02F40. Molecules 2015, 21, 34. [Google Scholar] [CrossRef]
- Cai, S.; Zhu, T.; Du, L.; Zhao, B.; Li, D.; Gu, Q. Sterigmatocystins from the deep-sea-derived fungus Aspergillus versicolor. J. Antibiot. 2011, 64, 193–196. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, G.-Q.; Tong, X.-P.; Chen, G.-D.; Huang, Y.-F.; Cui, J.-Y.; Kong, M.-Z.; Guo, L.-D.; Zheng, Y.-Z.; Yao, X.-S.; et al. Diphenyl ethers from Aspergillus sp. and their anti-Aβ42 aggregation activities. Fitoterapia 2014, 98, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Rajachan, O.A.; Kanokmedhakul, K.; Soytong, K.; Kanokmedhakul, S. Mycotoxins from the Fungus Botryotrichum piluliferum. J. Agric. Food Chem. 2017, 65, 1337–1341. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.-Q.; Lin, S.-T.; Kumaravel, K.; Zhou, H.; Wang, S.-Y.; Liu, Y.-H. Polyketide-derived metabolites from the sponge-derived fungus Aspergillus sp. F40. Phytochem. Lett. 2018, 27, 74–77. [Google Scholar] [CrossRef]
- Capon, R.J.; Skene, C.; Stewart, M.; Ford, J.; O’Hair, R.A.J.; Williams, L.; Lacey, E.; Gill, J.H.; Heiland, K.; Friedel, T. Aspergillicins A–E: Five novel depsipeptides from the marine-derived fungus Aspergillus carneus. Org. Biomol. Chem. 2003, 1, 1856–1862. [Google Scholar] [CrossRef]
- Gouda, H.; Sunazuka, T.; Ui, H.; Handa, M.; Sakoh, Y.; Iwai, Y.; Hirono, S.; Omura, S. Stereostructure of luminamicin, an anaerobic antibiotic, via molecular dynamics, NMR spectroscopy, and the modified Mosher method. Proc. Natl. Acad. Sci. USA 2005, 102, 18286–18291. [Google Scholar] [CrossRef] [PubMed]
- Ashley, D.L.; Orti, D.L.; Hill, R.H. Proton nuclear magnetic resonance evidence for two configurations of the hemiacetals of aflatoxin B1 and sterigmatocystin. J. Agric. Food Chem. 1987, 35, 782–785. [Google Scholar] [CrossRef]
- Li, J.; Xie, S.; Ahmed, S.; Wang, F.; Gu, Y.; Zhang, C.; Chai, X.; Wu, Y.; Cai, J.; Cheng, G. Antimicrobial Activity and Resistance: Influencing Factors. Front. Pharm. 2017, 8, 364. [Google Scholar] [CrossRef] [PubMed]
- Baharuddin, A.A.; Roosli, R.A.J.; Zakaria, Z.A.; Md Tohid, S.F. Dicranopteris linearis extract inhibits the proliferation of human breast cancer cell line (MDA-MB-231) via induction of S-phase arrest and apoptosis. Pharm. Biol. 2018, 56, 422–432. [Google Scholar] [CrossRef]
- Gerl, R.; Vaux, D.L. Apoptosis in the development and treatment of cancer. Carcinogenesis 2005, 26, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Alabsi, A.M.; Ali, R.; Ali, A.M.; Al-Dubai, S.A.; Harun, H.; Abu Kasim, N.H.; Alsalahi, A. Apoptosis induction, cell cycle arrest and in vitro anticancer activity of gonothalamin in a cancer cell lines. Asian Pac. J. Cancer Prev. 2012, 13, 5131–5136. [Google Scholar] [CrossRef]
- Yeh, C.T.; Rao, Y.K.; Yao, C.J.; Yeh, C.F.; Li, C.H.; Chuang, S.E.; Luong, J.H.; Lai, G.M.; Tzeng, Y.M. Cytotoxic triterpenes from Antrodia camphorata and their mode of action in HT-29 human colon cancer cells. Cancer Lett. 2009, 285, 73–79. [Google Scholar] [CrossRef]
- Schmelz, E.M.; Dombrink-Kurtzman, M.A.; Roberts, P.C.; Kozutsumi, Y.; Kawasaki, T.; Merrill, A.H., Jr. Induction of apoptosis by fumonisin B1 in HT29 cells is mediated by the accumulation of endogenous free sphingoid bases. Toxicol. Appl. Pharmacol. 1998, 148, 252–260. [Google Scholar] [CrossRef]
