Molecular Targets and Related Biologic Activities of Fucoidan: A Review


Abstract
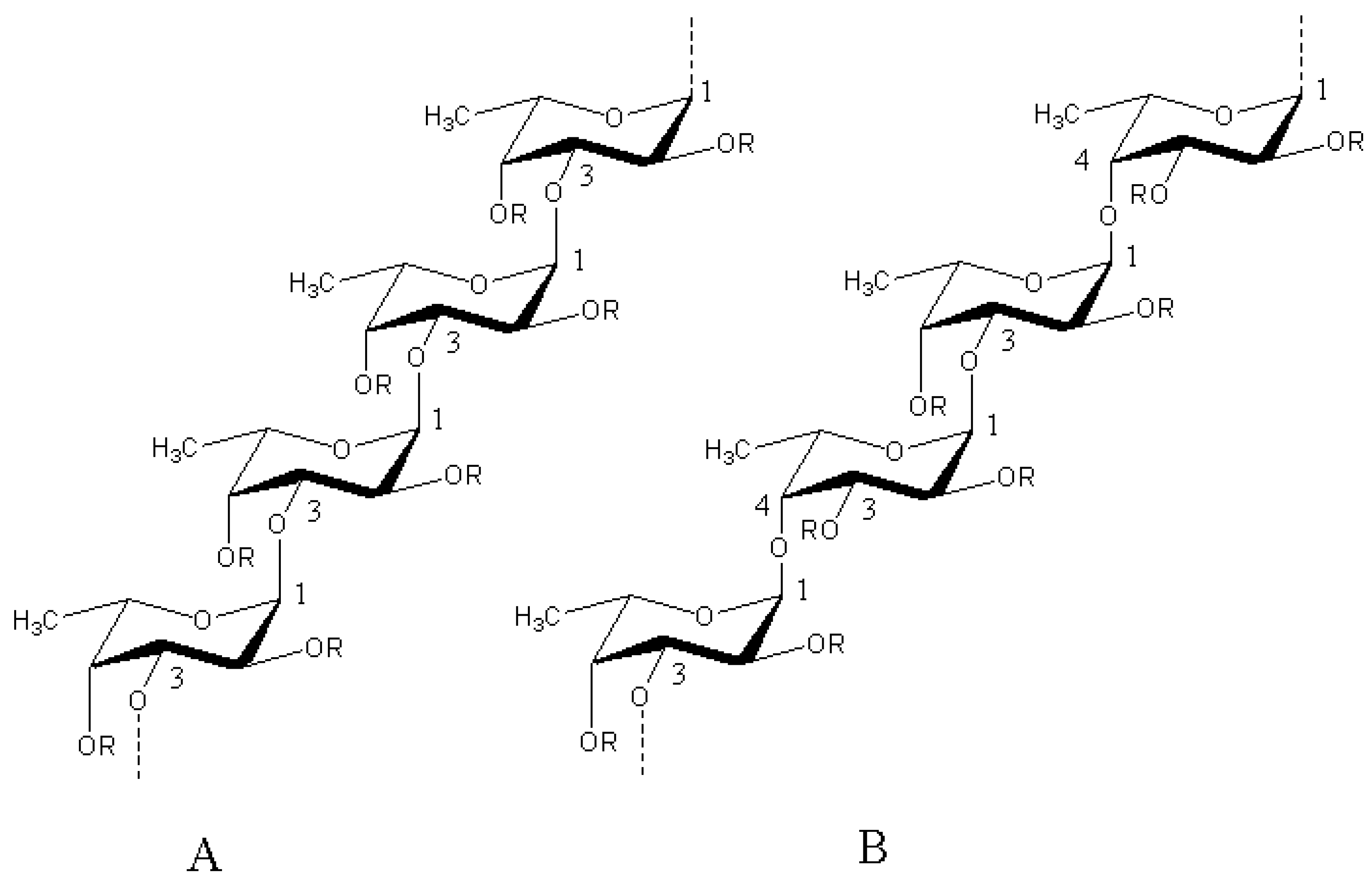
1. Introduction
2. Molecular Targets of Fucoidan
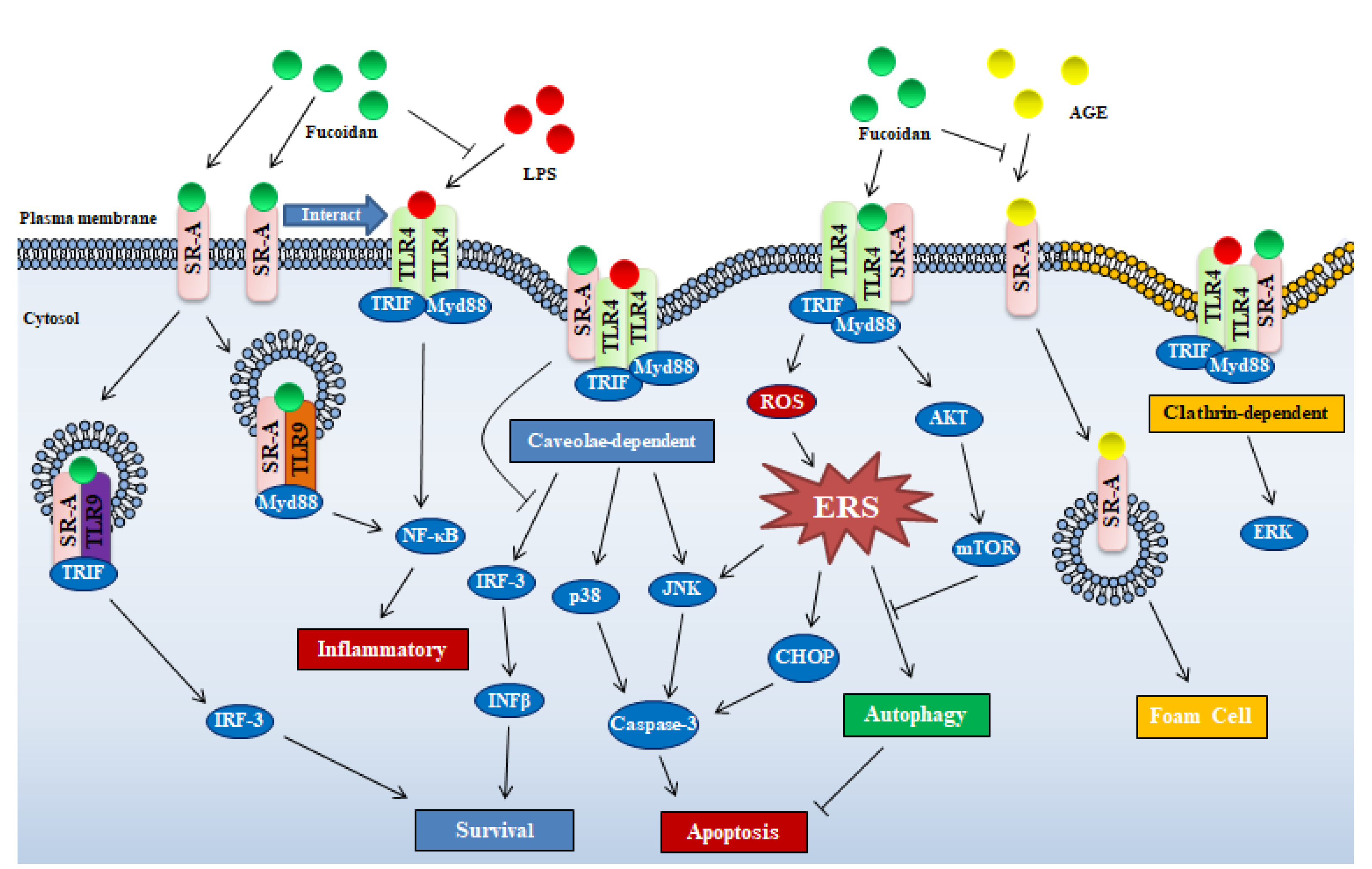
2.1. Class A Scavenger Receptor
2.1.1. Fucoidan/SR-A Involved Biological Functions and Related Molecular Mechanisms
Cooperation between SR-A and Other Receptors
SR-A Mediated Internalization of Ligands
SR-A Involved Endoplasmic Reticulum Stress Pathway
2.2. Toll-Like Receptors
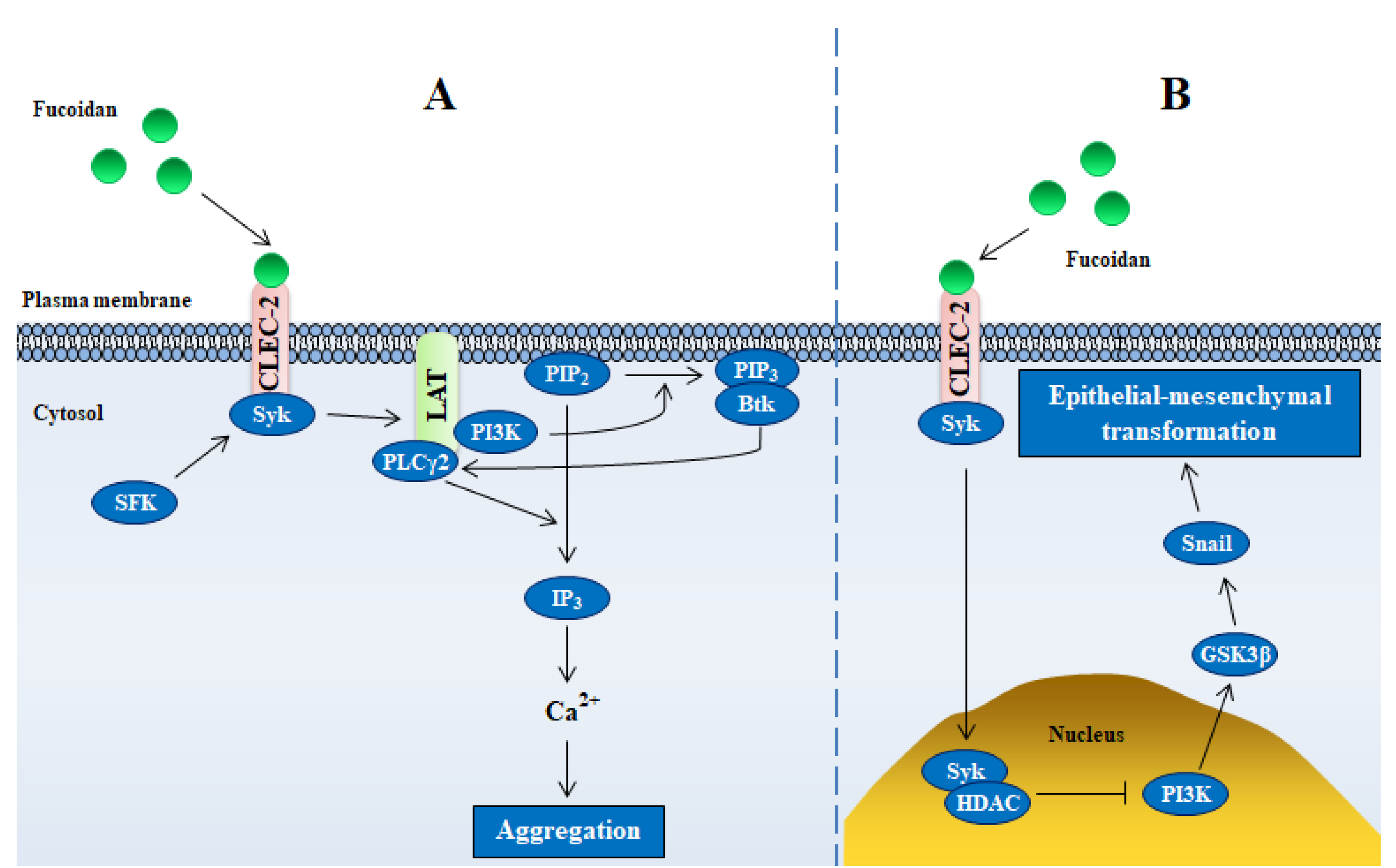
2.3. C-Type Lectins (CLEC)
2.3.1. C-Type Lectin Receptors (CLRs)
2.3.2. CLEC-2
2.3.3. Selectins
2.4. Other Affinity Molecules of Fucoidan
2.4.1. Integrins: CR3 and αVβ3
2.4.2. VEGF
2.4.3. CXCL12/CXCR4
2.4.4. Elastin Peptide Receptor
2.4.5. TGF-β1
2.4.6. ECM Proteins
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Xie, L.; Shen, M.; Hong, Y.; Ye, H.; Huang, L.; Xie, J. Chemical modifications of polysaccharides and their anti-tumor activities. Carbohydr. Polym. 2020, 229, 115436. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.H.; Zhu, L.; Jiang, J.G. Immunoregulatory actions of polysaccharides from Chinese herbal medicine. Expert Opin. Ther. Targets 2010, 14, 1367–1402. [Google Scholar] [CrossRef]
- Kim, Y.S.; Ryu, J.H.; Han, S.J.; Choi, K.H.; Nam, K.B.; Jang, I.H.; Lemaitre, B.; Brey, P.T.; Lee, W.J. Gram-negative bacteria-binding protein, a pattern recognition receptor for lipopolysaccharide and beta-1,3-glucan that mediates the signaling for the induction of innate immune genes in drosophila melanogaster cells. J. Biol. Chem. 2000, 275, 32721–32727. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Lan, Y.; Liu, J.; Zhang, F.; Zhang, L.; Li, B.; Zhao, X. The structure property and endothelial protective activity of fucoidan from laminaria japonica. Int. J. Biol. Macromol. 2017, 105, 1421–1429. [Google Scholar] [CrossRef] [PubMed]
- Narayani, S.S.; Saravanan, S.; Ravindran, J.; Ramasamy, M.S.; Chitra, J. In vitroanticancer activity of fucoidan extracted from Sargassum cinereum against Caco-2 cells. Int. J. Biol. Macromol. 2019, 138, 618–628. [Google Scholar] [CrossRef]
- Oliveira, C.; Granja, S.; Neves, N.M.; Reis, R.L.; Baltazar, F.; Silva, T.H.; Martins, A. Fucoidan from Fucus vesiculosus inhibits new blood vessel formation and breast tumor growth in vivo. Carbohydr. Polym. 2019, 223, 115034. [Google Scholar] [CrossRef]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef]
