Sea Cucumber Derived Type I Collagen: A Comprehensive Review




Abstract
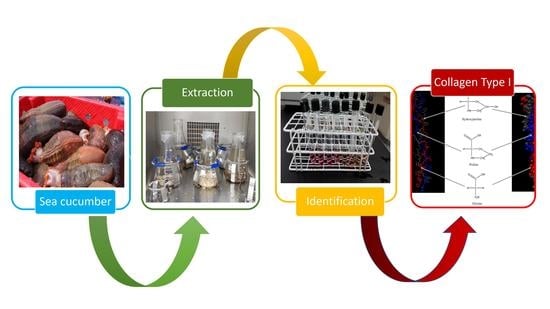
1. Introduction
2. Definition and History of Collagen
2.1. Basic Structure and Synthesis
2.2. Nomenclature, Types, and Classifications
3. Sources of Collagen
3.1. Land Animal By-Products
3.2. Marine Organisms
3.3. Sea Cucumber as A Source of Collagen
4. Characteristics and Properties of Collagen Type I
4.1. Thermal Stability
4.2. Enzymatic Resistance and Digestion
4.3. Isoelectric Point of Collagen
4.4. Bioactive Properties of Sea Cucumber Collagen
5. Industrial Applications
5.1. Biomedical Applications
5.2. Non-Biomedical Applications
6. Pre-Treatment, Extraction, Isolation, and Purification
6.1. Pre-Treatment
6.2. Extraction Methods
6.2.1. Conventional Methods
Salt Solubilization
Chemical Hydrolysis
6.2.2. Novel Methods
Enzymatic Hydrolysis
Ultrasound-Assisted Extraction
Microwave-Assisted Extraction and Other Methods
6.3. Isolation Methods
6.3.1. Chromatography
6.3.2. Centrifugation
6.3.3. Use of Non-Aqueous Solvents for Isolation and Purification
6.4. Assaying of Isolated Collagen
6.4.1. Western Blotting
6.4.2. SDS-PAGE
6.4.3. Spectrophotometric Analysis
6.5. Characterization of Isolated Collagen
6.5.1. Differential Scanning Calorimetry
6.5.2. Tyrosine Measurement
6.5.3. Hydroxyproline Determination
7. Functional Properties of Collagen
7.1. Gelling and Hydrophilic Properties
7.2. Emulsifying Properties
7.3. Film Forming Properties
8. Challenges and Future Perspectives of Sea Cucumber Collagen
Author Contributions
Funding
Conflicts of Interest
References
- Shahidi, F. Nutraceuticals and functional foods: Whole versus processed foods. Trends Food Sci. Technol. 2009, 20, 376–387. [Google Scholar] [CrossRef]
- Bordbar, S.; Anwar, F.; Saari, N. High-value components and bioactives from sea cucumbers for functional foods—A review. Mar. Drugs 2011, 9, 1761–1805. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Novel functional food ingredients from marine sources. Curr. Opin. Food Sci. 2015, 2, 123–129. [Google Scholar] [CrossRef]
- Rodríguez, F.; Morán, L.; González, G.; Troncoso, E.; Zúñiga, R.N. Collagen extraction from mussel byssus: A new marine collagen source with physicochemical properties of industrial interest. J. Food Sci. Technol. 2017, 54, 1228–1238. [Google Scholar] [CrossRef] [PubMed]
- García, J.; Méndez, D.; Álvarez, M.; Sanmartin, B.; Vazquez Sobrado, R.; Regueiro, L.; Atanassova, M. Design of novel functional food products enriched with bioactive extracts from holothurians for meeting the nutritional needs of the elderly. LWT 2019, 109, 55–62. [Google Scholar] [CrossRef]
- Hamed, I.; Özogul, F.; Özogul, Y.; Regenstein, J.M. Marine bioactive compounds and their health benefits: A review. Compr. Rev. Food Sci. Food Saf. 2015, 14, 446–465. [Google Scholar] [CrossRef]
- Sun, J.; Hamel, J.; Gianasi, B.; Graham, M.; Mercier, A. Growth, health and biochemical composition of the sea cucumber Cucumaria frondosa after multi-year holding in effluent waters of land-based salmon culture. Aquac. Environ. Interact. 2020, 12, 139–151. [Google Scholar] [CrossRef]
- Karim, A.A.; Bhat, R. Fish gelatin: Properties, challenges, and prospects as an alternative to mammalian gelatins. Food Hydrocoll. 2009, 23, 563–576. [Google Scholar] [CrossRef]
- Gomez-Guillen, M.C.; Gimenez, B.; Lopez-Caballero, M.E.; Montero, M.P. Functional and bioactive properties of collagen and gelatin from alternative sources: A review. Food Hydrocoll. 2011, 25, 1813–1827. [Google Scholar] [CrossRef]
- Pal, G.K.; Suresh, P.V. Sustainable valorisation of seafood by-products: Recovery of collagen and development of collagen-based novel functional food ingredients. Innov. Food Sci. Emerg. Technol. 2016, 37, 201–215. [Google Scholar] [CrossRef]
- Janakiram, N.B.; Mohammed, A.; Rao, C.V. Sea cucumbers metabolites as potent anti-cancer agents. Mar. Drugs 2015, 13, 2909–2923. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Feng, W.; Hu, S.; Liang, S.; An, N.; Mao, Y. Bioactive compounds of sea cucumbers and their therapeutic effects. Chin. J. Oceanol. Limnol. 2016, 34, 549–558. [Google Scholar] [CrossRef]
- Silchenko, A.S.; Kalinovsky, A.I.; Avilov, S.A.; Kalinin, V.I.; Andrijaschenko, P.V.; Dmitrenok, P.S.; Chingizova, E.A.; Ermakova, S.P.; Malyarenko, O.S.; Dautova, T.N. Nine new triterpene glycosides, magnumosides A1–A4, B1, B2, C1, C2 and C4, from the vietnamese sea cucumber neothyonidium (=massinium) magnum: Structures and activities against tumor cells independently and in synergy with radioactive irradiation. Mar. Drugs 2017, 15, 256. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Kunisaki, N.; Urano, N.; Kimura, S. Collagen as the major edible component of sea cucumber (Stichopus japonicus). J. Food Sci. 2002, 67, 1319–1322. [Google Scholar] [CrossRef]
- Pangestuti, R.; Arifin, Z. Medicinal and health benefit effects of functional sea cucumbers. J. Tradit. Complement. Med. 2018, 8, 341–351. [Google Scholar] [CrossRef]
- Cui, F.-x.; Xue, C.-h.; Li, Z.-j.; Zhang, Y.-q.; Dong, P.; Fu, X.-y.; Gao, X. Characterization and subunit composition of collagen from the body wall of sea cucumber Stichopus japonicus. Food Chem. 2007, 100, 1120–1125. [Google Scholar] [CrossRef]
- Subhan, F.; Ikram, M.; Shehzad, A.; Ghafoor, A. Marine Collagen: An emerging player in biomedical applications. J. Food Sci. Technol. 2015, 52, 4703–4707. [Google Scholar] [CrossRef]
- Hossain, A.; Dave, D.; Shahidi, F. Northern sea cucumber (cucumaria frondosa): A potential candidate for functional food, nutraceutical, and pharmaceutical sector. Mar. Drugs 2020, 18, 274. [Google Scholar] [CrossRef]
- Ferrario, C.; Rusconi, F.; Pulaj, A.; Macchi, R.; Landini, P.; Paroni, M.; Colombo, G.; Martinello, T.; Melotti, L.; Gomiero, C.; et al. From food waste to innovative biomaterial: Sea urchin-derived collagen for applications in skin regenerative medicine. Mar. Drugs 2020, 18, 414. [Google Scholar] [CrossRef]
- Silvipriya, K.S.; Krishna Kumar, K.; Bhat, A.R.; Dinesh Kumar, B.; John, A.; Lakshmanan, P. Collagen: Animal sources and biomedical application. J. Appl. Pharm. Sci. 2015, 5, 123–127. [Google Scholar] [CrossRef]
- Wang, L.; Liang, Q.; Chen, T.; Wang, Z.; Xu, J.; Ma, H. Characterization of collagen from the skin of amur sturgeon (acipenser schrenckii). Food Hydrocoll. 2014, 38, 104–109. [Google Scholar] [CrossRef]
- Wang, L.; Liang, Q.; Wang, Z.; Xu, J.; Liu, Y.; Ma, H. Preparation and characterisation of type i and v collagens from the skin of amur sturgeon (acipenser schrenckii). Food Chem. 2014, 148, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, M.H.; Suo, Z.; Avci, R.; Asara, J.M.; Allen, M.A.; Arce, F.T.; Horner, J.R. Analyses of soft tissue from tyrannosaurus rex suggest the presence of protein. Science 2007, 316, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, G.N.; Kartha, G. Structure of collagen. Nature 1954, 174, 269–270. [Google Scholar] [CrossRef] [PubMed]
- Cowan, P.M.; North, A.C.T.; Randall, J.T. X-ray diffraction studies of collagen fibres. Symp. Soc. Exp. Biol. 1955, 9, 115–126. [Google Scholar] [CrossRef]
- Rich, A.; Crick, F. The structure of collagen. Nature 1955, 176, 915–916. [Google Scholar] [CrossRef] [PubMed]
- Bhagwat, P.K.; Dandge, P.B. Collagen and collagenolytic proteases: A review. Biocatal. Agric. Biotechnol. 2018, 15, 43–55. [Google Scholar] [CrossRef]
- Sorushanova, A.; Delgado, L.M.; Wu, Z.; Shologu, N.; Kshirsagar, A.; Raghunath, R.; Mullen, A.M.; Bayon, Y.; Pandit, A.; Raghunath, M.; et al. The Collagen suprafamily: From biosynthesis to advanced biomaterial development. Adv. Mater. 2019, 31, 1–39. [Google Scholar] [CrossRef]
- Sherman, V.R.; Yang, W.; Meyers, M.A. The materials science of collagen. J. Mech. Behav. Biomed. Mater. 2015, 52, 22–50. [Google Scholar] [CrossRef]
- Feyzi, S.; Varidi, M.; Zare, F.; Varidi, M.J. Fenugreek (trigonella foenum graecum) seed protein isolate: Extraction optimization, amino acid composition, thermo and functional properties. J. Sci. Food Agric. 2015, 95, 3165–3176. [Google Scholar] [CrossRef]
- Persikov, A.V.; Ramshaw, J.A.M.; Brodsky, B. Prediction of collagen stability from amino acid sequence. J. Biol. Chem. 2005, 280, 19343–19349. [Google Scholar] [CrossRef] [PubMed]
