
Laminarin, a Major Polysaccharide in Stramenopiles

 ,
, 
Abstract
:1. Introduction
2. General Features of Laminarin in Stramenopiles
2.1. Laminarin Structure and Composition
2.2. Laminarin Content
2.3. Bioactive Potentials of Laminarin
3. Biosynthesis and Catabolism Pathways of Laminarin in Stramenopiles
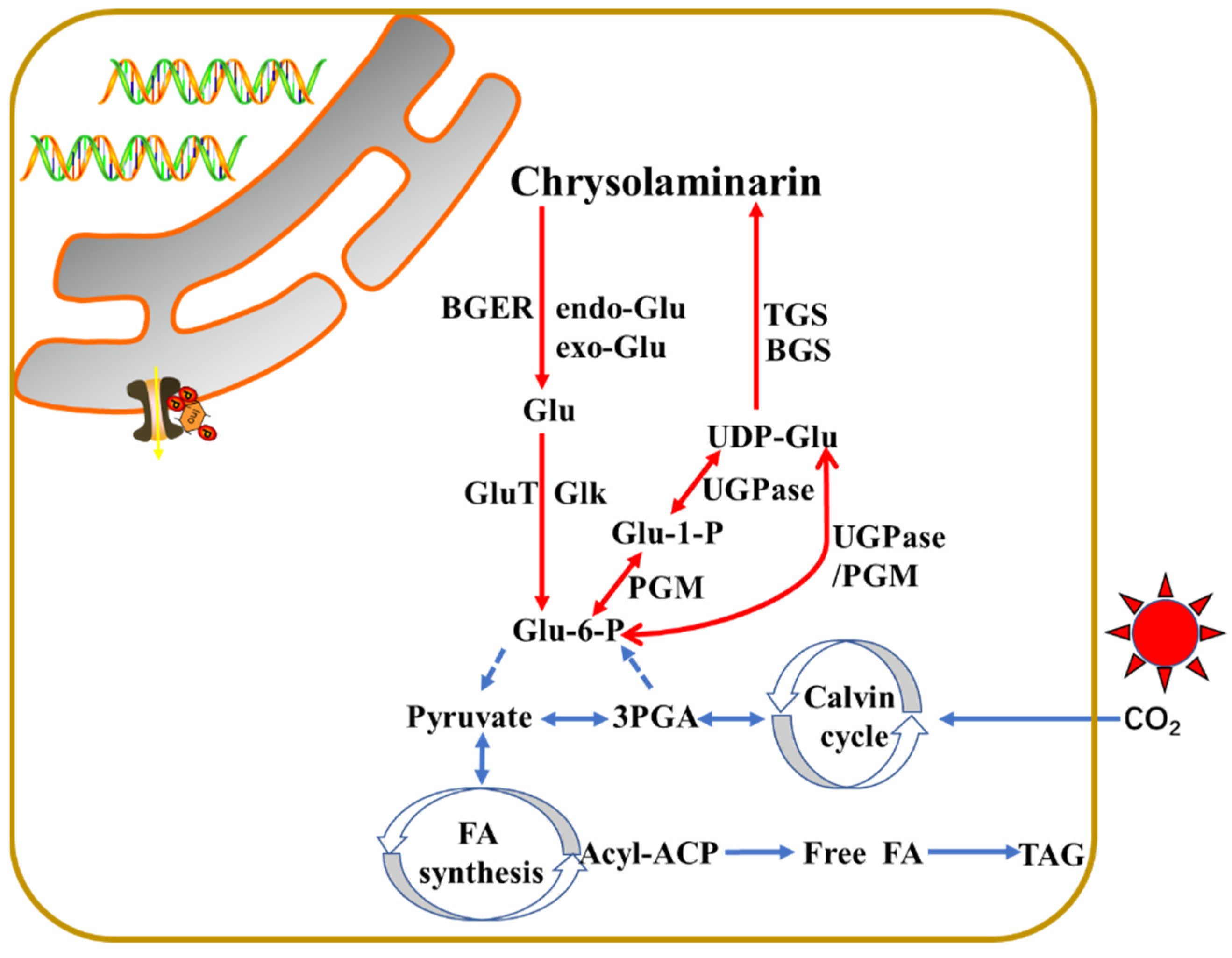
3.1. The Proposed Pathways of Laminarin
3.2. Manipulation of Key Genes Involved in Chrysolaminarin Biosynthesis
3.3. Diel Regulation of Chrysolaminarin
4. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Mathieu-Rivet, E.; Mati-Baouche, N.; Walet-Balieu, M.L.; Lerouge, P.; Bardor, M. N- and o-glycosylation pathways in the microalgae polyphyletic group. Front. Plant Sci. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Adl, S.M.; Bass, D.; Lane, C.E.; Lukeš, J.; Schoch, C.L.; Smirnov, A.; Agatha, S.; Berney, C.; Brown, M.W.; Burki, F.; et al. Revisions to the classification, nomenclature, and diversity of eukaryotes. J. Eukaryot. Microbiol. 2019, 66, 4–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irisarri, I.; Strassert, J.F.H.; Burki, F. Phylogenomic insights into the origin of primary plastids. Syst. Biol. 2021, 4, 1–36. [Google Scholar]
- Strassert, J.F.H.; Irisarri, I.; Williams, T.A.; Burki, F. A molecular timescale for eukaryote evolution with implications for the origin of red algal-derived plastids. Nat. Commun. 2021, 12, 1–13. [Google Scholar]
- Faktorová, D.; Nisbet, R.E.R.; Fernández Robledo, J.A.; Casacuberta, E.; Sudek, L.; Allen, A.E.; Ares, M.; Aresté, C.; Balestreri, C.; Barbrook, A.C.; et al. Genetic tool development in marine protists: Emerging model organisms for experimental cell biology. Nat Methods 2020, 17, 481–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falciatore, A.; Jaubert, M.; Bouly, J.P.; Bailleul, B.; Mock, T. Diatom molecular research comes of age: Model species for studying phytoplankton biology and diversity. Plant Cell. 2020, 32, 547–572. [Google Scholar] [CrossRef] [Green Version]