- Holtkamp, A.D.; Kelly, S.; Ulber, R.; Lang, S. Fucoidans and fucoidanases—Focus on techniques for molecular structure elucidation and modification of marine polysaccharides. Appl. Microbiol. Biotechnol. 2009, 82, 1–11. [Google Scholar] [CrossRef]
- Goldstein, J.L.; Ho, Y.K.; Basu, S.K.; Brown, M.S. Binding site on macrophages that mediates uptake and degradation of acetylated low density lipoprotein, producing massive cholesterol deposition. Proc. Natl. Acad. Sci. USA 1979, 76, 333–337. [Google Scholar] [CrossRef]
- Kelley, J.L.; Ozment, T.R.; Li, C.; Schweitzer, J.B.; Williams, D.L. Scavenger receptor-a (CD204): A two-edged sword in health and disease. Crit. Rev. Immunol. 2014, 34, 241–261. [Google Scholar] [CrossRef]
- Zhu, X.D.; Zhuang, Y.; Ben, J.J.; Qian, L.L.; Huang, H.P.; Bai, H.; Sha, J.H.; He, Z.G.; Chen, Q. Caveolae-dependent endocytosis is required for class A macrophage scavenger receptor-mediated apoptosis in macrophages. J. Biol. Chem. 2011, 286, 8231–8239. [Google Scholar] [CrossRef] [PubMed]
- Okamura, D.M.; Lopez-Guisa, J.M.; Koelsch, K.; Collins, S.; Eddy, A.A. Atherogenic scavenger receptor modulation in the tubulointerstitium in response to chronic renal injury. Am. J. Physiol. Ren. Physiol. 2007, 293, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Vitorino Carvalho, A.; Eozenou, C.; Richard, C.; Forde, N.; Healey, G.D.; Giraud-Delville, C.; Mansouri-Attia, N.; Lonergan, P.; Sheldon, I.M.; Sandra, O. Bovine scavenger receptor class A (SR-A) exhibit specific patterns of regulation in the endometrium during the oestrous cycle and early pregnancy. Reprod. Fertil. Dev. 2019, 3, 1078–1090. [Google Scholar] [CrossRef]
- Yu, H.; Ha, T.; Liu, L.; Wang, X.; Gao, M.; Kelley, J.; Kao, R.; Williams, D.; Li, C. Scavenger receptor A (SR-A) is required for LPS-induced TLR4 mediated NF-κB activation in macrophages. Biochim. Biophys. Acta 2012, 1823, 1192–1198. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.Y.; Chiu, S.L.; Wen, M.H.; Chen, K.Y.; Hua, K.F. Ligands of macrophage scavenger receptor induce cytokine expression via differential modulation of protein kinase signaling pathways. J. Biol. Chem. 2001, 276, 28719–28730. [Google Scholar] [CrossRef]
- Hsu, H.Y.; Hajjar, D.P.; Khan, K.M.F.; Falcone, D.J. Ligand binding to macrophage scavenger receptor-a induces urokinase-type plasminogen activator expression by a protein kinase-dependent signaling pathway. J. Biol. Chem. 1998, 273, 1240–1246. [Google Scholar] [CrossRef]
- Nakamura, T.; Suzuki, H.; Wada, Y.; Kodama, T.; Doi, T. Fucoidan induces nitric oxide production via p38 mitogen-activated protein kinase and NF-κB-dependent signaling pathways through macrophage scavenger receptors. Biochem. Biophys. Res. Commun. 2006, 343, 286–294. [Google Scholar] [CrossRef]
