Marine Terpenoids from Polar Latitudes and Their Potential Applications in Biotechnology




 and
and 
Abstract
1. Foreword
2. Marine Biotechnology: Governance, Access and Benefit Sharing in the Antarctic and Arctic Environments
3. Marine Compounds from Polar Regions
3.1. Marine chemical diversity in Polar Regions
3.2. Marine Polar Terpenoids
3.2.1. Monoterpenes and Monoterpenoids
3.2.2. Sesquiterpenes and Sesquiterpenoids
3.2.3. Diterpenes and Diterpenoids
3.2.4. Sesterterpenes and Sesterterpenoids
3.2.5. Triterpenes and Triterpenoids
Steroids
3.2.6. Tetraterpenes and Tetraterpenoids
Carotenoids
3.2.7. Triterpene and Triterpenoid Derivatives
Triterpenoid Conjugates
Triterpenoid Saponins
Meroterpenes
4. Perspectives and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hurst, D.; Børresen, T.; Almesjö, L.; De Raedemaecker, F.; Bergseth, S. Marine Biotechnology Strategic Research and Innovation Roadmap: Insights to the Future Direction of European Marine Biotechnology; Marine Biotechnology ERA-NET: Oostende, Belgium, 2016. [Google Scholar]
- Nunez-Pons, L.; Avila, C.; Romano, G.; Verde, C.; Giordano, D. UV-protective compounds in marine organisms from the Southern Ocean. Mar. Drugs 2018, 16, 336. [Google Scholar] [CrossRef] [PubMed]
- Angulo-Preckler, C.; Spurkland, T.; Avila, C.; Iken, K. Antimicrobial activity of selected benthic Arctic invertebrates. Polar Biol. 2015, 38, 1941–1948. [Google Scholar] [CrossRef]
- Bruno, S.; Coppola, D.; di Prisco, G.; Giordano, D.; Verde, C. Enzymes from marine polar regions and their biotechnological applications. Mar. Drugs 2019, 17, 544. [Google Scholar] [CrossRef] [PubMed]
- Soldatou, S.; Baker, B.J. Cold-water marine natural products, 2006 to 2016. Nat. Prod. Rep. 2017, 34, 585–626. [Google Scholar] [CrossRef]
- Tian, Y.; Li, Y.L.; Zhao, F.C. Secondary metabolites from polar organisms. Mar. Drugs 2017, 15, 28. [Google Scholar] [CrossRef]
- Eakins, B.W.; Sharman, G.F. Volumes of the World’s Oceans from ETOPO1; NOAA National Geophysical Data Center: Boulder, CO, USA, 2010.
- Polyak, L.; Alley, R.B.; Andrews, J.T.; Brigham-Grette, J.; Cronin, T.M.; Darby, D.A.; Dyke, A.S.; Fitzpatrick, J.J.; Funder, S.; Holland, M.; et al. History of sea ice in the Arctic. Quat. Sci. Rev. 2010, 29, 1757–1778. [Google Scholar] [CrossRef]
- Convey, P.; Bindschadler, R.; di Prisco, G.; Fahrbach, E.; Gutt, J.; Hodgson, D.A.; Mayewski, P.A.; Summerhayes, C.P.; Turner, J. Antarctic climate change and the environment. Antarct. Sci. 2009, 21, 541–563. [Google Scholar] [CrossRef]
- Jayatilake, G.S.; Baker, B.J.; McClintock, J.B. Isolation and identification of a stilbene derivative from the Antarctic sponge Kirkpatrickia variolosa. J. Nat. Prod. 1995, 58, 1958–1960. [Google Scholar] [CrossRef]
- Gordon, A. Oceanography of Antarctic Waters. In Antarctic Oceanology I; Reid, J.L., Ed.; AGU: Hoboken, NJ, USA, 1971; pp. 169–203. [Google Scholar]
- Barker, P.F.; Filippelli, G.M.; Florindo, F.; Martin, E.E.; Scher, H.D. Onset and role of the Antarctic Circumpolar Current. Deep Sea Res. II Top. Stud. Ocean. 2007, 54, 2388–2398. [Google Scholar] [CrossRef]
- Avila, C.; Taboada, S.; Núñez-Pons, L. Antarctic marine chemical ecology: What is next? Mar. Ecol. 2008, 29, 1–71. [Google Scholar] [CrossRef]
- Núñez-Pons, L.; Avila, C. Natural products mediating ecological interactions in Antarctic benthic communities: A mini-review of the known molecules. Nat. Prod. Rep. 2015, 32, 1114–1130. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.T.; Lu, X.L.; Liu, X.Y.; Gao, Y.; Hu, B.; Jiao, B.H.; Zheng, H. Bioactive natural products from the antarctic and arctic organisms. Mini Rev. Med. Chem. 2013, 13, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Lebar, M.D.; Heimbegner, J.L.; Baker, B.J. Cold-water marine natural products. Nat. Prod. Rep. 2007, 24, 774–797. [Google Scholar] [CrossRef] [PubMed]
- Klemm, E.J.; Wong, V.K.; Dougan, G. Emergence of dominant multidrug-resistant bacterial clades: Lessons from history and whole-genome sequencing. Proc. Nat. Acad. Sci. USA 2018, 115, 12872–12877. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.; Vieira, H.; Gaspar, H.; Santos, S. Marketed marine natural products in the pharmaceutical and cosmeceutical industries: Tips for success. Mar. Drugs 2014, 12, 1066–1101. [Google Scholar] [CrossRef] [PubMed]
- Market Research Engine. Available online: https://www.marketresearchengine.com/marine-derived-drugs-market (accessed on 15 May 2020).
- Mayer, A.S.M. Marine Pharmaceuticals: The Clinical Pipeline. Available online: https://www.midwestern.edu/departments/marinepharmacology.xml (accessed on 27 July 2020).
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef] [PubMed]
- Jaspars, M.; De Pascale, D.; Andersen, J.H.; Reyes, F.; Crawford, A.D.; Ianora, A. The marine biodiscovery pipeline and ocean medicines of tomorrow. J. Mar. Biol. Assoc. U. K. 2016, 96, 151–158. [Google Scholar] [CrossRef]
- Schoffski, P.; Dumez, H.; Wolter, P.; Stefan, C.; Wozniak, A.; Jimeno, J.; Van Oosterom, A.T. Clinical impact of trabectedin (ecteinascidin-743) in advanced/metastatic soft tissue sarcoma. Expert Opin. Pharm. 2008, 9, 1609–1618. [Google Scholar] [CrossRef]
- Karsch-Mizrachi, I.; Takagi, T.; Cochrane, G.; International Nucleotide Sequence Database Collaboration. The international nucleotide sequence database collaboration. Nucleic Acids Res. 2018, 46, D48–D51. [Google Scholar] [CrossRef]
- Costello, M.J.; Chaudhary, C. Marine biodiversity, biogeography, deep-sea gradients, and conservation. Curr. Biol. 2017, 27, R511–R527. [Google Scholar] [CrossRef]
- Puig-Marcó, R. Access and benefit sharing of Antarctica’s biological material. Mar. Genom. 2014, 17, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Dodds, K.J. Sovereignty watch: Claimant states, resources, and territory in contemporary Antarctica. Polar Rec. 2010, 47, 231–243. [Google Scholar] [CrossRef]
- Petra, D.; Alex, G.O.E.; Bert, V.; Tamara, T. Marine genetic resources in areas beyond national jurisdiction: Access and benefit-sharing. Int. J. Mar. Coast. Law 2012, 27, 375–433. [Google Scholar] [CrossRef]
- Dodds, K. Governing Antarctica: Contemporary challenges and the enduring legacy of the 1959 Antarctic Treaty. Glob. Policy 2010, 1, 108–115. [Google Scholar] [CrossRef]
- Joyner, C.C. Bioprospecting as a challenge to the Antarctic Treaty. In Antarctic Security in the Twenty-First Century; Hemmings, A.D., Rothwell, D.R., Scott, K.N., Eds.; Routledge: Abingdon, UK, 2012; pp. 197–214. [Google Scholar]
- Tvedt, M.W. Patent law and bioprospecting in Antarctica. Polar Rec. 2010, 47, 46–55. [Google Scholar] [CrossRef]
- Rogan-Finnemore, M. What bioprospecting means for Antarctica and the Southern Ocean. In International Law Issues in the South Pacific; von Tigerstrom, B., Ed.; Ashgate: Hampshire, UK, 2005; pp. 199–228. [Google Scholar]
- Summerhayes, C.P. International collaboration in Antarctica: The International Polar Years, the International Geophysical Year, and the Scientific Committee on Antarctic Research. Polar Rec. 2008, 44, 321–334. [Google Scholar] [CrossRef]
- Oldham, P.; Hall, S.; Forero, O. Biological diversity in the patent system. PLoS ONE 2013, 8, e78737. [Google Scholar] [CrossRef]
- Eritja, M.C. Bio-prospecting in the Arctic: An overview of the interaction between the rights of indigenous peoples and access and benefit sharing. Boston Coll. Environ. Aff. Law Rev. 2017, 44, 223. [Google Scholar]
- Hoag, H. The cold rush. Nat. Biotech. 2009, 27, 690–692. [Google Scholar] [CrossRef]
- Leary, D. UNU-IAS Report: Bioprospecting in the Arctic 8, 2008; UNU-IAS: Minato, Japan, 2013. [Google Scholar]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2020, 37, 175–223. [Google Scholar] [CrossRef]
- Alcaino, J.; Cifuentes, V.; Baeza, M. Physiological adaptations of yeasts living in cold environments and their potential applications. World J. Microbiol. Biotechnol. 2015, 31, 1467–1473. [Google Scholar] [CrossRef] [PubMed]
- Munoz, P.A.; Marquez, S.L.; Gonzalez-Nilo, F.D.; Marquez-Miranda, V.; Blamey, J.M. Structure and application of antifreeze proteins from Antarctic bacteria. Microb. Cell Fact. 2017, 16, 138. [Google Scholar] [CrossRef] [PubMed]
- Bang, J.K.; Lee, J.H.; Murugan, R.N.; Lee, S.G.; Do, H.; Koh, H.Y.; Shim, H.E.; Kim, H.C.; Kim, H.J. Antifreeze peptides and glycopeptides, and their derivatives: Potential uses in biotechnology. Mar. Drugs 2013, 11, 2013–2041. [Google Scholar] [CrossRef] [PubMed]
- Venketesh, S.; Dayananda, C. Properties, potentials, and prospects of antifreeze proteins. Crit. Rev. Biotechnol. 2008, 28, 57–82. [Google Scholar] [CrossRef] [PubMed]
- Qadeer, S.; Khan, M.A.; Ansari, M.S.; Rakha, B.A.; Ejaz, R.; Iqbal, R.; Younis, M.; Ullah, N.; DeVries, A.L.; Akhter, S. Efficiency of antifreeze glycoproteins for cryopreservation of Nili-Ravi (Bubalus bubalis) buffalo bull sperm. Anim. Reprod. Sci. 2015, 157, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.G.; Koh, H.Y.; Lee, J.H.; Kang, S.H.; Kim, H.J. Cryopreservative effects of the recombinant ice-binding protein from the arctic yeast Leucosporidium sp. on red blood cells. Appl. Biochem. Biotechnol. 2012, 167, 824–834. [Google Scholar] [CrossRef]
- Kumar, A.S.; Mody, K.; Jha, B. Bacterial exopolysaccharides-A perception. J. Basic Microbiol. 2007, 47, 103–117. [Google Scholar] [CrossRef]
- Freitas, F.; Alves, V.D.; Reis, M.A. Advances in bacterial exopolysaccharides: From production to biotechnological applications. Trends Biotechnol. 2011, 29, 388–398. [Google Scholar] [CrossRef]
- Caruso, C.; Rizzo, C.; Mangano, S.; Poli, A.; Di Donato, P.; Nicolaus, B.; Di Marco, G.; Michaud, L.; Lo Giudice, A. Extracellular polymeric substances with metal adsorption capacity produced by Pseudoalteromonas sp. MER144 from Antarctic seawater. Environ. Sci. Pollut. Res. Int. 2018, 25, 4667–4677. [Google Scholar] [CrossRef]
- Poli, A.; Anzelmo, G.; Nicolaus, B. Bacterial exopolysaccharides from extreme marine habitats: Production, characterization and biological activities. Mar. Drugs 2010, 8, 1779–1802. [Google Scholar] [CrossRef]