- Ricard-Blum, S. The collagen family. Cold Spring Harb. Perspect. Biol. 2011, 3, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Terzi, A.; Gallo, N.; Bettini, S.; Sibillano, T.; Altamura, D.; Campa, L.; Natali, M.L.; Salvatore, L.; Madaghiele, M.; De Caro, L.; et al. Investigations of processing–induced structural changes in horse type-i collagen at sub and supramolecular levels. Front. Bioeng. Biotechnol. 2019, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kadler, K.E.; Baldock, C.; Bella, J.; Boot-Handford, R.P. Collagens at a glance. J. Cell Sci. 2007, 120, 1955–1958. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.J.; Woods, P.E.; Eyre, D.R. Identification of cross-linking sites in bovine cartilage type IX collagen reveals an antiparallel type II-type IX molecular relationship and type IX to type IX bonding. J. Biol. Chem. 1992, 267, 23007–23014. [Google Scholar]
- Mizuno, K.; Boudko, S.P.; Engel, J.; Peter Bächinger, H. Kinetic hysteresis in collagen folding. Biophys. J. 2010, 98, 3004–3014. [Google Scholar] [CrossRef]
- Ertem, F.; Watson, A.R.; Rivers, C.R.; Babichenko, D.; Tang, G.; Schwartz, M.; Proksell, S.; Johnston, E.; Hashash, J.G.; Barrie, A.; et al. Mo1823 – endoscopic patterns and location of post-operative recurrence in crohn’s disease patients with side to side anastomosis following ileocecal resection. Gastroenterology 2019, 156, S-850–S-851. [Google Scholar] [CrossRef]
- Franzke, C.W.; Bruckner, P.; Bruckner-Tuderman, L. Collagenous transmembrane proteins: Recent insights into biology and pathology. J. Biol. Chem. 2005, 280, 4005–4008. [Google Scholar] [CrossRef]
- Van Der Rest, M.; Garrone, R. Collagen family of proteins. FASEB J. 1991, 5, 2814–2823. [Google Scholar] [CrossRef]
- Ramshaw, J.A.M.; Werkmeister, J.A.; Glattauer, V. Collagen-based biomaterials. Biotechnol. Genet. Eng. Rev. 1996, 13, 335–382. [Google Scholar] [CrossRef]
- Friess, W. Collagen—Biomaterial for drug delivery. Eur. J. Pharm. Biopharm. 1998, 45, 113–136. [Google Scholar] [CrossRef]
- Kittiphattanabawon, P.; Benjakul, S.; Visessanguan, W.; Shahidi, F. Isolation and characterization of collagen from the cartilages of brownbanded bamboo shark (chiloscyllium punctatum) and blacktip shark (carcharhinus limbatus). LWT Food Sci. Technol. 2010, 43, 792–800. [Google Scholar] [CrossRef]
- Liu, D.C.; Lin, Y.K.; Chen, M.T. Optimum condition of extracting collagen from chicken feet and its characteristics. Asian Anim. Sci. 2001, 14, 1638–1644. [Google Scholar] [CrossRef]
- Conway, J.R.W.; Vennin, C.; Cazet, A.S.; Herrmann, D.; Murphy, K.J.; Warren, S.C.; Wullkopf, L.; Boulghourjian, A.; Zaratzian, A.; Da Silva, A.M.; et al. Three-dimensional organotypic matrices from alternative collagen sources as pre-clinical models for cell biology. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef]
- Lausch, A.J.; Chong, L.C.; Uludag, H.; Sone, E.D. Multiphasic collagen scaffolds for engineered tissue interfaces. Adv. Funct. Mater. 2018, 28, 1–9. [Google Scholar] [CrossRef]
- Huda, N.; Seow, E.K.; Normawati, M.N.; Nik Aisyah, N.M. Preliminary study on physicochemical properties of duck feet collagen. Int. J. Poult. Sci. 2013, 12, 615–621. [Google Scholar] [CrossRef]
- Wood, A.; Ogawa, M.; Portier, R.J.; Schexnayder, M.; Shirley, M.; Losso, J.N. Biochemical properties of alligator (alligator mississippiensis) bone collagen. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 151, 246–249. [Google Scholar] [CrossRef]
- Lin, Y.K.; Liu, D.C. Comparison of physical-chemical properties of type I collagen from different species. Food Chem. 2006, 99, 244–251. [Google Scholar] [CrossRef]
- Cheng, F.Y.; Hsu, F.W.; Chang, H.S.; Lin, L.C.; Sakata, R. Effect of different acids on the extraction of pepsin-solubilised collagen containing melanin from silky fowl feet. Food Chem. 2009, 113, 563–567. [Google Scholar] [CrossRef]
- Zhou, C.; Li, Y.; Yu, X.; Yang, H.; Ma, H.; Yagoub, A.E.G.A.; Cheng, Y.; Hu, J.; Otu, P.N.Y. Extraction and characterization of chicken feet soluble collagen. LWT-Food Sci. Technol. 2016, 74, 145–153. [Google Scholar] [CrossRef]
- Busra, F.M.; Chowdhury, S.R.; Saim, A.B.; Idrus, R.B. Genotoxicity and cytotoxicity of ovine collagen on human dermal fibroblasts. Saudi Med. J. 2011, 32, 1311–1312. [Google Scholar] [PubMed]
- Amri, M.; Firdaus, M.; Fauzi, M.; Chowdhury, S.; Fadilah, N.; Hamirul, W.W.; Reusmaazran, M.; Aminuddin, B.; Ruszymah, B. Cytotoxic evaluation of biomechanically improved crosslinked ovine collagen on human dermal fibroblasts. Bio-Med. Mater. Eng. 2014, 24, 1715–1724. [Google Scholar] [CrossRef]
- Fauzi, M.B.; Lokanathan, Y.; Aminuddin, B.S.; Ruszymah, B.H.I.; Chowdhury, S.R. Ovine tendon collagen: Extraction, characterisation and fabrication of thin films for tissue engineering applications. Mater. Sci. Eng. C 2016, 68, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Fauzi, M.B.; Lokanathan, Y.; Nadzir, M.M.; Aminuddin, S.; Ruszymah, B.H.I.; Chowdhury, S.R. Attachment, proliferation, and morphological properties of human dermal fibroblasts on ovine tendon collagen scaffolds: A comparative study. Malays. J. Med. Sci. 2017, 24, 33–43. [Google Scholar] [CrossRef]
- Cersoy, S.; Zazzo, A.; Lebon, M.; Rofes, J.; Zirah, S. Collagen extraction and stable isotope analysis of small vertebrate bones: A comparative approach. Radiocarbon 2017, 59, 679–694. [Google Scholar] [CrossRef]
- Zhang, J.; Duan, R. Characterisation of acid-soluble and pepsin-solubilised collagen from frog (rana nigromaculata) skin. Int. J. Biol. Macromol. 2017, 101, 638–642. [Google Scholar] [CrossRef]
- de Almeida, P.F.; de Araújo, M.G.O.; Santana, J.C.C. Obtenção de colágeno a partir de tarsos de frangos para produção de geleias. Acta Sci. Technol. 2012, 34, 345–351. [Google Scholar] [CrossRef]
- Hashim, P.; Ridzwan, M.S.M.; Bakar, J. Isolation and characterization of collagen from chicken Feet. Int. J. Biol. Biomol. Agric. Food Biotechnol. Eng. 2014, 8, 147–151. [Google Scholar]
- Lee, J.H.; Lee, J.; Song, K. Bin Development of a chicken feet protein film containing essential oils. Food Hydrocoll. 2015, 46, 208–215. [Google Scholar] [CrossRef]
- Dhakal, D.; Koomsap, P.; Lamichhane, A.; Sadiq, M.B.; Anal, A.K. Optimization of collagen extraction from chicken feet by papain hydrolysis and synthesis of chicken feet collagen based biopolymeric fibres. Food Biosci. 2018, 23, 23–30. [Google Scholar] [CrossRef]
- Huda, N.; Seow, E.K.; Normawati, M.N.; Nik Aisyah, N.M.; Fazilah, A.; Easa, A.M. Effect of duck feet collagen addition on physicochemical properties of surimi. Int. Food Res. J. 2013, 20, 537–544. [Google Scholar]
- Nogueira, F.G.E.; do Prado, N.T.; Oliveira, L.C.A.; Bastos, A.R.R.; Lopes, J.H.; de Carvalho, J.G. Incorporation of mineral phosphorus and potassium on leather waste (collagen): A new NcollagenPK-fertilizer with slow liberation. J. Hazard. Mater. 2010, 176, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Rizk, M.A.; Mostafa, N.Y. Extraction and characterization of collagen from buffalo skin for biomedical applications. Orient. J. Chem. 2016, 32, 1601–1609. [Google Scholar] [CrossRef]
- Zahrani, A.R. Extraction and Isolation of Collagen Type I from Fish Skin. Master’s Thesis, The University of Otago, Dunedin, New Zealand, November 2011. [Google Scholar]
- Abraham, G.A.; Murray, J.; Billiar, K.; Sullivan, S.J. Evaluation of the porcine intestinal collagen layer as a biomaterial. J. Biomed. Mater. Res. 2000, 51, 442–452. [Google Scholar] [CrossRef]
- Chen, X.; Wu, J.-H.; Li, L.; Wang, S.-Y. The cryoprotective effects of antifreeze peptides from pigskin collagen on texture properties and water mobility of frozen dough subjected to freeze–thaw cycles. Eur. Food Res. Technol. 2017, 243, 1149–1156. [Google Scholar] [CrossRef]
- Maione-Silva, L.; de Castro, E.G.; Nascimento, T.L.; Cintra, E.R.; Moreira, L.C.; Cintra, B.A.S.; Valadares, M.C.; Lima, E.M. Ascorbic acid encapsulated into negatively charged liposomes exhibits increased skin permeation, retention and enhances collagen synthesis by fibroblasts. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Munasinghe, K.A.; Schwarz, J.G.; Whittiker, M. Utilization of chicken by-products to form collagen films. J. Food Process. 2015, 2015, 1–6. [Google Scholar] [CrossRef]
- Shahidi, F.; Varatharajan, V.; Peng, H.; Senadheera, R. Utilization of marine by-products for the recovery of value-added products. J. Food Bioact. 2019, 6, 10–61. [Google Scholar] [CrossRef]
- Dave, D.; Liu, Y.; Clark, L.; Dave, N.; Trenholm, S.; Westcott, J. Availability of marine collagen from Newfoundland fisheries and aquaculture waste resources. Bioresour. Technol. Reports 2019, 7, 100271. [Google Scholar] [CrossRef]
