Sortase A-Inhibitory Metabolites from a Marine-Derived Fungus Aspergillus sp.

 and
and
Abstract
1. Introduction
2. Results and Discussion
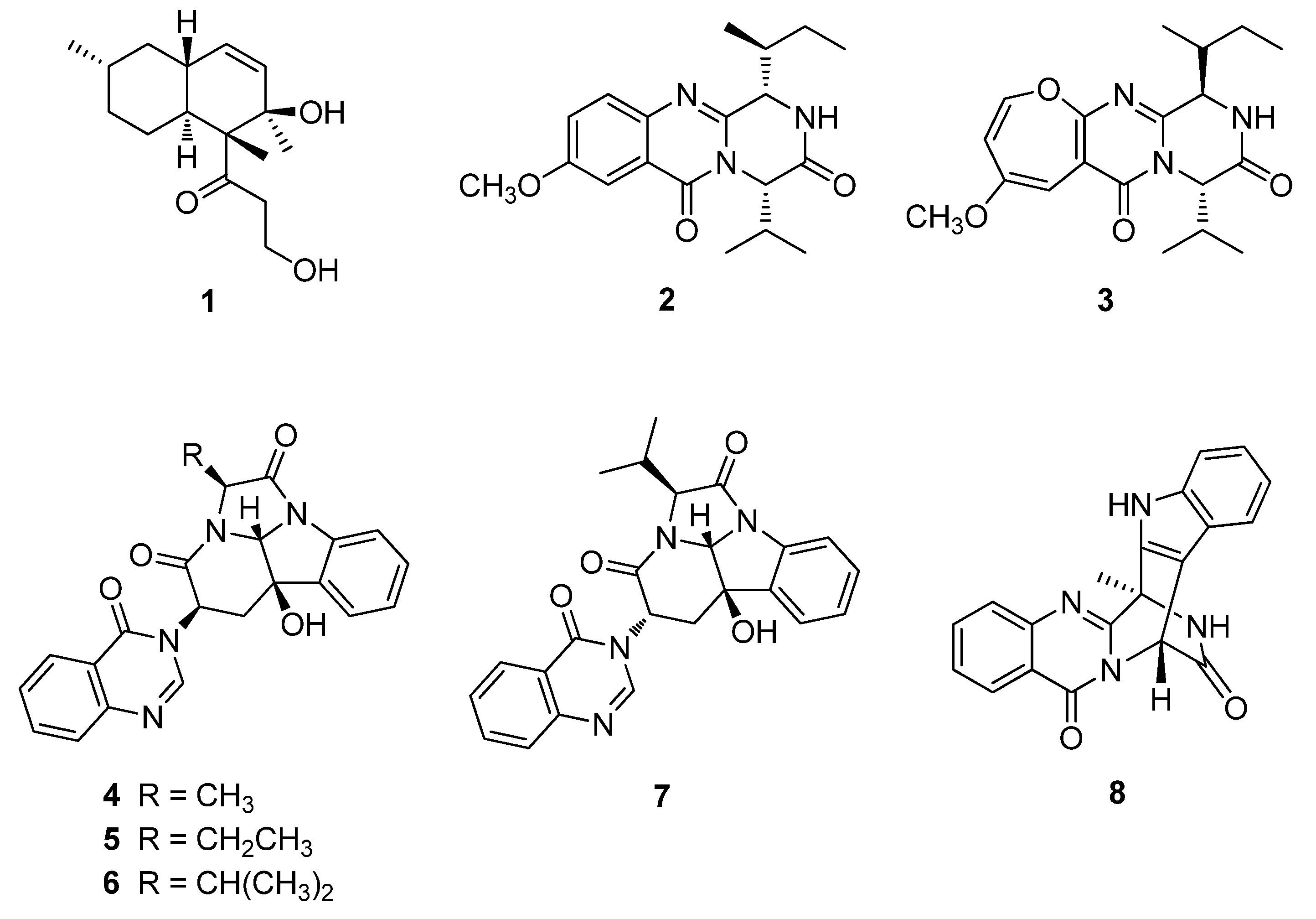
2.1. Isolation and Structure Elucidation of Compounds 1–8
2.2. SrtA Inhibitory Activity of Compounds 1–8
2.3. Antibacterial Activity and Cytotoxicity of Compounds 1–8
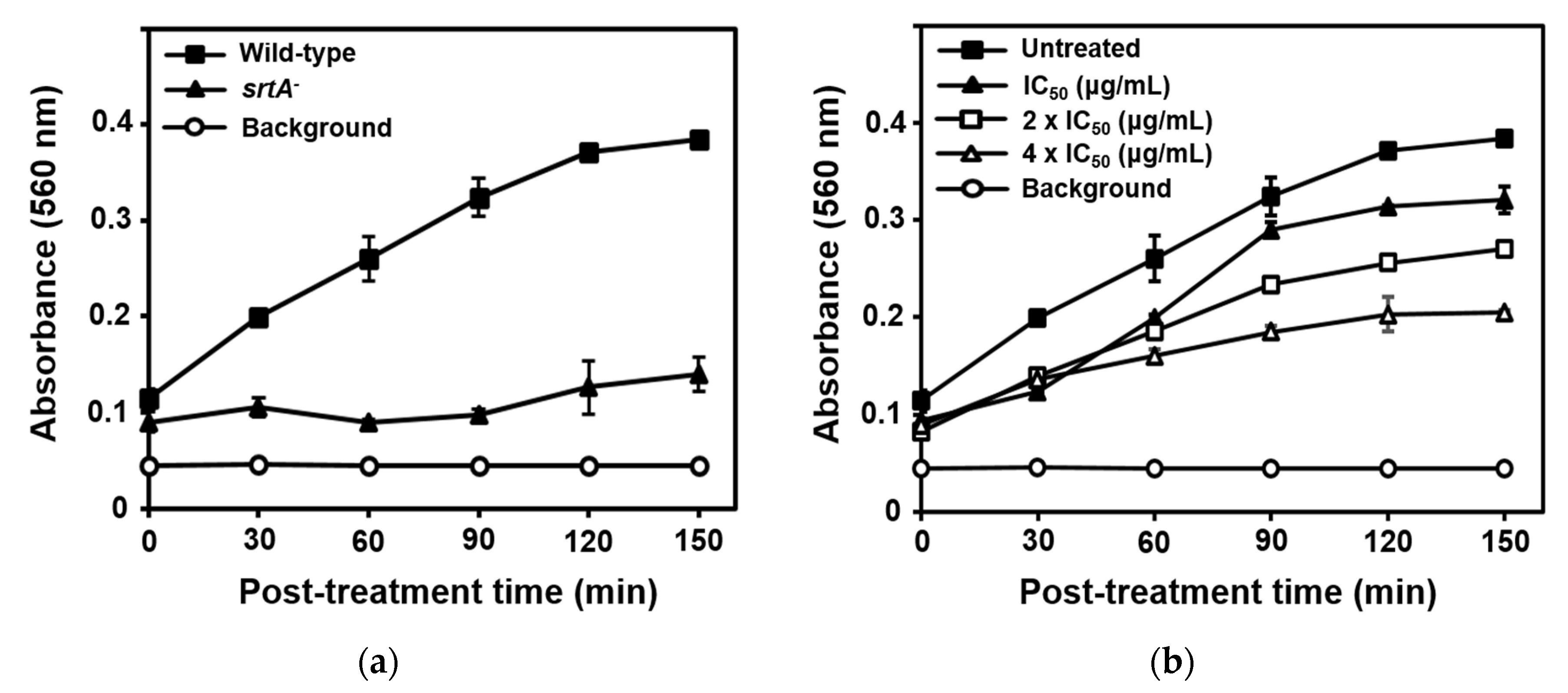
2.4. Inhibition of SrtA-mediated S. aureus Adhesion to Fibronectin by Compound 1
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Fungal Material
3.3. Extraction and Isolation
3.4. SrtA Inhibition Assay
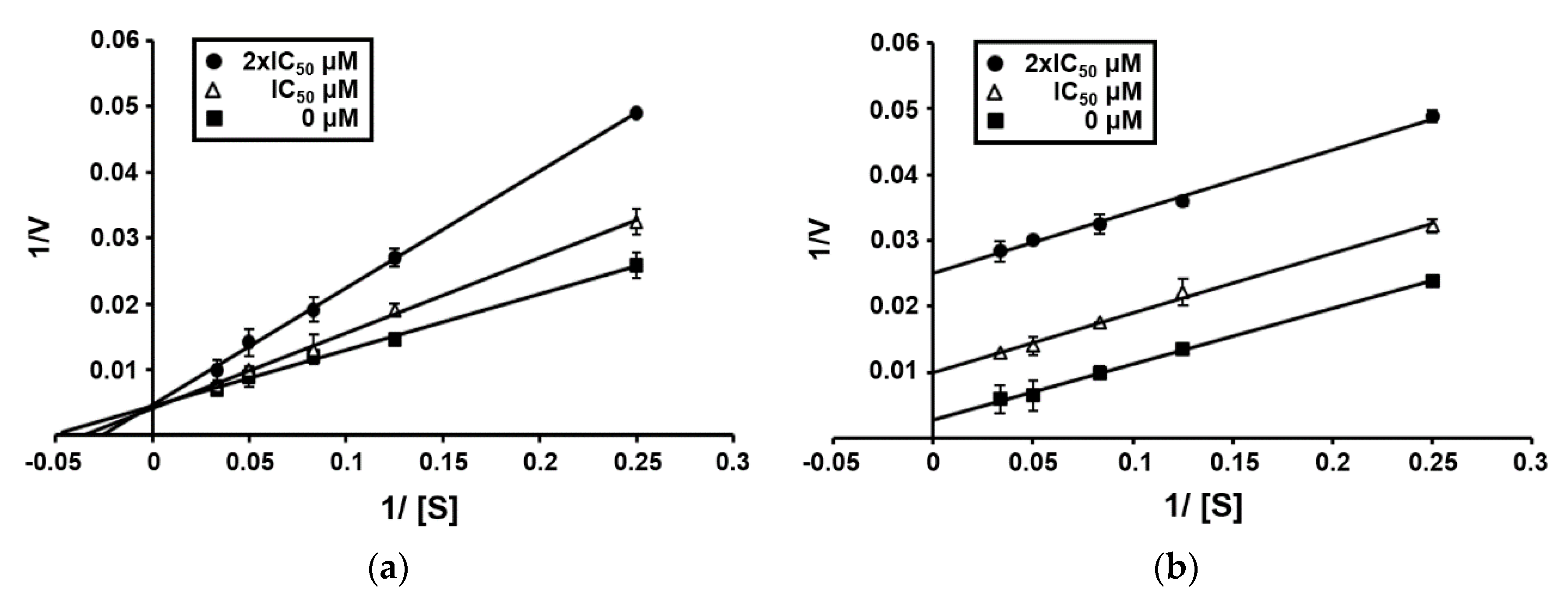
3.5. Enzyme Kinetics
3.6. Antibacterial Activity Assay
3.7. Cytotoxicity Assay
3.8. Fibronectin-Binding Assay
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gould, I.M. Antibiotic resistance: The perfect storm. Int. J. Antimicrob Agents 2009, 34, S2–S5. [Google Scholar] [CrossRef]
- Maresso, A.W.; Schneewind, O. Sortase as a target of anti-infective therapy. Pharm. Rev. 2008, 60, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Cascioferro, S.; Totsika, M.; Schillaci, D. Sortase A: An ideal target for anti-virulence drug development. Microb. Pathog. 2014, 77, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Rasko, D.A.; Sperandio, V. Anti-virulence strategies to combat bacteria-mediated disease. Nat. Rev. Drug. Discov. 