- Bianchi, A.C.; Olazabal, L.; Torre, A.; Loperena, L. Antarctic microorganisms as source of the omega-3 polyunsaturated fatty acids. World J. Microbiol. Biotechnol. 2014, 30, 1869–1878. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Sharpe, P.L.; Hong, S.P.; Yadav, N.S.; Xie, D.; Short, D.R.; Damude, H.G.; Rupert, R.A.; Seip, J.E.; Wang, J.; et al. Production of omega-3 eicosapentaenoic acid by metabolic engineering of Yarrowia lipolytica. Nat. Biotechnol. 2013, 31, 734–740. [Google Scholar] [CrossRef]
- Abbas, S.; Kelly, M.; Bowling, J.; Sims, J.; Waters, A.; Hamann, M. Advancement into the Arctic region for bioactive sponge secondary metabolites. Mar. Drugs 2011, 9, 2423–2437. [Google Scholar] [CrossRef] [PubMed]
- Hansen, Ø.K.; Isaksson, J.; Glomsaker, E.; Andersen, H.J.; Hansen, E. Ponasterone A and F, ecdysteroids from the Arctic bryozoan Alcyonidium gelatinosum. Molecules 2018, 23, 1481. [Google Scholar] [CrossRef]
- Di, X.; Rouger, C.; Hardardottir, I.; Freysdottir, J.; Molinski, T.F.; Tasdemir, D.; Omarsdottir, S. 6-Bromoindole derivatives from the Icelandic marine sponge Geodia barretti: Isolation and anti-inflammatory activity. Mar. Drugs 2018, 16, 437. [Google Scholar] [CrossRef]
- Figuerola, B.; Sala-Comorera, L.; Angulo-Preckler, C.; Vazquez, J.; Jesus Montes, M.; Garcia-Aljaro, C.; Mercade, E.; Blanch, A.R.; Avila, C. Antimicrobial activity of Antarctic bryozoans: An ecological perspective with potential for clinical applications. Mar. Environ. Res. 2014, 101, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Berne, S.; Kalauz, M.; Lapat, M.; Savin, L.; Janussen, D.; Kersken, D.; Avguštin, J.A.; Jokhadar, Š.Z.; Jaklič, D.; Gunde-Cimerman, N.; et al. Screening of the Antarctic marine sponges (Porifera) as a source of bioactive compounds. Polar Biol. 2016, 39, 947–959. [Google Scholar] [CrossRef]
- Moles, J.; Nunez-Pons, L.; Taboada, S.; Figueroa, B.; Cristobo, J.; Avila, C. Anti-predatory chemical defences in Antarctic benthic fauna. Mar. Biol. 2015, 162, 1813–1821. [Google Scholar] [CrossRef]
- Li, F.; Janussen, D.; Peifer, C.; Perez-Victoria, I.; Tasdemir, D. Targeted isolation of tsitsikammamines from the Antarctic deep-sea sponge Latrunculia biformis by molecular networking and anticancer activity. Mar. Drugs 2018, 16, 268. [Google Scholar] [CrossRef]
- Thomas, T.R.; Kavlekar, D.P.; LokaBharathi, P.A. Marine drugs from sponge-microbe association—A review. Mar. Drugs 2010, 8, 1417–1468. [Google Scholar] [CrossRef]
- Blockley, A.; Elliott, D.; Roberts, A.P.; Sweet, M. Symbiotic microbes from marine invertebrates: Driving a new era of natural product drug discovery. Div. Distrib. 2017, 9, 49. [Google Scholar] [CrossRef]
- Lo Giudice, A.; Rizzo, C. Bacteria associated with marine benthic invertebrates from polar environments: Unexplored frontiers for biodiscovery? Diversity 2018, 10, 80. [Google Scholar] [CrossRef]
- Furbino, L.E.; Godinho, V.M.; Santiago, I.F.; Pellizari, F.M.; Alves, T.M.; Zani, C.L.; Junior, P.A.; Romanha, A.J.; Carvalho, A.G.; Gil, L.H.; et al. Diversity patterns, ecology and biological activities of fungal communities associated with the endemic macroalgae across the Antarctic peninsula. Microb. Ecol. 2014, 67, 775–787. [Google Scholar] [CrossRef]
- Henríquez, M.; Vergara, K.; Norambuena, J.; Beiza, A.; Maza, F.; Ubilla, P.; Araya, I.; Chávez, R.; San-Martín, A.; Darias, J.; et al. Diversity of cultivable fungi associated with Antarctic marine sponges and screening for their antimicrobial, antitumoral and antioxidant potential. World J. Microbiol. Biotech. 2014, 30, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Godinho, V.M.; Furbino, L.E.; Santiago, I.F.; Pellizzari, F.M.; Yokoya, N.S.; Pupo, D.; Alves, T.M.; Junior, P.A.; Romanha, A.J.; Zani, C.L.; et al. Diversity and bioprospecting of fungal communities associated with endemic and cold-adapted macroalgae in Antarctica. ISME J. 2013, 7, 1434–1451. [Google Scholar] [CrossRef]
- Maida, I.; Bosi, E.; Fondi, M.; Perrin, E.; Orlandini, V.; Papaleo, M.; Mengoni, A.; de Pascale, D.; Tutino, M.L.; Michaud, L.; et al. Antimicrobial activity of Pseudoalteromonas strains isolated from the Ross Sea (Antarctica) versus Cystic Fibrosis opportunistic pathogens. Hydrobiologia 2015, 761, 443–457. [Google Scholar] [CrossRef]
- Jayatilake, G.S.; Thornton, M.P.; Leonard, A.C.; Grimwade, J.E.; Baker, B.J. Metabolites from an Antarctic sponge-associated bacterium, Pseudomonas aeruginosa. J. Nat. Prod. 1996, 59, 293–296. [Google Scholar] [CrossRef]
- Mangano, S.; Michaud, L.; Caruso, C.; Brilli, M.; Bruni, V.; Fani, R.; Lo Giudice, A. Antagonistic interactions between psychrotrophic cultivable bacteria isolated from Antarctic sponges: A preliminary analysis. Res. Microbiol. 2009, 160, 27–37. [Google Scholar] [CrossRef]
- Orlandini, V.; Maida, I.; Fondi, M.; Perrin, E.; Papaleo, M.C.; Bosi, E.; de Pascale, D.; Tutino, M.L.; Michaud, L.; Lo Giudice, A.; et al. Genomic analysis of three sponge-associated Arthrobacter Antarctic strains, inhibiting the growth of Burkholderia cepacia complex bacteria by synthesizing volatile organic compounds. Microbiol. Res. 2014, 169, 593–601. [Google Scholar] [CrossRef]
- Papaleo, M.C.; Fondi, M.; Maida, I.; Perrin, E.; Lo Giudice, A.; Michaud, L.; Mangano, S.; Bartolucci, G.; Romoli, R.; Fani, R. Sponge-associated microbial Antarctic communities exhibiting antimicrobial activity against Burkholderia cepacia complex bacteria. Biotechnol. Adv. 2012, 30, 272–293. [Google Scholar] [CrossRef]
- Papaleo, M.C.; Romoli, R.; Bartolucci, G.; Maida, I.; Perrin, E.; Fondi, M.; Orlandini, V.; Mengoni, A.; Emiliani, G.; Tutino, M.L.; et al. Bioactive volatile organic compounds from Antarctic (sponges) bacteria. New Biotechnol. 2013, 30, 824–838. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, P.; Maida, I.; Palma Esposito, F.; Tortorella, E.; Subko, K.; Ezeofor, C.C.; Zhang, Y.; Tabudravu, J.; Jaspars, M.; Fani, R.; et al. Antimicrobial activity of monoramnholipids produced by bacterial strains isolated from the Ross Sea (Antarctica). Mar. Drugs 2016, 14, 83. [Google Scholar] [CrossRef] [PubMed]
- Wietz, M.; Månsson, M.; Bowman, J.S.; Blom, N.; Ng, Y.; Gram, L. Wide distribution of closely related, antibiotic-producing Arthrobacter strains throughout the Arctic Ocean. Appl. Environ. Microbiol. 2012, 78, 2039–2042. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Lu, Y.; Xing, Y.; Ma, Y.; Lu, J.; Bao, W.; Wang, Y.; Xi, T. A novel anticancer and antifungus phenazine derivative from a marine actinomycete BM-17. Microbiol. Res. 2012, 167, 616–622. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Saurav, K.; Yu, Z.; Mándi, A.; Kurtán, T.; Li, J.; Tian, X.; Zhang, Q.; Zhang, W.; Zhang, C. α-Pyrones with diverse hydroxy substitutions from three marine-derived Nocardiopsis strains. J. Nat. Prod. 2016, 79, 1610–1618. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xie, B.; Yang, J.; Chen, J.; Sun, Z. Identification of microbial carotenoids and isoprenoid quinones from Rhodococcus sp. B7740 and its stability in the presence of iron in model gastric conditions. Food Chem. 2018, 240, 204–211. [Google Scholar] [CrossRef]
- Chen, Y.; Guo, M.; Yang, J.; Chen, J.; Xie, B.; Sun, Z. Potential TSPO ligand and photooxidation quencher isorenieratene from Arctic Ocean Rhodococcus sp. B7740. Mar. Drugs 2019, 17, 316. [Google Scholar] [CrossRef]
- Chen, Y.; Mu, Q.; Hu, K.; Chen, M.; Yang, J.; Chen, J.; Xie, B.; Sun, Z. Characterization of MK(8)(H(2)) from Rhodococcus sp. B7740 and its potential antiglycation capacity measurements. Mar. Drugs 2018, 16, 391. [Google Scholar] [CrossRef]
- Moon, K.; Ahn, C.-H.; Shin, Y.; Won, T.H.; Ko, K.; Lee, S.K.; Oh, K.-B.; Shin, J.; Nam, S.-I.; Oh, D.-C. New benzoxazine secondary metabolites from an Arctic Actinomycete. Mar. Drugs 2014, 12, 2526–2538. [Google Scholar] [CrossRef]
- Ivanova, V.; Oriol, M.; Montes, M.-J.; Garcia, A.; Guinea, J. Secondary metabolites from a Streptomyces strain isolated from Livingston Island, Antarctica. Z. Naturforsch. C Biosci. 2001, 56, 1–5. [Google Scholar] [CrossRef]
- Kim, D.; Lee, E.J.; Lee, J.; Leutou, A.S.; Shin, Y.H.; Choi, B.; Hwang, J.S.; Hahn, D.; Choi, H.; Chin, J.; et al. Antartin, a cytotoxic zizaane-type sesquiterpenoid from a Streptomyces sp. isolated from an Antarctic marine sediment. Mar. Drugs 2018, 16, 130. [Google Scholar] [CrossRef] [PubMed]
- Macherla, V.R.; Liu, J.; Bellows, C.; Teisan, S.; Nicholson, B.; Lam, K.S.; Potts, B.C.M. Glaciapyrroles A, B, and C, pyrrolosesquiterpenes from a Streptomyces sp. isolated from an Alaskan marine sediment. J. Nat. Prod. 2005, 68, 780–783. [Google Scholar] [CrossRef] [PubMed]
- Sannino, F.; Parrilli, E.; Apuzzo, G.A.; de Pascale, D.; Tedesco, P.; Maida, I.; Perrin, E.; Fondi, M.; Fani, R.; Marino, G.; et al. Pseudoalteromonas haloplanktis produces methylamine, a volatile compound active against Burkholderia cepacia complex strains. New Biotechnol. 2017, 35, 13–18. [Google Scholar] [CrossRef]
- Sannino, F.; Sansone, C.; Galasso, C.; Kildgaard, S.; Tedesco, P.; Fani, R.; Marino, G.; de Pascale, D.; Ianora, A.; Parrilli, E.; et al. Pseudoalteromonas haloplanktis TAC125 produces 4-hydroxybenzoic acid that induces pyroptosis in human A459 lung adenocarcinoma cells. Sci. Rep. 2018, 8, 1190. [Google Scholar] [CrossRef]
- Mitova, M.; Tutino, M.L.; Infusini, G.; Marino, G.; De Rosa, S. Exocellular peptides from Antarctic psychrophile Pseudoalteromonas Haloplanktis. Mar. Biotechnol. 2005, 7, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Casillo, A.; Papa, R.; Ricciardelli, A.; Sannino, F.; Ziaco, M.; Tilotta, M.; Selan, L.; Marino, G.; Corsaro, M.M.; Tutino, M.L.; et al. Anti-biofilm activity of a long-chain fatty aldehyde from Antarctic Pseudoalteromonas haloplanktis TAC125 against Staphylococcus epidermidis biofilm. Front. Cell. Infect. Microb. 2017, 7, 46. [Google Scholar] [CrossRef] [PubMed]