- Hashim, P.; Mohd Ridzwan, M.S.; Bakar, J.; Mat Hashim, D. Collagen in food and beverage industries. Int. Food Res. J. 2015, 22, 1–8. [Google Scholar]
- Li, B. Beneficial effects of collagen hydrolysate: A review on recent developments. Biomed. J. Sci. Tech. Res. 2017, 1, 2–5. [Google Scholar] [CrossRef]
- Dave, D.; Routray, W. Current scenario of canadian fishery and corresponding underutilized species and fishery byproducts: A potential source of omega-3 fatty acids. J. Clean. Prod. 2018, 180, 617–641. [Google Scholar] [CrossRef]
- Kim, H.K.; Kim, Y.H.; Kim, Y.J.; Park, H.J.; Lee, N.H. Effects of ultrasonic treatment on collagen extraction from skins of the sea bass Lateolabrax japonicus. Fish. Sci. 2012, 78, 485–490. [Google Scholar] [CrossRef]
- Kittiphattanabawon, P.; Benjakul, S.; Sinthusamran, S.; Kishimura, H. Characteristics of collagen from the skin of clown featherback (Chitala ornata). Int. J. Food Sci. Technol. 2015, 50, 1972–1978. [Google Scholar] [CrossRef]
- Kaewdang, O.; Benjakul, S.; Kaewmanee, T.; Kishimura, H. Characteristics of collagens from the swim bladders of yellowfin tuna (Thunnus albacares). Food Chem. 2014, 155, 264–270. [Google Scholar] [CrossRef]
- Liu, D.; Liang, L.; Regenstein, J.M.; Zhou, P. Extraction and characterisation of pepsin-solubilised collagen from fins, scales, skins, bones and swim bladders of bighead carp (hypophthalmichthys nobilis). Food Chem. 2012, 133, 1441–1448. [Google Scholar] [CrossRef]
- Manchinasetty, N.V.L.; Oshima, S.; Kikuchi, M. Preparation of flexible bone tissue scaffold utilizing sea urchin test and collagen. J. Mater. Sci. Mater. Med. 2017, 28, 4–12. [Google Scholar] [CrossRef]
- Nagai, T.; Nagamori, K.; Yamashita, E.; Suzuki, N. Collagen of octopus callistoctopus arakawai arm. Int. J. Food Sci. Technol. 2002, 37, 285–289. [Google Scholar] [CrossRef]
- Kittiphattanabawon, P.; Nalinanon, S.; Benjakul, S.; Kishimura, H. Characteristics of pepsin-solubilised collagen from the skin of splendid squid (loligo formosana). J. Chem. 2015, 2015. [Google Scholar] [CrossRef]
- Jridi, M.; Bardaa, S.; Moalla, D.; Rebaii, T.; Souissi, N.; Sahnoun, Z.; Nasri, M. Microstructure, rheological and wound healing properties of collagen-based gel from cuttlefish skin. Int. J. Biol. Macromol. 2015, 77, 369–374. [Google Scholar] [CrossRef]
- Exposito, J.Y.; Larroux, C.; Cluzel, C.; Valcourt, U.; Lethias, C.; Degnan, B.M. Demosponge and sea anemone fibrillar collagen diversity reveals the early emergence of A/C clades and the maintenance of the modular structure of type V/XI collagens from sponge to human. J. Biol. Chem. 2008, 283, 28226–28235. [Google Scholar] [CrossRef] [PubMed]
- Li, P.H.; Lu, W.C.; Chan, Y.J.; Ko, W.C.; Jung, C.C.; Le Huynh, D.T.; Ji, Y.X. Extraction and characterization of collagen from sea cucumber (holothuria cinerascens) and its potential application in moisturizing cosmetics. Aquaculture 2020, 515, 734590. [Google Scholar] [CrossRef]
- Abdollahi, M.; Rezaei, M.; Jafarpour, A.; Undeland, I. Sequential extraction of gel-forming proteins, collagen and collagen hydrolysate from gutted silver carp (hypophthalmichthys molitrix), a biorefinery approach. Food Chem. 2018, 242, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Kobayashi, S.; Yamashita, M. Comparison of connective tissue structure and muscle toughness of spotted mackerel Scomber australasicus and Pacific mackerel S. japonicus during chilled and frozen storage. Fish. Sci. 2017, 83, 133–139. [Google Scholar] [CrossRef]
- Berillis, P.; Environment, A.; Envi-, A.; Ionia, N. Marine Collagen: Extraction and applications. SM Gr. Open Access eBooks 2015, 1–13. [Google Scholar]
- Arumugam, G.K.S.; Sharma, D.; Balakrishnan, R.M.; Ettiyappan, J.B.P. Extraction, optimization and characterization of collagen from sole fish skin. Sustain. Chem. Pharm. 2018, 9, 19–26. [Google Scholar] [CrossRef]
- Yu, D.; Chi, C.F.; Wang, B.; Ding, G.F.; Li, Z.R. Characterization of acid-and pepsin-soluble collagens from spines and skulls of skipjack tuna (Katsuwonus pelamis). Chin. J. Nat. Med. 2014, 12, 712–720. [Google Scholar] [CrossRef]
- Kittiphattanabawon, P.; Sriket, C.; Kishimura, H.; Benjakul, S. Characteristics of acid and pepsin solubilized collagens from nile tilapia (oreochromis niloticus) scale. Emirates J. Food Agric. 2019, 31, 95–101. [Google Scholar] [CrossRef]
- Hickman, D.; Sims, T.J.; Miles, C.A.; Bailey, A.J.; De Mari, M.; Koopmans, M. Isinglass/collagen: Denaturation and functionality. J. Biotechnol. 2000, 79, 245–257. [Google Scholar] [CrossRef]
- Wasswa, J.; Tang, J.; Gu, X.-H.; Yuan, X.-Q. Influence of the extent of enzymatic hydrolysis on the functional properties of protein hydrolysate from grass carp (ctenopharyngodon idella) skin. Food Chem. 2007, 104, 1698–1704. [Google Scholar] [CrossRef]
- Hoyer, B.; Bernhardt, A.; Lode, A.; Heinemann, S.; Sewing, J.; Klinger, M.; Notbohm, H.; Gelinsky, M. Jellyfish collagen scaffolds for cartilage tissue engineering. Acta Biomater. 2014, 10, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Swatschek, D.; Schatton, W.; Kellermann, J.; Müller, W.E.G.; Kreuter, J. Marine sponge collagen: Isolation, characterization and effects on the skin parameters surface-pH, moisture and sebum. Eur. J. Pharm. Biopharm. 2002, 53, 107–113. [Google Scholar] [CrossRef]
- Suhre, M.H.; Gertz, M.; Steegborn, C.; Scheibel, T. Structural and functional features of a collagen-binding matrix protein from the mussel byssus. Nat. Commun. 2014, 5, 3392. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Dong, X.; Zhou, D.; Gao, Y.; Yang, J.; Li, D.; Zhao, X.; Ren, T.; Ye, W.; Tan, H.; et al. Physicochemical properties and radical scavenging capacities of pepsin-solubilized collagen from sea cucumber Stichopus japonicus. Food Hydrocoll. 2012, 28, 182–188. [Google Scholar] [CrossRef]
- Park, S.Y.; Lim, H.K.; Lee, S.; Hwang, H.C.; Cho, S.K.; Cho, M. Pepsin-solubilised collagen (PSC) from Red Sea cucumber (Stichopus japonicus) regulates cell cycle and the fibronectin synthesis in HaCaT cell migration. Food Chem. 2012, 132, 487–492. [Google Scholar] [CrossRef]
- Wu, H.T.; Li, D.M.; Zhu, B.W.; Sun, J.J.; Zheng, J.; Wang, F.L.; Konno, K.; Jiang, X. Proteolysis of noncollagenous proteins in sea cucumber, stichopus japonicus, body wall: Characterisation and the effects of cysteine protease inhibitors. Food Chem. 2013, 141, 1287–1294. [Google Scholar] [CrossRef]
- Adibzadeh, N.; Aminzadeh, S.; Jamili, S.; Karkhane, A.A.; Farrokhi, N. Purification and characterization of pepsin-solubilized collagen from skin of sea cucumber holothuria parva. Appl. Biochem. Biotechnol. 2014, 173, 143–154. [Google Scholar] [CrossRef]
- Abedin, M.Z.; Karim, A.A.; Ahmed, F.; Latiff, A.A.; Gan, C.Y.; Che Ghazali, F.; Islam Sarker, M.Z. Isolation and characterization of pepsin-solubilized collagen from the integument of sea cucumber (stichopus vastus). J. Sci. Food Agric. 2013, 93, 1083–1088. [Google Scholar] [CrossRef]
- Abedin, M.Z.; Karim, A.A.; Gan, C.Y.; Ghazali, F.C.; Barzideh, Z.; Zzaman, W.; Zaidul, I.S.M. Identification of angiotensin I converting enzyme inhibitory and radical scavenging bioactive peptides from sea cucumber (stichopus vastus) collagen hydrolysates through optimization. Int. Food Res. J. 2015, 22, 1074–1082. [Google Scholar]
- Zhong, M.; Chen, T.; Hu, C.; Ren, C. Isolation and characterization of collagen from the body wall of sea cucumber stichopus monotuberculatus. J. Food Sci. 2015, 80, C671–C679. [Google Scholar] [CrossRef]
- Liu, Y.X.; Zhou, D.Y.; Ma, D.D.; Liu, Z.Q.; Liu, Y.F.; Song, L.; Dong, X.P.; Li, D.M.; Zhu, B.W.; Konno, K.; et al. Effects of endogenous cysteine proteinases on structures of collagen fibres from dermis of sea cucumber (Stichopus japonicus). Food Chem. 2017, 232, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.X.; Zhou, D.Y.; Liu, Z.Q.; Lu, T.; Song, L.; Li, D.M.; Dong, X.P.; Qi, H.; Zhu, B.W.; Shahidi, F. Structural and biochemical changes in dermis of sea cucumber (stichopus japonicus) during autolysis in response to cutting the body wall. Food Chem. 2018, 240, 1254–1261. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Xue, C.; Chang, Y.; Shen, J.; Zhang, Y.; Li, Z.; Wang, Y. Collagen fibrils of sea cucumber (apostichopus japonicus) are heterotypic. Food Chem. 2020, 316, 126272. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Shen, P.; Yu, M.; Yu, C.; Zhu, B.; Qi, H. (−)-Epigallocatechin gallate protected molecular structure of collagen fibers in sea cucumber apostichopus japonicus body wall during thermal treatment. LWT 2020, 123, 1–7. [Google Scholar] [CrossRef]
