Collagen Peptides from Swim Bladders of Giant Croaker (Nibea japonica) and Their Protective Effects against H2O2-Induced Oxidative Damage toward Human Umbilical Vein Endothelial Cells

Abstract

1. Introduction
2. Results and Discussion
2.1. Single Factor Results
2.1.1. Selection of the Optimal Enzyme
2.1.2. Single Factor Experiments
2.2. Optimization of Extraction Parameters by Response Surface Methodology (RSM)
2.2.1. Response Surface Analysis
2.2.2. Validation of the Models
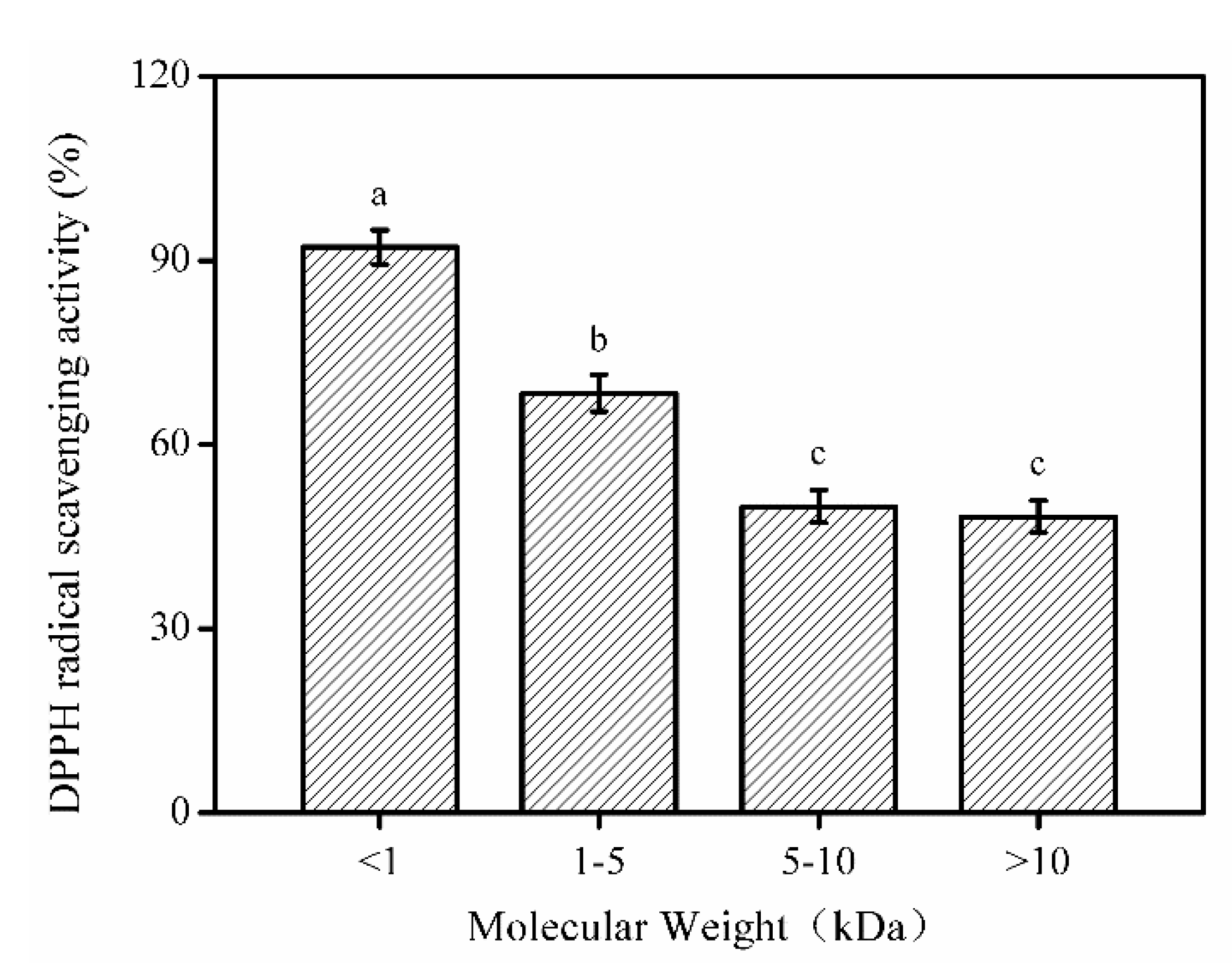
2.3. Molecular Weight Distribution of SNNHs
2.4. Fractionation of SNNH-1 from SNNHs
2.5. Amino Acid Composition of SNNH-1
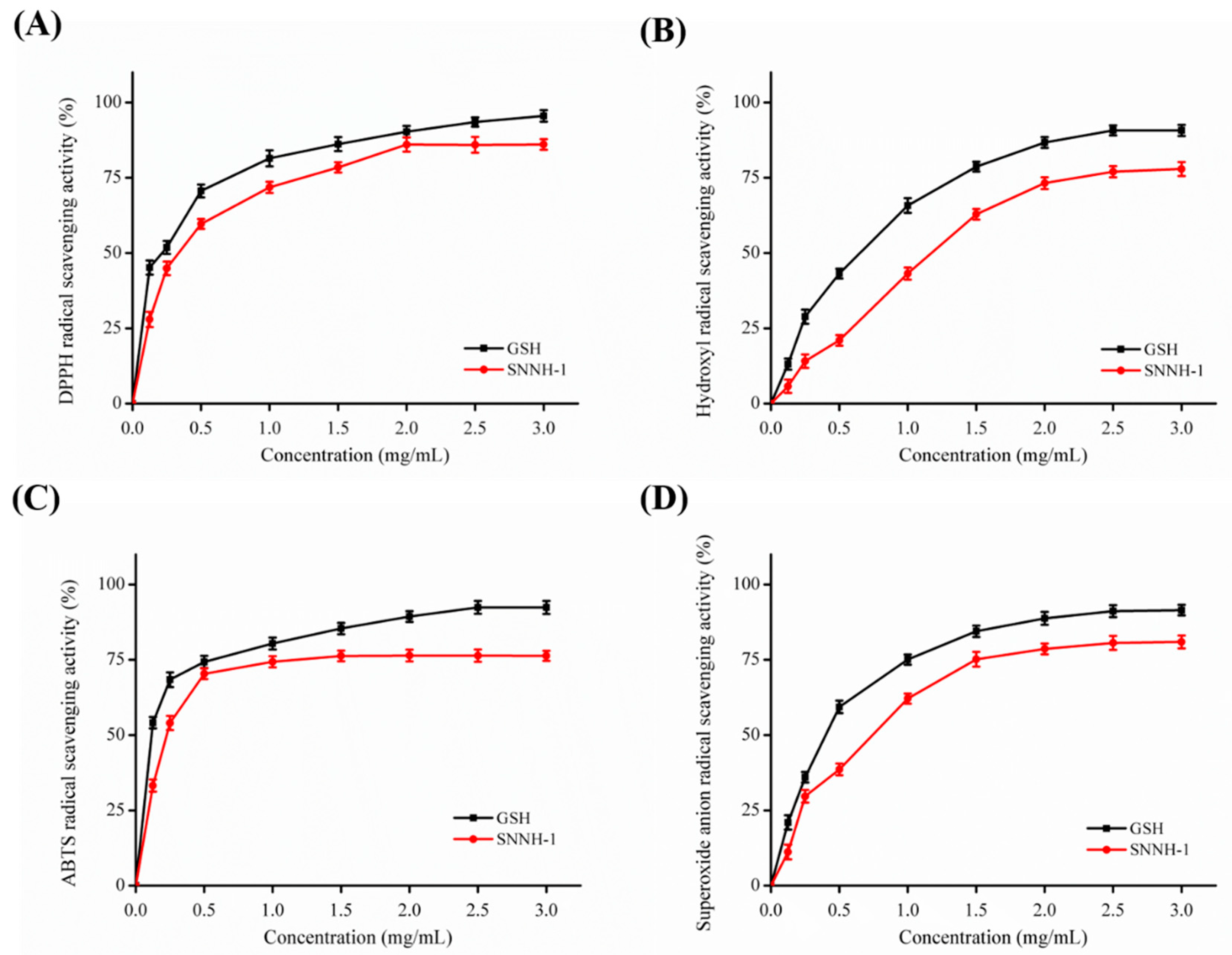
2.6. Antioxidant Activity of SNNH-1
2.7. Cytotoxic and Allergenic Potential of SNNH-1
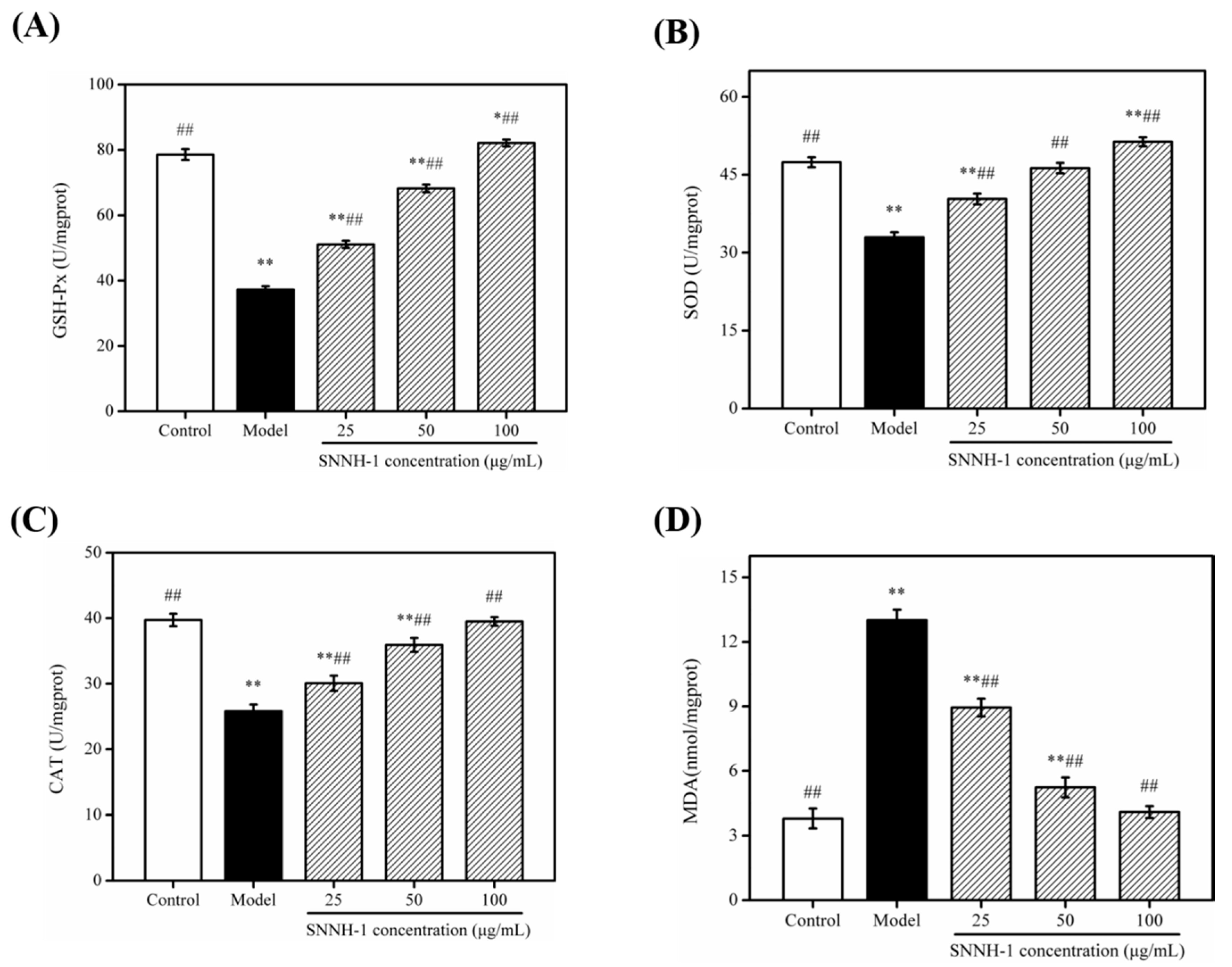
2.8. Effects of SNNH-1 on the Levels of GSH-Px, SOD, CAT and Malondialdehyde (MDA) in an H2O2-Induced HUVECs Injury Model
2.9. Effects of SNNH-1 on ROS Levels in a H2O2-Induced HUVECs Injury Model
3. Materials and Methods
3.1. Materials and Chemicals
3.2. Optimization of Preparative Conditions
3.3. Determination of the Mw Distribution of SNNHs
3.4. Fractionation of SNNH-1 by Ultrafiltration
3.5. Amino Acid Composition Measurement of SNNH-1
3.6. Antioxidant Activity of SNNH-1
3.7. Cytotoxic and Allergenic Properties of SNNH-1
3.8. Assays for Antioxidant Enzymatic Activity of SNNH-1 in H2O2-Induced HUVECs
3.9. Determination of the Levels of ROS in H2O2-Induced HUVECs
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ward, C.W.; Prosser, B.L.; Lederer, W.J. Mechanical Stretch-Induced Activation of ROS/RNS Signaling in Striated Muscle. Antioxid. Redox Signal. 2014, 20, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.B.; Wu, C.; Liu, D.; Yang, X.; Huang, J.; Zhang, J.; Liao, B.; He, H. Antioxidant and anti-freezing peptides from salmon collagen hydrolysate prepared by bacterial extracellular protease. Food Chem. 2018, 248, 342–356. [Google Scholar] [CrossRef] [PubMed]
