In Vitro Chemopreventive Potential of Phlorotannins-Rich Extract from Brown Algae by Inhibition of Benzo[a]pyrene-Induced P2X7 Activation and Toxic Effects

 , , and
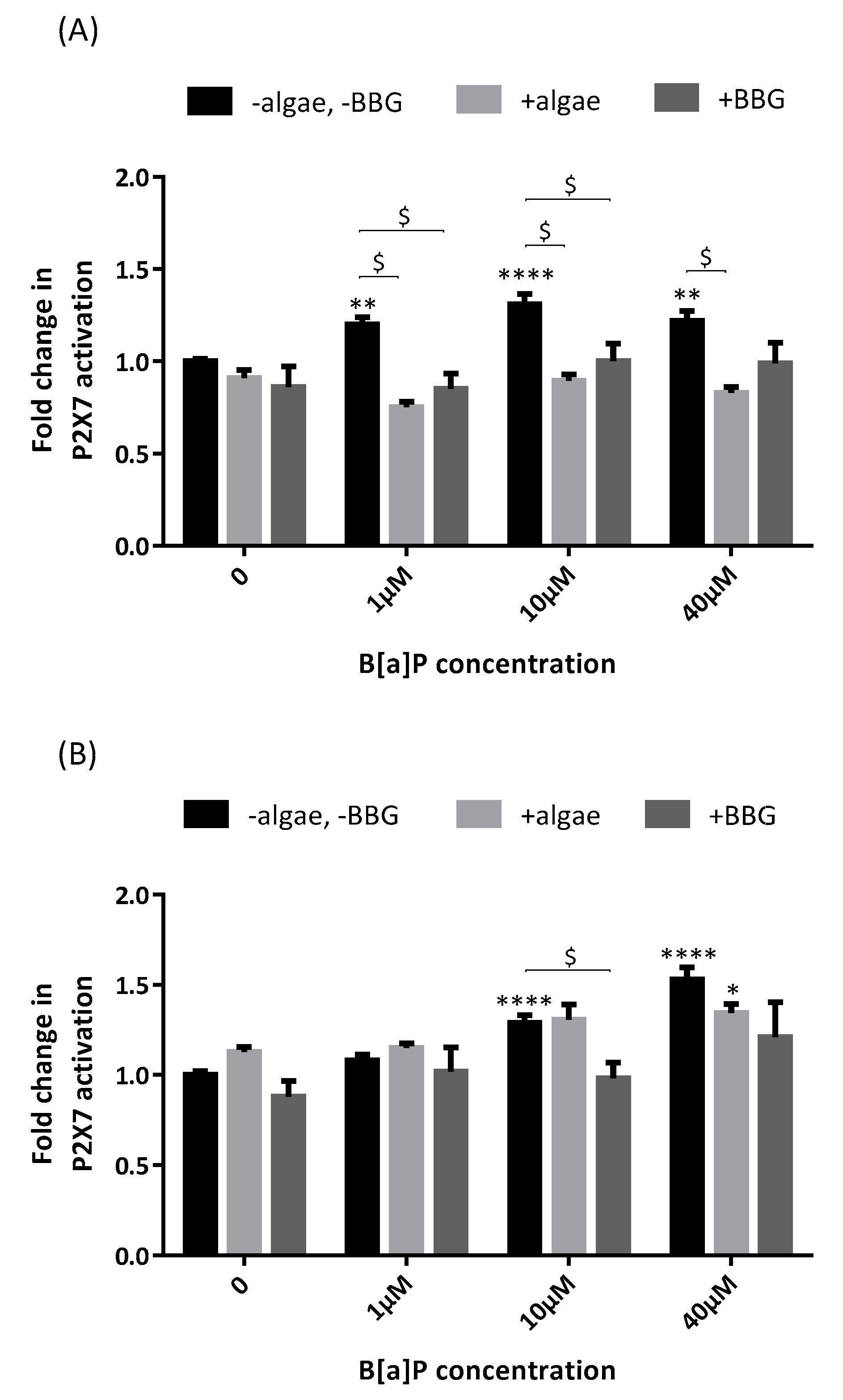
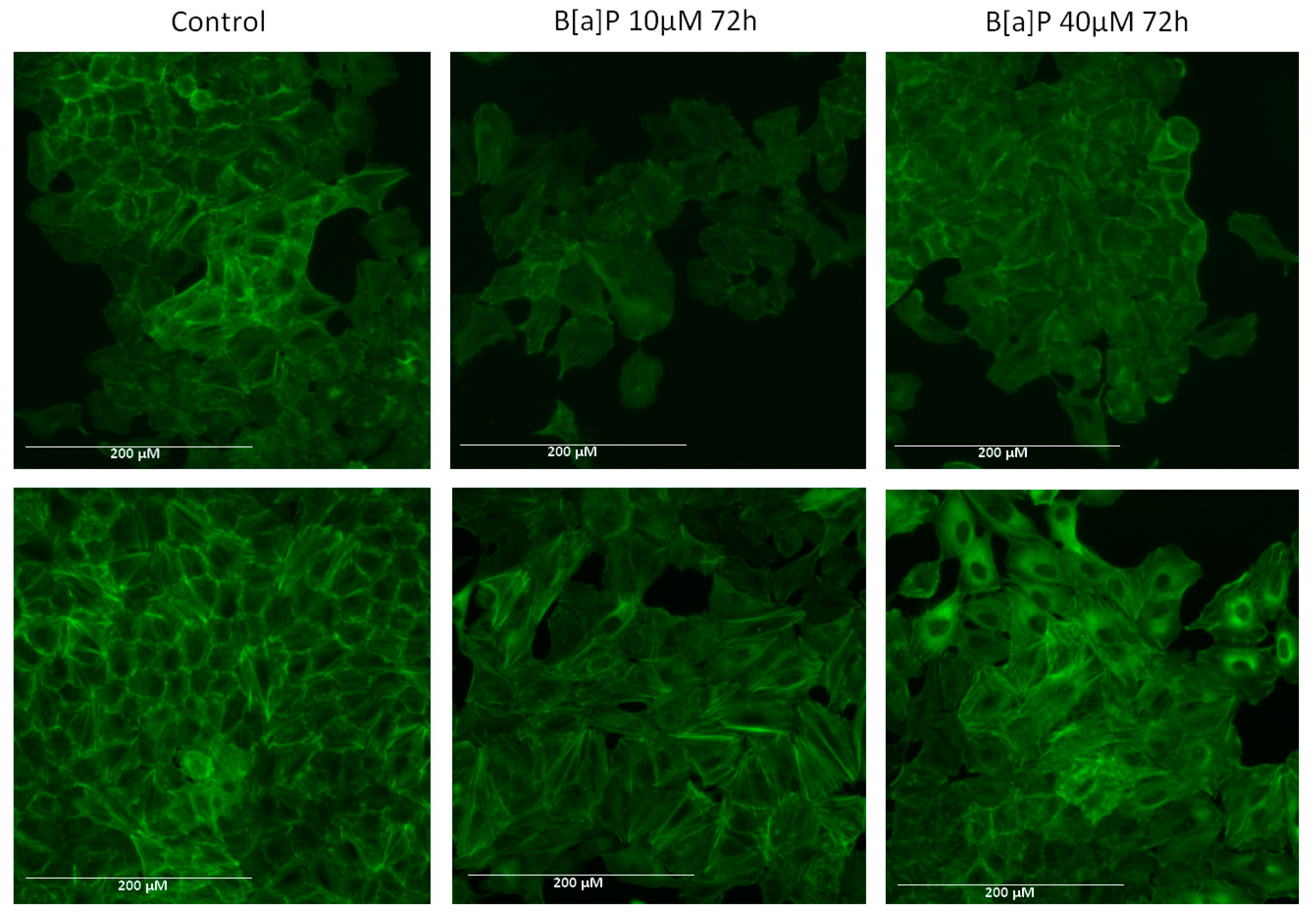
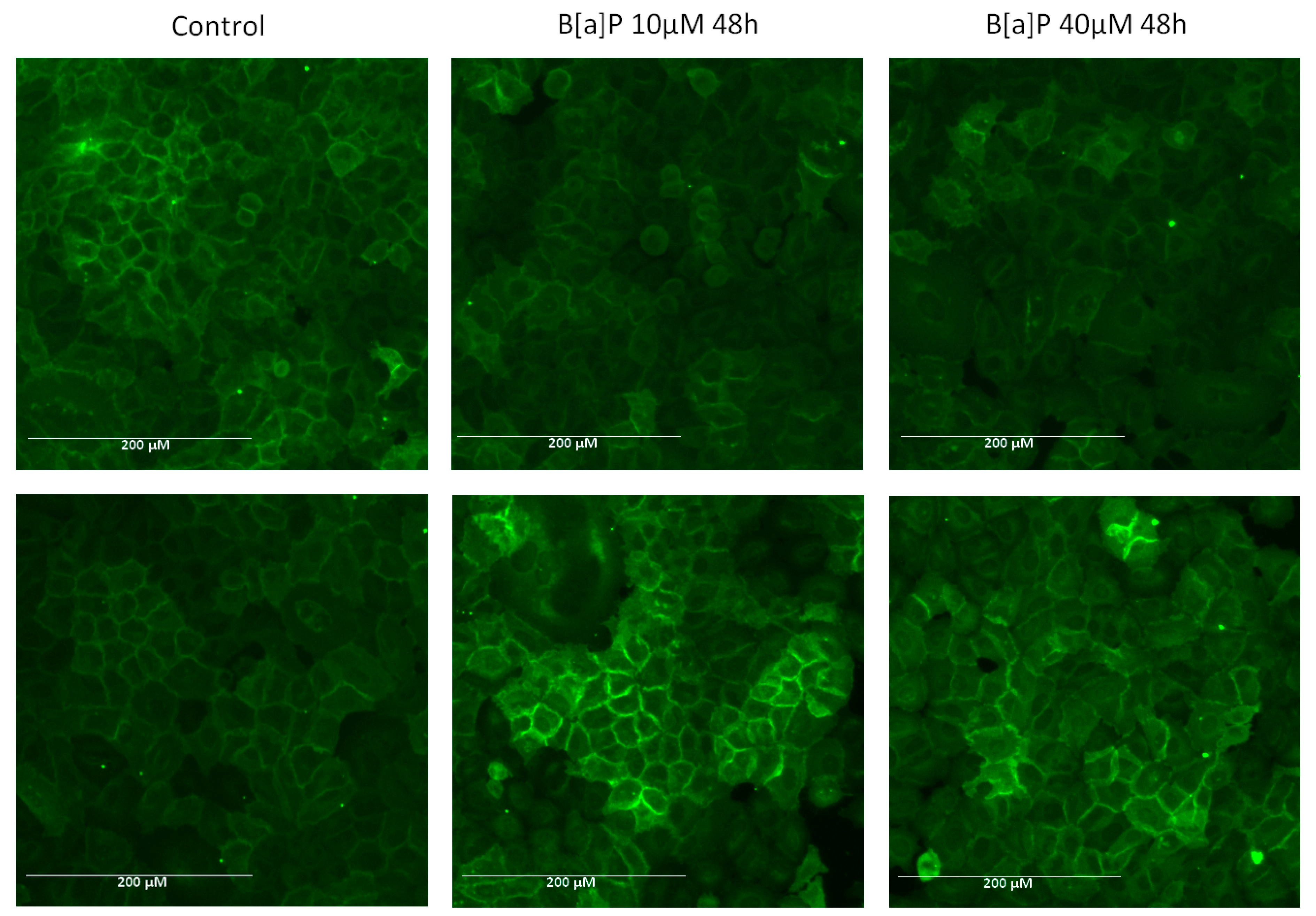
, , and 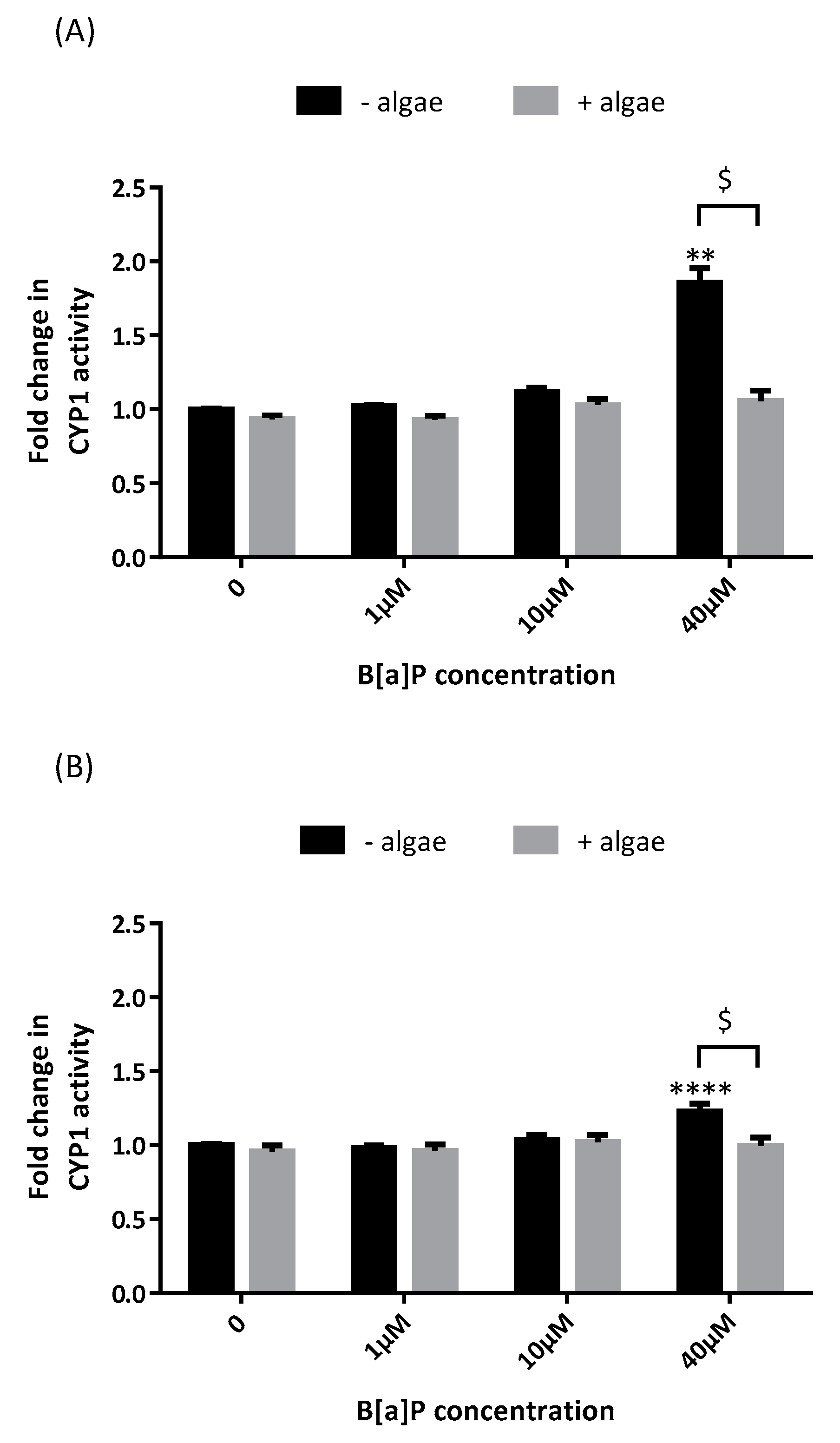{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