- Eyre, D.; Glimcher, M. Comparative biochemistry of collagen crosslinks: Reducible bonds in invertebrate collagens. Biochim. Biophys. Acta, Protein Struct. 1971, 243, 525–529. [Google Scholar] [CrossRef]
- Matsumura, T.; Shinmei, M.; Nagai, Y. Disaggregation of connective tissue: Preparation of fibrous components from sea cucumber body wall and calf skin. J. Biochem. 1973, 73, 155–162. [Google Scholar] [CrossRef]
- Eyre, D.R.; Glimcher, M.J. Evidence for glycosylated crosslinks in body-wall collagen of the sea cucumber, thyone briareus. Proc. Soc. Exp. Biol. Med. 1973, 144, 400–403. [Google Scholar] [CrossRef]
- Trotter, J.A.; Lyons-Levy, G.; Thurmond, F.A.; Koob, T.J. Covalent composition of collagen fibrils from the dermis of the sea cucumber, cucumaria frondosa, a tissue with mutable mechanical properties. Comp. Biochem. Physiol. Part A Physiol. 1995, 112, 463–478. [Google Scholar] [CrossRef]
- Trotter, J.A.; Chapman, J.A.; Kadler, K.E.; Holmes, D.F. Growth of sea cucumber collagen fibrils occurs at the tips and centers in a coordinated manner. J. Mol. Biol. 1998, 284, 1417–1424. [Google Scholar] [CrossRef]
- Trotter, J.A.; Kadler, K.E.; Holmes, D.F. Echinoderm collagen fibrils grow by surface-nucleation-and-propagation from both centers and ends. J. Mol. Biol. 2000, 300, 531–540. [Google Scholar] [CrossRef]
- Trotter, J.A.; Lyons-Levy, G.; Luna, D.; Koob, T.J.; Keene, D.R.; Atkinson, M.A.L. Stiparin: A glycoprotein from sea cucumber dermis that aggregates collagen fibrils. Matrix Biol. 1996, 15, 99–110. [Google Scholar] [CrossRef]
- Trotter, J.A.; Lyons-Levy, G.; Chino, K.; Koob, T.J.; Keene, D.R.; Atkinson, M.A.L. Collagen fibril aggregation-inhibitor from sea cucumber dermis. Matrix Biol. 1999, 18, 569–578. [Google Scholar] [CrossRef]
- Thurmond, F.A.; Trotter, J.A. Morphology and biomechanics of the microfibrillar network of sea cucumber dermis. J. Exp. Biol. 1996, 199, 1817–1828. [Google Scholar] [PubMed]
- Dong, X.; Zhu, B.; Sun, L.; Zheng, J.; Jiang, D.; Zhou, D.; Wu, H.; Murata, Y. Changes of collagen in sea cucumber (stichopus japonicas) during cooking. Food Sci. Biotechnol. 2011, 20, 1137–1141. [Google Scholar] [CrossRef]
- Liu, Y.X.; Zhou, D.Y.; Ma, D.D.; Liu, Y.F.; Li, D.M.; Dong, X.P.; Tan, M.Q.; Du, M.; Zhu, B.W. Changes in collagenous tissue microstructures and distributions of cathepsin L in body wall of autolytic sea cucumber (stichopus japonicus). Food Chem. 2016, 212, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Si, L.; Fan, Y.; Wang, Y.; Sun, L.; Li, B.; Xue, C.; Hou, H. Thermal degradation behavior of collagen from sea cucumber (stichopus japonicus) using TG-FTIR analysis. Thermochim. Acta 2018, 659, 166–171. [Google Scholar] [CrossRef]
- Liu, Z.; Tuo, F.; Song, L.; Liu, Y.; Dong, X.; Li, D.; Zhou, D.; Shahidi, F. Action of trypsin on structural changes of collagen fibres from sea cucumber (Stichopus japonicus). Food Chem. 2018, 256, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-; Liu, Z.; Song, L.; Ma, Q.; Zhou, D.; Zhu, B.; Shahidi, F. Effects of collagenase type I on the structural features of collagen fibres from sea cucumber (stichopus japonicus) body wall. Food Chem. 2019, 301, 125302. [Google Scholar] [CrossRef]
- Liu, Z.; Oliveira, A.C.M.; Su, Y.C. Purification and characterization of pepsin-solubilized collagen from skin and connective tissue of giant red sea cucumber (parastichopus californicus). J. Agric. Food Chem. 2010, 58, 1270–1274. [Google Scholar] [CrossRef]
- Siddiqui, Y.D.; Arief, E.M.; Yusoff, A.; Suzina, A.H.; Abdullah, S.Y. Isolation of pepsin solubilized collagen (PSC) from crude collagen extracted from body wall of sea cucumber (bohadschia spp.). Int. J. Pharm. Pharm. Sci. 2013, 5, 555–559. [Google Scholar]
- Abedin, M.Z.; Karim, A.A.; Latiff, A.A.; Gan, C.-Y.; Ghazali, F.C.; Barzideh, Z.; Ferdosh, S.; Akanda, M.J.H.; Zzaman, W.; Karim, M.R.; et al. Biochemical and radical-scavenging properties of sea cucumber (stichopus vastus) collagen hydrolysates. Nat. Prod. Res. 2014, 28, 1302–1305. [Google Scholar] [CrossRef] [PubMed]
- Sari, P.A. Determination of nano-collagen quality from sea cucumber holothuria scabra. IOP Conf. Ser. Earth Environ. Sci. 2020, 430, 012005. [Google Scholar] [CrossRef]
- Liu, F.; Zamora, L.; Jeffs, A.; Quek, S.Y. Biochemical composition of the Australasian sea cucumber, australostichopus mollis, from a nutritional point of view. Nutrire 2017, 42, 1–11. [Google Scholar] [CrossRef]
- Abdillah, S.; Wijiyanti, G.; Setiawan, M.; Noor, S.U.; Nurilmala, M. In vitro anti-tyrosinase and anti-elastase activity of collagen from sea cucumber (holothuria leucospilota). Afr. J. Biotech 2017, 16, 771–776. [Google Scholar] [CrossRef][Green Version]
- Lin, S.; Xue, Y.-P.; San, E.; Keong, T.C.; Chen, L.; Zheng, Y.-G. Extraction and Characterization of pepsin soluble collagen from the body wall of sea cucumber acaudina leucoprocta. J. Aquat. Food Prod. Technol. 2017, 26, 502–515. [Google Scholar] [CrossRef]
- Jin, H.X.; Xu, H.P.; Li, Y.; Zhang, Q.W.; Xie, H. Preparation and evaluation of peptides with potential antioxidant activity by microwave assisted enzymatic hydrolysis of collagen from sea cucumber acaudina molpadioides obtained from Zhejiang province in China. Mar. Drugs 2019, 17, 169. [Google Scholar] [CrossRef] [PubMed]
- Yuniati, R.; Sulardiono, B. Exploration of the collagen of non commercial sea cucumber Holothuria atra and commercial sea cucumber stichopus vastus in the Karimunjawa Islands, Indonesia. Ocean Life 2019, 3, 18–23. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, W.; Li, G.; Shi, B.; Miao, Y.; Wu, X. Isolation and partial characterization of pepsin-soluble collagen from the skin of grass carp (ctenopharyngodon idella). Food Chem. 2007, 103, 906–912. [Google Scholar] [CrossRef]
- Ottani, V.; Martini, D.; Franchi, M.; Ruggeri, A.; Raspanti, M. Hierarchical structures in fibrillar collagens. Micron 2002, 33, 587–596. [Google Scholar] [CrossRef]
- Eyre, D.R.; Weis, M.A.; Rai, J. Analyses of lysine aldehyde cross-linking in collagen reveal that the mature cross-link histidinohydroxylysinonorleucine is an artifact. J. Biol. Chem. 2019, 294, 6578–6590. [Google Scholar] [CrossRef]
- Toroian, D.; Joo, E.L.; Price, P.A. The size exclusion characteristics of type I collagen: Implications for the role of noncollagenous bone constituents in mineralization. J. Biol. Chem. 2007, 282, 22437–22447. [Google Scholar] [CrossRef] [PubMed]
- Pucci, F.; Rooman, M. Physical and molecular bases of protein thermal stability and cold adaptation. Curr. Opin. Struct. Biol. 2017, 42, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Leikina, E.; Mertts, M.V.; Kuznetsova, N.; Leikin, S. Type I collagen is thermally unstable at body temperature. Proc. Natl. Acad. Sci. USA 2002, 99, 1314–1318. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Liu, Y.; Liu, A.; Wang, W. Improved thermal-stability and mechanical properties of type I collagen by crosslinking with casein, keratin and soy protein isolate using transglutaminase. Int. J. Biol. Macromol. 2017, 98, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, R.; Haq, M.; Chun, B.S. Characterization of marine derived collagen extracted from the by-products of bigeye tuna (thunnus obesus). Int. J. Biol. Macromol. 2019, 135, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Li, B.; Zhao, X. Isolation and characterization of collagen from squid (ommastrephes bartrami) skin. J. Ocean Univ. China 2009, 8, 191–196. [Google Scholar] [CrossRef]
- Qin, L.; Bi, J.R.; Li, D.M.; Dong, M.; Zhao, Z.Y.; Dong, X.P.; Zhou, D.Y.; Zhu, B.W. Unfolding/refolding study on collagen from sea cucumber based on 2D fourier transform infrared spectroscopy. Molecules 2016, 21, 1546. [Google Scholar] [CrossRef]
- Dong, X.; Qi, H.; Feng, D.; He, B.; Nakamura, Y.; Yu, C.; Zhu, B. Oxidative stress involved in textural changes of sea cucumber stichopus japonicus body wall during low-temperature treatment. Int. J. Food Prop. 2018, 21, 2646–2659. [Google Scholar] [CrossRef]
- Daboor, S.M.; Budge, S.M.; Ghaly, A.E.; Brooks, M.S.; Dave, D. Isolation and activation of collagenase from fish processing waste. Adv. Biosci. Biotechnol. 2012, 03, 191–203. [Google Scholar] [CrossRef]