- Seimon, T.A.; Obstfeld, A.; Moore, K.J.; Golenbock, D.T.; Tabas, I. Combinatorial pattern recognition receptor signaling alters the balance of life and death in macrophages. Proc. Natl. Acad. Sci. USA 2006, 103, 19794–19799. [Google Scholar] [CrossRef]
- Makarenkova, I.D.; Logunov, D.Y.; Tukhvatulin, A.I.; Semenova, I.B.; Besednova, N.N.; Zvyagintseva, T.N. Interactions between sulfated polysaccharides from sea brown algae and Toll-like receptors on HEK293 eukaryotic cellsin vitro. Bull. Exp. Biol. Med. 2012, 154, 241–244. [Google Scholar] [CrossRef]
- Pei, Z.; Pang, H.; Qian, L.; Yang, S.; Zhang, W.; Wu, X.; Dallas, S.; Wilson, B.; Reece, J.M.; Miller, D.S.; et al. Mac1 mediates LPS-induced production of superoxide by microglia: The role of pattern recognition receptors in dopaminergic neurotoxicity. Glia 2007, 55, 1362–1373. [Google Scholar] [CrossRef]
- Szabó, R.; Mezö, G.; Pállinger, E.; Kovács, P.; Köhidai, L.; Bösze, S.; Hudecz, F. In vitro cytotoxicity, chemotactic effect, and cellular uptake of branched polypeptides with poly(L-lys) backbone by J774 murine macrophage cell line. Bioconjugate Chem. 2008, 19, 1078–1086. [Google Scholar] [CrossRef] [PubMed]
- Hamasaki, S.; Kobori, T.; Yamazaki, Y.; Kitaura, A.; Niwa, A.; Nishinaka, T.; Nishibori, M.; Mori, S.; Nakao, S.; Takahashi, H. Effects of scavenger receptors-1 class a stimulation on macrophage morphology and highly modified advanced glycation end product-protein phagocytosis. Sci. Rep. 2018, 8, 5901. [Google Scholar] [CrossRef] [PubMed]
- Ben, J.; Zhang, Y.; Zhou, R.; Zhang, H.; Zhu, X.; Li, X.; Zhang, H.; Li, N.; Zhou, X.; Bai, H.; et al. Major vault protein regulates class A scavenger receptor-mediated TNF-alpha synthesis and apoptosis in macrophages. J. Biol. Chem. 2013, 288, 20076–20084. [Google Scholar] [CrossRef]
- Huang, H.; Li, X.; Zhuang, Y.; Li, N.; Chen, Q. Class a scavenger receptor activation inhibits endoplasmic reticulum stress-induced autophagy in macrophage. J. Biomed. Res. 2014, 28, 213–221. [Google Scholar] [PubMed]
- Patin, E.C.; Thompson, A.; Orr, S.J. Pattern recognition receptors in fungal immunity. Semin. Cell Dev. Biol. 2018, 89, 24–33. [Google Scholar] [CrossRef]
- Ji, C.F.; Chen, J.R.; Zhang, Z.Y. Research progress of receptors of plant polysaccharides. Chin. Pharm. J. 2019, 54, 1766–1772. [Google Scholar]
- Teruya, T.; Tatemoto, H.; Konishi, T.; Tako, M. Structural characteristics andin vitromacrophage activation of acetyl fucoidan fromcladosiphon okamuranus. Glycoconj. J. 2009, 26, 1019–1028. [Google Scholar] [CrossRef]