- Chianese, G.; Esposito, F.P.; Parrot, D.; Ingham, C.; de Pascale, D.; Tasdemir, D. Linear aminolipids with moderate antimicrobial activity from the Antarctic gram-negative bacterium Aequorivita sp. Mar. Drugs 2018, 16, 187. [Google Scholar] [CrossRef]
- Al-Zereini, W.; Schuhmann, I.; Laatsch, H.; Helmke, E.; Anke, H. New aromatic nitro compounds from Salegentibacter sp. T436, an Arctic sea ice bacterium: Taxonomy, fermentation, isolation and biological activities. J. Antibiot. 2007, 60, 301–308. [Google Scholar] [CrossRef]
- Schuhmann, I.; Yao, C.B.F.-F.; Al-Zereini, W.; Anke, H.; Helmke, E.; Laatsch, H. Nitro derivatives from the Arctic ice bacterium Salegentibacter sp. isolate T436. J. Antibiot. 2009, 62, 453–460. [Google Scholar] [CrossRef]
- Zhang, H.L.; Hua, H.M.; Pei, Y.H.; Yao, X.S. Three new cytotoxic cyclic acylpeptides from marine Bacillus sp. Chem. Pharm. Bull. 2004, 52, 1029–1030. [Google Scholar] [CrossRef]
- Corral, P.; Esposito, F.P.; Tedesco, P.; Falco, A.; Tortorella, E.; Tartaglione, L.; Festa, C.; D’Auria, M.V.; Gnavi, G.; Varese, G.C.; et al. Identification of a sorbicillinoid-producing Aspergillus strain with antimicrobial activity against Staphylococcus aureus: A new polyextremophilic marine fungus from Barents Sea. Mar. Biotechnol. 2018, 20, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.W.; Lin, A.Q.; Gu, Q.Q.; Zhu, T.J.; Li, D.H. Four new chloro-eremophilane sesquiterpenes from an Antarctic deep-sea derived fungus, Penicillium sp PR19N-1. Mar. Drugs 2013, 11, 1399–1408. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Wu, G.; Gu, Q.; Zhu, T.; Li, D. New eremophilane-type sesquiterpenes from an Antarctic deepsea derived fungus, Penicillium sp. PR19 N-1. Arch. Pharm. Res. 2014, 37, 839–844. [Google Scholar] [CrossRef]
- Niu, S.; Fan, Z.-W.; Xie, C.-L.; Liu, Q.; Luo, Z.-H.; Liu, G.; Yang, X.-W. Spirograterpene A, a tetracyclic spiro-diterpene with a fused 5/5/5/5 ring system from the deep-sea-derived fungus Penicillium granulatum MCCC 3A00475. J. Nat. Prod. 2017, 80, 2174–2177. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, Y.H.; Yu, H.B.; Liu, X.Y.; Lu, X.L.; Jiao, B.H. Furanone derivative and sesquiterpene from Antarctic marine-derived fungus Penicillium sp. S-1–18. J. Asian Nat. Prod. Res. 2018, 20, 1108–1115. [Google Scholar] [CrossRef]
- Hu, Z.-Y.; Li, Y.-Y.; Huang, Y.-J.; Su, W.-J.; Shen, Y.-M. Three new sesquiterpenoids from Xylaria sp. NCY2. Helv. Chim. Acta 2008, 91, 46–52. [Google Scholar] [CrossRef]
- Wu, G.W.; Ma, H.Y.; Zhu, T.J.; Li, J.; Gu, Q.Q.; Li, D.H. Penilactones A and B, two novel polyketides from Antarctic deep-sea derived fungus Penicillium crustosum PRB-2. Tetrahedron 2012, 68, 9745–9749. [Google Scholar] [CrossRef]
- Wu, B.; Wiese, J.; Labes, A.; Kramer, A.; Schmaljohann, R.; Imhoff, J.F. Lindgomycin, an unusual antibiotic polyketide from a marine fungus of the Lindgomycetaceae. Mar. Drugs 2015, 13, 4617–4632. [Google Scholar] [CrossRef]
- Shin, H.J.; Mondol, M.A.M.; Yu, T.K.; Lee, H.-S.; Lee, Y.-J.; Jung, H.J.; Kim, J.H.; Kwon, H.J. An angiogenesis inhibitor isolated from a marine-derived actinomycete, Nocardiopsis sp. 03N67. Phytochem. Lett. 2010, 3, 194–197. [Google Scholar] [CrossRef]
- Silva, T.R.; Canela-Garayoa, R.; Eras, J.; Rodrigues, M.V.N.; Dos Santos, F.N.; Eberlin, M.N.; Neri-Numa, I.A.; Pastore, G.M.; Tavares, R.S.N.; Debonsi, H.M.; et al. Pigments in an iridescent bacterium, Cellulophaga fucicola, isolated from Antarctica. Antonie Leeuwenhoek 2019, 112, 479–490. [Google Scholar] [CrossRef]
- Silva, T.R.; Tavares, R.S.N.; Canela-Garayoa, R.; Eras, J.; Rodrigues, M.V.N.; Neri-Numa, I.A.; Pastore, G.M.; Rosa, L.H.; Schultz, J.A.A.; Debonsi, H.M.; et al. Chemical characterization and biotechnological applicability of pigments i solated from Antarctic bacteria. Mar. Biotechnol. 2019, 21, 416–429. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, L.; Jiménez, C.; Rodríguez, J.; Areche, C.; Chávez, R.; Henríquez, M.; de la Cruz, M.; Díaz, C.; Segade, Y.; Vaca, I. 3-Nitroasterric acid derivatives from an Antarctic sponge-derived Pseudogymnoascus sp. fungus. J. Nat. Prod. 2015, 78, 919–923. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-C.; Lee, H.-S.; Ko, W.; Lee, D.-S.; Sohn, J.H.; Yim, J.H.; Kim, Y.-C.; Oh, H. Anti-inflammatory effect of methylpenicinoline from a marine isolate of Penicillium sp. (SF-5995): Inhibition of NF-κB and MAPK pathways in lipopolysaccharide-induced RAW264.7 macrophages and BV2 microglia. Molecules 2014, 19, 18073–18089. [Google Scholar] [CrossRef] [PubMed]
- Manzo, E.; Ciavatta, M.L.; Melck, D.; Schupp, P.; de Voogd, N.; Gavagnin, M. Aromatic cyclic peroxides and related keto-compounds from the Plakortis sp. component of a sponge consortium. J. Nat. Prod. 2009, 72, 1547–1551. [Google Scholar] [CrossRef] [PubMed]
- Carbone, M.; Núñez-Pons, L.; Castelluccio, F.; Avila, C.; Gavagnin, M. Illudalane sesquiterpenoids of the alcyopterosin series from the Antarctic marine soft coral Alcyonium grandis. J. Nat. Prod. 2009, 72, 1357–1360. [Google Scholar] [CrossRef]
- Palermo, J.A.; Brasco, M.F.; Spagnuolo, C.; Seldes, A.M. Illudalane sesquiterpenoids from the soft coral Alcyonium paessleri: The first natural nitrate esters. J. Org. Chem. 2000, 65, 4482–4486. [Google Scholar] [CrossRef]
- Brasco, M.F.R.; Seldes, A.M.; Palermo, J.A. Paesslerins A and B: Novel tricyclic sesquiterpenoids from the soft coral Alcyonium paessleri. Org. Lett. 2001, 3, 1415–1417. [Google Scholar] [CrossRef]
- Mellado, G.G.; Zubia, E.; Ortega, M.J.; Lopez-Gonzalez, P.J. Steroids from the Antarctic octocoral Anthomastus bathyproctus. J. Nat. Prod. 2005, 68, 1111–1115. [Google Scholar] [CrossRef]
- Thomas, S.A.L.; Sanchez, A.; Kee, Y.; Wilson, N.G.; Baker, B.J. Bathyptilones: Terpenoids from an Antarctic sea pen, Anthoptilum grandiflorum (Verrill, 1879). Mar. Drugs 2019, 17, 513. [Google Scholar] [CrossRef]
- Mellado, G.G.; Zubia, E.; Ortega, M.J.; Lopez-Gonzalez, P.J. New polyoxygenated steroids from the Antarctic octocoral Dasystenella Acanthina. Steroids 2004, 69, 291–299. [Google Scholar] [CrossRef]
- Koljak, R.; Lopp, A.; Pehk, T.; Varvas, K.; Muurisepp, A.-M.; Jarving, I.; Samel, N. New cytotoxic sterols from the soft coral Gersemia fruticosa. Tetrahedron 1998, 54, 179–186. [Google Scholar] [CrossRef]
- Lopp, A.; Pihlak, A.; Paves, H.; Samuel, K.; Koljak, R.; Samel, N. The effect of 9,11-secosterol, a newly discovered compound from the soft coral Gersemia fruticosa, on the growth and cell cycle progression of various tumor cells in culture. Steroids 1994, 59, 274–281. [Google Scholar] [CrossRef]
- Angulo-Preckler, C.; Genta-Jouve, G.; Mahajan, N.; de la Cruz, M.; de Pedro, N.; Reyes, F.; Iken, K.; Avila, C.; Thomas, O.P. Gersemiols A–C and eunicellol A, diterpenoids from the Arctic soft coral Gersemia fruticosa. J. Nat. Prod. 2016, 79, 1132–1136. [Google Scholar] [CrossRef]
- Thomas, S.A.L.; von Salm, J.L.; Clark, S.; Nemani, P.; Ferlita, S.; Wilson, N.G.; Baker, B.J. Keikipukalides, furanocembranoid aldehydes from the deep sea Antarctic coral Plumerella delicatissima. J. Nat. Prod. 2018, 81, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Von Salm, J.L.; Wilson, N.G.; Vesely, B.A.; Kyle, D.E.; Cuce, J.; Baker, B.J. Shagenes A and B, new tricyclic sesquiterpenes produced by an undescribed Antarctic octocoral. Org. Lett. 2014, 16, 2630–2633. [Google Scholar] [CrossRef] [PubMed]
- Tadesse, M.; Tabudravu, J.N.; Jaspars, M.; Strom, M.B.; Hansen, E.; Andersen, J.H.; Kristiansen, P.E.; Haug, T. The antibacterial ent-eusynstyelamide B and eusynstyelamides D, E, and F from the Arctic bryozoan Tegella cf. spitzbergensis. J. Nat. Prod. 2011, 74, 837–841. [Google Scholar] [CrossRef]
- Michael, P.; Hansen, E.; Isaksson, J.; Andersen, J.H.; Hansen, K.Ø. Dendrobeaniamine A, a new alkaloid from the Arctic marine bryozoan Dendrobeania murrayana. Nat. Prod. Res. 2019, 34, 2059–2064. [Google Scholar] [CrossRef]
- Maschek, J.A.; Mevers, E.M.; Diyabalanage, T.; Chen, L.; Ren, Y.; Amsler, C.D.; McClintock, J.B.; Wu, J.; Baker, B.J. Palmadorin chemodiversity from the Antarctic nudibranch Austrodoris kerguelenensis and inhibition of Jak2/STAT5-dependent HEL leukemia cells. Tetrahedron 2012, 68, 9095–9104. [Google Scholar] [CrossRef]
- Rodriguez, J.; Castro, R.; Riguera, R. Holothurinosides: New antitumour non-sulphated triterpenoid glycosides from the sea cucumber Holothuria forskalii. Tetrahedron 1991, 47, 4753–4762. [Google Scholar] [CrossRef]
- Maier, M.S.; Roccatagliata, A.J.; Kuriss, A.; Chludil, H.; Seldes, A.M.; Pujol, C.A.; Damonte, E.B. Two new cytotoxic and virucidal trisulfated triterpene glycosides from the Antarctic sea cucumber Staurocucumis liouvillei. J. Nat. Prod. 2001, 64, 732–736. [Google Scholar] [CrossRef]
- Kicha, A.A.; Ivanchina, N.V.; Malyarenko, T.V.; Kalinovsky, A.I.; Popov, R.S.; Stonik, V.A. Six new polyhydroxylated steroids conjugated with taurine, microdiscusols A-F, from the Arctic starfish Asterias microdiscus. Steroids 2019, 150, 108458. [Google Scholar] [CrossRef]
- D’Auria, M.V.; Paloma, L.G.; Minale, L.; Riccio, R.; Zampella, A.; Morbidoni, M. Isolation and structure characterization of two novel bioactive sulphated polyhydroxysteroids from the Antarctic ophiuroid Ophioderma longicaudum. Nat. Prod. Lett. 1993, 3, 197–201. [Google Scholar] [CrossRef]
- Comin, M.J.; Maier, M.S.; Roccatagliata, A.J.; Pujol, C.A.; Damonte, E.B. Evaluation of the antiviral activity of natural sulfated polyhydroxysteroids and their synthetic derivatives and analogs. Steroids 1999, 64, 335–340. [Google Scholar] [CrossRef]
- Ma, W.S.; Mutka, T.; Vesley, B.; Amsler, M.O.; McClintock, J.B.; Amsler, C.D.; Perman, J.A.; Singh, M.P.; Maiese, W.M.; Zaworotko, M.J.; et al. Norselic acids A-E, highly oxidized anti-infective steroids that deter mesograzer predation, from the Antarctic sponge Crella sp. J. Nat. Prod. 2009, 72, 1842–1846. [Google Scholar] [CrossRef] [PubMed]
- Ankisetty, S.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Further membranolide diterpenes from the antarctic sponge Dendrilla membranosa. J. Nat. Prod. 2004, 67, 1172–1174. [Google Scholar] [CrossRef] [PubMed]