- Zhou, D.Y.; Chang, X.N.; Bao, S.S.; Song, L.; Zhu, B.W.; Dong, X.P.; Zong, Y.; Li, D.M.; Zhang, M.M.; Liu, Y.X.; et al. Purification and partial characterisation of a cathepsin L-like proteinase from sea cucumber (stichopus japonicus) and its tissue distribution in body wall. Food Chem. 2014, 158, 192–199. [Google Scholar] [CrossRef]
- Zhong, M.; Hu, C.; Ren, C.; Luo, X.; Cai, Y. Characterization of a main extracellular matrix autoenzyme from the dermis of sea cucumber stichopus monotuberculatus: Collagenase. Int. J. Food Prop. 2016, 19, 2495–2509. [Google Scholar] [CrossRef]
- Angele, P.; Abke, J.; Kujat, R.; Faltermeier, H.; Schumann, D.; Nerlich, M.; Kinner, B.; Englert, C.; Ruszczak, Z.; Mehrl, R.; et al. Influence of different collagen species on physico-chemical properties of crosslinked collagen matrices. Biomaterials 2004, 25, 2831–2841. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, G.; Shi, B. Physicochemical properties of collagen, gelatin and collagen hydrolysate derived from bovine limed split wastes. J. Soc. Leather Technol. Chem. 2006, 90, 23–28. [Google Scholar]
- Muthukumar, T.; Sreekumar, G.; Sastry, T.P.; Chamundeeswari, M. Collagen as a potential biomaterial in biomedical applications. Rev. Adv. Mater. Sci. 2018, 53, 29–39. [Google Scholar] [CrossRef]
- Ferrario, C.; Leggio, L.; Leone, R.; Di Benedetto, C.; Guidetti, L.; Coccè, V.; Ascagni, M.; Bonasoro, F.; La Porta, C.A.M.; Candia Carnevali, M.D.; et al. Marine-derived collagen biomaterials from echinoderm connective tissues. Mar. Environ. Res. 2017, 128, 46–57. [Google Scholar] [CrossRef]
- Carvalho, A.M.; Marques, A.P.; Silva, T.H.; Reis, R.L. Evaluation of the potential of collagen from codfish skin as a biomaterial for biomedical applications. Mar. Drugs 2018, 16, 495. [Google Scholar] [CrossRef]
- Rastian, Z.; Pütz, S.; Wang, Y.J.; Kumar, S.; Fleissner, F.; Weidner, T.; Parekh, S.H. Type I Collagen from Jellyfish Catostylus mosaicus for Biomaterial Applications. ACS Biomater. Sci. Eng. 2018, 4, 2115–2125. [Google Scholar] [CrossRef]
- Di Benedetto, C.; Barbaglio, A.; Martinello, T.; Alongi, V.; Fassini, D.; Cullorà, E.; Patruno, M.; Bonasoro, F.; Barbosa, M.A.; Carnevali, M.D.C.; et al. Production, characterization and biocompatibility of marine collagen matrices from an alternative and sustainable source: The sea urchin Paracentrotus lividus. Mar. Drugs 2014, 12, 4912–4933. [Google Scholar] [CrossRef]
- Coppola, D.; Oliviero, M.; Vitale, G.A.; Lauritano, C.; D’Ambra, I.; Iannace, S.; de Pascale, D. Marine collagen from alternative and sustainable sources: Extraction, processing and applications. Mar. Drugs 2020, 18, 214. [Google Scholar] [CrossRef]
- Zhong, Y.; Khan, M.A.; Shahidi, F. Compositional characteristics and antioxidant properties of fresh and processed sea cucumber (cucumaria frondosa). J. Agric. Food Chem. 2007, 55, 1188–1192. [Google Scholar] [CrossRef]
- de Castro Santana, R.; Kawazoe Sato, A.C.; Lopes da Cunha, R. Emulsions stabilized by heat-treated collagen fibers. Food Hydrocoll. 2012, 26, 73–81. [Google Scholar] [CrossRef]
- Gennadios, A.; Hanna, M.A.; Kurth, L.B. Application of edible coatings on meats, poultry and seafoods: A review. LWT Food Sci. Technol. 1997, 30, 337–350. [Google Scholar] [CrossRef]
- Ruban, S. Biobased Packaging—Application in Meat Industry. Vet. World 2009, 2, 79. [Google Scholar] [CrossRef]
- Technology, E. Effects of fish collagen hydrolysate (FCH) as fat replacer in the production of buffalo patties. J. Adv. Res. Appl. Sci. Eng. Technol. 2018, 11, 108–117. [Google Scholar]
- Le, A.; Morales-peñaloza, A.; Mart, M. Hydrolyzed collagen-sources and applications. Molecules 2019, 24, 4031. [Google Scholar] [CrossRef]
- Bilek, S.E.; Bayram, S.K. Fruit juice drink production containing hydrolyzed collagen. J. Funct. Foods 2015, 14, 562–569. [Google Scholar] [CrossRef]
- Alves, A.L.; Marques, A.L.P.; Martins, E.; Silva, T.H.; Reis, R.L. Cosmetic potential of marine fish skin collagen. Cosmetics 2017, 4, 39. [Google Scholar] [CrossRef]
- Kim, S.J.; Park, S.Y.; Hong, S.M.; Kwon, E.H.; Lee, T.K. Skin whitening and anti-corrugation activities of glycoprotein fractions from liquid extracts of boiled sea cucumber. Asian Pac. J. Trop. Med. 2016, 9, 1002–1006. [Google Scholar] [CrossRef]
- Kupper, S.; Kłosowska-Chomiczewska, I.; Szumała, P. Collagen and hyaluronic acid hydrogel in water-in-oil microemulsion delivery systems. Carbohydr. Polym. 2017, 175, 347–354. [Google Scholar] [CrossRef]
- Silva, T.H.; Moreira-Silva, J.; Marques, A.L.P.; Domingues, A.; Bayon, Y.; Reis, R.L. Marine origin collagens and its potential applications. Mar. Drugs 2014, 12, 5881–5901. [Google Scholar] [CrossRef]
- Schmidt, M.M.; Dornelles, R.C.P.; Mello, R.O.; Kubota, E.H.; Mazutti, M.A.; Kempka, A.P.; Demiate, I.M. Collagen extraction process. Int. Food Res. J. 2016, 23, 913–922. [Google Scholar]
- Yang, H.; Shu, Z. The extraction of collagen protein from pigskin. J. Chem. Pharm. Res. 2014, 6, 683–687. [Google Scholar]
- Blanco, M.; Vázquez, J.A.; Pérez-Martín, R.I.; Sotelo, C.G. Collagen extraction optimization from the skin of the small-spotted catshark (S. Canicula) by response surface methodology. Mar. Drugs 2019, 17, 40. [Google Scholar] [CrossRef] [PubMed]
- Yd, S.; Em, A.; Yusoff, A.; Ssa, H.; Ty, N.; Mys, A. Extraction, Purification and physical characterization of collagen from body wall of sea cucumber bohadschia bivitatta. Heal. Environ. J. 2013, 4, 53–65. [Google Scholar]
- Benjakul, S.; Nalinanon, S.; Shahidi, F. Fish Collagen. In Food Biochemistry and Food Processing, 2nd ed.; Simpson, B.K., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2012; pp. 365–387. [Google Scholar] [CrossRef]
- Hukmi, N.M.M.; Sarbon, N.M. Isolation and characterization of acid soluble collagen (ASC) and pepsin soluble collagen (PSC) extracted from silver catfish (pangasius sp.) skin. Int. Food Res. J. 2018, 25, 2601–2607. [Google Scholar]
- Hadfi, N.H.; Sarbon, N.M. Influenced By Acetic Acid Concentration. Food Res. 2019, 3, 783–790. [Google Scholar] [CrossRef]
- Ran, X.G.; Wang, L.Y. Use of ultrasonic and pepsin treatment in tandem for collagen extraction from meat industry by-products. J. Sci. Food Agric. 2014, 94, 585–590. [Google Scholar] [CrossRef]
- Wang, L.; Yang, B.; Du, X.; Yang, Y.; Liu, J. Optimization of conditions for extraction of acid-soluble collagen from grass carp (ctenopharyngodon idella) by response surface methodology. Innov. Food Sci. Emerg. Technol. 2008, 9, 604–607. [Google Scholar] [CrossRef]
- Liu, D.; Nikoo, M.; Boran, G.; Zhou, P.; Regenstein, J.M. Collagen and gelatin. Annu. Rev. Food Sci. Technol. 2015, 6, 527–557. [Google Scholar] [CrossRef]
- de Moraes, M.C.; Cunha, R.L. Gelation property and water holding capacity of heat-treated collagen at different temperature and pH values. Food Res. Int. 2013, 50, 213–223. [Google Scholar] [CrossRef]
- Wu, S.-L.; Kang, H.-B.; Li, D.-J. Technology for extracting effective components from fish scale. J. Food Sci. Eng. 2017, 7, 351–358. [Google Scholar] [CrossRef]
- Chang, H.J.; Xu, X.L.; Zhou, G.H.; Li, C.B.; Huang, M. Effects of characteristics changes of collagen on meat physicochemical properties of beef semitendinosus muscle during ultrasonic processing. Food Bioprocess Technol. 2012, 5, 285–297. [Google Scholar] [CrossRef]
- Kim, H.K.; Kim, Y.H.; Park, H.J.; Lee, N.H. Application of ultrasonic treatment to extraction of collagen from the skins of sea bass lateolabrax japonicus. Fish. Sci. 2013, 79, 849–856. [Google Scholar] [CrossRef]
- Ali, A.M.M.; Kishimura, H.; Benjakul, S. Extraction efficiency and characteristics of acid and pepsin soluble collagens from the skin of golden carp (probarbus Jullieni) as affected by ultrasonication. Process Biochem. 2018, 66, 237–244. [Google Scholar] [CrossRef]
- Khong, N.M.H.; Yusoff, F.M.; Jamilah, B.; Basri, M.; Maznah, I.; Chan, K.W.; Armania, N.; Nishikawa, J. Improved collagen extraction from jellyfish (acromitus hardenbergi) with increased physical-induced solubilization processes. Food Chem. 2018, 251, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, M.; Yi, R.; Bai, K.; Wang, G.; Tan, R.; Sun, S.; Xu, N. Electrodialysis extraction of pufferfish skin (takifugu flavidus): A promising source of collagen. Mar. Drugs 2019, 17, 25. [Google Scholar] [CrossRef]
- Yan, L.J.; Zhan, C.L.; Cai, Q.F.; Weng, L.; Du, C.H.; Liu, G.M.; Su, W.J.; Cao, M.J. Purification, characterization, cdna cloning and in vitro expression of a serine proteinase from the intestinal tract of sea cucumber (stichopus japonicus) with collagen degradation activity. J. Agric. Food Chem. 2014, 62, 4769–4777. [Google Scholar] [CrossRef]
