
Marine-Derived Compounds for the Potential Treatment of Glucocorticoid Resistance in Severe Asthma

 ,
,  , and
, and
Abstract
:1. Introduction
2. Pathobiological Features Associated with Glucocorticoid Resistance
3. Mechanisms of Glucocorticoid Resistance as Potential Therapeutic Targets
3.1. Mechanisms of Action of Glucocorticoids
3.2. Glucocorticoids Resistance: Cellular and Molecular Basis
- Deficient binding between the GC and the GR or between the GR complex and DNA may be a cause [39].
- Increased antagonism is determined either by increased GR-β expression [40] or by diminished GR-α expression [41]. This can be explained by the IL-2/IL-4-induced suppression of GR-α (and not GR-β) expression in peripheral blood mononuclear cells (PBMCs) [42]. Additionally, IL-2 and IL-4 can synergistically reduce (via the p38MAPK pathway) nuclear translocation and binding affinity in T-cells (reversible by a p38 inhibitor) [43]. Furthermore, IL-17 and IL-23 cytokines were reported to significantly upregulate GR-β [42].
- Inflammation or oxidative stress has the potential to negatively affect GC signaling [22].
- The upregulation of certain cytokines, such as IL-2, IL-4, and IL-13, was detected in the lungs of patients with GC unresponsiveness [45,46,47]; in vitro, the overexpression of these cytokines was associated with the phosphorylation of GR and a decrease in nuclear translocation in inflammatory cells through the activation of p38 mitogen-activated protein kinase [48]. p38MAPK activity was demonstrated to be higher in alveolar macrophages from patients with impaired response to GCs compared to ‘responders’. Furthermore, the expression of MKP-1 (DUSP1 gene), an endogenous inhibitor of the MAPK pathway, was significantly diminished in alveolar macrophages after GCS exposure, leading to an increase in p38MAPK activity [49]. Furthermore, p38MAPK inhibitors, such as AZD7624 or SB203580, have recently been investigated in corticosteroid-resistant asthmatic populations [50,51].
- Increased HDAC activity using theophylline, PI3K, and p38 MAPK inhibitors demonstrated beneficial effects [52,53,54], especially in glucocorticoid-resistant asthmatic smokers, where increased antagonism of the GR-α resulted from a reduced ratio of GR-α to GR-β isoforms [55]. Moreover, reduced total HDAC activity in PBMCs isolated from prednisone-dependent asthmatics compared to ICS-maintained moderate asthmatics and healthy volunteers was reported [56].
- GC resistance has been associated with Haemophilus influenzae, Chlamydia pneumoniae, Influenza A virus (IAV), rhinovirus, and Respiratory syncytial virus (RSV) infections [57,58,59,60,61]. The molecular mechanism proposed for glucocorticoid insensitivity in rhinovirus-infected primary human bronchial epithelial cells is the activation of NF-κB and c-Jun N-terminal kinase, which leads to a decrease in GR-α nuclear translocation [62]. The influence of NF-κB activity on GC resistance has also been confirmed by research on the blockade of this pathway [63,64].
- Using mouse models of steroid-resistant asthma driven by bacterial (Chlamydia and Haemophilus influenzae) and viral (influenza and RSV) respiratory tract infections, Kim et al. demonstrated that steroid insensitivity can be induced through PI3K-mediated phosphorylation and the nuclear translocation of pAKT [65].
- By upregulating miR-9 expression in pulmonary macrophages, IFN-γ can increase GR phosphorylation and, consequently, inhibit GR nuclear translocation in experimental models of steroid-resistant airway hyperresponsiveness [66].
- In a study of human fetal airway smooth muscle cells, TNF-α and IFN-γ cytokines were shown to sustain GC resistance by promoting the Nuclear factor-κB (NF-κB) pathway and Stat1 phosphorylation [67]. TNF-α also demonstrated the potential to activate the c-Jun N-terminal kinase (JNK), which directly phosphorylated GR-α at Ser226 and inhibited GRE-binding [68].
- The nitrosylation of the glucocorticoid receptor at the HSP90 (chaperone) binding site can be caused by high levels of nitric oxide generated in situ as a result of eosinophilic inflammation. This can decrease its affinity with chaperone proteins that protect it from cytoplasmic degradation. The binding affinity to GCs (ligand) in structural cells, such as fibroblasts, can also be reduced by nitrosylation [69]. In conclusion, asthmatics with persistent airway eosinophilia with increased localized nitric oxide production and possibly increased remodeling may develop GC resistance through the repeated nitrosylation of GR.
- Increased NLR Family Pyrin Domain Containing 3 (NLRP3) inflammasome/IL-1β activation contributed to glucocorticoid resistance in murine models of steroid-resistant allergic airway disease [70].
- The Th2 cytokines IL-13 and IL-5 each possess the ability to induce diminished GR-binding affinity. The effect of hydrocortisone in suppressing LPS-induced IL-6 production by monocytes was demonstrated to be significantly hindered when the cells were primed by IL-13 [71]. Additionally, IL-5-primed eosinophils were unresponsive to GS-induced apoptosis (via synergistic upregulation of nuclear-factor IL-3 due to a cross-talk between GCS-induced trans-activation signaling and IL-5 antiapoptotic pathway) [72].
- The adoptive transfer of Th17 cells in mice resulted in the development of steroid insensitivity, and Th17 cells and IL-17A levels are frequently associated with CG resistance in asthmatic patients [73,74,75]. Accordingly, the expression of GR-β has been reported to increase Th17 responses [76]. In the obesity phenotype of asthma, the associated steroid resistance may be induced by IL-17 produced by the pulmonary type 3 innate lymphoid cells [77]. The role of IL-7 in GC resistance has been confirmed by the augmentation of dexamethasone anti-inflammatory action in diesel exhaust particle-induced neutrophilic steroid insensitivity secondary to anti-IL-17 therapy [78].
- Bhavsar et al. showed that dexamethasone could not suppress the lipopolysaccharide (LPS)-induced release of pro-inflammatory cytokines [49]. This finding was supported by Li et al., who simulated an airway infection in a mouse model of steroid-resistant asthma through the concomitant administration of LPS + IFNγ; consequently, PP2A activity (that induced JNK) was attenuated and led to the phosphorylation of GR-α at Ser226, thereby hindering glucocorticoid receptor nuclear translocation in pulmonary macrophages [66].
- LPS promoted a shift from Th2-derived airway eosinophilic inflammation to Th17-drived neutrophilic inflammation in an ovalbumin-sensitized murine asthma model [79].
- Dysregulated IL-10 production is associated with GC insensitivity. This is probably due to impaired IL-10 production, according to Hawrylowicz et al., who compared in vitro stimulated T lymphocytes from corticosteroid-resistant asthmatic with dexamethasone to T lymphocytes from steroid-sensitive asthmatics [80].
- The induction of Th2/Th17 responses in fungus-exposed patients has the potential to develop GC resistance [65]. More precisely, in neonatal mice, Aspergillus alternata exposure induced IL-33 dependent GC resistant asthma, mediated by ILC2 and Th2 cells [81]. The suggested mechanism underlying glucocorticoid insensitivity is the activation of p38-MAPK in CD4 + T cells and induction of phosphorylation of GR by IL-33 [82].
4. Potentially Therapeutic Effect of Marine-Derived Biologically Active Compounds in Severe Asthma
4.1. Cellular Signal/Corticoresistance
4.2. Cytokine Levels
4.2.1. Th2 Cytokines
4.2.2. Th17/Non Th2 Cytokines
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Borna, E.; Nwaru, B.I.; Bjerg, A.; Mincheva, R.; Rådinger, M.; Lundbäck, B.; Ekerljung, L. Changes in the Prevalence of Asthma and Respiratory Symptoms in Western Sweden between 2008 and 2016. Allergy 2019, 74, 1703–1715. [Google Scholar] [CrossRef]
- Hekking, P.-P.W.; Wener, R.R.; Amelink, M.; Zwinderman, A.H.; Bouvy, M.L.; Bel, E.H. The Prevalence of Severe Refractory Asthma. J. Allergy Clin. Immunol. 2015, 135, 896–902. [Google Scholar] [CrossRef]
- Chung, K.F.; Wenzel, S.E.; Brozek, J.L.; Bush, A.; Castro, M.; Sterk, P.J.; Adcock, I.M.; Bateman, E.D.; Bel, E.H.; Bleecker, E.R.; et al. International ERS/ATS Guidelines on Definition, Evaluation and Treatment of Severe Asthma. Eur. Respir. J. 2014, 43, 343–373. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, S.; Sweeney, J.; Patterson, C.C.; Menzies-Gow, A.; Niven, R.; Mansur, A.H.; Bucknall, C.; Chaudhuri, R.; Thomson, N.C.; Brightling, C.E.; et al. The Cost of Treating Severe Refractory Asthma in the UK: An Economic Analysis from the British Thoracic Society Difficult Asthma Registry. Thorax 2015, 70, 376–378. [Google Scholar] [CrossRef] [Green Version]
- Global Initiative for Asthma. Available online: https://ginasthma.org/ (accessed on 27 September 2021).