- Shilling, A.J.; Witowski, C.G.; Maschek, J.A.; Azhari, A.; Vesely, B.; Kyle, D.E.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Spongian diterpenoids derived from the Antarctic sponge Dendrilla antarctica are potent inhibitors of the Leishmania parasite. J. Nat. Prod. 2020, 83, 1553–1562. [Google Scholar] [CrossRef]
- Von Salm, J.L.; Witowski, C.G.; Fleeman, R.M.; McClintock, J.B.; Amsler, C.D.; Shaw, L.N.; Baker, B.J. Darwinolide, a new diterpene scaffold thati inhibits methicillin-resistant Staphylococcus aureus biofilm from the Antarctic sponge Dendrilla membranosa. Org. Lett. 2016, 18, 2596–2599. [Google Scholar] [CrossRef]
- Bory, A.; Shilling, A.J.; Allen, J.; Azhari, A.; Roth, A.; Shaw, L.N.; Kyle, D.E.; Adams, J.H.; Amsler, C.D.; McClintock, J.B.; et al. Bioactivity of spongian diterpenoid scaffolds from the Antarctic sponge Dendrilla Antarctica. Mar. Drugs. 2020, 18, 327. [Google Scholar] [CrossRef]
- Ciaglia, E.; Malfitano, A.M.; Laezza, C.; Fontana, A.; Nuzzo, G.; Cutignano, A.; Abate, M.; Pelin, M.; Sosa, S.; Bifulco, M.; et al. Immuno-modulatory and anti-inflammatory effects of dihydrogracilin A, a terpene derived from the marine sponge Dendrilla membranosa. Int. J. Mol. Sci. 2017, 18, 1643. [Google Scholar] [CrossRef]
- Lidgren, G.; Bohlin, L.; Bergman, J. Studies of Swedish marine organisms. VII. A novel biologically active indole alkaloid from the sponge Geodia baretti. Tetrahedron Lett. 1986, 27, 3283–3284. [Google Scholar] [CrossRef]
- Lind, K.F.; Hansen, E.; Østerud, B.; Eilertsen, K.-E.; Bayer, A.; Engqvist, M.; Leszczak, K.; Jørgensen, T.Ø.; Andersen, J.H. Antioxidant and anti-inflammatory activities of barettin. Mar. Drugs 2013, 11, 2655–2666. [Google Scholar] [CrossRef] [PubMed]
- Volk, C.A.; Koeck, M. Viscosaline: New 3-alkyl pyridinium alkaloid from the Arctic sponge Haliclona viscosa. Org. Biomol. Chem. 2004, 2, 1827–1830. [Google Scholar] [CrossRef] [PubMed]
- Lippert, H.; Brinkmeyer, R.; Mulhaupt, T.; Iken, K. Antimicrobial activity in sub-Arctic marine invertebrates. Polar Biol. 2003, 26, 591–600. [Google Scholar] [CrossRef]
- Perry, N.B.; Ettouati, L.; Litaudon, M.; Blunt, J.W.; Munro, M.H.G.; Parkin, S.; Hope, H. Alkaloids from the antarctic sponge Kirkpatrickia varialosa. Part 1: Variolin B, a new antitumor and antiviral compound. Tetrahedron 1994, 50, 3987–3992. [Google Scholar] [CrossRef]
- Trimurtulu, G.; Faulkner, D.J.; Perry, N.B.; Ettouati, L.; Litaudon, M.; Blunt, J.W.; Munro, M.H.G.; Jameson, G.B. Alkaloids from the antarctic sponge Kirkpatrickia varialosa. Part 2: Variolin A and N(3’)-methyl tetrahydrovariolin B. Tetrahedron 1994, 50, 3993–4000. [Google Scholar] [CrossRef]
- Na, M.; Ding, Y.; Wang, B.; Tekwani, B.L.; Schinazi, R.F.; Franzblau, S.; Kelly, M.; Stone, R.; Li, X.-C.; Ferreira, D.; et al. Anti-infective discorhabdins from a deep-water Alaskan sponge of the senus Latrunculia. J. Nat. Prod. 2010, 73, 383–387. [Google Scholar] [CrossRef]
- Cutignano, A.; De Palma, R.; Fontana, A. A chemical investigation of the Antarctic sponge Lyssodendoryx flabellata. Nat. Prod. Res. 2012, 26, 1240–1248. [Google Scholar] [CrossRef]
- Cutignano, A.; Nuzzo, G.; D’Angelo, D.; Borbone, E.; Fusco, A.; Fontana, A. Mycalol: A natural lipid with promising cytotoxic properties against human anaplastic thyroid carcinoma cells. Angew. Chem. Int. Ed. 2013, 52, 9256–9260. [Google Scholar] [CrossRef]
- Holzwarth, M.; Trendel, J.-M.; Albrecht, P.; Maier, A.; Michaelis, W. Cyclic peroxides derived from the marine sponge Plakortis simplex. J. Nat. Prod. 2005, 68, 759–761. [Google Scholar] [CrossRef]
- Kong, F.; Andersen, R.J. Polymastiamide A, a novel steroid/amino acid conjugate isolated from the Norwegian marine sponge Polymastia boletiformis (Lamarck, 1815). J. Org. Chem. 1993, 58, 6924–6927. [Google Scholar] [CrossRef]
- Kong, F.; Andersen, R.J. Polymastiamides B-F, novel steroid/amino acid conjugates isolated from the Norwegian marine sponge Polymastia boletiformis. J. Nat. Prod. 1996, 59, 379–385. [Google Scholar] [CrossRef]
- Smyrniotopoulos, V.; Rae, M.; Soldatou, S.; Ding, Y.; Wolff, C.W.; McCormack, G.; Coleman, C.M.; Ferreira, D.; Tasdemir, D. Sulfated steroid-amino acid conjugates from the Irish marine sponge Polymastia boletiformis. Mar. Drugs 2015, 13, 1632–1646. [Google Scholar] [CrossRef] [PubMed]
- Hanssen, K.O.; Andersen, J.H.; Stiberg, T.; Engh, R.A.; Svenson, J.; Geneviere, A.M.; Hansen, E. Antitumoral and mechanistic studies of ianthelline isolated from the Arctic sponge Stryphnus fortis. Anticancer Res. 2012, 32, 4287–4297. [Google Scholar] [PubMed]
- Shin, J.; Seo, Y.; Rho, J.R.; Baek, E.; Kwon, B.-M.; Jeong, T.S.; Bok, S.H. Suberitenones A and B: Sesterterpenoids of an unprecedented skeletal class from the Antarctic sponge Suberites sp. J. Org. Chem. 1995, 60, 7582–7588. [Google Scholar] [CrossRef]
- Gompel, M.; Leost, M.; De Joffe, E.B.K.; Puricelli, L.; Franco, L.H.; Palermo, J.; Meijer, L. Meridianins, a new family of protein kinase inhibitors isolated from the ascidian Aplidium meridianum. Bioorg. Med. Chem. Lett. 2004, 14, 1703–1707. [Google Scholar] [CrossRef] [PubMed]
- Appleton, D.R.; Chuen, C.S.; Berridge, M.V.; Webb, V.L.; Copp, B.R. Rossinones A and B, biologically active meroterpenoids from the Antarctic ascidian, Aplidium species. J. Org. Chem. 2009, 74, 9195–9198. [Google Scholar] [CrossRef]
- Steffan, B. Lepadin A, a decahydroquinoline alkaloid from the tunicate Clavelina lepadiformis. Tetrahedron 1991, 47, 8729–8732. [Google Scholar] [CrossRef]
- Kubanek, J.; Williams, D.E.; de Silva, E.D.; Allen, T.; Andersen, R.J. Cytotoxic alkaloids from the flatworm Prostheceraeus villatus and its tunicate prey Clavelina lepadiformis. Tetrahedron Lett. 1995, 36, 6189–6192. [Google Scholar] [CrossRef]
- Diyabalanage, T.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Palmerolide A, a cytotoxic macrolide from the Antarctic tunicate Synoicum adareanum. J. Am. Chem. Soc. 2006, 128, 5630–5631. [Google Scholar] [CrossRef]
- Noguez, J.H.; Diyabalanage, T.; Miyata, Y.; Xie, X.-S.; Valeriote, F.A.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Palmerolide macrolides from the Antarctic tunicate Synoicum adareanum. Bioorg. Med. Chem. 2011, 19, 6608–6614. [Google Scholar] [CrossRef]
- Miyata, Y.; Diyabalanage, T.; Amsler, C.D.; McClintock, J.B.; Valeriote, F.A.; Baker, B.J. Ecdysteroids from the Antarctic tunicate Synoicum adareanum. J. Nat. Prod. 2007, 70, 1859–1864. [Google Scholar] [CrossRef] [PubMed]
- Tadesse, M.; Strøm, M.B.; Svenson, J.; Jaspars, M.; Milne, B.F.; Tørfoss, V.; Andersen, J.H.; Hansen, E.; Stensvåg, K.; Haug, T. Synoxazolidinones A and B: Novel bioactive alkaloids from the ascidian Synoicum pulmonaria. Org. Lett. 2010, 12, 4752–4755. [Google Scholar] [CrossRef]
- Tadesse, M.; Svenson, J.; Jaspars, M.; Strøm, M.B.; Abdelrahman, M.H.; Andersen, J.H.; Hansen, E.; Kristiansen, P.E.; Stensvåg, K.; Haug, T. Synoxazolidinone C; a bicyclic member of the synoxazolidinone family with antibacterial and anticancer activities. Tetrahedron Lett. 2011, 52, 1804–1806. [Google Scholar] [CrossRef]
- Tadesse, M.; Svenson, J.; Sepcic, K.; Trembleau, L.; Engqvist, M.; Andersen, J.H.; Jaspars, M.; Stensvag, K.; Haug, T. Isolation and synthesis of pulmonarins A and B, acetylcholinesterase inhibitors from the colonial ascidian Synoicum pulmonaria. J. Nat. Prod. 2014, 77, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Pettus, J.A., Jr.; Wing, R.M.; Sims, J.J. Marine natural products. XII. Isolation of a family of multihalogenated gamma-methylene lactones from the red seaweed Delisea fimbriata. Tetrahedron Lett. 1977, 18, 41–44. [Google Scholar] [CrossRef]
- Shilling, A.J.; Von Salm, J.L.; Sanchez, A.R.; Kee, Y.; Amsler, C.D.; McClintock, J.B.; Baker, B.J. Anverenes B-E, new polyhalogenated monoterpenes from the Antarctic red alga Plocamium cartilagineum. Mar. Drugs 2019, 17, 230. [Google Scholar] [CrossRef]
- Ankisetty, S.; Nandiraju, S.; Win, H.; Park, Y.C.; Amsler, C.D.; McClintock, J.B.; Baker, J.A.; Diyabalanage, T.K.; Pasaribu, A.; Singh, M.P.; et al. Chemical investigation of predator-deterred macroalgae from the Antarctic peninsula. J. Nat. Prod. 2004, 67, 1295–1302. [Google Scholar] [CrossRef]
- Nedialkova, D.; Naidenova, M. Screening the antimicrobial activity of actinomycetes strains isolated from Antarctica. J. Cult. Collect. 2005, 4, 29–35. [Google Scholar]
- Silber, J.; Ohlendorf, B.; Labes, A.; Erhard, A.; Imhoff, J.F. Calcarides A-E, antibacterial macrocyclic and linear polyesters from a Calcarisporium strain. Mar. Drugs 2013, 11, 3309–3323. [Google Scholar] [CrossRef]
- Encheva-Malinova, M.; Stoyanova, M.; Avramova, H.; Pavlova, Y.; Gocheva, B.; Ivanova, I.; Moncheva, P. Antibacterial potential of streptomycete strains from Antarctic soils. Biotechnol. Biotechnol. Equip. 2014, 28, 721–727. [Google Scholar] [CrossRef]
- Giudice, A.L.; Fani, R. Antimicrobial potential of cold-adapted bacteria and fungi from polar regions. In Biotechnology of Extremophiles: Advances and Challenges; Rampelotto, P.H., Ed.; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Parrilli, E.; Papa, R.; Carillo, S.; Tilotta, M.; Casillo, A.; Sannino, F.; Cellini, A.; Artini, M.; Selan, L.; Corsaro, M.M.; et al. Anti-biofilm activity of Pseudoalteromonas haloplanktis tac125 against Staphylococcus epidermidis biofilm: Evidence of a signal molecule involvement? Int. J. Immunopathol. Pharmacol. 2015, 28, 104–113. [Google Scholar] [CrossRef]
- Bosi, E.; Fondi, M.; Orlandini, V.; Perrin, E.; Maida, I.; de Pascale, D.; Tutino, M.L.; Parrilli, E.; Lo Giudice, A.; Filloux, A.; et al. The pangenome of (Antarctic) Pseudoalteromonas bacteria: Evolutionary and functional insights. BMC Genom. 2017, 18, 93. [Google Scholar] [CrossRef] [PubMed]
- Lipotec Antarcticine. Available online: https://www.lipotec.com/en/products/antarcticine-reg-marine-ingredient/ (accessed on 15 March 2020).
- Lipotec SeaCode. Available online: https://www.lipotec.com/en/products/seacode-trade-marine-ingredient/ (accessed on 15 March 2020).