- Bäumgen, M.; Dutschei, T.; Bornscheuer, U.T. Marine polysaccharides: Occurrence, enzymatic degradation and utilization. ChemBioChem 2021, 22, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.; Tebben, J.; Coffinet, S.; Wiltshire, K.; Iversen, M.H.; Harder, T.; Hinrichs, K.U.; Hehemann, J.H. Laminarin is a major molecule in the marine carbon cycle. Proc. Natl. Acad. Sci. USA 2020, 117, 6599–6607. [Google Scholar] [CrossRef] [Green Version]
- Vogler, B.W.; Brannum, J.; Chung, J.W.; Seger, M.; Posewitz, M.C. Characterization of the Nannochloropsis gaditana storage carbohydrate: A 1,3-beta glucan with limited 1,6-branching. Algal Res. 2018, 36, 152–158. [Google Scholar] [CrossRef]
- Ma, M.; Li, Y.; Chen, J.; Wang, F.; Yuan, L.; Li, Y.; Zhang, B.; Ye, D.; Han, D.; Jin, H.; et al. High-cell-density cultivation of the flagellate alga Poterioochromonas malhamensis for biomanufacturing the water-soluble β-1,3-glucan with multiple biological activities. Bioresour. Technol. 2021, 337, 125447. [Google Scholar] [CrossRef]
- Gügi, B.; Le Costaouec, T.; Burel, C.; Lerouge, P.; Helbert, W.; Bardor, M. Diatom-specific oligosaccharide and polysaccharide structures help to unravel biosynthetic capabilities in diatoms. Mar. Drugs 2015, 13, 5993–6018. [Google Scholar] [CrossRef] [Green Version]
- Kamoun, S. Molecular genetics of pathogenic oomycetes. Eukaryot. Cell 2003, 2, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Vogler, B.W.; Ashford, A.; Posewitz, M.C. CRISPR/Cas9 disruption of glucan synthase in Nannochloropsis gaditana attenuates accumulation of β-1,3-glucose oligomers. Algal Res. 2021, 58, 1–7. [Google Scholar] [CrossRef]
- Butler, T.; Kapoore, R.V.; Vaidyanathan, S. Phaeodactylum tricornutum: A diatom cell factory. Trends Biotechnol. 2020, 38, 606–622. [Google Scholar] [CrossRef] [PubMed]
- Daboussi, F.; Leduc, S.; Maréchal, A.; Dubois, G.; Guyot, V.; Perez-Michaut, C.; Amato, A.; Falciatore, A.; Juillerat, A.; Beurdeley, M.; et al. Genome engineering empowers the diatom Phaeodactylum tricornutum for biotechnology. Nat. Commun. 2014, 5, 1–7. [Google Scholar] [CrossRef]