- Sila, A.; Bougatef, A. Antioxidant peptides from marine by-products: Isolation, identification and application in food systems. J. Funct. Foods 2015, 21, 10–26. [Google Scholar] [CrossRef]
- Jin, Y.; Liu, K.; Peng, J.; Wang, C.; Kang, L.; Chang, N.; Sun, H. Rhizoma Dioscoreae Nipponicae polysaccharides protect HUVECs from H2O2-induced injury by regulating PPARγ factor and the NADPH oxidase/ROS–NF-κB signal pathway. Toxicol. Lett. 2015, 232, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Bougatef, A.; Hajji, M.; Balti, R.; Lassoued, I.; Triki-Ellouz, Y.; Nasri, M. Antioxidant and free radical-scavenging activities of smooth hound (Mustelus mustelus) muscle protein hydrolysates obtained by gastrointestinal proteases. Food Chem. 2009, 114, 1198–1205. [Google Scholar] [CrossRef]
- Zhao, W.H.; Luo, Q.B.; Pan, X.; Chi, C.F.; Sun, K.L.; Wang, B. Preparation, identification, and activity evaluation of ten antioxidant peptides from protein hydrolysate of swim bladders of miiuy croaker (Miichthys miiuy). J. Funct. Foods 2018, 47, 503–511. [Google Scholar] [CrossRef]
- Johansen, J.S.; Harris, A.K.; Rychly, D.J.; Ergul, A. Oxidative stress and the use of antioxidants in diabetes: Linking basic science to clinical pratice. Cardiovasc. Diabetol. 2005, 4, 5–16. [Google Scholar] [CrossRef]
- Huang, Y.; Ruan, G.; Qin, Z.; Li, H.; Zheng, Y. Antioxidant activity measurement and potential antioxidant peptides exploration from hydrolysates of novel continuous microwave-assisted enzymolysis of the Scomberomorus niphonius protein. Food Chem. 2017, 223, 89–95. [Google Scholar] [CrossRef]
- Yu, Z.P.; Yin, Y.G.; Zhao, W.Z.; Wang, F.; Yu, Y.D.; Liu, B.Q.; Liu, J.B.; Chen, F. Characterization of ACE-Inhibitory Peptide Associated with Antioxidant and Anticoagulation Properties. J. Food Sci. 2011, 76, 1149–1155. [Google Scholar] [CrossRef]