- Carmichael, J.; Paterson, I.C.; Diaz, P.; Crompton, G.K.; Kay, A.B.; Grant, I.W. Corticosteroid Resistance in Chronic Asthma. Br. Med. J. (Clin. Res. Ed.) 1981, 282, 1419–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, P.J.; Greening, A.P.; Crompton, G.K. Glucocorticoid Resistance in Asthma. Am. J. Respir. Crit. Care Med. 1995, 152 Pt 2, S125–S140. [Google Scholar] [CrossRef] [PubMed]
- Löwhagen, O. Diagnosis of Asthma—New Theories. J. Asthma 2015, 52, 538–544. [Google Scholar] [CrossRef]
- Barreca, M.; Spanò, V.; Montalbano, A.; Cueto, M.; Díaz Marrero, A.R.; Deniz, I.; Erdoğan, A.; Lukić Bilela, L.; Moulin, C.; Taffin-de-Givenchy, E.; et al. Marine Anticancer Agents: An Overview with a Particular Focus on Their Chemical Classes. Mar. Drugs 2020, 18, 619. [Google Scholar] [CrossRef]
- Chen, G.; Seukep, A.J.; Guo, M. Recent Advances in Molecular Docking for the Research and Discovery of Potential Marine Drugs. Mar. Drugs 2020, 18, 545. [Google Scholar] [CrossRef] [PubMed]
- Grosso, C.; Valentão, P.; Ferreres, F.; Andrade, P.B. Bioactive Marine Drugs and Marine Biomaterials for Brain Diseases. Mar. Drugs 2014, 12, 2539–2589. [Google Scholar] [CrossRef] [Green Version]
- Mayer, A.M.S.; Guerrero, A.J.; Rodríguez, A.D.; Taglialatela-Scafati, O.; Nakamura, F.; Fusetani, N. Marine Pharmacology in 2014–2015: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis, Antiviral, and Anthelmintic Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action. Mar. Drugs 2019, 18, 5. [Google Scholar] [CrossRef] [Green Version]
- Mayer, A.M.S.; Guerrero, A.J.; Rodríguez, A.D.; Taglialatela-Scafati, O.; Nakamura, F.; Fusetani, N. Marine Pharmacology in 2016–2017: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis and Antiviral Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action. Mar. Drugs 2021, 19, 49. [Google Scholar] [CrossRef] [PubMed]
- Yun, C.W.; Kim, H.J.; Lee, S.H. Therapeutic Application of Diverse Marine-Derived Natural Products in Cancer Therapy. Anticancer Res. 2019, 39, 5261–5284. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.-B.; Luo, R.; Zheng, Y.-L.; Pang, J.-Y. Recent Advances in the Discovery and Development of Marine Natural Products with Cardiovascular Pharmacological Effects. Mini Rev. Med. Chem. 2018, 18, 527–550. [Google Scholar] [CrossRef] [PubMed]
- Bilal, M.; Qindeel, M.; Nunes, L.V.; Duarte, M.T.S.; Ferreira, L.F.R.; Soriano, R.N.; Iqbal, H.M.N. Marine-Derived Biologically Active Compounds for the Potential Treatment of Rheumatoid Arthritis. Mar. Drugs 2021, 19, 10. [Google Scholar] [CrossRef]
- Bălașa, A.F.; Chircov, C.; Grumezescu, A.M. Marine Biocompounds for Neuroprotection—A Review. Mar. Drugs 2020, 18, 290. [Google Scholar] [CrossRef]
- Wenzel, S.E. Asthma Phenotypes: The Evolution from Clinical to Molecular Approaches. Nat. Med. 2012, 18, 716–725. [Google Scholar] [CrossRef]
- Chambers, E.S.; Nanzer, A.M.; Pfeffer, P.E.; Richards, D.F.; Timms, P.M.; Martineau, A.R.; Griffiths, C.J.; Corrigan, C.J.; Hawrylowicz, C.M. Distinct Endotypes of Steroid-Resistant Asthma Characterized by IL-17Ahigh and IFN-Γhigh Immunophenotypes: Potential Benefits of Calcitriol. J. Allergy Clin. Immunol. 2015, 136, 628–637.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keenan, C.R.; Radojicic, D.; Li, M.; Radwan, A.; Stewart, A.G. Heterogeneity in Mechanisms Influencing Glucocorticoid Sensitivity: The Need for a Systems Biology Approach to Treatment of Glucocorticoid-Resistant Inflammation. Pharmacol. Ther. 2015, 150, 81–93. [Google Scholar] [CrossRef]
- Barnes, P.J.; Adcock, I.M. Glucocorticoid Resistance in Inflammatory Diseases. Lancet 2009, 373, 1905–1917. [Google Scholar] [CrossRef]
- Barnes, P.J. Corticosteroid Resistance in Patients with Asthma and Chronic Obstructive Pulmonary Disease. J. Allergy Clin. Immunol. 2013, 131, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Keenan, C.R.; Salem, S.; Fietz, E.R.; Gualano, R.C.; Stewart, A.G. Glucocorticoid-Resistant Asthma and Novel Anti-Inflammatory Drugs. Drug Discov. Today 2012, 17, 1031–1038. [Google Scholar] [CrossRef]
- Scheschowitsch, K.; Leite, J.A.; Assreuy, J. New Insights in Glucocorticoid Receptor Signaling-More Than Just a Ligand-Binding Receptor. Front. Endocrinol. (Lausanne) 2017, 8, 16. [Google Scholar] [CrossRef]
- Oakley, R.H.; Sar, M.; Cidlowski, J.A. The Human Glucocorticoid Receptor β Isoform: Expression, Biochemical Properties, and Putative Function. J. Biol. Chem. 1996, 271, 9550–9559. [Google Scholar] [CrossRef] [Green Version]
- Charmandari, E.; Chrousos, G.P.; Ichijo, T.; Bhattacharyya, N.; Vottero, A.; Souvatzoglou, E.; Kino, T. The Human Glucocorticoid Receptor (HGR) β Isoform Suppresses the Transcriptional Activity of HGRα by Interfering with Formation of Active Coactivator Complexes. Mol. Endocrinol. 2005, 19, 52–64. [Google Scholar] [CrossRef] [Green Version]
- Stahn, C.; Buttgereit, F. Genomic and Nongenomic Effects of Glucocorticoids. Nat. Rev. Rheumatol. 2008, 4, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Galigniana, M.D.; Echeverría, P.C.; Erlejman, A.G.; Piwien-Pilipuk, G. Role of Molecular Chaperones and TPR-Domain Proteins in the Cytoplasmic Transport of Steroid Receptors and Their Passage through the Nuclear Pore. Nucleus 2010, 1, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Galon, J.; Franchimont, D.; Hiroi, N.; Frey, G.; Boettner, A.; Ehrhart-Bornstein, M.; O’Shea, J.J.; Chrousos, G.P.; Bornstein, S.R. Gene Profiling Reveals Unknown Enhancing and Suppressive Actions of Glucocorticoids on Immune Cells. FASEB J. 2002, 16, 61–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, M.; Itkin, I.H. Effectiveness and Complications of Aerosol Dexamethasone Phosphate in Severe Asthma. J. Allergy 1964, 35, 70–76. [Google Scholar] [CrossRef]
- Schwiebert, L.A.; Beck, L.A.; Stellato, C.; Bickel, C.A.; Bochner, B.S.; Schleimer, R.P. Glucocorticosteroid Inhibition of Cytokine Production: Relevance to Antiallergic Actions. J. Allergy Clin. Immunol. 1996, 97 Pt 2, 143–152. [Google Scholar] [CrossRef]
- Alangari, A.A. Genomic and Non-Genomic Actions of Glucocorticoids in Asthma. Ann. Thorac. Med. 2010, 5, 133. [Google Scholar] [CrossRef]
- Hirst, S.J.; Lee, T.H. Airway Smooth Muscle as a Target of Glucocorticoid Action in the Treatment of Asthma. Am. J. Respir. Crit. Care Med. 1998, 158 (Suppl. S2), S201–S206. [Google Scholar] [CrossRef]
- Zhou, J.; Liu, D.-F.; Liu, C.; Kang, Z.-M.; Shen, X.-H.; Chen, Y.-Z.; Xu, T.; Jiang, C.-L. Glucocorticoids Inhibit Degranulation of Mast Cells in Allergic Asthma via Nongenomic Mechanism. Allergy 2008, 63, 1177–1185. [Google Scholar] [CrossRef]
- Ayroldi, E.; Cannarile, L.; Migliorati, G.; Nocentini, G.; Delfino, D.V.; Riccardi, C. Mechanisms of the Anti-Inflammatory Effects of Glucocorticoids: Genomic and Nongenomic Interference with MAPK Signaling Pathways. FASEB J. 2012, 26, 4805–4820. [Google Scholar] [CrossRef] [PubMed]