- Knestrick, M.A.; Wilson, N.G.; Roth, A.; Adams, J.H.; Baker, B.J. Friomaramide, a highly modified linear hexapeptide from an Antarctic sponge, inhibits Plasmodium falciparum liver-stage development. J. Nat. Prod. 2019, 82, 2354–2358. [Google Scholar] [CrossRef]
- Ebada, S.S.; Lin, W.; Proksch, P. Bioactive sesterterpenes and triterpenes from marine sponges: Occurrence and pharmacological significance. Mar. Drugs 2010, 8, 313–346. [Google Scholar] [CrossRef] [PubMed]
- Elissawy, A.M.; El-Shazly, M.; Ebada, S.S.; Singab, A.B.; Proksch, P. Bioactive terpenes from marine-derived fungi. Mar. Drugs 2015, 13, 1966–1992. [Google Scholar] [CrossRef] [PubMed]
- Hegazy, M.E.; Mohamed, T.A.; Alhammady, M.A.; Shaheen, A.M.; Reda, E.H.; Elshamy, A.I.; Aziz, M.; Pare, P.W. Molecular architecture and biomedical leads of terpenes from red sea marine invertebrates. Mar. Drugs 2015, 13, 3154–3181. [Google Scholar] [CrossRef]
- Wu, Q.; Sun, J.; Chen, J.; Zhang, H.; Guo, Y.W.; Wang, H. Terpenoids from marine soft coral of the genus Lemnalia: Chemistry and biological activities. Mar. Drugs 2018, 16, 320. [Google Scholar] [CrossRef]
- McNaught, A.D.; Wilkinson, A. IUPAC Compendium of Chemical Terminology; International Union of Pure and Applied Chemistry: Hoboken, NJ, USA, 1997. [Google Scholar]
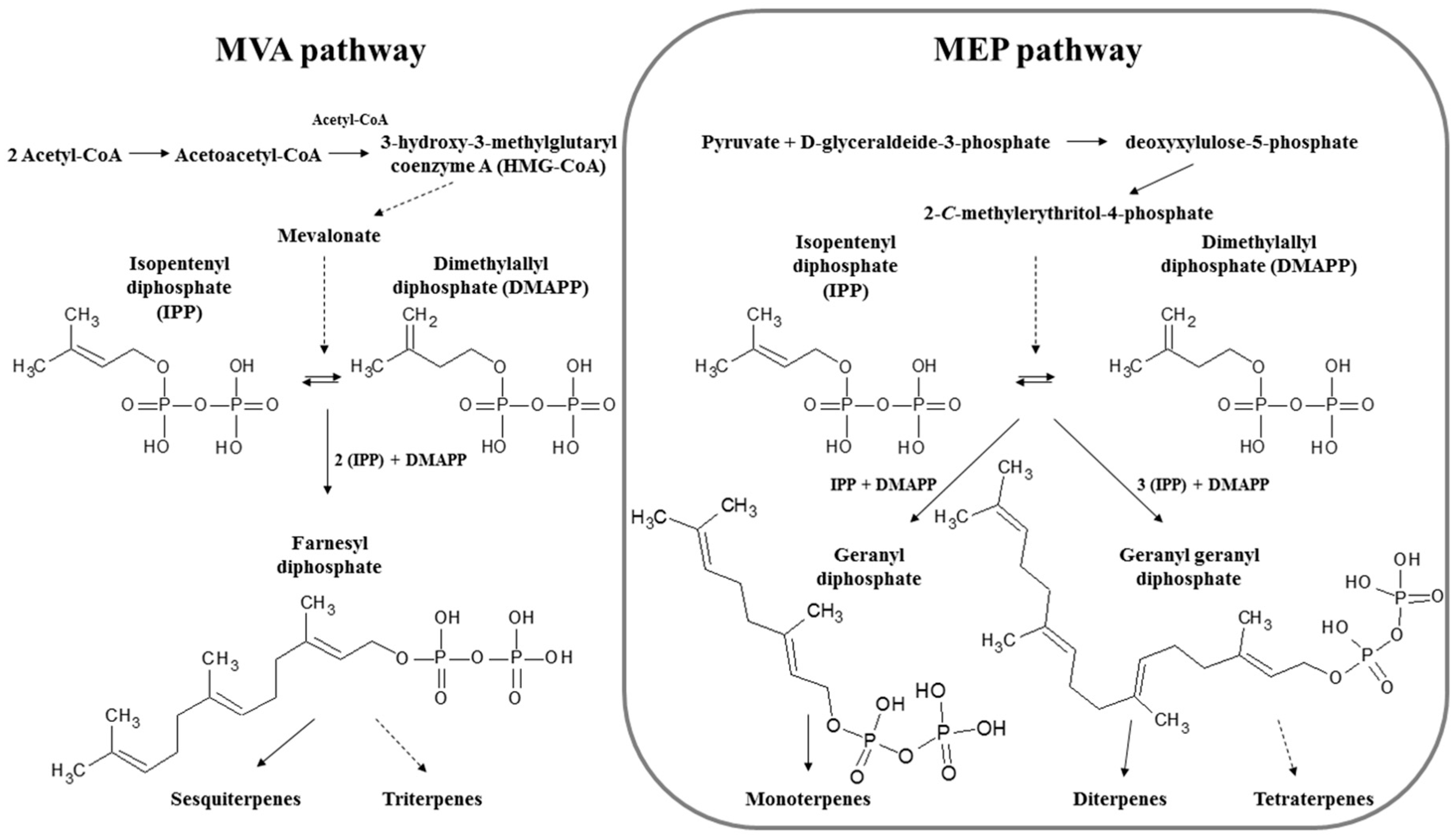
- Ruzicka, L. The isoprene rule and the biogenesis of terpenic compounds. Experientia 1953, 9, 357–367. [Google Scholar] [CrossRef]
- Davis, E.M.; Croteau, R. Cyclization enzymes in the biosynthesis of monoterpenes, sesquiterpenes, and diterpenes. In Biosynthesis: Aromatic Polyketides, Isoprenoids, Alkaloids; Leeper, F.J., Vederas, J.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 53–95. [Google Scholar]
- Leal, M.C.; Madeira, C.; Brandao, C.A.; Puga, J.; Calado, R. Bioprospecting of marine invertebrates for new natural products-a chemical and zoogeographical perspective. Molecules 2012, 17, 9842–9854. [Google Scholar] [CrossRef]
- Amsler, C.D.; McClintock, J.B.; Baker, B.J. Chemical mediation of mutualistic interactions between macroalgae and mesograzers structure unique coastal communities along the western Antarctic Peninsula. J. Phycol. 2014, 50, 1–10. [Google Scholar] [CrossRef]
- Kladi, M.; Vagias, C.; Roussis, V. Volatile halogenated metabolites from marine red algae. Phytochem. Rev. 2004, 3, 337–366. [Google Scholar] [CrossRef]
- Amsler, C.D.; Iken, K.B.; McClintock, J.B.; Amsler, M.O.; Peters, K.J.; Hubbard, J.M.; Furrow, F.B.; Baker, B.J. A comprehensive evaluation of the palatability and chemical defenses of subtidal macroalgae from the Antarctic peninsula. Mar. Ecol. Prog. Ser. 2005, 294, 141–159. [Google Scholar] [CrossRef]
- McClintock, J.B.; Baker, B.J. Marine Chemical Ecology; CRC Press: New York, NY, USA, 2001. [Google Scholar]
- Pérez, M.J.; Falqué, E.; Domínguez, H. Antimicrobial action of compounds from marine seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef] [PubMed]
- MarinLit, a Database of the Marine Natural Products Literature. Available online: http://pubs.rsc.org/marinlit/ (accessed on 3 March 2020).
- Fraga, B.M. Natural sesquiterpenoids. Nat. Prod. Rep. 2013, 30, 1226–1264. [Google Scholar] [CrossRef]
- Duran-Pena, M.J.; Ares, J.M.B.; Hanson, J.R.; Collado, I.G.; Hernandez-Galan, R. Biological activity of natural sesquiterpenoids containing a gem-dimethylcyclopropane unit. Nat. Prod. Rep. 2015, 32, 1236–1248. [Google Scholar] [CrossRef]
- Choudhary, A.; Naughton, L.M.; Montanchez, I.; Dobson, A.D.W.; Rai, D.K. Current status and future prospects of marine natural products (MNPs) as antimicrobials. Mar. Drugs 2017, 15, 272. [Google Scholar] [CrossRef]
- Bideau, F.L.; Kousara, M.; Chen, L.; Wei, L.; Dumas, F. Tricyclic sesquiterpenes from marine origin. Chem. Rev. 2017, 117, 6110–6159. [Google Scholar] [CrossRef]
- Kato, S.; Shindo, K.; Kawai, H.; Odagawa, A.; Matsuoka, M.; Mochizuki, J. Pyrrolostatin, a novel lipid peroxidation inhibitor from Streptomyces chrestomyceticus. Taxonomy, fermentation, isolation, structure elucidation and biological properties. J. Antibiot. (Tokyo) 1993, 46, 892–899. [Google Scholar] [CrossRef][Green Version]
- Liu, D.Z.; Liang, B.W. Structural elucidation and NMR assignments of two new pyrrolosesquiterpenes from Streptomyces sp. Hd7–21. Magn. Reson. Chem. 2014, 52, 57–59. [Google Scholar] [CrossRef]
- Bradfield, A.E.; Penfold, A.R.; Simonsen, J.L. 414. The constitution of eremophilone and of two related hydroxy-ketones from the wood oil of Eremophila mitchelli. J. Chem. Soc. 1932, 2744–2759. [Google Scholar] [CrossRef]
- Riche, C.; Pascard-Billy, C.; Devys, M.; Gaudemer, A.; Barbier, M.; Bousquet, J.-F. Structure cristalline et moleculaire de la phomenone, phytotoxine produite par le champignon Phoma exigua var. non oxydabilis. Tetrahedron Lett. 1974, 15, 2765–2766. [Google Scholar] [CrossRef]
- Yuyama, K.T.; Fortkamp, D.; Abraham, W.R. Eremophilane-type sesquiterpenes from fungi and their medicinal potential. Biol. Chem. 2018, 399, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, M.; Tang, J.; Li, X. Eremophilane sesquiterpenes from a deep marine-derived fungus, Aspergillus sp. SCSIOW2, cultivated in the presence of epigenetic modifying agents. Molecules 2016, 21, 473. [Google Scholar] [CrossRef]
- McDonald, L.A.; Barbieri, L.R.; Bernan, V.S.; Janso, J.; Lassota, P.; Carter, G.T. 07H239-A, a new cytotoxic eremophilane sesquiterpene from the marine-derived Xylariaceous fungus LL-07H239. J. Nat. Prod. 2004, 67, 1565–1567. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wang, J.; Huang, H.; Ma, L.; Wang, J.; Gu, Y.; Liu, L.; Lin, Y. Four eremophilane sesquiterpenes from the mangrove endophytic fungus Xylaria sp. BL321. Mar. Drugs 2012, 10, 340–348. [Google Scholar] [CrossRef]
- Torres, P.; Ayala, J.; Grande, C.; Anaya, J.; Grande, M. Furanoeremophilane derivatives from Senecio flavus. Phytochemistry 1999, 52, 1507–1513. [Google Scholar] [CrossRef]
- Mohamed, A.E.-H.; Ahmed, H. Eremophilane-type sesquiterpene derivatives from Senecio aegyptius var. discoideus. J. Nat. Prod. 2005, 68, 439–442. [Google Scholar] [CrossRef]
- Tori, M.; Otose, K.; Fukuyama, H.; Murata, J.; Shiotani, Y.; Takaoka, S.; Nakashima, K.; Sono, M.; Tanaka, M. New eremophilanes from Farfugium japonicum. Tetrahedron 2010, 66, 5235–5243. [Google Scholar] [CrossRef]
- Harinantenaina, L.; Matsunami, K.; Otsuka, H. Chemical and biologically active constituents of Pteris multifida. J. Nat. Med. 2008, 62, 452–455. [Google Scholar] [CrossRef]
- Fabian, K.; Lorenzen, K.; Anke, T.; Johansson, M.; Sterner, O. Five new bioactive sesquiterpenes from the fungus Radulomyces confluens (Fr.) Christ. Z. Naturforsch. C. J. Biosci. 1998, 53, 939–945. [Google Scholar] [CrossRef]
- Mogi, Y.; Inanaga, K.; Tokuyama, H.; Ihara, M.; Yamaoka, Y.; Yamada, K.-i.; Takasu, K. Rapid assembly of protoilludane skeleton through tandem catalysis: Total synthesis of paesslerin A and its structural revision. Org. Lett. 2019, 21, 3954–3958. [Google Scholar] [CrossRef] [PubMed]
- Iken, K.B.; Baker, B.J. Ainigmaptilones, sesquiterpenes from the Antarctic gorgonian coral Ainigmaptilon antarcticus. J. Nat. Prod. 2003, 66, 888–890. [Google Scholar] [CrossRef]
- Cano, L.P.P.; Manfredi, R.Q.; Pérez, M.; García, M.; Blustein, G.; Cordeiro, R.; Pérez, C.D.; Schejter, L.; Palermo, J.A. Isolation and antifouling activity of azulene derivatives from the Antarctic gorgonian Acanthogorgia laxa. Chem. Biodivers. 2018, 15, e1700425. [Google Scholar] [CrossRef] [PubMed]
- Iken, K.; Avila, C.; Ciavatta, M.L.; Fontana, A.; Cimino, G. Hodgsonal, a new drimane sesquiterpene from the mantle of the Antarctic nudibranch Bathydoris hodgsoni. Tetrahedron Lett. 1998, 39, 5635–5638. [Google Scholar] [CrossRef]
- Avila, C. Terpenoids in marine heterobranch molluscs. Mar. Drugs 2020, 18, 162. [Google Scholar] [CrossRef]
- Hortelano, S. Molecular basis of the anti-inflammatory effects of terpenoids. Inflamm. Allergy Drug Targets 2009, 8, 28–39. [Google Scholar]
- Gao, S.S.; Li, X.M.; Zhang, Y.; Li, C.S.; Wang, B.G. Conidiogenones H and I, two new diterpenes of cyclopiane class from a marine-derived endophytic fungus Penicillium chrysogenum QEN-24S. Chem. Biodivers. 2011, 8, 1748–1753. [Google Scholar] [CrossRef]
- Du, L.; Li, D.; Zhu, T.; Cai, S.; Wang, F.; Xiao, X.; Gu, Q. New alkaloids and diterpenes from a deep ocean sediment derived fungus Penicillium sp. Tetrahedron 2009, 65, 1033–1039. [Google Scholar] [CrossRef]
- Roncal, T.; Cordobes, S.; Sterner, O.; Ugalde, U. Conidiation in Penicillium cyclopium is induced by conidiogenone, an endogenous diterpene. Eukaryot. Cell 2002, 1, 823–829. [Google Scholar] [CrossRef]