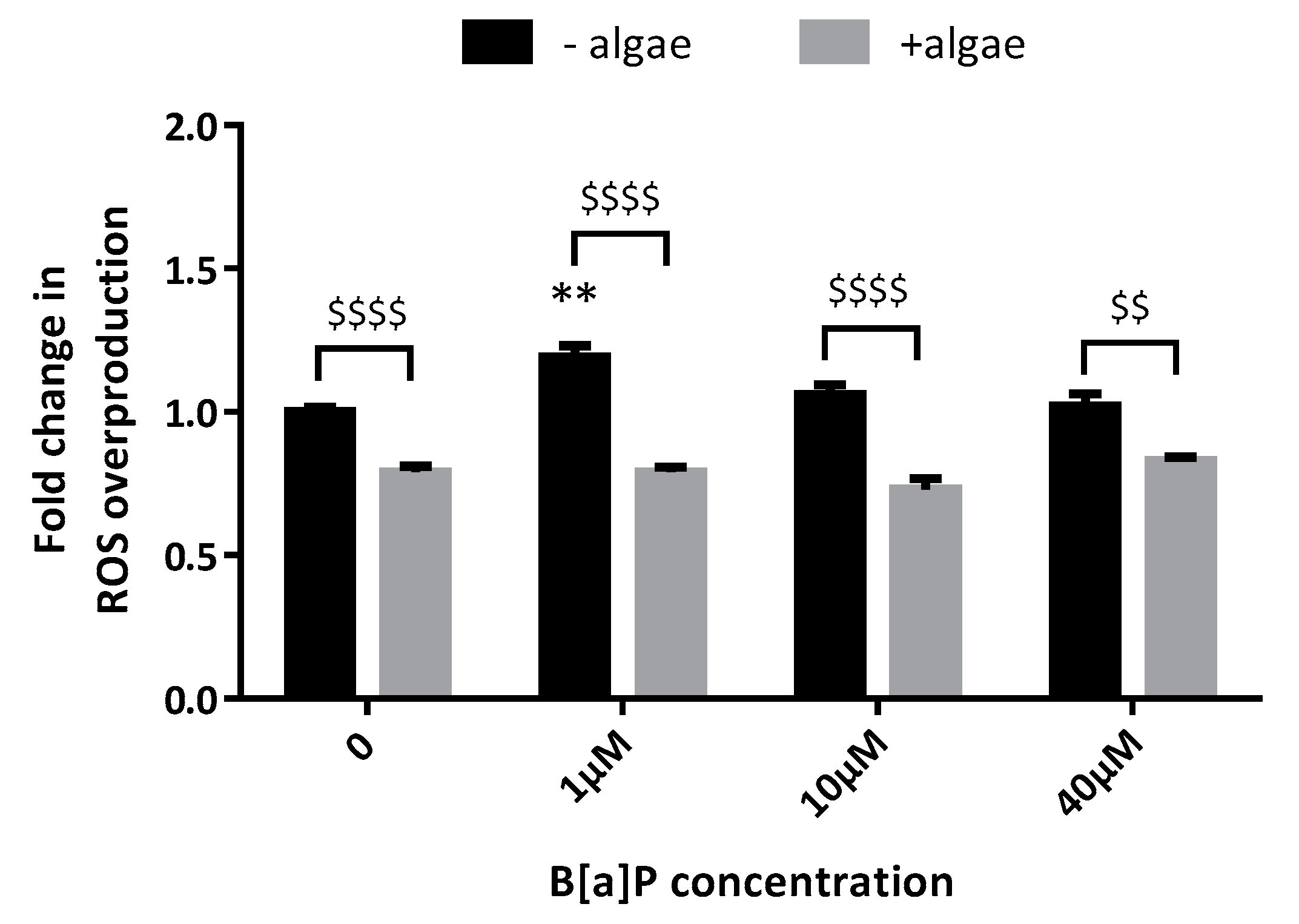
Abstract
Share and Cite
Dutot, M.; Olivier, E.; Fouyet, S.; Magny, R.; Hammad, K.; Roulland, E.; Rat, P.; Fagon, R. In Vitro Chemopreventive Potential of Phlorotannins-Rich Extract from Brown Algae by Inhibition of Benzo[a]pyrene-Induced P2X7 Activation and Toxic