- Tantisira, K.G.; Lasky-Su, J.; Harada, M.; Murphy, A.; Litonjua, A.A.; Himes, B.E.; Lange, C.; Lazarus, R.; Sylvia, J.; Klanderman, B.; et al. Genomewide Association between GLCCI1 and Response to Glucocorticoid Therapy in Asthma. N. Engl. J. Med. 2011, 365, 1173–1183. [Google Scholar] [CrossRef] [Green Version]
- Keskin, O.; Uluca, Ü.; Birben, E.; Coşkun, Y.; Ozkars, M.Y.; Keskin, M.; Kucukosmanoglu, E.; Kalayci, O. Genetic Associations of the Response to Inhaled Corticosteroids in Children during an Asthma Exacerbation. Pediatr. Allergy Immunol. 2016, 27, 507–513. [Google Scholar] [CrossRef]
- Hu, C.; Xun, Q.; Li, X.; He, R.; Lu, R.; Zhang, S.; Hu, X.; Feng, J. GLCCI1 Variation Is Associated with Asthma Susceptibility and Inhaled Corticosteroid Response in a Chinese Han Population. Arch. Med. Res. 2016, 47, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Weigel, N.L.; Moore, N.L. Steroid Receptor Phosphorylation: A Key Modulator of Multiple Receptor Functions. Mol. Endocrinol. 2007, 21, 2311–2319. [Google Scholar] [CrossRef] [Green Version]
- Adcock, I.M.; Lane, S.J.; Brown, C.R.; Lee, T.H.; Barnes, P.J. Abnormal Glucocorticoid Receptor-Activator Protein 1 Interaction in Steroid-Resistant Asthma. J. Exp. Med. 1995, 182, 1951–1958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramamoorthy, S.; Cidlowski, J.A. Ligand-Induced Repression of the Glucocorticoid Receptor Gene Is Mediated by an NCoR1 Repression Complex Formed by Long-Range Chromatin Interactions with Intragenic Glucocorticoid Response Elements. Mol. Cell. Biol. 2013, 33, 1711–1722. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-Tello, A.; Halwani, R.; Hamid, Q.; Al-Muhsen, S. Glucocorticoid Receptor-Beta up-Regulation and Steroid Resistance Induction by IL-17 and IL-23 Cytokine Stimulation in Peripheral Mononuclear Cells. J. Clin. Immunol. 2013, 33, 466–478. [Google Scholar] [CrossRef]
- Irusen, E.; Matthews, J.G.; Takahashi, A.; Barnes, P.J.; Chung, K.F.; Adcock, I.M. P38 Mitogen-Activated Protein Kinase–Induced Glucocorticoid Receptor Phosphorylation Reduces Its Activity: Role in Steroid-Insensitive Asthma. J. Allergy Clin. Immunol. 2002, 109, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Adcock, I.M. Glucocorticoid-Regulated Transcription Factors. Pulm. Pharmacol. Ther. 2001, 14, 211–219. [Google Scholar] [CrossRef]
- Leung, D.Y.; Martin, R.J.; Szefler, S.J.; Sher, E.R.; Ying, S.; Kay, A.B.; Hamid, Q. Dysregulation of Interleukin 4, Interleukin 5, and Interferon Gamma Gene Expression in Steroid-Resistant Asthma. J. Exp. Med. 1995, 181, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Gurgone, D.; McShane, L.; McSharry, C.; Guzik, T.J.; Maffia, P. Cytokines at the Interplay Between Asthma and Atherosclerosis? Front. Pharmacol. 2020, 11, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, M.C.; Wenzel, S.E. Intersection of Biology and Therapeutics: Type 2 Targeted Therapeutics for Adult Asthma. Lancet 2020, 395, 371–383. [Google Scholar] [CrossRef]
- Ito, K.; Yamamura, S.; Essilfie-Quaye, S.; Cosio, B.; Ito, M.; Barnes, P.J.; Adcock, I.M. Histone Deacetylase 2–Mediated Deacetylation of the Glucocorticoid Receptor Enables NF-ΚB Suppression. J. Exp. Med. 2005, 203, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Bhavsar, P.; Hew, M.; Khorasani, N.; Torrego, A.; Barnes, P.J.; Adcock, I.; Chung, K.F. Relative Corticosteroid Insensitivity of Alveolar Macrophages in Severe Asthma Compared with Non-Severe Asthma. Thorax 2008, 63, 784–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, N.R.; Cunoosamy, D.M.; Fagerås, M.; Taib, Z.; Asimus, S.; Hegelund-Myrbäck, T.; Lundin, S.; Pardali, K.; Kurian, N.; Ersdal, E.; et al. The Development of AZD7624 for Prevention of Exacerbations in COPD: A Randomized Controlled Trial. Int. J. Chron. Obstruct. Pulm. Dis. 2018, 13, 1009–1019. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Leung, D.Y.M.; Goleva, E. Activated P38 MAPK in Peripheral Blood Monocytes of Steroid Resistant Asthmatics. PLoS ONE 2015, 10, e0141909. [Google Scholar] [CrossRef] [Green Version]
- Spears, M.; Donnelly, I.; Jolly, L.; Brannigan, M.; Ito, K.; McSharry, C.; Lafferty, J.; Chaudhuri, R.; Braganza, G.; Adcock, I.M.; et al. Effect of Low-Dose Theophylline plus Beclometasone on Lung Function in Smokers with Asthma: A Pilot Study. Eur. Respir. J. 2009, 33, 1010–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercado, N.; Hakim, A.; Kobayashi, Y.; Meah, S.; Usmani, O.S.; Chung, K.F.; Barnes, P.J.; Ito, K. Restoration of Corticosteroid Sensitivity by P38 Mitogen Activated Protein Kinase Inhibition in Peripheral Blood Mononuclear Cells from Severe Asthma. PLoS ONE 2012, 7, e41582. [Google Scholar] [CrossRef] [Green Version]
- Bi, J.; Min, Z.; Yuan, H.; Jiang, Z.; Mao, R.; Zhu, T.; Liu, C.; Zeng, Y.; Song, J.; Du, C.; et al. PI3K Inhibitor Treatment Ameliorates the Glucocorticoid Insensitivity of PBMCs in Severe Asthma. Clin. Transl. Med. 2020, 9, 22. [Google Scholar] [CrossRef] [Green Version]
- Ito, K.; Lim, S.; Caramori, G.; Chung, K.F.; Barnes, P.J.; Adcock, I.M. Cigarette Smoking Reduces Histone Deacetylase 2 Expression, Enhances Cytokine Expression, and Inhibits Glucocorticoid Actions in Alveolar Macrophages. FASEB J. 2001, 15, 1110–1112. [Google Scholar] [CrossRef] [Green Version]
- Zuccaro, L.; Cox, A.; Pray, C.; Radford, K.; Novakowski, K.; Dorrington, M.; Surette, M.G.; Bowdish, D.; Nair, P. Histone Deacetylase Activity and Recurrent Bacterial Bronchitis in Severe Eosinophilic Asthma. Allergy 2016, 71, 571–575. [Google Scholar] [CrossRef]
- Essilfie, A.-T.; Simpson, J.L.; Horvat, J.C.; Preston, J.A.; Dunkley, M.L.; Foster, P.S.; Gibson, P.G.; Hansbro, P.M. Haemophilus influenzae Infection Drives IL-17-Mediated Neutrophilic Allergic Airways Disease. PLoS Pathog. 2011, 7, e1002244. [Google Scholar] [CrossRef]
- Yang, X.; Wang, Y.; Zhao, S.; Wang, R.; Wang, C. Long-Term Exposure to Low-Dose Haemophilus influenzae during Allergic Airway Disease Drives a Steroid-Resistant Neutrophilic Inflammation and Promotes Airway Remodeling. Oncotarget 2018, 9, 24898–24913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paróczai, D.; Mosolygó, T.; Kókai, D.; Endrész, V.; Virok, D.P.; Somfay, A.; Burián, K. Chlamydia Pneumoniae Influence on Cytokine Production in Steroid-Resistant and Steroid-Sensitive Asthmatics. Pathogens 2020, 9, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beale, J.; Jayaraman, A.; Jackson, D.J.; Macintyre, J.D.R.; Edwards, M.R.; Walton, R.P.; Zhu, J.; Ching, Y.M.; Shamji, B.; Edwards, M.; et al. Rhinovirus-Induced IL-25 in Asthma Exacerbation Drives Type 2 Immunity and Allergic Pulmonary Inflammation. Sci. Transl. Med. 2014, 6, 256ra134. [Google Scholar] [CrossRef] [Green Version]