2010, 9, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Hendrickx, A.P.; Budzik, J.M.; Oh, S.Y.; Schneewind, O. Architects at the bacterial surface-sortases and the assembly of pili with isopeptide bonds. Nat. Rev. Microbiol. 2011, 9, 166–176. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Skaar, E.P.; Gaspar, A.H.; Humayun, M.; Gornicki, P.; Jelenska, J.; Joachmiak, A.; Missiakas, D.M.; Schneewind, O. Passage of heme-iron across the envelope of Staphylococcus aureus. Science 2003, 299, 906–909. [Google Scholar] [CrossRef]
- Clancy, K.W.; Melvin, J.A.; McCafferty, D.G. Sortase transpeptidases: Insights into mechanism, substrate specificity and inhibition. Biopolymers 2010, 94, 385–396. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Liu, G.; Jensen, E.R.; Lenoy, E.; Schneewind, O. Staphylococcus aureus sortase mutants defective in the display of surface proteins and in the pathogenesis of animal infections. Proc. Natl. Acad. Sci. USA 2000, 97, 5510–5515. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Ton-That, H.; Su, K.; Schneewind, O. An iron-regulated sortase anchors a class of surface protein during Staphylococcus aureus pathogenesis. Proc. Natl. Acad. Sci. USA 2002, 99, 2293–2298. [Google Scholar] [CrossRef]
- Weiss, W.J.; Lenoy, E.; Murphy, T.; Tardio, L.; Burgio, P.; Projan, S.J.; Schneewind, O.; Alksne, L. Effect of srtA and srtB gene expression on the virulence of Staphylococcus aureus in animal models of infection. J. Antimicrob. Chemother 2004, 53, 480–486. [Google Scholar] [CrossRef]