- Jara, A.M.R.; Liggieri, C.S.; Bruno, M.A. Preparation of soy protein hydrolysates with antioxidant activity by using peptidases from latex of Maclura pomifera fruits. Food Chem. 2018, 246, 326–333. [Google Scholar] [CrossRef]
- Senphan, T.; Benjakul, S. Antioxidative activities of hydrolysates from seabass skin prepared using protease from hepatopancreas of Pacific white shrimp. J. Funct. Foods 2014, 6, 147–156. [Google Scholar] [CrossRef]
- Xia, Y.; Bamdad, F.; Gaenzle, M.; Chen, L. Fractionation and characterization of antioxidant peptides derived from barley glutelin by enzymatic hydrolysis. Food Chem. 2012, 134, 1509–1518. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.F.; Wang, B.; Hu, F.Y.; Wang, Y.M.; Zhang, B.; Deng, S.G.; Wu, C.W. Purification and identification of three novel antioxidant peptides from protein hydrolysate of bluefin leatherjacket (Navodon septentrionalis) skin. Food Res. Int. 2015, 73, 124–129. [Google Scholar] [CrossRef]
- Kumar, N.S.S.; Nazeer, R.A.; Jaiganesh, R. Purification and biochemical characterization of antioxidant peptide from horse mackerel (Magalaspis cordyla) viscera protein. Peptides 2011, 32, 1496–1501. [Google Scholar] [CrossRef]
- You, L.; Zhao, M.; Regenstein, J.M.; Ren, J. In vitro antioxidant activity and in vivo anti-fatigue effect of loach (Misgurnus anguillicaudatus) peptides prepared by papain digestion. Food Chem. 2011, 124, 188–194. [Google Scholar] [CrossRef]
- Ding, J.F.; Li, Y.Y.; Xu, J.J.; Su, X.R.; Gao, X.; Yue, F.P. Study on effect of jellyfish collagen hydrolysate on anti-fatigue and anti-oxidation. Food Hydrocoll. 2011, 25, 1350–1353. [Google Scholar] [CrossRef]
- Ren, J.; Zhao, M.; Wang, H.; Cui, C.; You, L. Effects of supplementation with grass carp protein versus peptide on swimming endurance in mice. Nutrition 2011, 27, 789–795. [Google Scholar] [CrossRef]