- Størseth, T.R.; Hansen, K.; Reitan, K.I.; Skjermo, J. Structural characterization of β-D-(1→3)-glucans from different growth phases of the marine diatoms Chaetoceros mülleri and Thalassiosira weissflogii. Carbohydr. Res. 2005, 340, 1159–1164. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Gao, B.; Li, A.; Xiong, J.; Ao, Z.; Zhang, C. Preliminary characterization, antioxidant properties and production of chrysolaminarin from marine diatom Odontella aurita. Mar. Drugs 2014, 12, 4883–4897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirokawa, Y.; Fujiwara, S.; Suzuki, M.; Akiyama, T.; Sakamoto, M.; Kobayashi, S.; Tsuzuki, M. Structural and physiological studies on the storage β-polyglucan of haptophyte Pleurochrysis haptonemofera. Planta 2008, 227, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Menshova, R.V.; Ermakova, S.P.; Anastyuk, S.D.; Isakov, V.V.; Dubrovskaya, Y.V.; Kusaykin, M.I.; Um, B.H.; Zvyagintseva, T.N. Structure, enzymatic transformation and anticancer activity of branched high molecular weight laminaran from brown alga Eisenia bicyclis. Carbohydr. Polym. 2014, 99, 101–109. [Google Scholar] [CrossRef]
- Hentati, F.; Tounsi, L.; Djomdi, D.; Pierre, G.; Delattre, C.; Ursu, A.V.; Fendri, I.; Abdelkafi, S.; Michaud, P. Bioactive polysaccharides from seaweeds. Molecules 2020, 25, 3152. [Google Scholar] [CrossRef]
- Lomartire, S.; Marques, J.C.; Gonçalves, A.M.M. An overview to the health benefits of seaweeds consumption. Mar. Drugs 2021, 19, 341. [Google Scholar] [CrossRef]
- Rioux, L.E.; Turgeon, S.L.; Beaulieu, M. Structural characterization of laminaran and galactofucan extracted from the brown seaweed Saccharina longicruris. Phytochemistry 2010, 71, 1586–1595. [Google Scholar] [CrossRef]
- Ermakova, S.; Men’shova, R.; Vishchuk, O.; Kim, S.M.; Um, B.H.; Isakov, V.; Zvyagintseva, T. Water-soluble polysaccharides from the brown alga Eisenia bicyclis: Structural characteristics and antitumor activity. Algal Res. 2013, 2, 51–58. [Google Scholar] [CrossRef]
- Gao, B.; Chen, A.; Zhang, W.; Li, A.; Zhang, C. Co-production of lipids, eicosapentaenoic acid, fucoxanthin, and chrysolaminarin by Phaeodactylum tricornutum cultured in a flat-plate photobioreactor under varying nitrogen conditions. J. Ocean Univ. China 2017, 16, 916–924. [Google Scholar] [CrossRef]
- Caballero, M.A.; Jallet, D.; Shi, L.; Rithner, C.; Zhang, Y.; Peers, G. Quantification of chrysolaminarin from the model diatom Phaeodactylum tricornutum. Algal Res. 2016, 20, 180–188. [Google Scholar] [CrossRef] [Green Version]
