
Fucoidan and Derived Oligo-Fucoses: Structural Features with Relevance in Competitive Inhibition of Gastrointestinal Norovirus Binding



Abstract
:
1. Introduction
2. Results
2.1. Fucoidans Compete with Blood Group Fucosyl Residues on Gastric Mucins for Binding to Noroviral VLPs
2.2. Sulfate Residues on Fucoidan Are Not Essential for the Blocking of VLP Binding to Gastric Mucins
2.3. Desulfated Fucoidan Oligosaccharides from FvF Reveal Unbranched Chains in the Size Range Up to Nonasaccharides
2.4. Hydrothermal Degradation of Fucoidans
2.5. Structural Data on UpF and Its Hydrothermal Degradation Products
3. Discussion
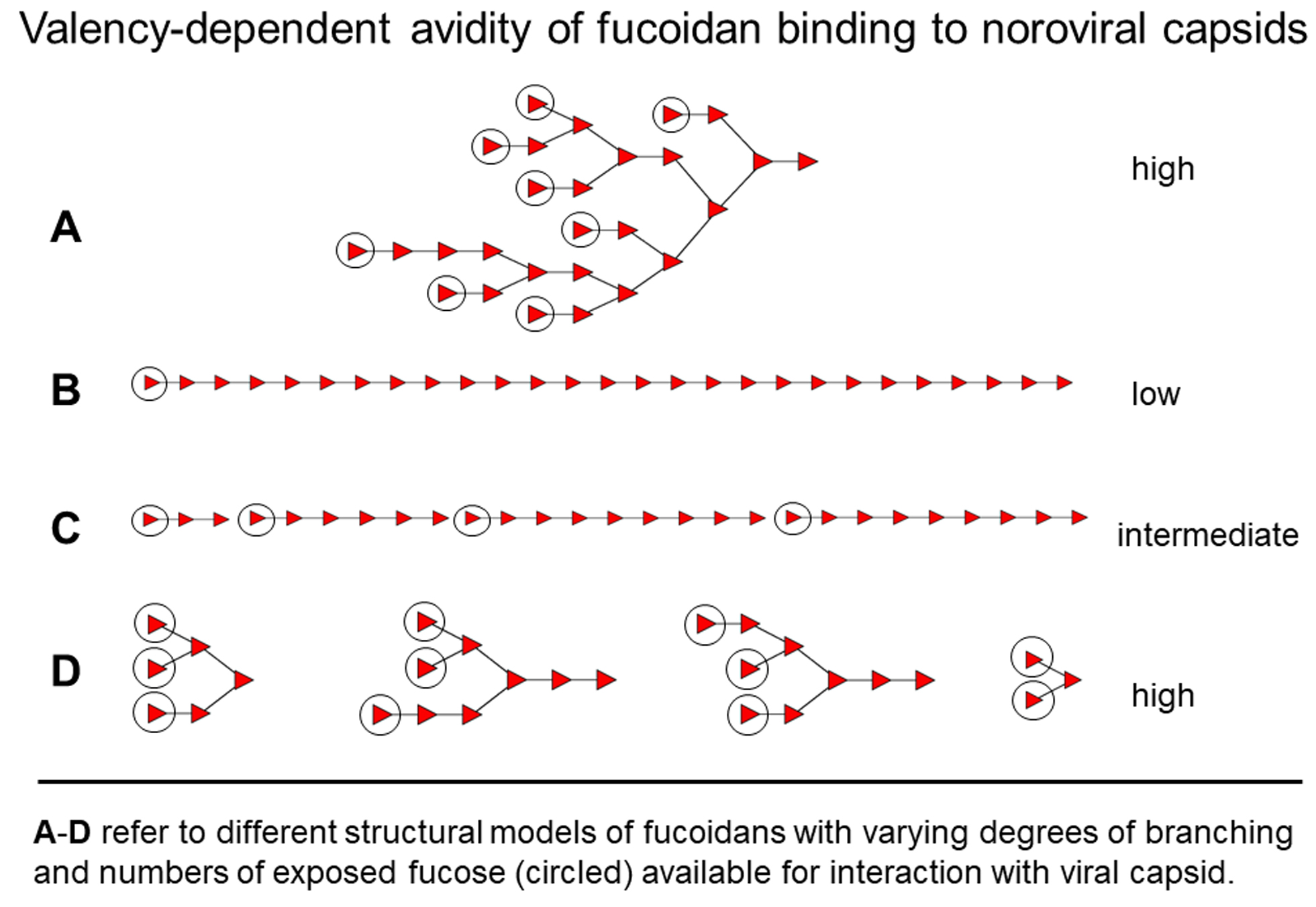
3.1. Fucose Multi-Valency as an Approach for Norovirus Drug Design
3.2. Essential Structural Features of UpF
4. Experimental Procedures
4.1. Materials
4.2. Processing of Fucoidan by Partial Acid Hydrolysis with Polystyrenesulfonic Acid
4.3. Fractionation of Fucoidan Processing Products and Alternative Workup Strategies
4.4. Hydrothermal Degradation of Fucoidans
4.5. Production of Norovirus VLPs
4.6. Norovirus Capsid Binding and Binding Inhibition Assays
4.7. Structural Studies by MALDI Mass Spectrometry
4.8. Linkage Analysis by GCMS
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Prasad, B.V.; Hardy, M.E.; Dokland, T.; Bella, J.; Rossmann, M.G.; Estes, M.K. X-ray crystallographic structure of the Norwalk virus capsid. Science 1999, 286, 287–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, D.P.; Ando, T.; Fankhauser, R.L.; Beard, R.S.; Glass, R.I.; Monroe, S.S. Norovirus classification and proposed strain nomenclature. Virology 2006, 346, 312–323. [Google Scholar] [CrossRef] [Green Version]
- Fankhauser, R.L.; Monroe, S.S.; Noel, J.S.; Humphrey, C.D.; Bresee, J.S.; Parashar, U.D.; Ando, T.; Glass, R.I. Epidemiologic and molecular trends of “Norwalk-like Viruses” associated with outbreaks of gastroenteritis in the United States. J. Infect. Dis. 2002, 186, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Marionneau, S.; Ruvoën, N.; Le Moullac-Vaidye, B.; Clement, M.; Cailleau-Thomas, A.; Ruiz-Palacois, G.; Huang, P.; Jiang, X.; Le Pendu, J. Norwalk virus binds to histo-blood group antigens present on gastroduodenal epithelial cells of secretor individuals. Gastroenterology 2002, 122, 1967–1977. [Google Scholar] [CrossRef]
- Schroten, H.; Hanisch, F.G.; Hansman, G.S. Human norovirus interactions with histo-blood group antigens and human milk oligosaccharides. J. Virol. 2016, 90, 5855–5859. [Google Scholar] [CrossRef] [Green Version]
- Lindesmith, L.; Moe, C.; Marionneau, S.; Ruvoen, N.; Jiang, X.; Lindblad, L.; Stewart, P.; LePendu, J.; Baric, R. Human susceptibility and resistance to Norwalk virus infection. Nat. Med. 2003, 9, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Lopman, B.A.; Trivedi, T.; Vicuna, Y.; Costantini, V.; Collins, N.; Gregoricus, N.; Parashar, U.; Sandoval, C.; Broncano, N.; Vaca, M.; et al. Norovirus infection and disease in an Ecuadorian Birth Cohort: Association of certain norovirus genotypes with host FUT2 secretor status. J. Infect. Dis. 2015, 211, 1813–1821. [Google Scholar] [CrossRef] [PubMed]