- Cheng, Z.; Li, Y.; Xu, W.; Liu, W.; Liu, L.; Zhu, D.; Kang, Y.; Luo, Z.; Li, Q. Three new cyclopiane-type diterpenes from a deep-sea derived fungus Penicillium sp. YPGA11 and their effects against human esophageal carcinoma cells. Bioorg. Chem. 2019, 91, 103129. [Google Scholar] [CrossRef]
- Gonzalez, Y.; Torres-Mendoza, D.; Jones, G.E.; Fernandez, P.L. Marine diterpenoids as potential anti-inflammatory agents. Mediat. Inflamm. 2015, 2015, 263543. [Google Scholar] [CrossRef] [PubMed]
- Dayton, P.K.; Bobilliard, G.A.; Paine, R.T.; Dayton, L.B. Biological accomodation in the benthic community at McMurdo Sound, Antarctica. Ecol. Monograph. 1974, 44, 105–128. [Google Scholar] [CrossRef]
- Nunez-Pons, L.; Carbone, M.; Vazquez, J.; Gavagnin, M.; Avila, C. Lipophilic defenses from alcyonium soft corals of Antarctica. J. Chem. Ecol. 2013, 39, 675–685. [Google Scholar] [CrossRef]
- Cattaneo-Vietti, R.; Chiantore, M.; Schiaparelli, S.; Albertelli, G. Shallow- and deep-water mollusc distribution at Terra Nova Bay (Ross Sea, Antarctica). Polar Biol. 2000, 23, 173–182. [Google Scholar] [CrossRef]
- Gavagnin, M.; De Napoli, A.; Castelluccio, F.; Cimino, G. Austrodorin-A and -B: First tricyclic diterpenoid 2’-monoglyceryl esters from an Antarctic nudibranch. Tetrahedron Lett. 1999, 40, 8471–8475. [Google Scholar] [CrossRef]
- Davies-Coleman, M.T.; Faulkner, D.J. New diterpenoic acid glycerides from the Antarctic nudibranch Austrodoris kerguelensis. Tetrahedron 1991, 47, 9743–9750. [Google Scholar] [CrossRef]
- Arens, J.; Bergs, D.; Mewes, M.; Merz, J.; Schembecker, G.; Schulz, F. Heterologous fermentation of a diterpene from Alternaria brassisicola. Mycology 2014, 5, 207–219. [Google Scholar] [CrossRef]
- Xu, R.; Fazio, G.C.; Matsuda, S.P. On the origins of triterpenoid skeletal diversity. Phytochemistry 2004, 65, 261–291. [Google Scholar] [CrossRef]
- Lednicer, D. Steroid Chemistry at a Glance; Wiley: Hoboken, NJ, USA, 2010. [Google Scholar]
- Mijts, B.N.; Lee, P.C.; Schmidt-Dannert, C. Engineering carotenoid biosynthetic pathways. Methods Enzymol. 2004, 388, 315–329. [Google Scholar]
- Fiedor, J.; Burda, K. Potential role of carotenoids as antioxidants in human health and disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef]
- Lohr, M. Carotenoids. In The Chlamydomonas Sourcebook, 2nd ed.; Harris, E.H., Stern, D.B., Witman, G.B., Eds.; Academic Press: London, UK, 2009; Chapter 21; pp. 799–817. [Google Scholar]
- Gonzalez, S.; Gilaberte, Y.; Philips, N.; Juarranz, A. Current trends in photoprotection-A new generation of oral photoprotectors. Open Dermatol. J. 2011, 5, 6–14. [Google Scholar] [CrossRef]
- Galasso, C.; Corinaldesi, C.; Sansone, C. Carotenoids from marine organisms: Biological Ffunctions and industrial applications. Antioxidants 2017, 6, 96. [Google Scholar] [CrossRef]
- Matsuno, T. Aquatic animal carotenoids. Fish. Sci. 2001, 67, 771–783. [Google Scholar] [CrossRef]
- Liao, L.; Su, S.; Zhao, B.; Fan, C.; Zhang, J.; Li, H.; Chen, B. Biosynthetic potential of a novel Antarctic actinobacterium Marisediminicola antarctica ZS314(T) revealed by genomic data mining and pigment characterization. Mar. Drugs 2019, 17, 388. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.B.; Flores, E.M.M.; Barin, J.S.; Mercadante, A.Z.; Jacob-Lopes, E.; Zepka, L.Q. Production of carotenoids from microalgae cultivated using agroindustrial wastes. Food Res. Int. 2014, 65, 144–148. [Google Scholar] [CrossRef]
- Da Silva, N.A.; Rodrigues, E.; Mercadante, A.Z.; de Rosso, V.V. Phenolic compounds and carotenoids from four fruits native from the Brazilian Atlantic forest. J. Agri. Food Chem. 2014, 62, 5072–5084. [Google Scholar] [CrossRef] [PubMed]
- Gharibzahedi, S.M.T.; Razavi, S.H.; Mousavi, S.M. Characterization of bacteria of the genus Dietzia: An updated review. Ann. Microbiol. 2014, 64, 1–11. [Google Scholar] [CrossRef]
- Guyomarc’h, F.; Binet, A.; Dufosse, L. Production of carotenoids by Brevibacterium linens: Variation among strains, kinetic aspects and HPLC profiles. J. Ind. Microbiol. Biotech. 2000, 24, 64–70. [Google Scholar] [CrossRef]
- Wagener, S.; Volker, T.; De Spirt, S.; Ernst, H.; Stahl, W. 3,3’-Dihydroxyisorenieratene and isorenieratene prevent UV-induced DNA damage in human skin fibroblasts. Free Radic. Biol. Med. 2012, 53, 457–463. [Google Scholar] [CrossRef]
- Kalinin, V.I.; Silchenko, A.S.; Avilov, S.A.; Stonik, V.A.; Smirnov, A.V. Sea cucumbers triterpene glycosides, the recent progress in structural elucidation and chemotaxonomy. Phytochem. Rev. 2005, 4, 221–236. [Google Scholar] [CrossRef]
- Antonov, A.S.; Avilov, S.A.; Kalinovsky, A.I.; Anastyuk, S.D.; Dmitrenok, P.S.; Evtushenko, E.V.; Kalinin, V.I.; Smirnov, A.V.; Taboada, S.; Ballesteros, M.; et al. Triterpene glycosides from Antarctic Sea cucumbers. 1. Structure of liouvillosides A1, A2, A3, B1, and B2 from the sea cucumber Staurocucumis liouvillei: New procedure for separation of highly polar glycoside fractions and taxonomic revision. J. Nat. Prod. 2008, 71, 1677–1685. [Google Scholar] [CrossRef] [PubMed]
- Howes, F.N. Fish-poison plants. Bull. Misc. Infor. (R. Bot. Gard. Kew) 1930, 1930, 129–153. [Google Scholar] [CrossRef]
- Augustin, J.M.; Kuzina, V.; Andersen, S.B.; Bak, S. Molecular activities, biosynthesis and evolution of triterpenoid saponins. Phytochemistry 2011, 72, 435–457. [Google Scholar] [CrossRef] [PubMed]
- Lorent, J.H.; Quetin-Leclercq, J.; Mingeot-Leclercq, M.P. The amphiphilic nature of saponins and their effects on artificial and biological membranes and potential consequences for red blood and cancer cells. Org. Biomol. Chem. 2014, 12, 8803–8822. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.X.; Xie, Y.; Ye, Y.P. Advances in saponin-based adjuvants. Vaccine 2009, 27, 1787–1796. [Google Scholar] [CrossRef]
- Bahrami, Y.; Zhang, W.; Chataway, T.; Franco, C. Structure elucidation of five novel isomeric saponins from the viscera of the sea cucumber Holothuria lessoni. Mar. Drugs 2014, 12, 4439–4473. [Google Scholar] [CrossRef]
- Antonov, A.S.; Avilov, S.A.; Kalinovsky, A.I.; Anastyuk, S.D.; Dmitrenok, P.S.; Kalinin, V.I.; Taboada, S.; Bosh, A.; Avila, C.; Stonik, V.A. Triterpene glycosides from Antarctic sea cucumbers. 2. Structure of achlioniceosides A(1), A(2), and A(3) from the sea cucumber Achlionice violaecuspidata (=Rhipidothuria racowitzai). J. Nat. Prod. 2009, 72, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Silchenko, A.S.; Kalinovsky, A.I.; Avilov, S.A.; Andryjashchenko, P.V.; Fedorov, S.N.; Dmitrenok, P.S.; Yurchenko, E.A.; Kalinin, V.I.; Rogacheva, A.V.; Gebruk, A.V. Kolgaosides A and B, two new triterpene glycosides from the Arctic deep water sea cucumber Kolga hyalina (Elasipodida: Elpidiidae). Nat. Prod. Commun. 2014, 9, 1259–1264. [Google Scholar] [CrossRef]
- Menna, M.; Imperatore, C.; D’Aniello, F.; Aiello, A. Meroterpenes from marine invertebrates: Structures, occurrence, and ecological implications. Mar. Drugs 2013, 11, 1602–1643. [Google Scholar] [CrossRef]
- Hemmings, A. A question of politics: Bioprospecting and the Antarctic Treaty System. Antarctic Bioprospecting; University of Canterbury: Christchurch, New Zealand, 2005; pp. 98–129. [Google Scholar]
- Hemmings, A. Re-Justifying the Antarctic Treaty System for the 21st Century: Rights, Expectations and Global Equity. In Polar Geopolitics: Knowledges, Resources and Legal Regimes; Edward Elgar Publishing: Cheltenham, UK, 2014; pp. 55–73. [Google Scholar]
- Greiber, T. Access and Benefit Sharing in Relation to Marine Genetic Resources from Areas Beyond National Jurisdiction: A Possible Way Forward; Study in Preparation of the Informal Workshop on Conservation of Biodiversity Beyond National Jurisdiction. In Research Project of the Federal Agency for Nature Conservation; Bundesamt für Naturschutz: Bonn, Germany, 2011. [Google Scholar]
- Greiber, T. An Explanatory Guide to the Nagoya Protocol on Access and Benefit-Sharing; IUCN: Gland, Switzerland, 2012. [Google Scholar]
- Wilkinson, M.D.; Dumontier, M.; Aalbersberg, I.J.; Appleton, G.; Axton, M.; Baak, A.; Blomberg, N.; Boiten, J.-W.; da Santos, L.B.S.; Bourne, P.E.; et al. The FAIR Guiding Principles for scientific data management and stewardship. Sci. Data 2016, 3, 160018. [Google Scholar] [CrossRef]
- Thompson, C.C.; Kruger, R.H.; Thompson, F.L. Unlocking marine biotechnology in the developing world. Trends Biotech. 2017, 35, 1119–1121. [Google Scholar] [CrossRef] [PubMed]
- Blasiak, R.; Jouffray, J.-B.; Wabnitz, C.C.C.; Sundström, E.; Österblom, H. Corporate control and global governance of marine genetic resources. Sci. Adv. 2018, 4, eaar5237. [Google Scholar] [CrossRef] [PubMed]
- Hofland, P. Plitidepsin–A novel anti-cancer agent possibly active against COVID19. Onco’Zine. 2020. Available online: https://www.oncozine.com/plitidepsin-a-novel-anti-cancer-agent-possibly-active-against-covid18/ (accessed on 27 July 2020).