- Rateb, M.E.; Ebel, R. Secondary metabolites of fungi from marine habitats. Nat. Prod. Rep. 2011, 28, 290–344. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Quan, C.; Hou, X.; Fan, S. Potential pharmacological resources: Natural bioactive compounds from marine-derived fungi. Mar. Drugs 2016, 14, 76. [Google Scholar] [CrossRef]
- Youssef, F.S.; Ashour, M.L.; Singab, A.N.B.; Wink, M. A comprehensive review of bioactive peptides from marine fungi and their biological significance. Mar. Drugs 2019, 17, 559. [Google Scholar] [CrossRef] [PubMed]
- Julianti, E.; Lee, J.-H.; Liao, L.; Park, W.; Park, S.; Oh, D.-C.; Oh, K.-B.; Shin, J. New polyaromatic metabolites from a marine-derived fungus Penicillium sp. Org. Lett. 2013, 15, 1286–1289. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.; Bae, S.Y.; Won, T.H.; You, M.; Kim, S.-H.; Oh, D.-C.; Lee, S.K.; Oh, K.-B.; Shin, J. Asperphenins A and B, lipopeptidyl benzophenones from a marine-derived Aspergillus sp. fungus. Org. Lett. 2017, 19, 2066–2069. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.-Y.; Lee, J.-H.; Park, S.C.; Lee, J.; Oh, D.-C.; Oh, K.-B.; Shin, J. New peptides from the marine-derived fungi Aspergillus allahabadii and Aspergillus ochraceopetaliformis. Mar. Drugs 2019, 17, 488. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Miura, S.; Yamashita, Y.; Ohta, T. Aspermytin A: A new neurotrophic polyketide isolated from a marine-derived fungus of the genus Aspergillus. Bioorg. Med. Chem. Lett. 2004, 14, 417–420. [Google Scholar] [CrossRef]