- Kadam, S.U.; Tiwari, B.K.; Álvarez, C.; O’Donnell, C.P. Ultrasound applications for the extraction, identification and delivery of food proteins and bioactive peptides. Trends Food Sci. Technol. 2015, 46, 60–67. [Google Scholar] [CrossRef]
- Song, K.M.; Jung, S.K.; Kim, Y.H.; Kim, Y.E.; Lee, N.H. Development of industrial ultrasound system for mass production of collagen and biochemical characteristics of extracted collagen. Food Bioprod. Process. 2018, 110, 96–103. [Google Scholar] [CrossRef]
- Li, D.; Mu, C.; Cai, S.; Lin, W. Ultrasonic irradiation in the enzymatic extraction of collagen. Ultrason. Sonochem. 2009, 16, 605–609. [Google Scholar] [CrossRef]
- Lin, Y.J.; Le, G.W.; Wang, J.Y.; Li, Y.X.; Shi, Y.H.; Sun, J. Antioxidative peptides derived from enzyme hydrolysis of bone collagen after microwave assisted acid pre-treatment and nitrogen protection. Int. J. Mol. Sci. 2010, 11, 4297–4308. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; An, B.; Ramshaw, J.A.M.; Brodsky, B. Bacterial collagen-like proteins that form triple-helical structures. J. Struct. Biol. 2014, 186, 451–461. [Google Scholar] [CrossRef]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Application of novel extraction technologies for bioactives from marine algae. J. Agric. Food Chem. 2013, 61, 4667–4675. [Google Scholar] [CrossRef]
- Gómez-Guillén, M.C.; Giménez, B.; Montero, P. Extraction of gelatin from fish skins by high pressure treatment. Food Hydrocoll. 2005, 19, 923–928. [Google Scholar] [CrossRef]
- Fernández-Martín, F.; Pérez-Mateos, M.; Montero, P. Effect of Pressure/Heat Combinations on blue whiting (micromesistius poutassou) washed mince: Thermal and mechanical properties. J. Agric. Food Chem. 1998, 46, 3257–3264. [Google Scholar] [CrossRef]
- Blanco, M.; Sanz, N.; Valcarcel, J.; Pérez-Martín, R.I.; Sotelo, C.G. Does subunit composition influence the intermolecular crosslinking of fish collagen? A study with hake and blue shark skin collagens. Polymers 2020, 12, 1734. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Chance, R.R.; Graessley, W.W.; Lohse, D.J. A study of the separation principle in size exclusion chromatography. Macromolecules 2004, 37, 4304–4312. [Google Scholar] [CrossRef]
- Fekete, S.; Goyon, A.; Veuthey, J.L.; Guillarme, D. Size exclusion chromatography of protein biopharmaceuticals: Past, present and future. Am. Pharm. Rev. 2018, 21, 1–4. [Google Scholar]
- Naffa, R.; Edwards, P.J.B.; Norris, G. Isolation and characterization of collagen type I crosslink from skin: High-resolution NMR reveals diastereomers of hydroxylysinonorleucine crosslink. Amino Acids 2019, 51, 705–715. [Google Scholar] [CrossRef]
- Fekete, S.; Beck, A.; Veuthey, J.L.; Guillarme, D. Ion-exchange chromatography for the characterization of biopharmaceuticals. J. Pharm. Biomed. Anal. 2015, 113, 43–55. [Google Scholar] [CrossRef]
- Chow, W.Y. Investigation of triple-helix collagen hydroxylation by solid-state NMR spectroscopy. Methods Mol. Biol. 2019, 1944, 57–77. [Google Scholar] [CrossRef] [PubMed]
- Mocan, E.; Tagadiuc, O.; Nacu, V. Aspects of collagen isolation procedure. Clin. Res. Stud. 2011, 8–10. [Google Scholar]
- Liu, Z.Q.; Mahmood, T.; Yang, P.C. Western blot: Technique, theory and trouble shooting. N. Am. J. Med. Sci. 2014, 6, 160. [Google Scholar] [CrossRef] [PubMed]
- Cliche, S.; Amiot, J.; Avezard, C.; Gariépy, C. Extraction and characterization of collagen with or without telopeptides from chicken skin. Poult. Sci. 2003, 82, 503–509. [Google Scholar] [CrossRef]
- Quiones, J.L.; Rosa, R.; Ruiz, D.L. Extracellular matrix remodeling and metalloproteinase involvement during intestine regeneration in the sea cucumber Holothuria glaberrima. Dev. Biol. 2002, 250, 181–197. [Google Scholar] [CrossRef]
- Hong, H.; Fan, H.; Chalamaiah, M.; Wu, J. Preparation of low-molecular-weight, collagen hydrolysates (peptides): Current progress, challenges, and future perspectives. Food Chem. 2019, 301, 125222. [Google Scholar] [CrossRef]
- Magdeldin, S.; Enany, S.; Yoshida, Y.; Xu, B.; Zhang, Y.; Zureena, Z.; Lokamani, I.; Yaoita, E.; Yamamoto, T. Basics and recent advances of two dimensional-polyacrylamide gel electrophoresis. Clin. Proteomics 2014, 11, 1–10. [Google Scholar] [CrossRef]
- Domon, B.; Aebersold, R. Mass spectrometry and protein analysis. Science 2006, 312, 212–217. [Google Scholar] [CrossRef]
- Eyre, D.R.; Weis, M.A.; Wu, J.J. Advances in collagen cross-link analysis. Methods 2008, 45, 65–74. [Google Scholar] [CrossRef]
- Heu, M.S.; Lee, J.H.; Kim, H.J.; Jee, S.J.; Lee, J.S.; Jeon, Y.J.; Shahidi, F.; Kim, J.S. Characterization of acid- and pepsin-soluble collagens from flatfish skin. Food Sci. Biotechnol. 2010, 19, 27–33. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, J.; Zhao, D.; Xue, C.; Liu, Z.; Mao, X. Characterization of turbot (scophthalmus maximus) skin and the extracted acid-soluble collagen. J. Ocean Univ. China 2019, 18, 687–692. [Google Scholar] [CrossRef]
- Hofman, K.; Hall, B.; Cleaver, H.; Marshall, S. High-throughput quantification of hydroxyproline for determination of collagen. Anal. Biochem. 2011, 417, 289–291. [Google Scholar] [CrossRef] [PubMed]
- Sirbu, R.; Stanciu, G.; Cadar, E.; Tomescu, A.; Cherim, M. Validation of a quantitative analysis method for collagen extracted from grey mullet marine fish. Rev. Chim. 2019, 70, 835–842. [Google Scholar] [CrossRef]
- Sadowska, M.; Kołodziejska, I.; Niecikowska, C. Isolation of collagen from the skins of Baltic cod (gadus morhua). Food Chem. 2003, 81, 257–262. [Google Scholar] [CrossRef]
- Shahidi, F.; Synowiecki, J. Protein hydrolyzates from seal meat as phosphate alternatives in food processing applications. Food Chem. 1997, 60, 29–32. [Google Scholar] [CrossRef]
- Erge, A.; Zorba, Ö. Optimization of gelatin extraction from chicken mechanically deboned meat residue using alkaline pre-treatment. Lwt 2018, 97, 205–212. [Google Scholar] [CrossRef]
- Zayas, J.F. Emulsifying Properties of proteins. Functionality of Proteins in Food; Springer: Berlin/Heidelberg, Germany, 1997; pp. 134–227. [Google Scholar] [CrossRef]
- Du, Y.N.; Guo, X.K.; Han, Y.T.; Han, J.R.; Yan, J.N.; Shang, W.H.; Wu, H.T. Physicochemical and functional properties of protein isolate from sea cucumber (stichopus japonicus) guts. J. Food Process. Preserv. 2019, 43, 1–11. [Google Scholar] [CrossRef]
- Abdelmalek, B.E.; Gómez-Estaca, J.; Sila, A.; Martinez-Alvarez, O.; Gómez-Guillén, M.C.; Chaabouni-Ellouz, S.; Ayadi, M.A.; Bougatef, A. Characteristics and functional properties of gelatin extracted from squid (loligo vulgaris) skin. LWT Food Sci. Technol. 2016, 65, 924–931. [Google Scholar] [CrossRef]
- Bhuimbar, M.V.; Bhagwat, P.K.; Dandge, P.B. Extraction and characterization of acid soluble collagen from fish waste: Development of collagen-chitosan blend as food packaging film. J. Environ. Chem. Eng. 2019, 7, 102983. [Google Scholar] [CrossRef]
- Nor, M.H.M.; Nazmi, N.N.M.; Sarbon, N.M. Effects of plasticizer concentrations on functional properties of chicken skin gelatin films. Int. Food Res. J. 2017, 24, 1910–1918. [Google Scholar]
- Slimane, E.B.; Sadok, S. Collagen from cartilaginous fish by-products for a potential application in bioactive film composite. Mar. Drugs 2018, 16, 211. [Google Scholar] [CrossRef] [PubMed]
- Avena-Bustillos, R.J.; Olsen, C.W.; Olson, D.A.; Chiou, B.; Yee, E.; Bechtel, P.J.; McHugh, T.H. Water vapor permeability of mammalian and fish gelatin films. J. Food Sci. 2006, 71, 202–207. [Google Scholar] [CrossRef]
- Muyonga, J.H.; Cole, C.G.B.; Duodu, K.G. Fourier transform infrared (FTIR) spectroscopic study of acid soluble collagen and gelatin from skins and bones of young and adult Nile perch (lates niloticus). Food Chem. 2004, 86, 325–332. [Google Scholar] [CrossRef]
- Teixeira, B.; Marques, A.; Pires, C.; Ramos, C.; Batista, I.; Saraiva, J.A.; Nunes, M.L. Characterization of fish protein films incorporated with essential oils of clove, garlic and origanum: Physical, antioxidant and antibacterial properties. LWT-Food Sci. Technol. 2014, 59, 533–539. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Collagen Type | Chains | Sub Family | Distribution |
|---|---|---|---|
| I | α1(I) α2(I) | Fibrillar collagen | Skin, tendon, bone, dermis, intestine, uterus |
| II | α1(II) | Fibrillar collagen | Hyaline cartilage, vitreous, nucleus pulposus |