- Papi, A.; Contoli, M.; Adcock, I.M.; Bellettato, C.; Padovani, A.; Casolari, P.; Stanciu, L.A.; Barnes, P.J.; Johnston, S.L.; Ito, K.; et al. Rhinovirus Infection Causes Steroid Resistance in Airway Epithelium through Nuclear Factor ΚB and C-Jun N-Terminal Kinase Activation. J. Allergy Clin. Immunol. 2013, 132, 1075–1085. [Google Scholar] [CrossRef]
- Reidl, J.; Monsó, E. Glucocorticoids and Antibiotics, How Do They Get Together? EMBO Mol. Med. 2015, 7, 992–993. [Google Scholar] [CrossRef]
- Shimizu, K.; Konno, S.; Ozaki, M.; Umezawa, K.; Yamashita, K.; Todo, S.; Nishimura, M. Dehydroxymethylepoxyquinomicin (DHMEQ), a Novel NF-KappaB Inhibitor, Inhibits Allergic Inflammation and Airway Remodelling in Murine Models of Asthma. Clin. Exp. Allergy 2012, 42, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Newton, R.; Holden, N.S.; Catley, M.C.; Oyelusi, W.; Leigh, R.; Proud, D.; Barnes, P.J. Repression of Inflammatory Gene Expression in Human Pulmonary Epithelial Cells by Small-Molecule IκB Kinase Inhibitors. J. Pharmacol. Exp. Ther. 2007, 321, 734–742. [Google Scholar] [CrossRef] [PubMed]
- Castanhinha, S.; Sherburn, R.; Walker, S.; Gupta, A.; Bossley, C.J.; Buckley, J.; Ullmann, N.; Grychtol, R.; Campbell, G.; Maglione, M.; et al. Pediatric Severe Asthma with Fungal Sensitization Is Mediated by Steroid-Resistant IL-33. J. Allergy Clin. Immunol. 2015, 136, 312–322.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.J.; Tay, H.L.; Maltby, S.; Xiang, Y.; Eyers, F.; Hatchwell, L.; Zhou, H.; Toop, H.D.; Morris, J.C.; Nair, P.; et al. MicroRNA-9 Regulates Steroid-Resistant Airway Hyperresponsiveness by Reducing Protein Phosphatase 2A Activity. J. Allergy Clin. Immunol. 2015, 136, 462–473. [Google Scholar] [CrossRef] [PubMed]
- Britt, R.D.; Thompson, M.A.; Sasse, S.; Pabelick, C.M.; Gerber, A.N.; Prakash, Y.S. Th1 Cytokines TNF-α and IFN-γ Promote Corticosteroid Resistance in Developing Human Airway Smooth Muscle. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2019, 316, L71–L81. [Google Scholar] [CrossRef]
- Ismaili, N.; Garabedian, M.J. Modulation of Glucocorticoid Receptor Function via Phosphorylation. Ann. N. Y. Acad. Sci. 2004, 1024, 86–101. [Google Scholar] [CrossRef] [PubMed]
- Galigniana, M.D.; Piwien-Pilipuk, G.; Assreuy, J. Inhibition of Glucocorticoid Receptor Binding by Nitric Oxide. Mol. Pharmacol. 1999, 55, 317–323. [Google Scholar] [CrossRef]
- Kim, R.Y.; Horvat, J.C.; Pinkerton, J.W.; Starkey, M.R.; Essilfie, A.T.; Mayall, J.R.; Nair, P.M.; Hansbro, N.G.; Jones, B.; Haw, T.J.; et al. MicroRNA-21 Drives Severe, Steroid-Insensitive Experimental Asthma by Amplifying Phosphoinositide 3-Kinase–Mediated Suppression of Histone Deacetylase 2. J. Allergy Clin. Immunol. 2017, 139, 519–532. [Google Scholar] [CrossRef] [Green Version]
- Spahn, J.D.; Szefler, S.J.; Surs, W.; Doherty, D.E.; Nimmagadda, S.R.; Leung, D.Y. A Novel Action of IL-13: Induction of Diminished Monocyte Glucocorticoid Receptor-Binding Affinity. J. Immunol. 1996, 157, 2654–2659. [Google Scholar]
- Pazdrak, K.; Moon, Y.; Straub, C.; Stafford, S.; Kurosky, A. Eosinophil Resistance to Glucocorticoid-Induced Apoptosis Is Mediated by the Transcription Factor NFIL3. Apoptosis 2016, 21, 421–431. [Google Scholar] [CrossRef] [PubMed]
- McKinley, L.; Alcorn, J.F.; Peterson, A.; DuPont, R.B.; Kapadia, S.; Logar, A.; Henry, A.; Irvin, C.G.; Piganelli, J.D.; Ray, A.; et al. TH17 Cells Mediate Steroid-Resistant Airway Inflammation and Airway Hyperresponsiveness in Mice. J. Immunol. 2008, 181, 4089–4097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chesné, J.; Braza, F.; Mahay, G.; Brouard, S.; Aronica, M.; Magnan, A. IL-17 in Severe Asthma. Where Do We Stand? Am. J. Respir. Crit. Care Med. 2014, 190, 1094–1101. [Google Scholar] [CrossRef]
- Morishima, Y.; Ano, S.; Ishii, Y.; Ohtsuka, S.; Matsuyama, M.; Kawaguchi, M.; Hizawa, N. Th17-Associated Cytokines as a Therapeutic Target for Steroid-Insensitive Asthma. Clin. Dev. Immunol. 2013, 2013, 609395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez-Tello, A.; Semlali, A.; Chakir, J.; Martin, J.G.; Leung, D.Y.; Eidelman, D.H.; Hamid, Q. Induction of Glucocorticoid Receptor-β Expression in Epithelial Cells of Asthmatic Airways by T-Helper Type 17 Cytokines. Clin. Exp. Allergy 2010, 40, 1312–1322. [Google Scholar] [CrossRef]
- Kim, H.Y.; Lee, H.J.; Chang, Y.-J.; Pichavant, M.; Shore, S.A.; Fitzgerald, K.A.; Iwakura, Y.; Israel, E.; Bolger, K.; Faul, J.; et al. Interleukin-17–Producing Innate Lymphoid Cells and the NLRP3 Inflammasome Facilitate Obesity-Associated Airway Hyperreactivity. Nat. Med. 2014, 20, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Brandt, E.B.; Khurana Hershey, G.K. A Combination of Dexamethasone and Anti-IL-17A Treatment Can Alleviate Diesel Exhaust Particle-Induced Steroid Insensitive Asthma. J. Allergy Clin. Immunol. 2016, 138, 924–928.e2. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Jiang, Y.; Yang, X.; Guo, D.; Wang, Y.; Wang, J.; Wang, R.; Wang, C. Lipopolysaccharides Promote a Shift from Th2-Derived Airway Eosinophilic Inflammation to Th17-Derived Neutrophilic Inflammation in an Ovalbumin-Sensitized Murine Asthma Model. J. Asthma 2017, 54, 447–455. [Google Scholar] [CrossRef]
- Hawrylowicz, C.; Richards, D.; Loke, T.-K.; Corrigan, C.; Lee, T. A Defect in Corticosteroid-Induced IL-10 Production in T Lymphocytes from Corticosteroid-Resistant Asthmatic Patients. J. Allergy Clin. Immunol. 2002, 109, 369. [Google Scholar] [CrossRef]
- Hirahara, K.; Mato, N.; Hagiwara, K.; Nakayama, T. The Pathogenicity of IL-33 on Steroid-Resistant Eosinophilic Inflammation via the Activation of Memory-Type ST2+CD4+ T Cells. J. Leukoc. Biol. 2018, 104, 895–901. [Google Scholar] [CrossRef]
- Bray, P.J.; Cotton, R.G.H. Variations of the Human Glucocorticoid Receptor Gene (NR3C1): Pathological and in Vitro Mutations and Polymorphisms. Hum. Mutat. 2003, 21, 557–568. [Google Scholar] [CrossRef]
- Cruz-Topete, D.; Cidlowski, J.A. One Hormone, Two Actions: Anti- and Pro-Inflammatory Effects of Glucocorticoids. Neuroimmunomodulation 2015, 22, 20–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herath, K.H.I.N.M.; Kim, H.J.; Jang, J.-H.; Kim, H.-S.; Kim, H.J.; Jeon, Y.-J.; Jee, Y. Mojabanchromanol Isolated from Sargassum horneri Attenuates Particulate Matter Induced Inflammatory Responses via Suppressing TLR2/4/7-MAPK Signaling in MLE-12 Cells. Mar. Drugs 2020, 18, 355. [Google Scholar] [CrossRef] [PubMed]
- Sanjeewa, K.K.A.; Jayawardena, T.U.; Lee, H.G.; Herath, K.H.I.N.M.; Jee, Y.; Jeon, Y.-J. The Protective Effect of Sargassum horneri against Particulate Matter-Induced Inflammation in Lung Tissues of an in Vivo Mouse Asthma Model. Food Funct. 2019, 10, 7995–8004. [Google Scholar] [CrossRef] [PubMed]