- Kikuchi, H.; Hoshikawa, T.; Fujimura, S.; Sakata, N.; Kurata, S.; Katou, Y.; Oshima, Y. Isolation of a Cyclic Depsipetide, Aspergillicin F, and Synthesis of Aspergillicins with Innate Immune-Modulating Activity. J. Nat. Prod. 2015, 78, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 1 | 2 | 3 | |||
|---|---|---|---|---|---|---|
| δC, Type | δH (J in Hz) | δC, Type | δH (J in Hz) | δC, Type | δH (J in Hz) | |
| 1 | 162.5, C | 161.9, C | 162.1, C | |||
| 2 | 91.0, CH | 6.62, s | 90.3, CH | 6.50, s | 90.3, CH | 6.55, s |
| 3 | 163.5, C | 163.0, C | 162.9, C | |||
| 4 | 108.3, C | 107.0, C | 107.9, C | |||
| 5 | 106.8, CH | 6.94, dd (8.5, 0.9) | 108.9, CH | 7.02, dd (8.4, 0.9) | 108.7, CH | 7.04, d (8.3) |
| 6 | 134.3, CH | 7.65, t (8.4) | 134.1, CH | 7.61, t (8.3) | 133.9, CH | 7.62, t (8.4) |
| 7 | 109.1, CH | 7.08, dd (8.4, 0.9) | 106.8, CH | 6.93, dd (8.4, 0.9) | 106.6, CH | 6.93, d (8.3) |
| 8 | 155.9, C | 159.6, C | 159.6, C | |||
| 9 | 173.0, C | 173.2, C | 173.1, C | |||
| 10 | 159.6, C | 156.0, C | 155.8, C | |||
| 11 | 113.9, C | 112.9, C | 113.1, C | |||
| 12 | 108.2, C | 107.5, C | 106.0, C | |||
| 13 | 152.7, C | 152.2, C | 152.7, C | |||
| 1′ | 117.3, CH | 6.41, s | 113.3, CH | 6.56, d (6.1) | 111.0, CH | 6.48, d (5.9) |
| 2′ | 89.2, C | 42.0, CH | 4.19, dd (9.3, 6.2) | 41.8, CH | 4.24, ddd (9.2, 5.9, 3.4) | |
| 3′ | 148.3, CH | 6.77, d (2.8) | 36.7, CH2 | 2.39, ddd (13.4, 9.5, 5.2) 2.23, d (13.3) | 36.0, CH2 | 2.35, dd (13.2, 4.8, 3.4) 2.25, ddd (13.5, 9.0, 5.3) |
| 4′ | 107.0, CH | 5.71, d (2.8) | 106.3, CH | 5.25, d (5.0) | 106.1, CH | 5.17, t (5.0) |
| 1-OMe | 56.5, CH3 | 3.84, s | 56.3, CH3 | 3.81, s | 56.1, CH3 | 3.82, s |
| 8-OMe | 56.1, CH3 | 3.85, s | 56.1, CH3 | 3.85, s | 55.8, CH3 | 3.85, s |
| 4′-OH | 6.50, s | |||||
| 4′-OMe | 54.5, CH3 | 3.09, s | 55.7, CH3 | 3.36, s | ||
| Compound No | Compound Name | IC50 (µM) |
|---|---|---|
| 1 | oxisterigmatocystin J | 6.28 |
| 2 | oxisterigmatocystin K | 15.14 |
| 3 | oxisterigmatocystin L | 988.05 |
| 4 | aspergillicin A | 1.63 |
| Taxol | 0.48 |
| Cytotoxic Compound | Viable Cell (%) | Apoptotic Cell (%) | Necrotic Cell (%) |
|---|---|---|---|
| 1 | 64.53 ± 3.05 | 30.65 ± 3.22 | 4.81 ± 1.02 |
| 2 | 36.49 ± 4.08 | 59.38 ± 4.88 | 4.12 ± 1.37 |
| 4 | 25.24 ± 4.21 | 4.80 ± 0.98 | 69.95 ± 4.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Artasasta, M.A.; Yanwirasti, Y.; Taher, M.; Djamaan, A.; Ariantari, N.P.; Edrada-Ebel, R.A.; Handayani, D. Apoptotic Activity of New Oxisterigmatocystin Derivatives from the Marine-Derived Fungus Aspergillus nomius NC06. Mar. Drugs 2021, 19, 631. https://doi.org/10.3390/md19110631
Artasasta MA, Yanwirasti Y, Taher M, Djamaan A, Ariantari NP, Edrada-Ebel RA, Handayani D. Apoptotic Activity of New Oxisterigmatocystin Derivatives from the Marine-Derived Fungus Aspergillus nomius NC06. Marine Drugs. 2021; 19(11):631. https://doi.org/10.3390/md19110631
Chicago/Turabian StyleArtasasta, Muh. Ade, Yanwirasti Yanwirasti, Muhammad Taher, Akmal Djamaan, Ni Putu Ariantari, Ru Angelie Edrada-Ebel, and Dian Handayani. 2021. "Apoptotic Activity of New Oxisterigmatocystin Derivatives from the Marine-Derived Fungus Aspergillus nomius NC06" Marine Drugs 19, no. 11: 631. https://doi.org/10.3390/md19110631
APA StyleArtasasta, M. A., Yanwirasti, Y., Taher, M., Djamaan, A., Ariantari, N. P., Edrada-Ebel, R. A., & Handayani, D. (2021). Apoptotic Activity of New Oxisterigmatocystin Derivatives from the Marine-Derived Fungus Aspergillus nomius NC06. Marine Drugs, 19(11), 631. https://doi.org/10.3390/md19110631