- Hsu, H.Y.; Lin, T.Y.; Lu, M.K.; Leng, P.J.; Tsao, S.M.; Wu, Y.C. Fucoidan induces Toll-like receptor 4-regulated reactive oxygen species and promotes endoplasmic reticulum stress-mediated apoptosis in lung cancer. Sci. Rep. 2017, 7, 44990. [Google Scholar] [CrossRef]
- Hsu, H.Y.; Lin, T.Y.; Hu, C.H.; Shu, D.T.F.; Lu, M.K. Fucoidan upregulates TLR4/CHOP-mediated caspase-3 and parp activation to enhance cisplatin-induced cytotoxicity in human lung cancer cells. Cancer Lett. 2018, 432, 112–120. [Google Scholar] [CrossRef]
- Park, H.Y.; Han, M.H.; Park, C.; Jin, C.Y.; Kim, G.Y.; Choi, I.W.; Kim, N.D.; Nam, T.J.; Kwon, T.K.; Choi, Y.H. Anti-inflammatory effects of fucoidan through inhibition of NF-κB, MAPKand AKTactivation in lipopolysaccharide-induced BV2 microglia cells. Food Chem. Toxicol. 2011, 49, 1745–1752. [Google Scholar] [CrossRef]
- Yi, H.; Yu, X.; Gao, P.; Wang, Y.; Baek, S.H.; Chen, X.; Kim, H.L.; Subjeck, J.R.; Wang, X.Y. Pattern recognition scavenger receptor SRA/CD204 down-regulates Toll-like receptor 4 signaling-dependent CD8 T-cell activation. Blood 2009, 113, 5819–5828. [Google Scholar] [CrossRef] [PubMed]
- Brennan, J.; Takei, F.; Wong, S.; Mager, D.L. Carbohydrate recognition by a natural killer cell receptor, Ly-49C. J. Biol. Chem. 1995, 270, 9691–9694. [Google Scholar] [CrossRef] [PubMed]
- Daniels, B.F.; Nakamura, M.C.; Rosen, S.D.; Yokoyama, W.M.; Seaman, W.E. Ly-49A, a receptor for H-2Dd, has a functional carbohydrate recognition domain. Immunity 1994, 1, 785–792. [Google Scholar] [CrossRef]
- Gange, C.T.; Quinn, J.M.W.; Zhou, H.; Kartsogiannis, V.; Gillespie, M.T.; Ng, K.W. Characterization of sugar binding by osteoclast inhibitory lectin. J. Biol. Chem. 2004, 279, 29043–29049. [Google Scholar] [CrossRef]
- Xu, L.; Liu, F.; Li, C.; Li, S.; Wu, H.; Guo, B.; Gu, J.; Wang, L. Fucoidan suppresses the gastric cancer cell malignant phenotype and production of TGF-β1 via CLEC-2. Glycobiology 2019, 30, 301–311. [Google Scholar] [CrossRef]
- Alshehri, O.M.; Montague, S.; Watson, S.; Carter, P.; Watson, S.P. Activation of Glycoprotein VI (GPVI) and C-type Lectin-like receptor-2 (CLEC-2) underlies platelet activation by diesel exhaust particles and other charged/hydrophobic ligands. Biochem. J. 2015, 468, 459–473. [Google Scholar] [CrossRef]
- Manne, B.K.; Getz, T.M.; Hughes, C.E.; Alshehri, O.; Dangelmaier, C.; Naik, U.P.; Watson, S.P.; Kunapuli, S.P. Fucoidan is a novel platelet agonist for the C-type Lectin-like receptor 2 (CLEC-2). J. Biol. Chem. 2013, 288, 7717–7726. [Google Scholar] [CrossRef]