- Pan, C.; Shi, Y.; Chen, X.; Chen, C.-T.A.; Tao, X.; Wu, B. New compounds from a hydrothermal vent crab-associated fungus Aspergillus versicolor XZ-4. Org. Biomol. Chem. 2017, 15, 1155–1163. [Google Scholar] [CrossRef]
- Wang, J.; He, W.; Huang, X.; Tian, X.; Liao, S.; Yang, B.; Wang, F.; Zhou, X.; Liu, Y. Antifungal new oxepine-containing alkaloids and xanthones from the deep-sea-derived fungus Aspergillus versicolor SCSIO 05879. J. Agric. Food Chem. 2016, 64, 2910–2916. [Google Scholar] [CrossRef]
- Liao, L.; You, M.; Chung, B.K.; Oh, D.-C.; Oh, K.-B.; Shin, J. Alkaloidal metabolites from a marine-derived Aspergillus sp. fungus. J. Nat. Prod. 2015, 78, 349–354. [Google Scholar] [CrossRef]
- Xie, F.; Li, X.-B.; Zhou, J.-C.; Xu, Q.-Q.; Wang, X.-N.; Yuan, H.-Q.; Lou, H.-X. Secondary metabolites from Aspergillus fumigatus, an endophytic fungus from the liverwort Heteroscyphus tener (Steph) Schiffn. Chem. Biodivers. 2015, 12, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Heredia, M.L.; de la Cuesta, E.; Avendano, C. Acid-promoted reactions in 1-hydroxy, 1-dimethylaminomethyl and 1-methylene-4-arylmethyl-2,4-dihydro-1H-pyrazino[2,1-b]-quinazoline-3,6-diones. Tetrahedron 2002, 58, 6163–6170. [Google Scholar] [CrossRef]
- Oh, K.-B.; Kim, S.-H.; Lee, J.; Cho, W.-J.; Lee, T.; Kim, S. Discovery of diarylacrylonitriles as a novel series of small molecule sortase A inhibitors. J. Med. Chem. 2004, 47, 2418–2421. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.-B.; Oh, M.-N.; Kim, J.-G.; Shin, D.