- Andrade-Ochoa, S.; Nevárez-Moorillón, G.V.; Sánchez-Torres, L.E.; Villanueva-García, M.; Sánchez-Ramírez, B.E.; Rodríguez-Valdez, L.M.; Rivera-Chavira, B.E. Quantitative structure-activity relationship of molecules constituent of different essential oils with antimycobacterial activity against Mycobacterium tuberculosis and Mycobacterium bovis. BMC Complement. Altern. Med. 2015, 15, 332. [Google Scholar] [CrossRef] [PubMed]






























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Collection Site (Distribution) | Compound | Molecule Type | Bioactivity | Reference |
|---|---|---|---|---|---|
| BACTERIA | |||||
| Plylum Actinobacteria | |||||
| Class Actinobacteria | |||||
| Arthrobacter sp. | Marine sediment, Terranova Bay, Ross Sea (Antarctica); | monoramnholipids | rhamnolipid | Antimicrobial activity against Burkholderia cepacia complex | [70] |
| Surface water, sea ice, zooplankton, the deep sea, and meltwater (Arctic Ocean) | arthrobacilins A–C | cyclic glycolipids | Antimicrobial activity against Vibrio anguillarum and Staphylococcus aureus | [71] | |
| Nocardia dassonvillei | Marine sediment (Arctic Ocean) | N-(2-hydroxyphenyl)-2-phenazinamine, 1,6-dihydroxyl-phenazine, 6-hydroxyl phenazine-1-carboxylic acid, 6-methoxy-1-phenazinol, 2-amino-1, 4-naphthoquinone, 2-amino-phenoxazin-3-one and 2-(N-methylamino)-3-phenoxazone | phenazine, phenoxazine, naphthoquinone | Antifungal activity against Candida albicans and high cancer cell cytotoxicity against HepG2, A549, HCT-116 and COC1 cells | [72] |
| Nocardiopsis SCSIO KS107 | Seashore sediment sample, China Great Wall station (Antarctica) | 7-hydroxymucidone, 4-hydroxymucidone, germicidin H | α-pyrones | Antibacterial activity against Micrococcus luteus and Bacillus subtilis | [73] |
| Rhodococcus sp. B7740 | Deep seawater (Arctic Ocean) | isorenieratene (116) | carotenoid | Antioxidant activity | [74,75] |
| menaquinone MK8(H2) (115) | isoprenoid quinone | Antioxidant and antiglycation activities | [76] | ||
| Streptomyces sp. | Marine surface sediment of the East Siberian continental margin (Arctic Ocean) | arcticoside, C-1027 chromophore-V, III, fijiolides A and B | benzoxazines, glycosylated paracyclophane | Cytotoxic activity against breast carcinoma MDA-MB231 cells and colorectal carcinoma cells (line HCT-116) | [77] |
| Streptomyces strain 1010 | Shallow sea sediment from the region of Livingston Island (Antarctica) | phthalic acid diethyl ester, 1,3-bis(3-phenoxyphenoxy)benzene, exanedioic acid dioctyl ester, 2-amino-9,13-dimethyl heptadecanoic acid | aromatic compounds | No activity tested * | [78] |
| Streptomyces sp. SCO736 | Marine sediment (Antarctica) | antartin (10) | zizaane-type sesquiterpene | Cytotoxic activity against A549, H1299 and U87 cancer cell lines by causing cell cycle arrest at the G1 phase | [79] |
| Streptomyces sp. NPS008187 | Alaskan marine sediment (Arctic Ocean) | glyciapyrroles A (11), B (12) and C (13), cyclo(leucyl-prolyl), cyclo(isoleucyl-prolyl), cyclo(phenylalanyl-prolyl) | pyrrolosesquiterpenes, diketopiperazines | Cytotoxic activity against colorectal adenocarcinoma HT-29 and melanoma B16-F10 | [80] |
| Plylum Proteobacteria | |||||
| Class γ-Proteobacteria | |||||
| Pseudoalteromonas haloplanktis TAC125 | French Antarctic station Dumont d’Urville, Terre Adélie (Antarctica) | methylamine | Volatile Organic Compounds (VOCs) | Antimicrobial activity against B. cepacia complex | [81] |
| 4-hydroxybenzoic acid | benzoic acid derivative | Antitumor activity against human A459 lung adenocarcinoma cells | [82] | ||
| cyclo-(D-pipecolinyl-L-isoleucine), cyclo-(L-prolyl-L-histidine), cyclo-(L-prolyl-L-alanine), cyclo-(L-prolyl-L-tyrosine), cyclo-(L-prolyl-L-proline), cyclo-(L-alanyl-L-isoleucine), cyclo-(D-pipecolinyl-L-leucine), cyclo-(L-pipecolinyl-L-phenylalanine), L-valyl-L-leucyl-Lprolyl-L-valyl-L-prolyl-L-glutamine and L-tyrosyl-L-valyl-L-prolyl-L-leucine | diketopiperazines | Antioxidant activity | [83] | ||
| pentadecanal | long-chain fatty aldehyde | Anti-biofilm activity against Staphylococcus epidermidis | [84] | ||
| Pseudomonas sp. | Terranova Bay, Ross Sea (Antarctica) | monoramnholipids | rhamnolipid | Antimicrobial activity against B. cepacia complex | [70] |
| Psychrobacter | Marine sediment from Terranova Bay, Ross Sea (Antarctica) | monoramnholipids | rhamnolipid | Antimicrobial activity against B. cepacia complex | [70] |
| Plylum Bacteroidetes | |||||
| Class Flavobacteriia | |||||
| Aequorivita | Marine sediments from Edmonson Point (Antarctica) | R-(+)-N-[15-methyl-3-(12-methyltridecanoyloxy)-hexadecanoyl]glycine and methyl ester derivatives; | aminolipids | Antimicrobial activity against S. aureus | [85] |
| Salegentibacter strain T436 | Bottom section of a sea ice floe (Arctic Ocean) | 4-Hydroxy-3-nitrobenzoic acid, 4,6-Dinitroguiacol, 4,5-Dinitro-3-methoxyphenol, (4-Hydroxy-3-nitrophenyl)-acetic acid methyl ester, (4-Hydroxy-3,5-dinitrophenyl)-acetic acid methyl ester, (4-Hydroxy-3-nitrophenyl)-acetic acid, (4-Hydroxy-3,5-dinitrophenyl)-acetic acid, (4-Hydroxy-3,5-dinitrophenyl)-propionic acid methyl ester, (4-Hydroxy-3-nitrophenyl)-propionic acid, (4-Hydroxy-3,5-dinitrophenyl)-propionic acid, 2-Chloro-3-(4-hydroxy-3,5-dinitrophenyl)-propionic acid methyl ester, 2-Hydroxy-3-(4-hydroxy-3-nitrophenyl)-propionic acid methyl ester, 2-(4-Hydroxy-3-nitrophenyl)-ethanol, 2-(4-Hydroxy-3,5-dinitrophenyl)-ethyl chloride, 2-(4-Hydroxy-3,5-dinitrophenyl)-ethanol, 2-Nitro-4-(2 -nitroethenyl)-phenol, 3,5-Dinitrogenistein, 3-Nitrogenistein, 3-Nitro-1H-indole | aromatic nitro compounds | Antimicrobial activity against C. albicans, Paecilomyces variotii, Penicillium notatumb, Mucor miehei Tü 284, Magnaporthe grisea, Nematospora coryli, Ustilago nuda, Bacillus brevis, B. subtilis, M. luteus, Escherichia coli K12b, Proteus vulgaris and cytotoxic activities | [86,87] |
| Plylum Firmicutes | |||||
| Class Bacilli | |||||
| Bacillus sp. | Sea mud (Arctic Ocean) | mixirins A, B and C | cyclopeptides | Cytotoxic activity against human colon tumor cells (HCT-116) | [88] |
| FUNGI | |||||
| Plylum Ascomycota | |||||
| Class Eurotiomycetes | |||||
| Aspergillus protuberus MUT 3638 | Sub-sea sediments, Barents Sea (Arctic Ocean) | bisvertinolone | sorbicillonoid | Antimicrobial activity against S. aureus | [89] |
| Penicillium sp. PR19N-1 | Deep-sea sediment, Prydz Bay (Antarctica) | chlorinated eremophilane sesquiterpenes (14–17), eremofortine C (18) | chloro-eremophilane sesquiterpenes | Cytotoxic activity against HL-60 and A549 cancer cell lines | [90] |
| eremophilane-type sesquiterpenes (19–23, 25, 26), eremophilane-type lactam (24, 27) | eremophilane-type sesquiterpenes | Cytotoxic activity against HL-60 and A549 cancer cell lines | [91] | ||
| Penicillium granulatum MCCC 3A00475 | Deep-sea sediment, Prydz Bay (Antarctica) | spirograterpene A (37), conidiogenone I (38) and conidiogenone C (39) | tetracyclic spiro-diterpene, cyclopiane diterpenes | Spirograterpene A: antiallergic effect on immunoglobulin E (IgE)-mediated rat mast RBL-2H3 cells | [92] |
| Penicillium sp. S-1-18 | Sea-bed sediments (Antarctica) | butanolide, guignarderemophilane F, xylarenone A (28) | furanone derivative, sesquiterpene | Butanolide: inhibitory activity of butanolide against tyrosine phosphatase 1B; xylarenone A: antitumor activity against HeLa and HepG2 cells and growth-inhibitory effects against pathogenic microbes | [93,94] |
| Penicillium crustosum PRB-2 | Deep-sea sediment, Prydz Bay (Antarctica) | penilactones A and B | oxygenated polyketides | Cytotoxic activity against HCT-8, Bel-7402, BGC-823, A549 and A2780 tumor cell lines | [95] |
| Class Dothideomycetes | |||||
| Strain KF970 (Lindgomycetaceae family) | Sea-water (Arctic Ocean) | lindgomycin, ascosetin | polyketides | Antimicrobial activity against methicillin-resistant S. aureus | [96] |
| ALGAE-ASSOCIATED MICROBES | |||||
| Bacteria-Plylum Actinobacteria-Class Actinobacteria | |||||
| Nocardiopsis sp. 03N67 | Arctic seaweed (Undaria pinnatifida) | cyclo-(L-Pro-L-Met) | diketopiperazine | Anti-angiogenesis activity against human umbilical vein endothelial cells (HUVECs) | [97] |
| SPONGE-ASSOCIATED MICROBES | |||||
| Bacteria-Plylum Bacteroidetes-Class Flavobacteriia | |||||
| Cellulophaga fucicola | Antarctic sea sponge | zeaxanthin (110), β-cryptoxanthin (111), β-carotene (112) | carotenoids | Antioxidant activity | [98] |
| Zobellia laminarie | Antarctic sea sponge | zeaxanthin (110), β-cryptoxanthin (111), β-carotene (112), phytoene (113) | carotenoids | Anti-UV and antioxidant activity and phototoxicity profile in murine fibroblasts | [99] |
| Bacteria-Plylum Proteobacteria-Class γ-Proteobacteria | |||||
| Pseudomonas aeruginosa | Isodictya setifera, Ross Island (Antarctica) | cyclo-(L-Pro-L-Val)/cyclo-(L-Pro-L-Leu)/cyclo-(L-Pro-L-Ile)/cyclo-(L-Pro-L-Phe)/cyclo-(L-Pro-L-Tyr)/cyclo-(L-Pro-L-Met)/diketopiperazines, phenazine-1-carboxylic acid, phenazine-1-carboxamide | diketopiperazines, phenazine alkaloids | Antimicrobial activity against B. subtilis, S. aureus and M. luteus | [65] |
| Fungi-Plylum Ascomycota-Class Leotiomycetes | |||||
| Pseudogymnoascus sp. F09-T18-1 | Antarctic sponge genus Hymeniacidon, Fildes Bay, King George Island (Antarctica) | pseudogymnoascin A, B, C, 3-nitroasterric acid | nitroasterric acid derivatives | Inactive in antimicrobial activity against P. aeruginosa, Acinetobacter baumannii, E. coli, S. aureus, a methicillin-sensitive S. aureus and methicillin-resistant S. aureus, C. albicans, Aspergillus fumigatu | [100] |
| CORAL-ASSOCIATED MICROBES | |||||
| Fungi-Plylum Ascomycota-Class Eurotiomycetes | |||||
| Penicillium sp. SF-5995 | Unidentified soft coral, Terra Nova Bay (Antarctica) | methylpenicinoline | pyrrolyl 4-quinoline alkaloid | Anti-inflammatory effect inhibiting NF-κB and MAPK pathways in lipopolysaccharide-induced RAW264.7 macrophages and BV2 microglia | [101] |
| CNIDARIA | |||||
| Plylum Cnidaria-Class Anthozoa | |||||
| Alcyonium antarcticum | Terra Nova Bay (Antarctica) | bulgarane sesquiterpene | sesquiterpene | No bioactivity tested (antipredation activity and ichthyotoxicity) * | [102] |
| Alcyonium antarcticum | Weddell Sea (Antarctica) | alcyopterosins | illudalane sesquiterpenoids | No bioactivity tested (antipredation activity) * | [103] |
| Alcyonium paessleri | South Georgia Islands (Antarctica) | alcyopterosin A (29), C (30), E (31), H (32) | illudalane sesquiterpenoids | Cytotoxic activity against Hep-2 (human larynx carcinoma) and HT-29 (human colon carcinoma) cell lines | [104] |
| Alcyonium paessleri | South Georgia Islands (Antarctica) | paesslerins A (33), B (34) | esquiterpenoids | Cytotoxic activity against human tumor cell lines | [105] |
| Anthomastus bathyproctus | Deception Island, South Shetland Islands (Antarctica) | conjugated cholestane, ergostane and 24-norcholestane steroids (79–82) | steroids | Cytotoxic activity against three human tumor cell lines. | [106] |
| Anthoptilum grandiflorum | Burdwood Bank, Scotia Arc (Antarctica) | bathyptilone A (48), B, C, enbepeanone A | briarane diterpenes, trinorditerpene | Bathyptilone A: cytotoxic activity against the neurogenic mammalian cell line Ntera-2 | [107] |