- Bhella, D. The role of cellular adhesion molecules in virus attachment and entry. Phil. Trans. R. Soc. B 2015, 370, 20140035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orchard, R.C.; Wilen, C.B.; Doench, J.G.; Baldrich, M.T.; McCune, B.T.; Lee, S.; Pruett-Miller, S.M.; Nelson, C.A.; Fremont, D.H.; Virgin, H.W. Discovery of a proteinaceous cellular receptor for a norovirus. Science 2016, 353, 933–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weichert, S.; Koromyslova, A.; Singh, B.K.; Hansman, S.; Jennewein, S.; Schroten, H.; Hansman, G.S. Structural basis for norovirus inhibition by human milk oligosaccharides. J. Virol. 2016, 90, 4843–4848. [Google Scholar] [CrossRef] [Green Version]
- Koromyslova, A.; Tripathi, S.; Morozov, V.; Schroten, H.; Hansman, G.S. Human norovirus inhibition by a human milk oligosaccharide. Virology 2017, 508, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Hanisch, F.G.; Hansman, G.S.; Morozov, V.; Kunz, C.; Schroten, S. Avidity of α-fucose on human milk oligosaccharides and blood group-unrelated oligo/polyfucoses is essential for potent norovirus-binding targets. J. Biol. Chem. 2018, 293, 11955–11965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Zheng, Y.; Wang, J.; Ma, S.; Yu, Y.; White, W.L.; Yang, S.; Yang, F.; Lu, J. Fucoidan extracted from Undaria pinnatifida: Source for nutraceuticals/functional foods. Mar. Drugs 2018, 16, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshino, T.; Hayashi, T.; Hayashi, K.; Hamada, J.; Lee, J.B.; Sankawa, U. An antivirally active sulfated polysaccharide from Sargassum horneri (TURNER) C. AGARDH. Biol. Pharm. Bull. 1998, 21, 730–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iqbal, M.; Flick-Smith, H.; McCauley, J.W. Interactions of bovine viral diarrhoea virus glycoprotein E (rns) with cell surface glycosaminoglycans. J. Gen. Virol. 2000, 81, 451–459. [Google Scholar]
- Lahrsen, E.; Liewert, I.; Alban, S. Gradual degradation of fucoidan from Fucus vesiculosus and its effect on structure, antioxidant and antiproliferative activities. Carbohydr. Polym. 2018, 192, 208–216. [Google Scholar] [CrossRef]
- Patankar, M.S.; Oehninger, S.; Barnett, T.; Williams, R.L.; Clark, G.F. A revised structure for fucoidan may explain some of its biological activities. J. Biol. Chem. 1993, 268, 21770–21776. [Google Scholar] [CrossRef]
- Domon, B.; Costello, C.E. A systematic nomenclature for carbohydrate fragmentations in FAB-MS/MS spectra of glycoconjugates. Glycoconj. J. 1988, 5, 397–409. [Google Scholar] [CrossRef]
- Haroun-Bouhedja, F.; Ellouali, M.; Sinquin, C.; Boisson-Vidal, C. Relationship between sulfate groups and biological activities of fucans. Thromb. Res. 2000, 100, 453–459. [Google Scholar] [CrossRef]
- Bücher, K.S.; Yan, H.; Creutznacher, R.; Ruoff, K.; Mallagaray, A.; Grafmüller, A.; Dirks, J.S.; Kilic, T.; Weickert, S.; Rubailo, A.; et al. Fucose-functionalized precision glycomacromolecules targeting human norovirus capsid protein. Biomacromolecules 2018, 19, 3714–3724. [Google Scholar] [CrossRef]