- Yu, F.; Zong, C.; Jin, S.; Zheng, J.; Nan, C.; Ju, H.; Yan, C.; Huang, F.; Yang, Z.; Tang, Y. Optimization of Extraction Conditions and Characterization of Pepsin-Solubilised Collagen from Skin of Giant Croaker (Nibea japonica). Mar. Drugs 2018, 16, 29. [Google Scholar] [CrossRef]
- Tang, Y.; Jin, S.; Li, X.; Li, X.; Hu, X.; Yan, C.; Huang, F.; Yang, Z.; Yu, F.; Ding, G. Physicochemical Properties and Biocompatibility Evaluation of Collagen from the Skin of Giant Croaker (Nibea japonica). Mar. Drugs 2018, 16, 222. [Google Scholar] [CrossRef]
- Chen, Y.; Jin, H.; Yang, F.; Jin, S.; Liu, C.; Zhang, L.; Huang, J.; Wang, S.; Yan, Z.; Cai, X.; et al. Physicochemical, antioxidant properties of giant croaker (Nibea japonica) swim bladders collagen and wound healing evaluation. Int. J. Biol. Macromol. 2019, 138, 483–491. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, P.; Zhou, C.; Li, S.; Hong, P. Marine Collagen Peptides from the Skin of Nile Tilapia (Oreochromis niloticus): Characterization and Wound Healing Evaluation. Mar. Drugs 2017, 15, 102–112. [Google Scholar]
- Li, Y.; Li, J.; Lin, S.J.; Yang, Z.S.; Jin, H.X. Preparation of Antioxidant Peptide by Microwave-Assisted Hydrolysis of Collagen and Its Protective Effect Against H2O2-Induced Damage of RAW264.7 Cells. Mar. Drugs 2019, 17, 642. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Zhao, H.; Zhao, Q.; Zhao, M.; Yang, B.; Wu, N.; Qian, Y. Structural characteristics of peptides extracted from Cantonese sausage during drying and their antioxidant activities. Innov. Food Sci. Emerg. Technol. 2009, 10, 558–563. [Google Scholar] [CrossRef]
- Je, J.; Qian, Z.; Kim, S. Antioxidant peptide isolated from muscle protein of bullfrog, Rana Catesbeiana Shaw. J. Med. Food 2007, 10, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Samaranayaka, A.G.P.; Lichan, E.C.Y. Food-derived peptidic antioxidants: A review of their production, assessment, and potential applications. J. Funct. Foods 2011, 3, 229–254. [Google Scholar] [CrossRef]