- Martyanov, A.A.; Balabin, F.A.; Maiorov, A.S.; Shamova, E.V.; Sveshnikova, A.N. Mathematical model of platelet intracellular signaling after activation by fucoidan. Biochem. Mosc. Suppl. Ser. A Membr. Cell Biol. 2018, 12, 333–343. [Google Scholar] [CrossRef]
- Wang, L.; Yin, J.; Wang, X.; Shao, M.; Duan, F.; Wu, W.; Peng, P.; Jin, J.; Tang, Y.; Ruan, Y.; et al. C-Type Lectin-like receptor 2 suppresses AKT signaling and invasive activities of gastric cancer cells by blocking expression of phosphoinositide 3-kinase subunits. Gastroenterology 2016, 150, 1183–1195. [Google Scholar] [CrossRef]
- Borsig, L. Selectins in cancer immunity. Glycobiology 2018, 28, 648–655. [Google Scholar] [CrossRef]
- Miura, T.; Nelson, D.P.; Schermerhorn, M.L.; Shin’oka, T.; Zund, G.; Hickey, P.R.; Neufeld, E.J.; Mayer, J.E. Blockade of selectin-mediated leukocyte adhesion improves postischemic function in lamb hearts. Ann. Thorac. Surg. 1996, 62, 1295–1300. [Google Scholar] [CrossRef]
- Sitrin, R.G.; Pan, P.M.; Blackwood, R.A.; Huang, J.; Petty, H.R. Cutting edge: Evidence for a signaling partnership between urokinase receptors (CD87) and L-selectin (CD62L) in human polymorphonuclear neutrophils. J. Immunol. 2001, 166, 4822–4825. [Google Scholar] [CrossRef]
- Barrabés, J.A.; Garcia-Dorado, D.; Mirabet, M.; Inserte, J.; Agulló, L.; Soriano, B.; Massaguer, A.; Padilla, F.; Lidón, R.M.; Soler-Soler, J. Antagonism of selectin function attenuates microvascular platelet deposition and platelet-mediated myocardial injury after transient ischemia. J. Am. Coll. Cardiol. 2005, 45, 293–299. [Google Scholar] [CrossRef]
- Bachelet, L.; Bertholon, I.; Lavigne, D.; Vassy, R.; Jandrot-Perrus, M.; Chaubet, F.; Letourneur, D. Affinity of low molecular weight fucoidan for P-selectin triggers its binding to activated human platelets. Biochim. Biophys. Acta (BBA) Gen. Subj. 2009, 1790, 141–146. [Google Scholar] [CrossRef]
- Rouzet, F.; Bachelet-Violette, L.; Alsac, J.M.; Suzuki, M.; Meulemans, A.; Louedec, L.; Petiet, A.; Jandrot-Perrus, M.; Chaubet, F.; Michel, J.B.; et al. Radiolabeled Fucoidan as a P-Selectin Targeting Agent for In Vivo Imaging of Platelet-Rich Thrombus and Endothelial Activation. J. Nucl. Med. 2011, 52, 1433–1440. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, X.M.; Heflin, K.E.; Lavigne, L.M.; Yu, K.; Kim, M.; Salomon, A.R.; Reichner, J.S. Lectin site ligation of CR3 induces conformational changes and signaling. J. Biol. Chem. 2012, 287, 3337–3348. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Li, C.; Surayot, U.; Yelithao, K.; Lee, S.; Park, W.; Tabarsa, M.; You, S. Molecular structures, chemical properties and biological activities of polysaccharide from Smilax glabra rhizome. Int. J. Biol. Macromol. 2018, 120, 1726–1733. [Google Scholar] [CrossRef]