Effects. Mar. Drugs 2021, 19, 34. https://doi.org/10.3390/md19010034
Dutot M, Olivier E, Fouyet S, Magny R, Hammad K, Roulland E, Rat P, Fagon R. In Vitro Chemopreventive Potential of Phlorotannins-Rich Extract from Brown Algae by Inhibition of Benzo[a]pyrene-Induced P2X7 Activation and Toxic Effects. Marine Drugs. 2021; 19(1):34. https://doi.org/10.3390/md19010034
Chicago/Turabian StyleDutot, Mélody, Elodie Olivier, Sophie Fouyet, Romain Magny, Karim Hammad, Emmanuel Roulland, Patrice Rat, and Roxane Fagon. 2021. "In Vitro Chemopreventive Potential of Phlorotannins-Rich Extract from Brown Algae by Inhibition of Benzo[a]pyrene-Induced P2X7 Activation and Toxic Effects" Marine Drugs 19, no. 1: 34. https://doi.org/10.3390/md19010034
APA StyleDutot, M., Olivier, E., Fouyet, S., Magny, R., Hammad, K., Roulland, E., Rat, P., & Fagon, R. (2021). In Vitro Chemopreventive Potential of Phlorotannins-Rich Extract from Brown Algae by Inhibition of Benzo[a]pyrene-Induced P2X7 Activation and Toxic Effects. Marine Drugs, 19(1), 34. https://doi.org/10.3390/md19010034