- Paulsen, B.S.; Myklestad, S. Structural studies of the reserve glucan produced by the marine diatom Skeletonema costatum (grev.) Cleve. Carbohydr. Res. 1978, 62, 386–388. [Google Scholar] [CrossRef]
- McConville, M.J.; Bacic, A.; Clarke, A.E. Structural studies of chrysolaminaran from the ice diatom Stauroneis amphioxys (Gregory). Carbohydr. Res. 1986, 153, 330–333. [Google Scholar] [CrossRef]
- Wustman, B.A.; Gretz, M.R.; Hoagland, K.D. Extracellular matrix assembly in diatoms (Bacillariophyceae): I. A model of adhesives based on chemical characterization and localization of polysaccharides from the marine diatom Achnanthes longipes and other diatoms. Plant Physiol. 1997, 113, 1059–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiovitti, A.; Bacic, A.; Burke, J.; Wetherbee, R. Heterogeneous xylose-rich glycans are associated with extracellular glycoproteins from the biofouling diatom Craspedostauros australis (Bacillariophyceae). Eur. J. Phycol. 2003, 38, 351–360. [Google Scholar] [CrossRef] [Green Version]
- Alekseeva, S.A.; Shevchenko, N.M.; Kusaǐkin, M.I.; Ponomorenko, L.P.; Isakov, V.V.; Zviagintseva, T.N.; Likhoshvai, E.V. Polysaccharides of diatoms occurring in Lake Baikal. Appl. Biochem. Microbiol. 2005, 41, 185–191. [Google Scholar] [CrossRef]
- Størseth, T.R.; Kirkvold, S.; Skjermo, J.; Reitan, K.I. A branched β-d-(1→3,1→6)-glucan from the marine diatom Chaetoceros debilis (Bacillariophyceae) characterized by NMR. Carbohydr. Res. 2006, 341, 2108–2114. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.F.; Li, D.W.; Chen, T.T.; Bin Hao, T.; Balamurugan, S.; Yang, W.D.; Liu, J.S.; Li, H.Y. Overproduction of bioactive algal chrysolaminarin by the critical carbon flux regulator phosphoglucomutase. Biotechnol. J. 2019, 14, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, M.; Manandhar-Shrestha, K.; Abbriano, R. Effects of chrysolaminarin synthase knockdown in the diatom Thalassiosira pseudonana: Implications of reduced carbohydrate storage relative to green algae. Algal. Res. 2017, 23, 66–77. [Google Scholar] [CrossRef]