- Herath, K.H.I.N.M.; Kim, H.J.; Mihindukulasooriya, S.P.; Kim, A.; Kim, H.J.; Jeon, Y.-J.; Jee, Y. Sargassum horneri Extract Containing Mojabanchromanol Attenuates the Particulate Matter Exacerbated Allergic Asthma through Reduction of Th2 and Th17 Response in Mice. Environ. Pollut. 2020, 265, 114094. [Google Scholar] [CrossRef]
- Huang, J.; Su, M.; Lee, B.-K.; Kim, M.-J.; Jung, J.H.; Im, D.-S. Suppressive Effect of 4-Hydroxy-2-(4-Hydroxyphenethyl) Isoindoline-1,3-Dione on Ovalbumin-Induced Allergic Asthma. Biomol. Ther. 2018, 26, 539–545. [Google Scholar] [CrossRef]
- Lee, D.-I.; Kang, S.A.; Md, A.; Jeong, U.-C.; Jin, F.; Kang, S.-J.; Lee, J.-Y.; Yu, H.S. Sea Cucumber Lipid-Soluble Extra Fraction Prevents Ovalbumin-Induced Allergic Airway Inflammation. J. Med. Food 2018, 21, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, T.U.; Sanjeewa, K.K.A.; Lee, H.-G.; Nagahawatta, D.P.; Yang, H.-W.; Kang, M.-C.; Jeon, Y.-J. Particulate Matter-Induced Inflammation/Oxidative Stress in Macrophages: Fucosterol from Padina boryana as a Potent Protector, Activated via NF-ΚB/MAPK Pathways and Nrf2/HO-1 Involvement. Mar. Drugs 2020, 18, 628. [Google Scholar] [CrossRef]
- Han, E.-J.; Jayawardena, T.U.; Jang, J.-H.; Fernando, I.P.S.; Jee, Y.; Jeon, Y.-J.; Lee, D.-S.; Lee, J.-M.; Yim, M.-J.; Wang, L.; et al. Sargachromenol Purified from Sargassum horneri Inhibits Inflammatory Responses via Activation of Nrf2/HO-1 Signaling in LPS-Stimulated Macrophages. Mar. Drugs 2021, 19, 497. [Google Scholar] [CrossRef]
- Chen, X.; Ni, L.; Fu, X.; Wang, L.; Duan, D.; Huang, L.; Xu, J.; Gao, X. Molecular Mechanism of Anti-Inflammatory Activities of a Novel Sulfated Galactofucan from Saccharina japonica. Mar. Drugs 2021, 19, 430. [Google Scholar] [CrossRef]
- Chakraborty, K.; Paulraj, R. Sesquiterpenoids with Free-Radical-Scavenging Properties from Marine Macroalga Ulva fasciata Delile. Food Chem. 2010, 122, 31–41. [Google Scholar] [CrossRef]
- Shah, S.A.A.; Hassan, S.S.U.; Bungau, S.; Si, Y.; Xu, H.; Rahman, M.H.; Behl, T.; Gitea, D.; Pavel, F.-M.; Corb Aron, R.A.; et al. Chemically Diverse and Biologically Active Secondary Metabolites from Marine Phylum Chlorophyta. Mar. Drugs 2020, 18, 493. [Google Scholar] [CrossRef]
- Manzoor, Z.; Koo, J.-E.; Ali, I.; Kim, J.-E.; Byeon, S.-H.; Yoo, E.-S.; Kang, H.-K.; Hyun, J.-W.; Lee, N.-H.; Koh, Y.-S. 4-Hydroxy-2,3-Dimethyl-2-Nonen-4-Olide Has an Inhibitory Effect on Pro-Inflammatory Cytokine Production in CpG-Stimulated Bone Marrow-Derived Dendritic Cells. Mar. Drugs 2016, 14, 88. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-J.; Dang, H.-T.; Kang, G.-J.; Yang, E.-J.; Park, S.-S.; Yoon, W.-J.; Jung, J.H.; Kang, H.-K.; Yoo, E.-S. Two Enone Fatty Acids Isolated from Gracilaria verrucosa Suppress the Production of Inflammatory Mediators by Down-Regulating NF-KappaB and STAT1 Activity in Lipopolysaccharide-Stimulated RAW 264.7 Cells. Arch. Pharm. Res. 2009, 32, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.; Ku, S.-K.; Min, G.; Choi, H.; Park, D.H.; Bae, J.-S. Suppressive Effects of Three Diketopiperazines from Marine-Derived Bacteria on Polyphosphate-Mediated Septic Responses. Chem. Biol. Interact. 2016, 257, 61–70. [Google Scholar] [CrossRef]
- Fürstner, A. Chemistry and Biology of Roseophilin and the Prodigiosin Alkaloids: A Survey of the Last 2500 Years. Angew. Chem. Int. Ed. 2003, 42, 3582–3603. [Google Scholar] [CrossRef]
- Kamata, K.; Okamoto, S.; Oka, S.; Kamata, H.; Yagisawa, H.; Hirata, H. Cycloprodigiosin Hydrocloride Suppresses Tumor Necrosis Factor (TNF) Alpha-Induced Transcriptional Activation by NF-KappaB. FEBS Lett. 2001, 507, 74–80. [Google Scholar] [CrossRef] [Green Version]
- Jüttner, F.; Todorova, A.K.; Walch, N.; von Philipsborn, W. Nostocyclamide M: A Cyanobacterial Cyclic Peptide with Allelopathic Activity from Nostoc 31. Phytochemistry 2001, 57, 613–619. [Google Scholar] [CrossRef]
- Anjum, K.; Abbas, S.Q.; Akhter, N.; Shagufta, B.I.; Shah, S.A.A.; Hassan, S.S.U. Emerging Biopharmaceuticals from Bioactive Peptides Derived from Marine Organisms. Chem. Biol. Drug Des. 2017, 90, 12–30. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.S.U.; Anjum, K.; Abbas, S.Q.; Akhter, N.; Shagufta, B.I.; Shah, S.A.A.; Tasneem, U. Emerging Biopharmaceuticals from Marine Actinobacteria. Environ. Toxicol. Pharmacol. 2017, 49, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Anjum, K.; Abbas, S.Q.; Shah, S.A.A.; Akhter, N.; Batool, S.; Hassan, S.S.U. Marine Sponges as a Drug Treasure. Biomol. Ther. 2016, 24, 347–362. [Google Scholar] [CrossRef] [Green Version]
- Hassan, S.S.U.; Shaikh, A.L. Marine Actinobacteria as a Drug Treasure House. Biomed. Pharmacother. 2017, 87, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Kondratyuk, T.P.; Park, E.-J.; Yu, R.; Van Breemen, R.B.; Asolkar, R.N.; Murphy, B.T.; Fenical, W.; Pezzuto, J.M. Novel Marine Phenazines as Potential Cancer Chemopreventive and Anti-Inflammatory Agents. Mar. Drugs 2012, 10, 451–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.-S.; Yoon, C.-S.; Jung, Y.-T.; Yoon, J.-H.; Kim, Y.-C.; Oh, H. Marine-Derived Secondary Metabolite, Griseusrazin A, Suppresses Inflammation through Heme Oxygenase-1 Induction in Activated RAW264.7 Macrophages. J. Nat. Prod. 2016, 79, 1105–1111. [Google Scholar] [CrossRef]
- Kwan, J.C.; Eksioglu, E.A.; Liu, C.; Paul, V.J.; Luesch, H. Grassystatins A-C from Marine Cyanobacteria, Potent Cathepsin E Inhibitors That Reduce Antigen Presentation. J. Med. Chem. 2009, 52, 5732–5747. [Google Scholar] [CrossRef] [Green Version]
- Kozuma, S.; Hirota-Takahata, Y.; Fukuda, D.; Kuraya, N.; Nakajima, M.; Ando, O. Identification and Biological Activity of Ogipeptins, Novel LPS Inhibitors Produced by Marine Bacterium. J. Antibiot. 2017, 70, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Toledo, T.R.; Dejani, N.N.; Monnazzi, L.G.S.; Kossuga, M.H.; Berlinck, R.G.S.; Sette, L.D.; Medeiros, A.I. Potent Anti-Inflammatory Activity of Pyrenocine A Isolated from the Marine-Derived Fungus Penicillium paxilli Ma(G)K. Mediat. Inflamm. 2014, 2014, e767061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; He, J.; Wu, Y.; Du, N.; Li, X.; Ju, J.; Hu, Z.; Umezawa, K.; Wang, L. Isolation and Characterization of New Anti-Inflammatory and Antioxidant Components from Deep Marine-Derived Fungus Myrothecium Sp. Bzo-L062. Mar. Drugs 2020, 18, 597. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, J.; Lin, X.; Zhao, B.; Wei, X.; Li, G.; Kaliaperumal, K.; Liao, S.; Yang, B.; Zhou, X.; et al. Chrysamides A–C, Three Dimeric Nitrophenyl Trans-Epoxyamides Produced by the Deep-Sea-Derived Fungus Penicillium chrysogenum SCSIO41001. Org. Lett. 2016, 18, 3650–3653. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Du, L.; Tang, X.; Jung, S.-Y.; Zheng, B.; Soh, B.Y.; Kim, S.-Y.; Gu, Q.; Park, H. Brevicompanine E Reduces Lipopolysaccharide-Induced Production of Proinflammatory Cytokines and Enzymes in Microglia by Inhibiting Activation of Activator Protein-1 and Nuclear Factor-ΚB. J. Neuroimmunol. 2009, 216, 32–38. [Google Scholar] [CrossRef]