| Dasystenella acanthina | Kapp Norvegia, Eastern Weddell Sea (Antarctica) | 7 polyoxygenated steroids (83–89) | steroids | Growth inhibition of several human tumor cell lines LN-caP and K-562 | [108] |
| Gersemia fruticosa | White Sea (circumpolar Arctic) | 6 polyoxygenated sterols (90–95) | sterols | Cytotoxic activity against human erythroleukemia K-562 cells, HL-60 and P388 | [109] |
| Gersemia fruticosa | White Sea (circumpolar Arctic) | 9,11-secosterol (96) | sterol | Cytotoxic activity against human leukemia K562, cervical cancer HeLa and Ehrlich ascites tumor cells | [110] |
| Gersemia fruticosa | Alaskan Beaufort Sea (Arctic Ocean) | gersemiols A–C, eunicellol A (40) | diterpenoids | Eunicellol A: antimicrobial activity against MRSA—methicillin resistant S. aureus | [111] |
| Plumarella delicatissima | Plateau of Fascination, Falkland Islands (Antarctica) | keikipukalides A–E (41–45), pukalide aldehyde (46), norditerpenoid ineleganolide (47) | diterpenes, diterpenoid | Cytotoxic activity against leishmaniasis causing a parasite, Leishmania donovani, with no cytotoxicity against the mammalian host | [112] |
| Undescribed soft coral | Scotia Arc (Antarctica) | shagenes A (35), B (36) | sesquiterpenoids | Cytotoxic activity against leishmaniasis causing a parasite, L. donovani, with no cytotoxicity against the mammalian host | [113] |
| BRYOZOA | |||||
| Plylum Bryozoa-Class Gymnolaemata | |||||
| Tegella cf. spitzbergensis | Bear Island (Arctic Ocean) | ent-eusynstyelamide B, eusynstyelamides D–F | brominated tryptophan-derived | Antimicrobial activity against bacteria; weak cytotoxicity against the human melanoma A2058 cell line | [114] |
| Dendrobeania murrayana | Vesterålsfjorden, Northern Norway (Arctic Ocean) | dendrobeaniamine A | guanidine alkaloid | Tested but inactive for cytotoxic, antimicrobial, anti-inflammatory or antioxidant activities | [115] |
| Alcyonidium gelatinosum | Hopenbanken, Svalbard (Arctic Ocean) | ponasterone A and F | ecdysteroids | Tested but inactive for cytotoxic, antimicrobial, estrogen receptor agonist activities | [52] |
| MOLLUSCA | |||||
| Plylum Mollusca-Class Gasteropoda | |||||
| Austrodoris kerguelenensis | Anvers Island (Circumpolar Antarctica) | palmadorin A (49), B (50), D (51), M (52), N (53), O (54) | diterpenoid glyceride esters | Inhibition of human erythroleukemia (HEL) cells; Palmadorin M inhibits Jak2, STAT5, and Erk1/2 activation in HEL cells | [116] |
| ECHINODERMATA | |||||
| Plylum Echinodermata- | |||||
| Class Holothuroidea | |||||
| Kolga hyalina | Deep sea, Amundsen Basin (Arctic Ocean) | holothurinoside B, kolgaosides A (120), B (121) | triterpene holostane nonsulfated pentaosides | Kolgaosides A–B: hemolytic activity against mouse erythrocytes and inhibition against Ehrlich ascite carcinoma cells | [117] |
| Staurocucumis liouvillei | South Georgia Islands (Antarctica) | liouvillosides A (118), B (119) | trisulfated triterpene glycosides | Activity against herpes simplex virus type 1 (HSV-1) | [118] |
| Class Astheroidea | |||||
| Asterias microdiscus | Chukchi Sea (Arctic Ocean) | polyhydroxylated steroids A–F | steroids | No activity tested * | [119] |
| Class Ophiuroidea | |||||
| Ophiosparte gigas | Ross Sea (Antarctica) | cholest-5-ene-2α,3α,4β,21-tetrao1-3,21-disulphate (102), cholest-5-ene-2β,3α, 21-triol-2,21-disulphate (103) | disulfated polyhydroxysteroids | cholest-5-ene-2α,3α,4β,21-tetrao1-3,21-disulphate: cytotoxic activity; cholest-5-ene-2β,3α, 21-triol-2,21-disulphate: cytoprotective activity against HIV-1 | [120] |
| Astrotoma agassizii | Antarctica | disulfated polyhydroxysteroids (104–106) | disulfated polyhydroxysteroids | Activity against one DNA (HSV-2) and two RNA (PV-3, JV) viruses | [121] |
| SPONGES | |||||
| Plylum Porifera | |||||
| Class Demospongiae | |||||
| Crella sp. | Norsel Point, Amsler Island (Antarctica) | norselic acid A (97), B (98), C (99), D (100), E (101) | oxidized steroids | Norselic acid A: activity against MRSA, methicillin-sensitive S. aureus (MSSA) and vancomycin-resistant Enterococci faecium (VREF) and C. albicans. All norselic acids were active against leishmaniasis | [122] |
| Dendrilla antarctica | Anvers Island (Antarctica) | aplysulphurin (55), membranoid A (56), B (57), C (58), D (59), E (60), G (61),H (62) | oxidized diterpenoids | Aplysulphurin: activity against C. albicans, and Gram-negative antibiotic activity against S. aureus and E. coli; membranoids: activity against the leishmaniasis | [123,124] |
| darwinolide (63), tetrahydroaplysulphurin-1 (65), membranolide (66), glaciolides (67–68), cadlinolide C (69), dendrillin A (70), B (71), C (72), D (73) and semisynthetic derivatives (74–76) | oxidized diterpenoids | Darwinolide: selectivity against the biofilm phase of MRSA compared to the planktonic phase; membranolide: activity against MRSA; dendrillin B: activity against L. donovani; 76, activity against Plasmodium falciparum | [125,126] | ||
| 9-11-dihydrogracilin A (64) | oxidized diterpenoid | Immuno-modulatory and anti-inflammatory activity in human cell lines | [127] | ||
| Geodia baretti | North Sea off the coast of Sweden and the northern coast of Iceland (Arctic Ocean) | barettin and the geobarrettins | diketopiperazine (likely produced by a symbiont) | Moderate antioxidant and anti-inflammatory activities | [53] |
| [128] | |||||
| [129] | |||||
| Haliclona viscosa | Svalbard Archipelago (Arctic Ocean) | viscosamine | 3-alkyl pyridinium alkaloids | Antibiotic activity against four separate sympatric bacterial strains | [130] |
| viscosaline | [131] | ||||
| Kirkpatrickia variolosa | Antarctica | variolins A-D (B most active) | pyridopyrrolopyrimidine | Cytotoxic activity against P388 murine leukemia cell line | [132] |
| [133] | |||||
| Latrunculia sp. | Aleutian Islands, Alaska (also found in Antarctic specimens) | discorhabdins A, C, R, dihydrodiscorhabdin B | spirocyclic imino-quinones | Anti-HCV (Hepatitis C virus) activity, antimalarial activity and selective antimicrobial activity against MRSA, Mycobacterium intracellulare and M. tuberculosis. | [134] |
| Antiprotozoal activity in vitro (P. falciparum) | |||||
| Weddell Sea (Antarctica) | tsitsikammamines | pyrroloiminoquinones | Anticancer and cytotoxic activities | [57] | |
| Lyssodendoryx flabellata | Terra Nova Bay (Antarctica) | terpioside | glycosphingolipid | Inhibition effect in mixed lymphocyte reactions on human cells | [135] |
| Mycale acerata | Terra Nova Bay (Antarctica) | mycalol | alkyl glyceryl ether lipid | Activity against human thyroid carcinoma cells | [136] |
| Plakortis simplex | Sula Ridge Reef, Norwegian Shelf (sub Arctic) | methyl 2-((3R,6S)-4,6-diethyl-6-hexyl-3,6-dihydro-1,2-dioxin- 3-yl)acetate | cyclic peroxide (fatty acids) | Selectively inhibited proliferation in gastric cancer (GXF 251L), non-small cell lung cancer (LXFL 529L) and melanoma (MEXF 462NL) cell lines. | [137] |
| Polymastia boletiformis | Norwegian coast, Western Irish Coast (sub Arctic) | polymastiamide A (117), B, C, D, E, F | sulfated steroid-amino acid conjugates | Polymastiamide A: antifungal activity against plant pathogens Cladosporium cucumerinum and Pythium ultimum and human yeast pathogen C. albicans | [138] |
| Polymastiamide A: antibacterial activity against S. aureus | [139] | ||||
| [140] | |||||
| Stryphus fortis | Spitsbergen, Svalbard, (Arctic Ocean) | ianthelline | bromotyrosine derivative | Antitumor properties against several malignant cell lines and inhibition of PK activity | [141] |
| Suberites sp. | King George Isalnd, McMurdo Sound (Antarctica) | suberitenones A (77), B (78) | oxidized sesterterpenoids | Inhibition of the cholesteryl ester transfer protein (CETP) | [142] |
| TUNICATES | |||||
| Plylum Chordata | |||||
| Class Ascidiacea | |||||
| Aplidium meridianum | South Georgia Islands (Antarctica) | meridianins | brominated 3-(2-aminopyrimidine)indoles | Prevention of cell proliferation and induction of cell apoptosis. Inhibition of CDKs, GSK-3, PKA and other kinases in the low micromolar range | [143] |
| Aplidium sp. | Ross Sea (Antarctica) | rossinones A (122), B (123) | meroterpenoids | Antiproliferative activity against several cell lines. Antiviral activity against the DNA virus HSV-1 as well as antibacterial and antifungal activity against B. subtilis and Trichophyton mentagrophytes | [144] |
| Clavelina lepadiformis | Bergen, Norway | lepadins | decahydroquinoline alkaloid | Lepadin A: anti-cancer activity against leukemia P388, breast cancer (MCF7), glioblastoma/astrocytoma (U373), ovarian (HEY), colon (LoVo) and lung (A549) | [145] |
| [146] | |||||
| Synoicum adareanum | Anvers Island (Antarctica) | palmerolides A–G | enamide-bearing macrolides | Palmerolide A: Activity against melanoma (UACC-62 LC50), by inhibition of vacuolar ATPase- | [147] |
| [148] | |||||
| hyousterones A (107), B, C (108), D, abeohyousterone (109) | ecdysteroids | Activity against colon cancer cells | [149] | ||
| Synoicum pulmonaria | Tromso, Northern Norway (Arctic Ocean) | synoxazolidinones A-C | brominated guanidinium oxazolidinones | Antibacterial activity against MSSA, MRSA and Corynebacterium glutamicum as well as antifungal properties against Saccharomyces cerevisiae. Active against human melanoma (A2058), breast adenocarcinoma (MCF-7) and colon carcinoma (HT-29) cell line, with noted cytoxicity | [150] |
| [151] | |||||
| pulmonarins A, B | brominated methoxybenzoylesters bearing quaternary ammonium mioeties | Acetylcholinesterase inhibitory activity and weak antibacterial activity against C. glutamicum | [152] | ||
| MACROALGAE | |||||
| Plylum Rhodophyta | |||||
| Class Florideophyceae | |||||
| Delisea fimbriata and pulchra | Anvers Island (Antarctica) | fimbrolides and analogues | polyhalogenated furanones | Antimicrobial activity against S. aureus, E. coli, C. albicans and Streptococcus sp. | [153] |
| Plocamium cartilagineum | Anvers Island (Antarctica) | oregonene (1) and similar compounds (2–4), anverenes A (5), B (6), C (7), D (8), E (9) | polyhalogenated monoterpenes | Cytotoxic activity against cervical cancer cells | [154] |
| Plylum Ochrophyta | |||||
| Class Phaeophyceae | |||||
| Desmarestia menziesii | Anvers Island (Antarctica) | Menzoquinone (124) | terpenoid-quinone | Antimicrobial activity against MRSA, MSSA, VREF | [155] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Núñez-Pons, L.; Shilling, A.; Verde, C.; Baker, B.J.; Giordano, D. Marine Terpenoids from Polar Latitudes and Their Potential Applications in Biotechnology. Mar. Drugs 2020, 18, 401. https://doi.org/10.3390/md18080401
Núñez-Pons L, Shilling A, Verde C, Baker BJ, Giordano D. Marine Terpenoids from Polar Latitudes and Their Potential Applications in Biotechnology. Marine Drugs. 2020; 18(8):401. https://doi.org/10.3390/md18080401
Chicago/Turabian StyleNúñez-Pons, Laura, Andrew Shilling, Cinzia Verde, Bill J. Baker, and Daniela Giordano. 2020. "Marine Terpenoids from Polar Latitudes and Their Potential Applications in Biotechnology" Marine Drugs 18, no. 8: 401. https://doi.org/10.3390/md18080401
APA StyleNúñez-Pons, L., Shilling, A., Verde, C., Baker, B. J., & Giordano, D. (2020). Marine Terpenoids from Polar Latitudes and Their Potential Applications in Biotechnology. Marine Drugs, 18(8), 401. https://doi.org/10.3390/md18080401