- Schreiber, V.; Dersch, J.; Puzik, K.; Bäcker, O.; Liu, X.; Stork, S.; Schulz, J.; Heimerl, T.; Klingl, A.; Zauner, S. The central vacuole of the diatom Phaeodactylum tricornutum: Identification of new vacuolar membrane proteins and of a functional di-leucine-based targeting motif. Protist 2017, 168, 271–282. [Google Scholar] [CrossRef]
- Costa, A.M.S.; Rodrigues, J.M.M.; Pérez-Madrigal, M.M.; Dove, A.P.; Mano, J.F. Modular functionalization of laminarin to create value-added naturally derived macromolecules. J. Am. Chem. Soc. 2020, 142, 19689–19697. [Google Scholar] [CrossRef]
- Maheshwari, G.; Sowrirajan, S.; Joseph, B. Extraction and isolation of β-glucan from grain sources—A review. J. Food Sci. 2017, 82, 1535–1545. [Google Scholar] [CrossRef] [Green Version]
- Abdullahi, A.S.; Underwood, G.J.C.; Gretz, M.R. Extracellular matrix assembly in diatoms (Bacillariophyceae). V. Environmental effects on polysaccharide synthesis in the model diatom, Phaeodactylum tricornutum. J. Phycol. 2006, 42, 363–378. [Google Scholar] [CrossRef]
- Zhang, H.; Row, K.H. Extraction and separation of polysaccharides from Laminaria japonica by size-exclusion chromatography. J. Chromatogr. Sci. 2015, 53, 498–502. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Ren, Y.; Li, Y.; Fan, M.; Qian, H.; Wang, L.; Wu, G.; Zhang, H.; Qi, X.; Xu, M.; et al. Physiological functionalities and mechanisms of β-glucans. Trends Food Sci. Technol. 2019, 88, 57–66. [Google Scholar] [CrossRef]
- Myklestad, S.M. Production, chemical structure, metabolism, and biological function of the (1→3)-linked, β3-D-glucans in diatoms. Biol. Oceanogr. 1989, 6, 313–326. [Google Scholar]
- Skogen, M.; Winge, P.; Brembu, T.; Vadstein, O.; Bones, A.M. Gene regulation of carbon fixation, storage, and utilization in the diatom Phaeodactylum tricornutum acclimated to light/dark cycles. Plant Physiol. 2013, 161, 1034–1048. [Google Scholar]
- Jensen, E.L.; Yangüez, K.; Carrière, F.; Gontero, B. Storage compound accumulation in diatoms as response to elevated CO2 concentration. Biology 2020, 9, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granum, E.; Myklestad, S.M. Mobilization of β-1,3-glucan and biosynthesis of amino acids induced by NH4+ addition to N-limited cells of the marine diatom Skeletonema costatum (Bacillariophyceae). J. Phycol. 2001, 37, 772–782. [Google Scholar] [CrossRef]