| III | α1(III) | Fibrillar collagen | Dermis, intestine, large vessels, heart valve |
| IV | α1(IV) α2(IV) α3(IV) α4(IV) α5(IV) α6(IV) | Basement membrane and associated collagen | Basement membranes |
| V | α1(V) α2(V) α3(V) | Fibrillar collagen | Cornea, placental membranes, bone, large vessels |
| VI | α1(VI) α2(VI) α3(VI) | Beaded filament forming collagen | Descement’s membrane, skin, heart muscles |
| VII | α1(VII) | Basement membrane and associated collagen | Skin, placenta, lung, cartilage, cornea |
| VIII | α1(VIII) α2(VIII) | Short chain collagen | Produced by endothelial cells, descemet’s membrane |
| IX | α1(IX) α2(IX) α3(IX) | Fibril associated and related collagen | Cartilage |
| X | α1(X) | Short chain collagen | Hypertrophic and mineralizing cartilage |
| XI | α1(XI) α2(XI) α3(XI) | Fibrillar collagen | Cartilage, intervertebral disc, vitreous humor |
| XII | α1(XII) | Fibril associated and related collagen | Chicken embryo tendon, bovine periodontal ligament |
| XIII | α1(XIII) | Trans membrane collagens and collagen like proteins | Cetal skin, bone, intestinal mucosa |
| Source | Extraction Method | Purpose of Extraction | Reference |
|---|---|---|---|
| Chicken feet | Acid extraction | Optimization of extraction condition | [43] |
| Enzyme extraction | Determination of pepsin digestion effect on the properties of extracted collagen | [48] | |
| Acid extraction | Preparation of edible films | [57] | |
| Enzyme extraction (using papain and pepsin) | Isolation and characterization of chicken feet originated collagen | [58] | |
| Acid extraction | Use of chicken feet for protein films | [59] | |
| Alkali, acid, and enzyme extraction | Identification of best method of collagen extraction method and characterization of chicken feet collagen | [50] | |
| Enzyme extraction | Optimization of extraction process and synthesis of chicken feet collagen based biopolymeric fibers | [60] | |
| Rat tail tendon | Acid extraction | Preparation of type I collagen for tissue engineering applications | [45] |
| Alligator bone | Acid and enzyme assisted extraction | Determination of biochemical properties of alligator bone collagen | [47] |
| Silky fowl feet | Combination of acid and enzyme extraction | Identification of best combination for high quality collagen extraction method | [49] |
| Ovine tendon | Acid extraction | Determination of the biocompatibility of ovine tendon originated collagen with human dermal fibroblast | [51] |
| Acid extraction | Determination of the biocompatibility of ovine tendon originated collagen with human dermal fibroblast Improve the mechanically strong ovine tendon originated collagen for tissue engineering purposes | [52] | |
| Acid extraction | Characterization and fabrication of thin films from ovine tendon collagen for tissue engineering applications | [53] | |
| Acid extraction | Investigation of attachment, proliferation, and morphological properties of human dermal fibroblasts on ovine tendon collagen | [54] | |
| Duck feet | Acid extraction | Investigation of physicochemical properties of collagen derived from duck feet | [46] |
| Acid extraction | Determination of feasibility of using duck feet collagen in improving physicochemical properties of surimi | [61] | |
| Kangaroo tail | Acid extraction | Identification of alternative collagen sources for pre-clinical models for cell biology | [44] |
| Sheep bone | Acid extraction | Determination of effect of different collagen extraction protocols | [55] |
| Equine tendon | Acid extraction | Evaluation of the effects of different extraction methods on the collagen structure of equine tendons | [33] |
| Sea Cucumber Species | Focus of Study | Major Findings | Reference |
|---|---|---|---|
| Stichopus japonicus | Chemical composition and subunit structure of collagen | Collagen was comprised of 2 distinct subunits (α1 and α2 and rich in glutamic acid compared to other fibrillar collagen | [14] |
| Characterization and subunit composition of collagen | Pepsin solubilized collagen resembled type I collagen and its amino acid composition was different from vertebrate collagen. | [16] | |
| Changes of collagen during cooking | Crude collagen fibers were more susceptible to heat treatment compared to pepsin-solubilized collagen | [115] | |
| Identification of physicochemical properties and radical scavenging capacities of pepsin-solubilized collagen | Extracted collagen maintained intact triple-stranded helices and high moisture retention and absorption capacities as well as exhibiting better radical scavenging ability compared to vitamins C and E. | [95] | |
| Wound-healing effects on human keratinocyte (HaCaT) cell line of pepsin-solubilized collagen | Pepsin-solubilized collagen has the potential to use as an alternative mammalian collagen in the nutraceutical and pharmaceutical industries | [96] | |
| Effect of autolysis of intact collagen fibers related to the distributions of cathepsin L | Lysosomal cathepsin L degrades the collagen fibers and speed and degree of autolysis is negatively correlated with the density of collagen. | [116] | |
| Structural characteristics of sea cucumber collagen fibers in the presence of endogenous cysteine proteinases | Collagen fibrils disaggregated into collagen fibrils by cysteine proteinases and the structural disorder of the native collagen fibers increased due to cysteine protease. | [102] | |
| Structural and biochemical changes of collagen related to autolysis | Collagen fibers and microfibrils gradually degraded with the autolysis and structural damage was less in monomeric collagen compared to other structural elements | [103] | |
| Structural and thermal properties of sea cucumber collagen | Distance between adjacent molecular chains in collagen molecules was decreased and CO2, NH3, H2O, CH4, NO2 and HCN gases released during the heat treatment | [117] | |
| Enzymatic hydrolysis of collagen to determine the structural changes of collagen fibrils | Collagen fibers were partially degraded into collagen fibrils by enzymatic (trypsin) treatments | [118] | |
| Investigate the effect of collagenase type I on the structural features of collagen fibers | Collagenase was responsible for partial depolymerization of collagen fibers into fibrils, uncoiled the fibrils, degrade monomeric collagen | [119] | |
| Parastichopus californicus | Purification and characterization of pepsin-solubilized collagen from skin and connective tissue | Collagen extracted from skin and connective tissue contains type I collagen with three α1 chain. Amino acid composition is different from the mammalian type I collagen | [120] |
| Bohadschia spp. | Analysis of isolated pepsin-solubilized collagen | Type I collagen was identified with three α1 chain | [121] |
| Stichopus vastus | Isolation and characterization of pepsin-solubilized collagen | Purified collagen belongs to type I collagen contains three α1 chain with triple helical structure | [99] |
| Molecular mass distribution, amino acid composition and radical-scavenging activity of collagen hydrolysates prepared from isolated collagen | β and α1 chains of the collagen were hydrolyzed by trypsin and molecular mass distribution ranged from 5 to 25 kDa. Hydrolysates contains high glycine, alanine, glutamate, proline and hydroxyproline residues and showed significant radical scavenging activity | [122] | |
| Physicochemical and biochemical properties of pepsin solubilized collagen | Glycine was the predominant amino acid present in purified collagen that possessed high moisture absorption and retention capacity | [122] | |
| Identification of Angiotensin I converting enzyme (ACE) inhibitory and radical scavenging activities from collagen hydrolysates | Novel bioactive peptides generated by optimized trypsin hydrolysis have the potential to use as ACE inhibitors and radical scavenging agents. | [100] | |
| Holothuria parva | Purification and characterization of pepsin-solubilized collagen | Isolated collagen constituted three α1 chain and was rich in glycine, proline, alanine and hydroxyproline | [98] |
| Stichopus monotuberculatus | Isolation and characterization of pepsin-solubilized collagen | Isolated collagen was classified as type I collagen consisted of three α1 chain | [101] |
| Holothuria scabra | Determination of nano-collagen quality and extraction of acid solubilized collagen | Extracted acid solubilized collagen had significant effect on physicochemical characteristics of nano-collagen particles | [123] |