- Yang, F.; Jin, S.; Tang, Y. Marine Collagen Peptides Promote Cell Proliferation of NIH-3T3 Fibroblasts via NF-kB Signaling Pathway. Molecules 2019, 24, 4201. [Google Scholar] [CrossRef]
- Elias, R.J.; Kellerby, S.S.; Decker, E.A. Antioxidant Activity of Proteins and Peptides. Crit. Rev. Food Sci. Nutr. 2008, 48, 430–441. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Aluko, R.E. Chemometric Analysis of the Amino Acid Requirements of Antioxidant Food Protein Hydrolysates. Int. J. Mol. Sci. 2011, 12, 3148–3161. [Google Scholar] [CrossRef]
- Luisi, G.; Stefanucci, A.; Zengin, G.; Dimmito, M.P.; Mollica, A. Anti-Oxidant and Tyrosinase Inhibitory In Vitro Activity of Amino Acids and Small Peptides: New Hints for the Multifaceted Treatment of Neurologic and Metabolic Disfunctions. Antioxidants 2018, 8, 7. [Google Scholar] [CrossRef]
- Yang, X.; Zhao, Y.; Qiu, Y.; Chi, C.; Wang, B. Preparation and Characterization of Gelatin and Antioxidant Peptides from Gelatin Hydrolysate of Skipjack Tuna (Katsuwonus pelamis) Bone Stimulated by in vitro Gastrointestinal Digestion. Mar. Drugs 2019, 17, 78. [Google Scholar] [CrossRef]
- Xu, Z.; Fang, Y.; Chen, Y.; Yang, W.; Ma, N.; Pei, F.; Kimatu, B.M.; Hu, Q.; Qiu, W. Protective effects of Se-containing protein hydrolysates from Se-enriched rice against Pb2+-induced cytotoxicity in PC12 and RAW264.7 cells. Food Chem. 2016, 202, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Shi, Z.; Xie, C.; Gong, W.; Hu, Z.; Peng, Y. A novel mechanism of Gamma-aminobutyric acid (GABA) protecting human umbilical vein endothelial cells (HUVECs) against H2O2-induced oxidative injury. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2019, 217, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Ahn, C.; Je, J. Purification and characterization of antioxidant peptides from enzymatically hydrolyzed ark shell (Scapharca subcrenata). Process Biochem. 2018, 72, 170–176. [Google Scholar] [CrossRef]