- Granum, E.; Kirkvold, S.; Myklestad, S.M. Cellular and extracellular production of carbohydrates and amino acids by the marine diatom Skeletonema costatum: Diel variations and effects of N depletion. Mar. Ecol. Prog. Ser. 2002, 242, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Roessler, P.G. Changes in the activities of various lipid and carbohydrate biosynthetic enzymes in the diatom Cyclotella cryptica in response to silicon deficiency. Arch. Biochem. Biophys. 1988, 267, 521–528. [Google Scholar] [CrossRef]
- Kusaikin, M.I.; Ermakova, S.P.; Shevchenko, N.M.; Isakov, V.V.; Gorshkov, A.G.; Vereshchagin, A.L.; Grachev, M.A.; Zvyagintseva, T.N. Structural characteristics and antitumor activity of a new chrysolaminaran from the diatom alga Synedra acus. Chem. Nat. Compd. 2010, 46, 1–4. [Google Scholar] [CrossRef]
- Park, H.K.; Kim, I.H.; Kim, J.; Nam, T.J. Induction of apoptosis and the regulation of ErbB signaling by laminarin in HT-29 human colon cancer cells. Int. J. Mol. Med. 2013, 32, 291–295. [Google Scholar] [CrossRef] [Green Version]
- Pradhan, B.; Nayak, R.; Patra, S.; Jit, B.P.; Ragusa, A.; Jena, M. Bioactive metabolites from marine algae as potent pharmacophores against oxidative stress-associated human diseases: A comprehensive review. Molecules 2020, 26, 37. [Google Scholar] [CrossRef] [PubMed]
- Sanjeewa, K.K.A.; Lee, J.S.; Kim, W.S.; Jeon, Y.J. The potential of brown-algae polysaccharides for the development of anticancer agents: An update on anticancer effects reported for fucoidan and laminaran. Carbohydr. Polym. 2017, 177, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Haferkamp, I.; Lepetit, B.; Molchanova, M.; Hou, S.; Jeblick, W.; Bártulos, C.R.; Kroth, P.G. Reduced vacuolar β-1,3-glucan synthesis affects carbohydrate metabolism as well as plastid homeostasis and structure in Phaeodactylum tricornutum. Proc. Natl. Acad. Sci. USA 2018, 115, 4791–4796. [Google Scholar] [CrossRef] [Green Version]
- Poliner, E.; Panchy, N.; Newton, L.; Wu, G.; Lapinsky, A.; Bullard, B.; Zienkiewicz, A.; Benning, C.; Shiu, S.H.; Farré, E.M. Transcriptional coordination of physiological responses in Nannochloropsis oceanica CCMP1779 under light/dark cycles. Plant J. 2015, 83, 1097–1113. [Google Scholar] [CrossRef]