| Australostichopus mollis | Biochemical composition of isolated collagen | Type I collagen was present with α1 and α2 chains, α chain dimers, β chains, and γ components. Most abundant amino acids were glycine, alanine, threonine, serine, and proline. | [124] |
| Holothuria leucospilota | In vitro activity of anti-tyrosinase and anti-elastase activity of isolated collagen | Isolated collagen exhibited weak anti-tyrosine activity and moderate anti-elastase activity | [125] |
| Acaudina leucoprocta | Extraction methods to remove heavy metals from the isolated collagen | Pepsi- solubilized collagen showed two isoforms and amount of heavy metals present in the collagen were below the contaminant limit | [126] |
| Acaudina molpadioides | Preparation and characterization of antioxidative peptides from collagen hydrolysates | Collagen peptides which showed highest antioxidant activity were rich in hydrophobic acid residues. | [127] |
| Stichopus vastus and Holothuria atra | Comparison of partial characteristics of two different sea cucumbers | No significant difference in amino acid composition, yield, or whiteness | [128] |
| Apostichopus japonicus | Type of constituent collagen using proteomics and bioinformatic strategies | Heterogenicity of the sea cucumber collagen fibrils was revealed for the first time that provides novel insight into the composition of sea cucumber collagen | [104] |
| Analysis of the effect of epigallocatechin gallate (EGCG) on preserving molecular structure of collagen fibers during heating | EGCG protects the structure of crude collagen fibers in a dosage dependent manner and effects hydrogen bonds on the collagen which promotes protein aggregation | [105] | |
| Holothuria cinerascens | Potential application of collagen in moisturizing cosmetics | Collagen showed better moisture retention and moisture absorption capacity. Abundant hydrophilic groups in collagen increases their ability for cosmetic formulations | [83] |
| Characteristics | Sea Cucumber Derived Collagen | Mammalian Collagen | Reference |
|---|---|---|---|
| Abundant type | Type I collagen | Type I collagen | [14,18,113] |
| Differences in amino acid composition | Low hydroxyproline content, high glutamic and aspartic acid residues | High hydroxyproline content, low glutamic acid and aspartic acid residues | [14,16,99,101,120] |
| Covalent cross links | Internally present and provide stabilization to the molecule | Internally present and provide stabilization to the molecule | [109,110,111] |
| Thermal stability | Low thermal stability with low denaturation temperature compared to mammalian collagen | High thermal stability compared with high denaturation temperatures | [98,101,120,122] |
| Resistance to protease digestion | Relatively low | Relatively high | [99] |
| Gel forming ability | Comparatively low | Comparatively high | [99] |
| Moisture absorption ability | Relatively high | Relatively high | [8,95] |
| Sea Cucumber Species | Body Parts | Pre-Treatment | Methods Used for Characterization of Collagen | Reference |
|---|---|---|---|---|
| Cucumaria frondosa | Inner dermis | Incubation with deionized water | Amino acid analysis SDS-PAGE Salt solubility determination | [109] |
| Stichopus japonicus | Body wall | Disaggregation with β-mercaptoethanol and 0.1 M NaOH treatment | Amino acid analysis SDS-PAGE DSC | [14] |
| Body wall | Incubation with water | Ultraviolet-visible (UV-vis) spectra SDS-PAGE Peptide mapping Amino acid composition DSC Gel filtration chromatography | [16] | |
| Stichopus vastus | Integument | Incubated with water | UV-vis spectra SDS-PAGE peptide mapping FTIR Gel forming capacity | [99] |
| Bohadshia spp. | Body wall | Washed in distilled water | SDS-PAGE | [165] |
| Holothuria parva | Skin | Washed in distilled water | SDS-PAGE DSC Gel-forming capacity UV-vis spectra Amino acid composition Scanning electron microscopy | [98] |
| Stichopus monotuberculatus | Body wall | Homogenization with water | UV-vis spectra SDS-PAGEA mino acid analysis FTIR Enzyme-digested peptide mapping DSC Solubility level | [142] |
| Parastichopus californicus | Skin and connective tissue | Washed in distilled water | DSC SDS-PAGE Enzyme-digested peptide mapping Gel-forming capability Amino acid composition | [120] |
| Australostichopus mollis | Body wall | Washed in distilled water | Scanning electron microscopy Electrophoretic analysis Peptide mapping UV-vis spectra DSC FTIR Amino acid analysis | [124] |
| Acaudina molpadioides | Body wall | Soaked in 0.2 M EDTA for 48 h | Gel-filtration chromatography Amino acid analysis RP-HPLC and identification of peptide sequence | [127] |
| Sea Cucumber Type | SDS Gel Composition | Collagen Type and Subunit Composition | Findings | Reference |
|---|---|---|---|---|
| Cucumaria frondosa | Linear polyacrylamide gradients of 4–20%, and 100 mM Tris, 3.3% SDS, 20% glycerol | Type I collagen (α1)3 | Covalent composition of collagen is α1 trimer and amino acid composition is similar to human collagen type I | [109] |
| Stichopus japonicus | Consisted with 9% polyacrylamide gels | Type I collagen, consisting of 1 α trimer (approximately 135 kDa) | Subunit structure of isolated collagen is similar to (α1)3 pattern that exists in the invertebrate collagen | [16] |
| Parastichopus californicus | Discontinuous Tris-HCl/glycine buffer system with 7.5% resolving gel and 4% stacking gel | Type I collagens, consisting of three α1 chains of approximately 138 kDa | Isolated collagen constituents were α1 and β dimers and similar to that reported for collagens from other sea cucumber species | [120] |
| Stichopus japonicus | Discontinuous Tris-HCl/glycine buffer system with 10% separating gel and a 5% stacking gel | Type I collagens, consisting of 1 α trimer | Electrophoresis pattern demonstrated a major single band on SDS-PAGE | [105] |
| Stichopus vastus | Discontinuous Tris-HCl-glycine buffer system with 75 g L−1 resolving gel and 40 g L−1 stacking gel | Type I collagen, consisting of three α1 chains of approximately 122 kDa each | Isolated collagen was consisted with major component (α1) of approximately 122 kDa and a small amount of β dimers (about 267 kDa each) similar to that reported for collagen from other sea cucumber species | [99] |
| Bohadschia spp. | Discontinuous Tris-HCl-glycine buffer system with 7.5% resolving gel and 4% stacking gel | Type I collagen with three α1 chains with approximately 138 kDa each | Collagen was formed with major component of α1 and smaller amount of β dimer | [165] |
| Stichopus monotuberculatus | Discontinuous tris-glycine buffer system electrophoresis with 7.5% precast gel | Type I collagen consists of three α1 with molecular weight of 137 kD | Collagen consisted of 3 homologous α1 chains as (α1)3. The molecular weight of isolated collagen was similar to the reported values of collagens from other species | [101] |
| Australostichopus mollis | Not included in detail | Type I collagens consist of α1 and α2 chains (approximately 116 kDa) | Collagen formed α1 and α2 chains with α chains dimer, β chains (around 212 kDa) and small amounts of γ components and electrophoresis pattern was similar to those of calf skin collagen | [124] |
| Holothuria cinerascens | 10% SDS separating gel and 5% stacking gel | Type I collagen with identical α1 chains (α1, α2 and α3) | Molecular weight of isolated α chains extracted was about 80–90 kDa, and the molecular weight of the β-chain was about 150–160 kDa. The reported molecular weights were significantly lower than those of tilapia and porcine skin collagen | [83] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Senadheera, T.R.L.; Dave, D.; Shahidi, F. Sea Cucumber Derived Type I Collagen: A Comprehensive Review. Mar. Drugs 2020, 18, 471. https://doi.org/10.3390/md18090471
Senadheera TRL, Dave D, Shahidi F. Sea Cucumber Derived Type I Collagen: A Comprehensive Review. Marine Drugs. 2020; 18(9):471. https://doi.org/10.3390/md18090471
Chicago/Turabian StyleSenadheera, Tharindu R.L., Deepika Dave, and Fereidoon Shahidi. 2020. "Sea Cucumber Derived Type I Collagen: A Comprehensive Review" Marine Drugs 18, no. 9: 471. https://doi.org/10.3390/md18090471
APA StyleSenadheera, T. R. L., Dave, D., & Shahidi, F. (2020). Sea Cucumber Derived Type I Collagen: A Comprehensive Review. Marine Drugs, 18(9), 471. https://doi.org/10.3390/md18090471