- Chi, C.; Cao, Z.; Wang, B.; Hu, F.; Li, Z.; Zhang, B. Antioxidant and Functional Properties of Collagen Hydrolysates from Spanish Mackerel Skin as Influenced by Average Molecular Weight. Molecules 2014, 19, 11211–11230. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ding, L.; Yu, Z.; Zhang, T.; Ma, S.; Liu, J. Intracellular ROS scavenging and antioxidant enzyme regulating capacities of corn gluten meal-derived antioxidant peptides in HepG2 cells. Food Res. Int. 2016, 90, 33–41. [Google Scholar] [CrossRef]
- Cai, S.Y.; Wang, Y.M.; Zhao, Y.Q.; Chi, C.F.; Wang, B. Cytoprotective Effect of Antioxidant Pentapeptides from the Protein Hydrolysate of Swim Bladders of Miiuy Croaker (Miichthys miiuy) against H2O2-Mediated Human Umbilical Vein Endothelial Cell (HUVEC) Injury. Int. J. Mol. Sci. 2019, 20, 5425. [Google Scholar] [CrossRef]
- Pelicano, H.; Carney, D.A.; Huang, P. ROS stress in cancer cells and therapeutic implications. Drug Resist. Updat. 2004, 7, 97–110. [Google Scholar] [CrossRef]
- Zheng, L.; Yu, H.; Wei, H.; Xing, Q.; Zou, Y.; Zhou, Y.; Peng, J. Antioxidative peptides of hydrolysate prepared from fish skin gelatin using ginger protease activate antioxidant response element-mediated gene transcription in IPEC-J2 cells. J. Funct. Foods 2018, 51, 104–112. [Google Scholar] [CrossRef]
- Li, W.; Ye, S.; Zhang, Z.; Tang, J.; Jin, H.; Huang, F.; Yang, Z.; Tang, Y.; Chen, Y.; Ding, G.; et al. Purification and Characterization of a Novel Pentadecapeptide from Protein Hydrolysates of Cyclina sinensis and Its Immunomodulatory Effects on RAW264.7 Cells. Mar. Drugs 2019, 17, 30. [Google Scholar] [CrossRef]
- Wu, Z.Z.; Ding, G.F.; Huang, F.F.; Yang, Z.S.; Yu, F.M.; Tang, Y.P.; Jia, Y.L.; Zheng, Y.Y.; Chen, R. Anticancer Activity of Anthopleura anjunae Oligopeptides in Prostate Cancer DU-145 Cells. Mar. Drugs 2018, 16, 125. [Google Scholar] [CrossRef]