- de Almeida Leone, P.; Redburn, J.; Hooper, J.N.; Quinn, R.J. Polyoxygenated Dysidea Sterols That Inhibit the Binding of [I125] IL-8 to the Human Recombinant IL-8 Receptor Type A. J. Nat. Prod. 2000, 63, 694–697. [Google Scholar] [CrossRef] [PubMed]
- Busserolles, J.; Payá, M.; D’Auria, M.V.; Gomez-Paloma, L.; Alcaraz, M.J. Protection against 2,4,6-Trinitrobenzenesulphonic Acid-Induced Colonic Inflammation in Mice by the Marine Products Bolinaquinone and Petrosaspongiolide M. Biochem. Pharmacol. 2005, 69, 1433–1440. [Google Scholar] [CrossRef] [PubMed]
- Jan, J.-S.; Yang, C.-H.; Wang, M.-H.; Lin, F.-L.; Yen, J.-L.; Hsieh, I.; Khotimchenko, M.; Lee, T.-H.; Hsiao, G. Hirsutanol A Attenuates Lipopolysaccharide-Mediated Matrix Metalloproteinase 9 Expression and Cytokines Production and Improves Endotoxemia-Induced Acute Sickness Behavior and Acute Lung Injury. Mar. Drugs 2019, 17, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, I.-S.; Kwak, J.H.; Pyo, S.; Lee, H.-W.; Kim, A.; Schmitz, F.J. Oscarellin, an Anthranilic Acid Derivative from a Philippine Sponge, Oscarella stillans, as an Inhibitor of Inflammatory Cytokines in Macrophages. J. Nat. Prod. 2017, 80, 149–155. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Lu, M.-C.; Su, J.-H.; Chu, C.-L.; Shiuan, D.; Weng, C.-F.; Sung, P.-J.; Huang, K.-J. Immunomodulatory Effect of Marine Cembrane-Type Diterpenoids on Dendritic Cells. Mar. Drugs 2013, 11, 1336–1350. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.-Y.; Hwang, T.-L.; Lin, M.-R.; Chen, Y.-H.; Chang, Y.-C.; Fang, L.-S.; Wang, W.-H.; Wu, Y.-C.; Sung, P.-J. Carijoside A, a Bioactive Sterol Glycoside from an Octocoral Carijoa Sp. (Clavulariidae). Mar. Drugs 2010, 8, 2014–2020. [Google Scholar] [CrossRef] [Green Version]
- Tseng, W.-R.; Huang, C.-Y.; Tsai, Y.-Y.; Lin, Y.-S.; Hwang, T.-L.; Su, J.-H.; Sung, P.-J.; Dai, C.-F.; Sheu, J.-H. New Cytotoxic and Anti-Inflammatory Steroids from the Soft Coral Klyxum flaccidum. Bioorg. Med. Chem. Lett. 2016, 26, 3253–3257. [Google Scholar] [CrossRef]
- Tsai, Y.-Y.; Huang, C.-Y.; Tseng, W.-R.; Chiang, P.-L.; Hwang, T.-L.; Su, J.-H.; Sung, P.-J.; Dai, C.-F.; Sheu, J.-H. Klyflaccisteroids K–M, Bioactive Steroidal Derivatives from a Soft Coral Klyxum flaccidum. Bioorg. Med. Chem. Lett. 2017, 27, 1220–1224. [Google Scholar] [CrossRef]
- Appleton, D.R.; Chuen, C.S.; Berridge, M.V.; Webb, V.L.; Copp, B.R. Rossinones A and B, Biologically Active Meroterpenoids from the Antarctic Ascidian, Aplidium Species. J. Org. Chem. 2009, 74, 9195–9198. [Google Scholar] [CrossRef]
- Ahmad, T.B.; Rudd, D.; Benkendorff, K.; Mahdi, L.K.; Pratt, K.-A.; Dooley, L.; Wei, C.; Kotiw, M. Brominated Indoles from a Marine Mollusc Inhibit Inflammation in a Murine Model of Acute Lung Injury. PLoS ONE 2017, 12, e0186904. [Google Scholar] [CrossRef] [Green Version]
- Kijjoa, A.; Sawangwong, P. Drugs and Cosmetics from the Sea. Mar. Drugs 2004, 2, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Ankisetty, S.; Khan, S.I.; Avula, B.; Gochfeld, D.; Khan, I.A.; Slattery, M. Chlorinated Didemnins from the Tunicate Trididemnum solidum. Mar. Drugs 2013, 11, 4478–4486. [Google Scholar] [CrossRef] [Green Version]
- Strangman, W.K.; Kwon, H.C.; Broide, D.; Jensen, P.R.; Fenical, W. Potent Inhibitors of Pro-Inflammatory Cytokine Production Produced by a Marine-Derived Bacterium. J. Med. Chem. 2009, 52, 2317–2327. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-Y.; Tseng, Y.-J.; Chokkalingam, U.; Hwang, T.-L.; Hsu, C.-H.; Dai, C.-F.; Sung, P.-J.; Sheu, J.-H. Bioactive Isoprenoid-Derived Natural Products from a Dongsha Atoll Soft Coral Sinularia erecta. J. Nat. Prod. 2016, 79, 1339–1346. [Google Scholar] [CrossRef] [PubMed]
- Herath, K.H.I.N.M.; Kim, H.J.; Lee, J.H.; Je, J.G.; Yu, H.-S.; Jeon, Y.-J.; Kim, H.J.; Jee, Y. Sargassum horneri (Turner) C. Agardh Containing Polyphenols Attenuates Particulate Matter-Induced Inflammatory Response by Blocking TLR-Mediated MYD88-Dependent MAPK Signaling Pathway in MLE-12 Cells. J. Ethnopharmacol. 2021, 265, 113340. [Google Scholar] [CrossRef]
- Jayawardena, T.U.; Asanka Sanjeewa, K.K.; Shanura Fernando, I.P.; Ryu, B.M.; Kang, M.-C.; Jee, Y.; Lee, W.W.; Jeon, Y.-J. Sargassum horneri (Turner) C. Agardh Ethanol Extract Inhibits the Fine Dust Inflammation Response via Activating Nrf2/HO-1 Signaling in RAW 264.7 Cells. BMC Complement. Altern. Med. 2018, 18, 249. [Google Scholar] [CrossRef] [Green Version]
- Sanjeewa, K.K.A.; Jayawardena, T.U.; Kim, S.-Y.; Lee, H.G.; Je, J.-G.; Jee, Y.; Jeon, Y.-J. Sargassum horneri (Turner) Inhibit Urban Particulate Matter-Induced Inflammation in MH-S Lung Macrophages via Blocking TLRs Mediated NF-ΚB and MAPK Activation. J. Ethnopharmacol. 2020, 249, 112363. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.E.; Jung, Y.C.; Jung, I.; Lee, H.-W.; Youn, H.-Y.; Lee, J.S. Anti-Inflammatory Effects of Ethanolic Extract from Sargassum horneri (Turner) C. Agardh on Lipopolysaccharide-Stimulated Macrophage Activation via NF-ΚB Pathway Regulation. Immunol. Investig. 2015, 44, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Na, H.-J.; Moon, P.-D.; Ko, S.-G.; Lee, H.-J.; Jung, H.-A.; Hong, S.-H.; Seo, Y.; Oh, J.-M.; Lee, B.-H.; Choi, B.-W.; et al. Sargassum hemiphyllum Inhibits Atopic Allergic Reaction via the Regulation of Inflammatory Mediators. J. Pharmacol. Sci. 2005, 97, 219–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Ni, L.; Fu, X.; Duan, D.; Xu, J.; Gao, X. A Sulfated Polysaccharide from Saccharina japonica Suppresses LPS-Induced Inflammation Both in a Macrophage Cell Model via Blocking MAPK/NF-ΚB Signal Pathways In Vitro and a Zebrafish Model of Embryos and Larvae In Vivo. Mar. Drugs 2020, 18, 593. [Google Scholar] [CrossRef]
- Wang, L.; Yang, H.-W.; Ahn, G.; Fu, X.; Xu, J.; Gao, X.; Jeon, Y.-J. In Vitro and In Vivo Anti-Inflammatory Effects of Sulfated Polysaccharides Isolated from the Edible Brown Seaweed, Sargassum fulvellum. Mar. Drugs 2021, 19, 277. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Xie, B.; Du, J.; Zhang, A.; Hao, J.; Wang, S.; Wang, J.; Cao, J. The Anti-Inflammatory Effect and Structure of EPCP1-2 from Crypthecodinium cohnii via Modulation of TLR4-NF-ΚB Pathways in LPS-Induced RAW 264.7 Cells. Mar. Drugs 2017, 15, 376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macagno, A.; Molteni, M.; Rinaldi, A.; Bertoni, F.; Lanzavecchia, A.; Rossetti, C.; Sallusto, F. A Cyanobacterial LPS Antagonist Prevents Endotoxin Shock and Blocks Sustained TLR4 Stimulation Required for Cytokine Expression. J. Exp. Med. 2006, 203, 1481–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorgersen, E.B.; Macagno, A.; Rossetti, C.; Mollnes, T.E. Cyanobacterial LPS Antagonist (CyP)-a Novel and Efficient Inhibitor of Escherichia Coli LPS-Induced Cytokine Response in the Pig. Mol. Immunol. 2008, 45, 3553–3557. [Google Scholar] [CrossRef] [PubMed]