- Yelithao, K.; Surayot, U.; Park, W.; Lee, S.; Lee, D.H.; You, S. Effect of sulfation and partial hydrolysis of polysaccharides from Polygonatum sibiricum on immune-enhancement. Int. J. Biol. Macromol. 2019, 122, 10–18. [Google Scholar] [CrossRef]
- Zen, K.; Liu, Y.; Cairo, D.; Parkos, C.A. CD11b/CD18-dependent interactions of neutrophils with intestinal epithelium are mediated by fucosylated proteoglycans. J. Immunol. 2002, 169, 5270–5278. [Google Scholar] [CrossRef]
- Pan, T.J.; Li, L.X.; Zhang, J.W.; Yang, Z.S.; Shi, D.M.; Yang, Y.K.; Wu, W.Z. Antimetastatic effect of fucoidan-sargassum against liver cancer cell invadopodia formation via targeting integrin αVβ3 and mediating αVβ3/Src/E2F1 signaling. J. Cancer 2019, 10, 4777–4792. [Google Scholar] [CrossRef]
- Chen, H.; Cong, Q.; Du, Z.; Liao, W.; Zhang, L.; Yao, Y.; Ding, K. Sulfated fucoidan FP08S2 inhibits lung cancer cell growth in vivo by disrupting angiogenesis via targeting VEGFR2/VEGF and blocking VEGFR2/Erk/VEGF signaling. Cancer Lett. 2016, 382, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Cui, N.; Bo, Z.; Xiang, F. Structural determinant and its underlying molecular mechanism of STPC2 related to anti-angiogenic activity. Mar. Drugs 2017, 15, 48. [Google Scholar] [CrossRef] [PubMed]
- Schneider, T.; Ehrig, K.; Liewert, I.; Alban, S. Interference with the CXCL12/CXCR4 axis as potential antitumor strategy: Superiority of a sulfated galactofucan from the brown alga Saccharina latissima and Fucoidan over heparins. Glycobiology 2015, 25, 812–824. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Nishiura, H.; Tokita, K.; Kouike, Y.; Taniguchi, C.; Iwahara, M.; Nishino, N.; Hama, Y.; Asakawa, M.; Yamamoto, T. Elastin peptide receptor-directed monocyte chemotactic polysaccharides derived from seaweed sporophyll and from infectious fungus. Microb. Pathog. 2008, 45, 423–434. [Google Scholar] [CrossRef]
- Larbi, A.; Levesque, G.; Robert, L.; Gagné, D.; Douziech, N.; Fülöp, T. Presence and active synthesis of the 67 kDa elastin-receptor in human circulating white blood cells. Biochem. Biophys. Res. Commun. 2005, 332, 787–792. [Google Scholar] [CrossRef]
- McCaffrey, T.A.; Falcone, D.J.; Vicente, D.; Du, B.; Consigli, S.; Borth, W. Protection of transforming growth factor-b1 activity by heparin and fucoidan. J. Cell. Physiol. 1994, 159, 51–59. [Google Scholar] [CrossRef]