- Liu, Y.; Wan, S.; Liu, J.; Zou, Y.; Liao, S. Antioxidant Activity and Stability Study of Peptides from Enzymatically Hydrolyzed Male Silkmoth. J. Food Process. Pres. 2017, 41, 1–10. [Google Scholar] [CrossRef]
- Lin, X.H.; Chen, Y.Y.; Jin, H.X.; Zhao, Q.L.; Liu, C.J. Collagen Extracted from Bigeye Tuna (Thunnus obesus) Skin by Isoelectric Precipitation: Physicochemical Properties, Proliferation, and Migration Activities. Mar. Drugs 2019, 17, 261. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Runs | Temperature (X1) | pH (X2) | Enzyme Concentration (X1) | DPPH Clearance Rate (%) (Y) |
|---|---|---|---|---|
| 1 | 40 | 7 | 700 | 84.43 |
| 2 | 50 | 7 | 1300 | 93.24 |
| 3 | 45 | 8 | 1300 | 93.09 |
| 4 | 45 | 7 | 1000 | 94.33 |
| 5 | 45 | 6 | 1300 | 90.92 |
| 6 | 50 | 7 | 700 | 91.03 |
| 7 | 45 | 7 | 1000 | 93.78 |
| 8 | 40 | 8 | 1000 | 86.98 |
| 9 | 45 | 6 | 700 | 84.61 |
| 10 | 50 | 8 | 1000 | 92.62 |
| 11 | 40 | 7 | 1300 | 86.06 |
| 12 | 50 | 6 | 1000 | 90.24 |
| 13 | 45 | 8 | 700 | 91.51 |
| 14 | 45 | 7 | 1000 | 94.36 |
| 15 | 40 | 6 | 1000 | 82.71 |
| Source | Sum of Squares | df | Mean Square | F Value | p Value |
|---|---|---|---|---|---|
| Model | 218.02 | 9 | 24.22 | 39.82 | 0.0004 |
| X1 | 90.79 | 1 | 90.79 | 149.22 | <0.0001 |
| X2 | 30.89 | 1 | 30.89 | 50.77 | 0.0008 |
| X3 | 17.20 | 1 | 17.20 | 28.27 | 0.0031 |
| X1X2 | 0.89 | 1 | 0.89 | 1.47 | 0.2798 |
| X1X3 | 0.084 | 1 | 0.084 | 0.14 | 0.7253 |
| X2X3 | 5.59 | 1 | 5.59 | 9.19 | 0.0290 |
| X12 | 50.03 | 1 | 50.03 | 82.22 | 0.0003 |
| X22 | 20.19 | 1 | 20.19 | 33.18 | 0.0022 |
| X32 | 11.78 | 1 | 11.78 | 19.35 | 0.0070 |
| Residual | 3.04 | 5 | 0.61 | ||
| Lack of fit | 2.83 | 3 | 0.94 | 8.84 | 0.1033 |
| Pure Error | 0.21 | 2 | 0.11 | ||
| Cor Total | 221.07 | 14 | |||
| R2 | 0.9862 | ||||
| Radj2 | 0.9615 |
| Amino Acid | SNNH-1 |
|---|---|