- Molteni, M.; Bosi, A.; Rossetti, C. The Effect of Cyanobacterial LPS Antagonist (CyP) on Cytokines and Micro-RNA Expression Induced by Porphyromonas gingivalis LPS. Toxins 2018, 10, 290. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.-N.; Lee, E.-H.; Kim, H.-M. Spirulina Platensis Inhibits Anaphylactic Reaction. Life Sci. 1997, 61, 1237–1244. [Google Scholar] [CrossRef]
- Appel, K.; Munoz, E.; Navarrete, C.; Cruz-Teno, C.; Biller, A.; Thiemann, E. Immunomodulatory and Inhibitory Effect of Immulina®, and Immunloges® in the Ig-E Mediated Activation of RBL-2H3 Cells. A New Role in Allergic Inflammatory Responses. Plants 2018, 7, 13. [Google Scholar] [CrossRef] [Green Version]
- Rod-in, W.; Monmai, C.; Lee, S.; Jung, S.-K.; You, S.; Park, W.J. Anti-Inflammatory Effects of Lipids Extracted from Arctoscopus japonicus Eggs on LPS-Stimulated RAW264.7 Cells. Mar. Drugs 2019, 17, 580. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Lee, S.-H.; Le, Q.-T.; Kim, M.-M.; Kim, S.-K. Anti-Allergic Effects of Phlorotannins on Histamine Release via Binding Inhibition between IgE and FcεRI. J. Agric. Food Chem. 2008, 56, 12073–12080. [Google Scholar] [CrossRef]
- Niu, S.; Liu, Q.; Xia, J.-M.; Xie, C.-L.; Luo, Z.-H.; Shao, Z.; Liu, G.; Yang, X.-W. Polyketides from the Deep-Sea-Derived Fungus Graphostroma Sp. MCCC 3A00421 Showed Potent Antifood Allergic Activities. J. Agric. Food Chem. 2018, 66, 1369–1376. [Google Scholar] [CrossRef]
- Gao, Y.-Y.; Liu, Q.-M.; Liu, B.; Xie, C.-L.; Cao, M.-J.; Yang, X.-W.; Liu, G.-M. Inhibitory Activities of Compounds from the Marine Actinomycete williamsia Sp. MCCC 1A11233 Variant on IgE-Mediated Mast Cells and Passive Cutaneous Anaphylaxis. J. Agric. Food Chem. 2017, 65, 10749–10756. [Google Scholar] [CrossRef]
- Lee, D.-I.; Park, M.-K.; Kang, S.A.; Choi, J.-H.; Kang, S.-J.; Lee, J.-Y.; Yu, H.S. Preventive Intra Oral Treatment of Sea Cucumber Ameliorate OVA-Induced Allergic Airway Inflammation. Am. J. Chin. Med. 2016, 44, 1663–1674. [Google Scholar] [CrossRef] [Green Version]
- Onodera, K.; Konishi, Y.; Taguchi, T.; Kiyoto, S.; Tominaga, A. Peridinin from the Marine Symbiotic Dinoflagellate, Symbiodinium Sp., Regulates Eosinophilia in Mice. Mar. Drugs 2014, 12, 1773–1787. [Google Scholar] [CrossRef] [Green Version]

| Degree of Glucocorticoid Resistance | Asthma Phenotypes | Pathobiologic Features |
|---|---|---|
| Severe corticosteroid resistance | Obesity-related asthma | Absence of Th2 specific response Increased oxidative stress |
| Neutrophilic asthma | Increased Th-17 response (increased IL-8, neutrophilia) | |
| Late-onset eosinophilic asthma | Increased IL-5 Eosinophilia | |
| Moderate corticosteroid resistance | Early-onset allergic asthma | Increased Th2 specific response Presence of antigen-specific IgE |
| Exercise-induced asthma | Increased Th2 specific response Increased mast cells degranulation Increased CysLTs |
| Molecular Targets | Pharmacological Effect | References |
|---|---|---|
| Decrease in activity of MAPK | Decrease in GR phosphorylation Increased ratio of GR-α to GR-β isoforms | [43,44,48,49,50,51,52,53,54,55] |
| Increase of activity of HDAC | Decrease in GR phosphorylation Increased ratio of GR-α to GR-β isoforms | [43,44,52,53,54,56] |
| Decrease in activation of JNK | Decrease in GR phosphorylation Increase in GR-α nuclear translocation | [68] [62] |
| Nitric oxide decrease | Decrease in nitrosylation of GR at HSP90 (chaperone binding site) | [69,84] |
| Decrease inactivation of NF-κB | Increase in GR-α nuclear translocation | [62,63,64,67] |
| Decrease in oxidative stress | Multiple | [22] |
| Downregulation of Th2 | ||
| IL-4 | Increase in GR-α expression and nuclear translocation Increase in GR binding affinity in T-cells Decrease in GR phosphorylation | [40,41,42,43,45,46,47,48] |
| IL-5 | Increased GR binding affinity | [71,72] |
| IL-13 | Decrease in GR phosphorylation Increased GR binding affinity | [45,46,47,48] [71] |
| Downregulation of non-Th2 | ||
| IL-17 | Decrease in GR-β expression | [42,76,77] |
| IL-23 | Decrease in GR-β expression | [42] |
| IFN-γ | Decreased GR phosphorylation and stimulation of GR nuclear translocation Increase in GR-α nuclear translocation (through downregulation of NF-κB) | [66] [67] |
| TNF-α | Increase in GR-α nuclear translocation (through downregulation of NF-κB) Decrease in GR-α phosphorylation at Ser226 and the inhibition of GRE-binding (through downregulation of JNK) | [67] [68] |
| IL-33 | Decreased GR phosphorylation | [81,82] |
| IL-1β | Unknown | [70] |
| Inhibition of inflammatory response shift: Th2 to Th17 † | Decreased GR phosphorylation and stimulate GR nuclear translocation Increase in GR-α nuclear translocation (through downregulation of NF-κB) Decrease in GR-α phosphorylation at Ser226 and the inhibition of GRE-binding (through downregulation of JNK) | [49,66] |
| Compound | Specie | Origin | Class | Molecular Targets | References | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Decrease Activity of MAPK | Increase of Activity of HDAC | Decrease Activation of JNK | Nitric Oxide Decrease | Decrease Activation of NF-κB | Decrease Oxidative Stress | Downregulation of Th2 | Downregulation of Non-Th2 | |||||||||||||
| IL-4 | IL-5 | IL-13 | IL-17 | IL-23 | IFN-γ | TNF-α | IL-33 | IL-1β | Inhibition of Inflammatory Response Shift: Th2 to Th17 † | |||||||||||
| Simple compounds | ||||||||||||||||||||
| Fucosterol | Padina boryana | brown algae | phytosterol | x | x | x | x | x | [89] | |||||||||||
| Mojabanchromanol | Sargassum horneri | brown algae | chromanol | x | x | x | x | x | [84] | |||||||||||
| Sargachromenol | Sargassum horneri | brown algae | chromenol | x | x | x | [90] | |||||||||||||
| Fucoidan | Saccharina japonica | brown algae | polysaccharides | x | x | x | x | [91] | ||||||||||||
| 3,4,5,5-Tetramethyl-4-(3′-oxopentyl)-2-cyclohexen-1-one | Ulva fasciata Deliles | green algae | sesquiterpenoids | x | [92,93] | |||||||||||||||
| 4-hydroxy-2,3-dimethyl-2-nonen-4-olide | Ulva pertusa | green algae | extract | x | [94] | |||||||||||||||
| (E)-9-Oxooctadec-10-enoic-acid and (E)-10-Oxooctadec-8-enoic-acid | Gracilaria verrucosa | red algae | fatty acids | x | x | x | [95] | |||||||||||||
| cyclo(L-Pro-D-Val), cyclo(L-Pro-L-Tyr), cyclo(L-pro-D-Leu) | Bacillus sp. HC001, Piscicoccus sp. 12L081 | bacteria | diketopiperazine | x | x | x | [96] | |||||||||||||
| Cycloprodigiosin | Pseudoalteromonas dentrificans | bacteria | prodigiosin | x | x | [97,98] | ||||||||||||||
| C-Phycocyanin | Nostoc Muscorum Cyanobacteria | bacteria | Polypeptide | x | [99,100] | |||||||||||||||
| Phenazines 1,2 | Streptomyces sp. | bacteria | aromatic secondary metabolites | x | x | x | [101,102,103,104] | |||||||||||||