- Huang, W.; Río Bártulos, C.; Kroth, P.G. Diatom vacuolar 1,6-β-transglycosylases can functionally complement the respective yeast mutants. J. Eukaryot. Microbiol. 2016, 63, 536–546. [Google Scholar] [CrossRef] [Green Version]
- Kroth, P.G.; Chiovitti, A.; Gruber, A.; Martin-Jezequel, V.; Mock, T.; Parker, M.S.; Stanley, M.S.; Kaplan, A.; Caron, L.; Weber, T.; et al. A model for carbohydrate metabolism in the diatom Phaeodactylum tricornutum deduced from comparative whole genome analysis. PLoS ONE 2008, 3, e1426. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Hempel, F.; Stork, S.; Bolte, K.; Moog, D.; Heimerl, T.; Maier, U.G.; Zauner, S. Addressing various compartments of the diatom model organism Phaeodactylum tricornutum via sub-cellular marker proteins. Algal Res. 2016, 20, 249–257. [Google Scholar] [CrossRef]
- Hildebrand, M.; Abbriano, R.M.; Polle, J.E.W.; Traller, J.C.; Trentacoste, E.M.; Smith, S.R.; Davis, A.K. Metabolic and cellular organization in evolutionarily diverse microalgae as related to biofuels production. Curr. Opin. Chem. Biol. 2013, 17, 506–514. [Google Scholar] [CrossRef]
- Zhu, B.H.; Shi, H.P.; Yang, G.P.; Lv, N.N.; Yang, M.; Pan, K.H. Silencing UDP-glucose pyrophosphorylase gene in Phaeodactylum tricornutum affects carbon allocation. N. Biotechnol. 2016, 33, 237–244. [Google Scholar] [CrossRef]
- McClung, C.R. The genetics of plant clocks. Adv. Genet. 2011, 74, 105–139. [Google Scholar] [PubMed]
- Litchman, E.; Klausmeier, C.A.; Bossard, P. Phytoplankton nutrient competition under dynamic light regimes. Limnol. Oceanogr. 2004, 49, 1457–1462. [Google Scholar] [CrossRef] [Green Version]
- Vårum, K.M.; Østgaard, K.; Grimsrud, K. Diurnal rhythms in carbohydrate metabolism of the marine diatom Skeletonema costatum (Grev.) Cleve. J. Exp. Mar. Bio. Ecol. 1986, 102, 249–256. [Google Scholar] [CrossRef]
- Jallet, D.; Caballero, M.A.; Gallina, A.A.; Youngblood, M.; Peers, G. Photosynthetic physiology and biomass partitioning in the model diatom Phaeodactylum tricornutum grown in a sinusoidal light regime. Algal Res. 2016, 18, 51–60. [Google Scholar] [CrossRef] [Green Version]
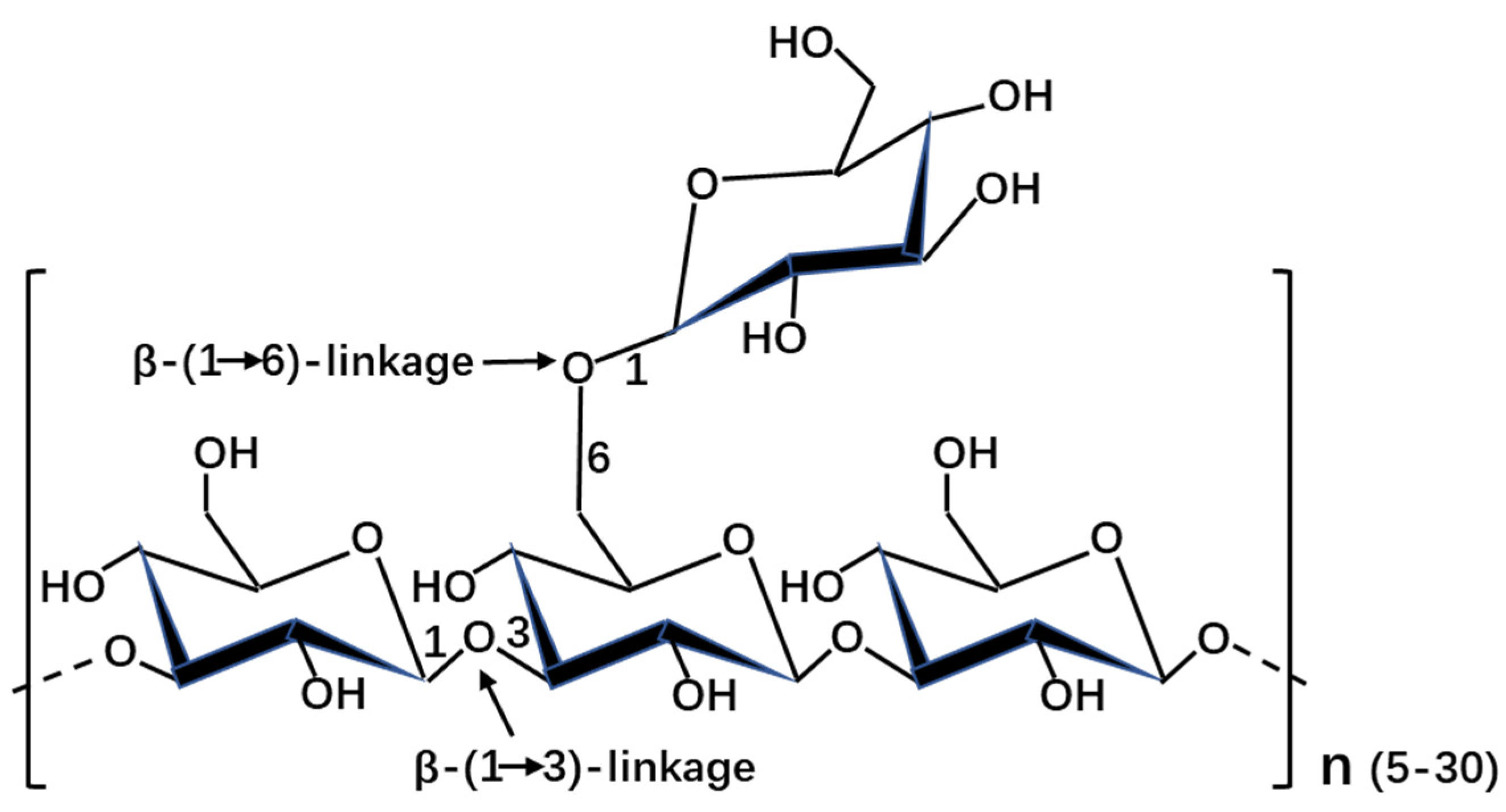