- Kim, T.H.; Lee, E.K.; Lee, M.J.; Kim, J.H.; Yang, W.S. Fucoidan inhibits activation and receptor binding of transforming growth factor-β1. Biochem. Biophys. Res. Commun. 2013, 432, 163–168. [Google Scholar] [CrossRef]
- Rocha, H.A.; Franco, C.R.; Trindade, E.S.; Veiga, S.S.; Leite, E.L.; Nader, H.B.; Dietrich, C.P. Fucan inhibits Chinese hamster ovary cell (CHO) adhesion to fibronectin by binding to the extracellular matrix. Planta Med. 2005, 71, 628–633. [Google Scholar] [CrossRef]
- Huang, Y.L.; Pai, F.S.; Tsou, Y.T.; Mon, H.C.; Hsu, T.L.; Wu, C.Y.; Chou, T.Y.; Yang, W.B.; Chen, C.H.; Wong, C.H.; et al. Human CLEC18 gene cluster contains C-type lectins with differential glycan-binding specificity. J. Biol. Chem. 2015, 290, 21252–21263. [Google Scholar] [CrossRef]
- Takano, T.; Motozono, C.; Imai, T.; Sonoda, K.H.; Nakanishi, Y.; Yamasaki, S. Dectin-1 intracellular domain determines species-specific ligand spectrum by modulating receptor sensitivity. J. Biol. Chem. 2017, 292, 16933–16941. [Google Scholar] [CrossRef]
- Surayot, U.; Lee, S.; You, S. Effects of sulfated fucan from the sea cucumber Stichopus japonicus on natural killer cell activation and cytotoxicity. Int. J. Biol. Macromol. 2018, 108, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Geng, L.; Yue, Y.; Zhang, Q. Use of fucoidan to treat renal diseases: A review of 15 years of clinic studies. Prog. Mol. Biol. Transl. Sci. 2019, 163, 95–111. [Google Scholar] [PubMed]
- Finlay, D.B.; Duffull, S.B.; Glass, M. 100 years of modelling ligand-receptor binding and response: A focus on GPCRs. Br. J. Pharmacol. 2020, 177, 1472–1484. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Molecular Targets | Cell Types | Biologic Activity of Interaction with Fucoidan | Reference |
|---|---|---|---|
| Scavenger receptors | |||
| SR-A | Macrophages, endothelial cells, lung epithelial cells, microglia, astrocytes, primary murine fibroblasts |
| [10,11,12,13,14,15,16,17,18,19,20,21,22,23,24] |
| Toll-like receptors | |||
| TLR2 | Macrophages, monocytes, dendritic cells, mast cells, neutrophils, natural killer (NK) cells fibroblasts, embryonic kidney cells | Activating the nuclear factor-κB (NF-κB) | [19,25,26] |
| TLR4 | Macrophages, monocytes, dendritic cells mast cells, neutrophils, b lymphocytes, intestinal epithelium cells, cardiomyocytes, renal tubular epithelial and endothelial cells, podocytes, Kupffer cells, lung cancer cells |
| [18,19,25,26,27,28,29,30,31] |
| C-type lectins | |||