| Griseusrazin A | Streptomyces griseus | bacteria | pyrazine-type molecules | x | x | x | x | [103,105] | ||||||||||||
| G rassystatin A | Lyngbya confervoides | bacteria | linear decadepsipeptide | x | [106] | |||||||||||||||
| Ogipeptins A-D | Pseudoalteromonas sp. SANK 71903 | bacteria | cyclic peptides | x | x | [107] | ||||||||||||||
| pyrenocine A | Penicillium paxilli | fungus | phytotoxins | x | x | [108] | ||||||||||||||
| (−)-1S-myrothecol and (+)-1R-myrothecol | Myrothecium sp. BZO-L062 | fungus | alkylresorcinol | x | [109] | |||||||||||||||
| Chrysamides A–C | Penicillium chrysogenum SCSIO41001 | fungus | dimeric nitrophenyl trans-epoxyamides | x | [110] | |||||||||||||||
| Brevicompanine E | Penicillium sp. | fungus | diketopiperazine derivatives | x | x | x | x | x | x | [111] | ||||||||||
| Polyoxygenated Sterols & bolinaquinone | Dysidea sp. | fungus | sterols | x | [112,113] | |||||||||||||||
| Hirsutanol A | Chondrostereum sp. NTOU4196 | fungus | Sesquiterpene compound | x | x | x | [114] | |||||||||||||
| Oscarellin | Oscarella stillans | sponge | anthralinic acids | x | x | x | [115] | |||||||||||||
| Lobocrassin B | Lobophytum crassum | coral | diterpenoids | x | x | [116] | ||||||||||||||
| Carijoside A | Carijoa sp. | soft coral | Steroid glycoside | x | [117] | |||||||||||||||
| Klyflaccisteroid J | Klyxum flaccidum | soft coral | steroidal derivatives | x | [118] | |||||||||||||||
| Klyflaccisteroid K-M | Klyxum flaccidum | soft coral | steroidal derivatives | x | [119] | |||||||||||||||
| Rossinones A & B | Aplidium speciesascidian | marine animals | Terpene derived metabolite | x | [120] | |||||||||||||||
| 6-bromoisatin | Dicathais orbita | gastropod mollusc | brominated indole derivatives | x | x | [121] | ||||||||||||||
| Didemnin B | Trididemnum solidum | ascidia | depsipeptides | x | x | [122,123] | ||||||||||||||
| Splenocins A-I | Streptomyces sp. | bacteria | depsipeptides | x | x | x | x | [124] | ||||||||||||
| Sinulerectol A & B | Sinularia erecta | soft coral | cembranoid | x | [125] | |||||||||||||||
| Complex composition | ||||||||||||||||||||
| Sargassum horneri extract | Sargassum horneri | brown algae | extract | x | x | x | x | x | [126] | |||||||||||
| Sargassum horneri (Turner) ethanol extract | Sargassum horneri (Turner) C. Agardh | brown algae | extract | x | x | x | x | x | [127] | |||||||||||
| Sargassum horneri (Turner) ethanol extract | Sargassum horneri | brown algae | extract | x | x | x | x | [128] | ||||||||||||
| Sargassum horneri ethanol extract | Sargassum horneri | brown alga | ethanol extract | x | x | x | x | x | [86] | |||||||||||
| Sargassum horneri (Turner) C. Agardh ethanol extract | Sargassum horneri (Turner) C. Agardh | brown algae | ethanolic extract | x | x | x | x | x | [129] | |||||||||||
| Sargassum hemiphyllum methanol extract | Sargassum hemiphyllum | brown alga | methanol extract | x | x | [130] | ||||||||||||||
| Sulfated polysaccharide | Saccharina japonica | brown algae | polysaccharides | x | x | x | x | x | x | [131] | ||||||||||
| Sulfated polysaccharide | Sargassum fulvellum | brown algae | polysaccharides | x | x | x | x | [132] | ||||||||||||
| Exopolysaccharide (EPCP1-2) | Crypthecodinium cohnii | microalgae | polysaccharide | x | x | [133] | ||||||||||||||
| Cyanobacterial lipopolysaccharide | Oscillatoria planktothrix FP1 | bacteria | lipopolysaccharides | x | x | [134,135,136] | ||||||||||||||
| Spirulina extract | Arthrospira platensis (Spirulina) | bacteria | lipoproteins | x | x | [137,138] | ||||||||||||||
| Arctoscopus japonicus egg extracted lipids | Arctoscopus japonicus | fish | fatty acid | x | x | x | x | x | x | [139] | ||||||||||
| Apostichopus japonicus extract | Apostichopus japonicus | marine animals | extract | x | x | x | x | [88] | ||||||||||||
| Compound | Chemical Structure |
|---|---|
| fucosterol | 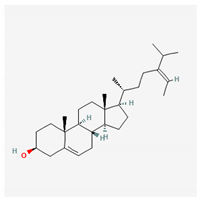 |
| mojabanchromanol | 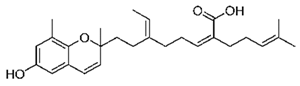 |
| sargachromenol | 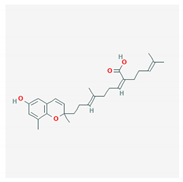 |
| fucoidan |  |
| 3,4,5,5-Tetramethyl-4-(3′-oxopentyl)-2-cyclohexen-1-one |  |
| 4-hydroxy-2,3-dimethyl-2-nonen-4-olide | 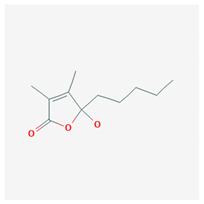 |
| (E)-9-Oxooctadec-10-enoic-acid and (E)-10-Oxooctadec-8-enoic-acid | 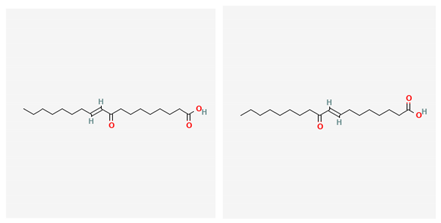 |
| cyclo(L-Pro-D-Val), cyclo(L-Pro-L-Tyr), cyclo(L-pro-D-Leu) | 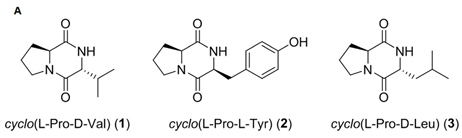 |
| cycloprodigiosin |  |
| C-phycocyanin |  |
| phenazines 1,2 | 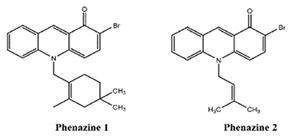 |
| griseusrazin A | 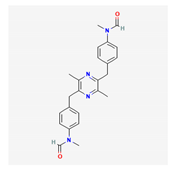 |
| grassystatin A | 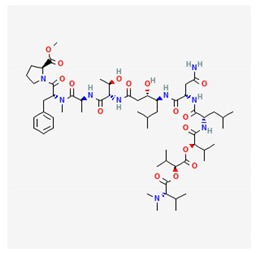 |
| ogipeptins A-D | 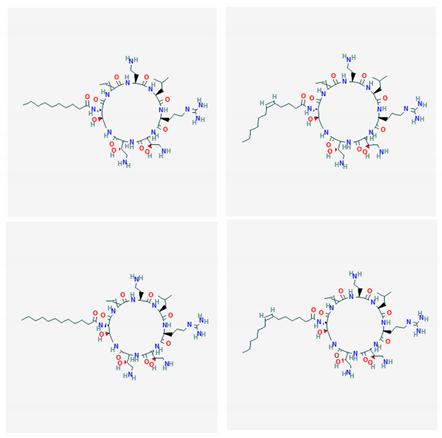 |
| pyrenocine A |  |
| (−)-1S-myrothecol and (+)-1R-myrothecol | 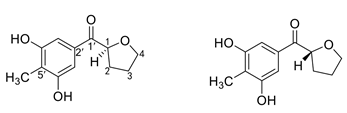 |
| chrysamides A–C | 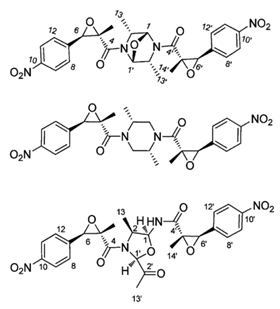 |
| brevicompanine E | 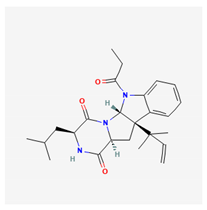 |
| polyoxygenated dysidea sterols | 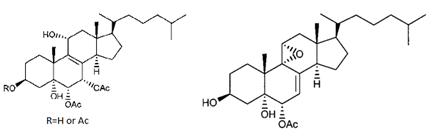 |
| bolinaquinone | 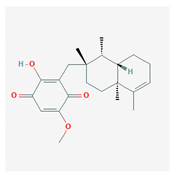 |
| hirsutanol A |  |
| oscarellin | 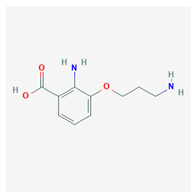 |
| lobocrassin B | 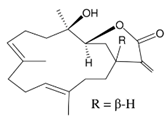 |
| carijoside A | 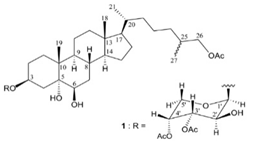 |
| klyflaccisteroid J | 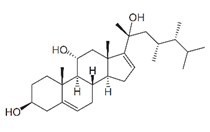 |
| klyflaccisteroid K-M | 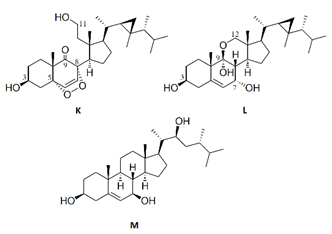 |
| rossinones A & B | 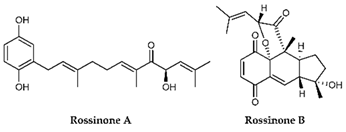 |
| 6-bromoisatin | 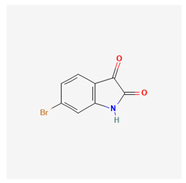 |
| didemnin B | 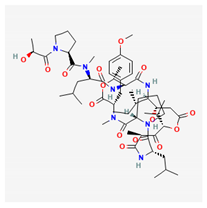 |
| splenocins A-I |  |
| sinulerectol A & B | 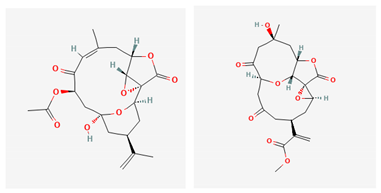 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghiciuc, C.M.; Vicovan, A.G.; Stafie, C.S.; Antoniu, S.A.; Postolache, P. Marine-Derived Compounds for the Potential Treatment of Glucocorticoid Resistance in Severe Asthma. Mar. Drugs 2021, 19, 586. https://doi.org/10.3390/md19110586
Ghiciuc CM, Vicovan AG, Stafie CS, Antoniu SA, Postolache P. Marine-Derived Compounds for the Potential Treatment of Glucocorticoid Resistance in Severe Asthma. Marine Drugs. 2021; 19(11):586. https://doi.org/10.3390/md19110586
Chicago/Turabian StyleGhiciuc, Cristina Mihaela, Andrei Gheorghe Vicovan, Celina Silvia Stafie, Sabina Antonela Antoniu, and Paraschiva Postolache. 2021. "Marine-Derived Compounds for the Potential Treatment of Glucocorticoid Resistance in Severe Asthma" Marine Drugs 19, no. 11: 586. https://doi.org/10.3390/md19110586
APA StyleGhiciuc, C. M., Vicovan, A. G., Stafie, C. S., Antoniu, S. A., & Postolache, P. (2021). Marine-Derived Compounds for the Potential Treatment of Glucocorticoid Resistance in Severe Asthma. Marine Drugs, 19(11), 586. https://doi.org/10.3390/md19110586