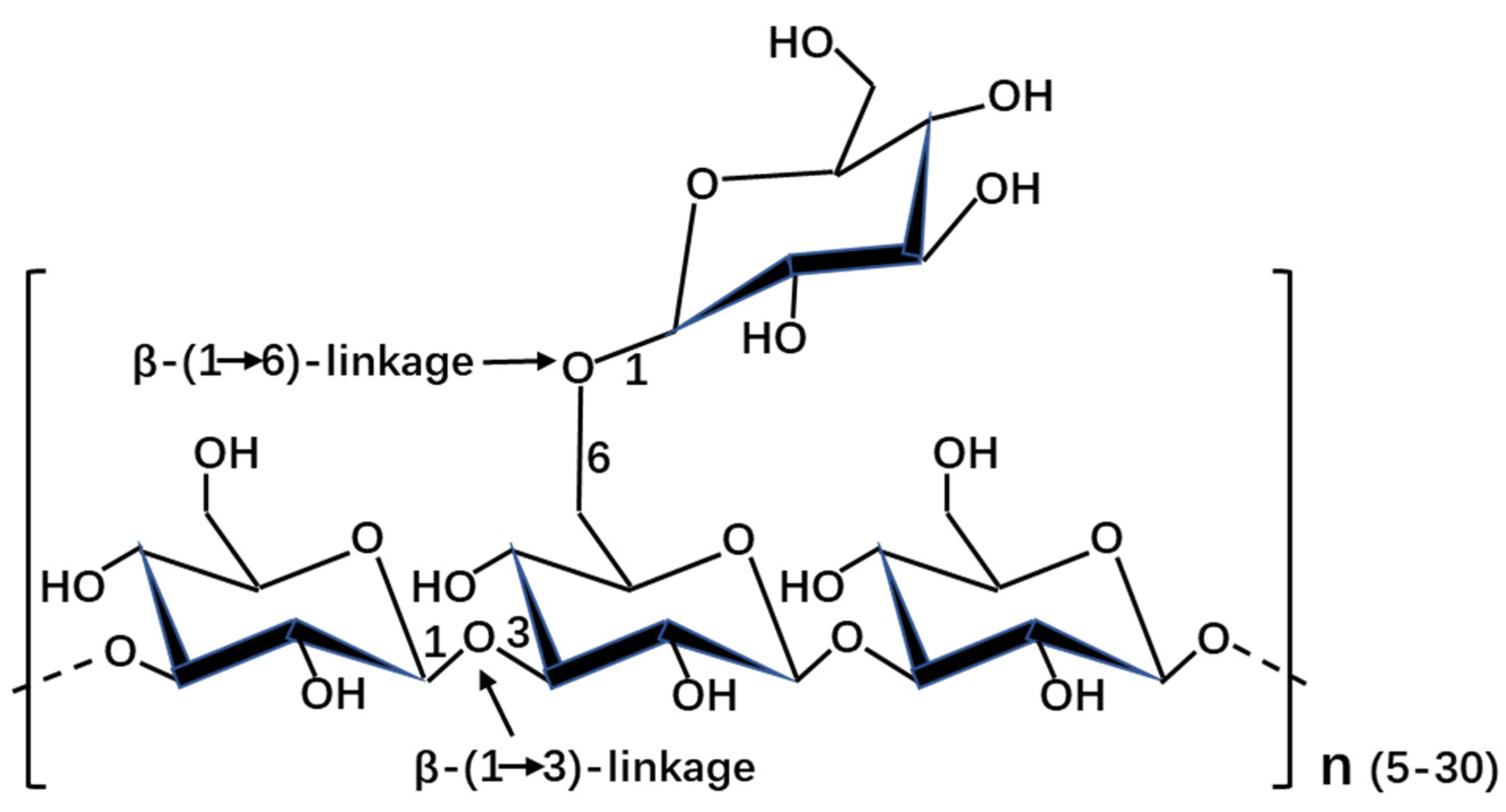{kind=link}
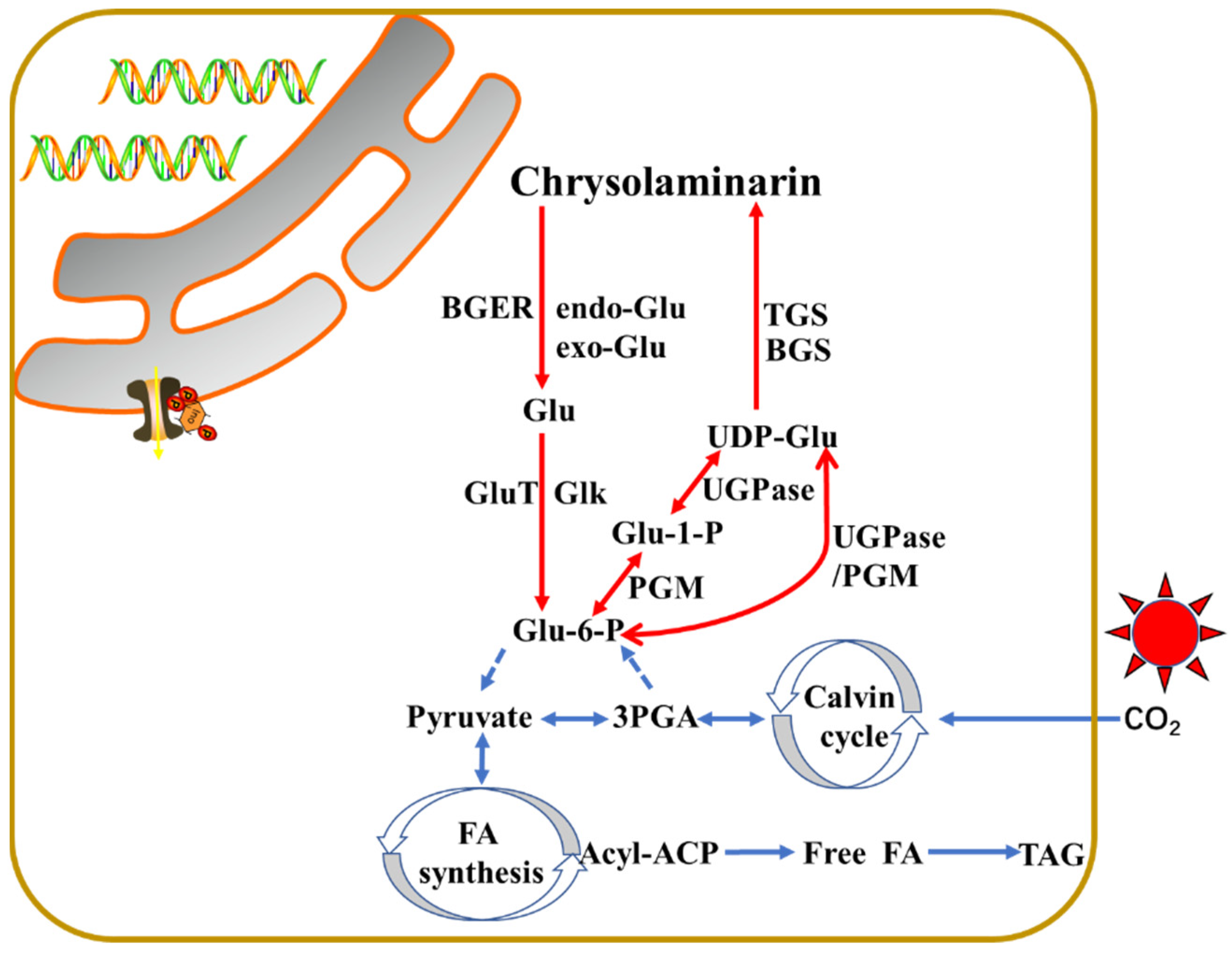{kind=link}
| Species | Mw/DP | DB | Branches | Yield | Reference |
|---|---|---|---|---|---|
| Phaeodactylum tricornutum | nd | β-1,6 | 14% | [24] | |
| Phaeodactylum tricornutum | DP 17 | 0.015 | β-1,6 | [25] | |
| Skeletonema costatum | 6–13kDa | β-1,6 β-1,2 | 32% | [26] | |
| Stauroneis amphixys | 4kDa DP~24 | β-1,6 β-1,2 | nd | [27] | |
| Achnanthes longipes | nd | β-1,6 β-1,2 | nd | [28] | |
| Craspedostauros australis | >10 kDa | β-1,6 | nd | [29] | |
| Aulacoseira baicalensis | 3–5 kDa | nd | 0.9% | [11] | |
| Stephanodiscus meyerii | 40 kDa | 0.053 | β-1,6 β-1,3 | 0.5% | [30] |
| Stephanodiscus meyerii | 2–6 kDa | 0.25 | β-1,6 β-1,3 | 0.4% | [30] |
| Aulacoseira baicalensis | nd | 0.11 | β-1,6 β-1,3 mannitol | 0.6% | [11] |
| Chaetoceros muelleri | DP 22–24 | 0.006–0.009 | β-1,6 β-1,3 | nd | [16] |
| Thalassiosira weissflogii | DP 5–13 | No branch | nd | [16] | |
| Chaetoceros debilis | 4.9 kDa, DP 30 | β-1,6 | 10% | [31] | |
| Odontella aurita | 7.75 kDa, | β-1,6 | 15.09% | [17] | |
| a Nannochloropsis gaditana | DP 8 | 0.028–0.105 | β-1,6 mannitol | 0.5% | [9,13] |
| b Nannochloropsis gaditana | DP 8.1–9.2 | 0.0036–0.0071 | β-1,6 mannitol | 0.5% | [9,13] |
| Poterioochromonas malhamensis | 16.7 kDa | β-1,6 | 55% | [10] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Yang, J.; Du, H.; Aslam, M.; Wang, W.; Chen, W.; Li, T.; Liu, Z.; Liu, X. Laminarin, a Major Polysaccharide in Stramenopiles. Mar. Drugs 2021, 19, 576. https://doi.org/10.3390/md19100576
Chen J, Yang J, Du H, Aslam M, Wang W, Chen W, Li T, Liu Z, Liu X. Laminarin, a Major Polysaccharide in Stramenopiles. Marine Drugs. 2021; 19(10):576. https://doi.org/10.3390/md19100576
Chicago/Turabian StyleChen, Jichen, Jianchao Yang, Hong Du, Muhmmad Aslam, Wanna Wang, Weizhou Chen, Tangcheng Li, Zhengyi Liu, and Xiaojuan Liu. 2021. "Laminarin, a Major Polysaccharide in Stramenopiles" Marine Drugs 19, no. 10: 576. https://doi.org/10.3390/md19100576
APA StyleChen, J., Yang, J., Du, H., Aslam, M., Wang, W., Chen, W., Li, T., Liu, Z., & Liu, X. (2021). Laminarin, a Major Polysaccharide in Stramenopiles. Marine Drugs, 19(10), 576. https://doi.org/10.3390/md19100576