- Kim, H.; Lim, C.Y.; Lee, D.B.; Seok, J.H.; Kim, K.H.; Chung, M.S. Inhibitory effects of Laminaria japonica fucoidans against noroviruses. Viruses 2020, 12, 997. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.T.H.; Li, Y.; Gorji, M.E.; Gong, Z.; Li, D. Fucoidan but not 2′-fucosyllactose inhibits human norovirus replication in zebrafish larvae. Viruses 2021, 13, 461. [Google Scholar] [CrossRef]
- Hemmingson, J.A.; Falshaw, R.; Furneaux, R.H. Structure and antiviral activity of the galactofucan sulfates extracted from Undaria pinnatifida (Phaeophyta). J. Appl. Phycol. 2006, 18, 185–193. [Google Scholar] [CrossRef]
- Fitton, J.H. Therapies from fucoidan; multifunctional marine polymers. Mar. Drugs 2011, 9, 1731–1760. [Google Scholar] [CrossRef] [PubMed]
- Clement, M.J.; Tissot, B.; Chevolot, L.; Adjadj, E.; Du, Y.; Curmi, P.A.; Daniel, R. NMR characterization and molecular modeling of fucoidan showing the importance of oligosaccharide branching in its anticomplementary activity. Glycobiology 2010, 20, 883–894. [Google Scholar] [CrossRef] [Green Version]
- Quitain, A.T.; Kai, T.; Sasaki, M.; Goto, M. Microwave-hydrothermal extraction and degradation of fucoidan from supercritical carbon dioxide deoiled Undaria pinnatifida. Ind. Eng. Chem. Res. 2013, 52, 7940–7946. [Google Scholar] [CrossRef]
- Hansman, G.S.; Saito, H.; Shibata, C.; Ishizuka, S.; Oseto, M.; Oka, T.; Takeda, N. Outbreak of gastroenteritis due to sapovirus. J. Clin. Microbiol. 2007, 45, 1347–1349. [Google Scholar] [CrossRef] [Green Version]
- Hanisch, F.G.; Kunz, C. Novel class of human milk oligosaccharides based on 6′-galactosyllactose containing N-acetylglucosamine branches extended by oligogalactoses. J. Proteome Res. 2021, 20, 3865–3874. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample * | Molar ratio l-Fuc/d-Gal | Estimated Molecular Masses (kD) |
|---|---|---|
| UpF (nutritional-grade) | 0.953 | n.d. |
| UpF (pure) | 1.226 | n.d. |
| UpF-HTD | 1.964 | <10 |
| UpF-PSSA | 0.876 | <6–8 |
| Sample | Fuc-ol/Gal-ol | Fuc/Fuc-ol |
|---|---|---|
| UpF native | - | - |
| UpF PSSA filtrate | >100 | 0.78 |
| UpF PSSA CG | >10 | 0.92 |
| UpF HTD-1 | >30 | 0.23 |
| UpF HTD-2 | >10 | 0.18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanisch, F.-G.; Aydogan, C.; Schroten, H. Fucoidan and Derived Oligo-Fucoses: Structural Features with Relevance in Competitive Inhibition of Gastrointestinal Norovirus Binding. Mar. Drugs 2021, 19, 591. https://doi.org/10.3390/md19110591
Hanisch F-G, Aydogan C, Schroten H. Fucoidan and Derived Oligo-Fucoses: Structural Features with Relevance in Competitive Inhibition of Gastrointestinal Norovirus Binding. Marine Drugs. 2021; 19(11):591. https://doi.org/10.3390/md19110591
Chicago/Turabian StyleHanisch, Franz-Georg, Cem Aydogan, and Horst Schroten. 2021. "Fucoidan and Derived Oligo-Fucoses: Structural Features with Relevance in Competitive Inhibition of Gastrointestinal Norovirus Binding" Marine Drugs 19, no. 11: 591. https://doi.org/10.3390/md19110591
APA StyleHanisch, F.-G., Aydogan, C., & Schroten, H. (2021). Fucoidan and Derived Oligo-Fucoses: Structural Features with Relevance in Competitive Inhibition of Gastrointestinal Norovirus Binding. Marine Drugs, 19(11), 591. https://doi.org/10.3390/md19110591