| Ly-49 | NK cells, subset of T lymphocytes | Blocking cell adhesion | [32,33] |
| OCIL | Macrophages, epithelial cells, mesenchymal cells, dendritic cells, lymphocytes | Cannot affect osteoclast inhibitory lectin (OCIL) inhibition of osteoclast formation | [34] |
| CLEC-2 | Platelets, (low levels in immune cells), gastric epithelial cells |
| [35,36,37,38,39] |
| L-selectin | Monocytes, dendritic cells, neutrophils, NK cells, B lymphocytes, T lymphocytes |
| [40,41,42] |
| P-selectin | Platelets, endothelial cell, Kupffer cells | Attenuating microvascular platelet deposition and platelet-mediated myocardial injury | [40,41,43,44,45] |
| Other affinity molecules of fucoidan | |||
| CR3 | Macrophages, monocytes, neutrophils, NK cells | Inhibiting adhesion of T84 cells to complement receptor 3 (CR3) | [46,47,48,49] |
| αVβ3 | Cancer cells, rapidly dividing endothelial cells | Suppressing metastasis of gastric cancer cells though Src/cortactin/ E2F transcription factor 1 (E2F1) signaling pathway | [50] |
| VEGF | N/A | Impeding vascular endothelial growth factors (VEGF)–VEGF receptor 2 (VEGFR2) interaction | [51,52] |
| CXCL12/CXCR4 | N/A | Blocking both C–X–C motif chemokine 12 (CXCL12) induced C–X–C motif chemokine receptor type 4 (CXCR4) activation | [53] |
| Elastin peptide receptor | Mesenchymal cells, vascular smooth muscle cells, skin fibroblasts | Recruiting monocytes through binding to elastin peptide receptor of monocytes | [54,55] |
| TGF-β1 | N/A | Exerting functions of anti-fibrosis through blocking transforming growth factor-β (TGF-β1) interacted with its receptor | [56,57] |
| ECM proteins | N/A | Suppressing metastasis of cancer cells though blocking extracellular matrix (ECM) proteins | [58] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Z.; Tan, X.; Zhang, Y.; Li, F.; Luo, P.; Liu, H. Molecular Targets and Related Biologic Activities of Fucoidan: A Review. Mar. Drugs 2020, 18, 376. https://doi.org/10.3390/md18080376
Lin Z, Tan X, Zhang Y, Li F, Luo P, Liu H. Molecular Targets and Related Biologic Activities of Fucoidan: A Review. Marine Drugs. 2020; 18(8):376. https://doi.org/10.3390/md18080376
Chicago/Turabian StyleLin, Zhen, Xiaohui Tan, Yu Zhang, Fangping Li, Ping Luo, and Huazhong Liu. 2020. "Molecular Targets and Related Biologic Activities of Fucoidan: A Review" Marine Drugs 18, no. 8: 376. https://doi.org/10.3390/md18080376
APA StyleLin, Z., Tan, X., Zhang, Y., Li, F., Luo, P., & Liu, H. (2020). Molecular Targets and Related Biologic Activities of Fucoidan: A Review. Marine Drugs, 18(8), 376. https://doi.org/10.3390/md18080376