| Aspartic acid | 38 |
| Threonine * | 14 |
| Serine | 24 |
| Glutamic acid | 76 |
| Glycine | 340 |
| Alanine | 134 |
| Cysteine | 0 |
| Valine * | 18 |
| Methionine * | 14 |
| Isoleucine * | 8 |
| Leucine * | 23 |
| Tyrosine | 7 |
| Phenylalanine * | 16 |
| Histidine | 6 |
| Lysine * | 28 |
| Arginine | 59 |
| Proline | 105 |
| Hydroxyproline | 86 |
| Imino acid | 191 |
| Independent Factors | Symbol | Level of Factor | ||
|---|---|---|---|---|
| −1 | 0 | 1 | ||
| Temperature (°C) | X1 | 40 | 45 | 50 |
| pH | X2 | 6 | 7 | 8 |
| Enzyme concentration (U/g) | X3 | 700 | 1000 | 1300 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, J.; Tian, X.; Xu, B.; Yuan, F.; Gong, J.; Yang, Z. Collagen Peptides from Swim Bladders of Giant Croaker (Nibea japonica) and Their Protective Effects against H2O2-Induced Oxidative Damage toward Human Umbilical Vein Endothelial Cells. Mar. Drugs 2020, 18, 430. https://doi.org/10.3390/md18080430
Zheng J, Tian X, Xu B, Yuan F, Gong J, Yang Z. Collagen Peptides from Swim Bladders of Giant Croaker (Nibea japonica) and Their Protective Effects against H2O2-Induced Oxidative Damage toward Human Umbilical Vein Endothelial Cells. Marine Drugs. 2020; 18(8):430. https://doi.org/10.3390/md18080430
Chicago/Turabian StyleZheng, Jiawen, Xiaoxiao Tian, Baogui Xu, Falei Yuan, Jianfang Gong, and Zuisu Yang. 2020. "Collagen Peptides from Swim Bladders of Giant Croaker (Nibea japonica) and Their Protective Effects against H2O2-Induced Oxidative Damage toward Human Umbilical Vein Endothelial Cells" Marine Drugs 18, no. 8: 430. https://doi.org/10.3390/md18080430
APA StyleZheng, J., Tian, X., Xu, B., Yuan, F., Gong, J., & Yang, Z. (2020). Collagen Peptides from Swim Bladders of Giant Croaker (Nibea japonica) and Their Protective Effects against H2O2-Induced Oxidative Damage toward Human Umbilical Vein Endothelial Cells. Marine Drugs, 18(8), 430. https://doi.org/10.3390/md18080430