-S.; Shin, J. Inhibition of sortase-mediated Staphylococcus aureus adhesion to fibronectin via fibronectin-binding protein by sortase inhibitors. Appl. Microbiol. Biotechnol. 2006, 70, 102–106. [Google Scholar] [CrossRef]
- Lineweaver, H.; Burk, D. The determination of enzyme dissociation constants. J. Am. Chem. Soc. 1934, 56, 658–666. [Google Scholar] [CrossRef]
- Alksne, L.E.; Projan, S.J. Bacterial virulence as a target for antimicrobial chemotherapy. Curr. Opin. Biotechnol. 2000, 11, 625–636. [Google Scholar] [CrossRef]
- Frankel, B.A.; Bentley, M.; Kruger, R.G.; McCafferty, D.G. Vinyl sulfones: Inhibitors of srtA, a transpeptidase required for cell wall protein anchoring and virulence in Staphylococcus aureus. J. Am. Chem. Soc. 2004, 126, 3404–3405. [Google Scholar] [CrossRef]
- Kim, T.S.; Shin, Y.-H.; Lee, H.-M.; Kim, J.K.; Choe, J.H.; Jang, J.-C.; Um, S.; Jin, H.S.; Komatsu, M.; Cha, G.-H.; et al. Ohmyungsamycins promote antimicrobial responses through autophagy activation via AMP-activated protein kinase pathway. Sci. Rep. 2017, 7, 3431. [Google Scholar] [CrossRef]
- Elgalai, I.; Foster, H.A. Comparison of adhesion of wound isolates of Staphylococcus aureus to immobilized proteins. J. Appl. Micrbiol. 2003, 94, 413–420. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Compounds | SrtA IC50 μM (μg/mL) | MIC μM (μg/mL) 1 |
|---|---|---|
| 1 | 146.0 ± 2.3 (38.9 ± 0.6) | >480.5 (>128) |
| 2 | 269.4 ± 3.9 (92.5 ± 1.4) | >372.7 (>128) |
| 3 | 193.5 ± 2.5 (69.5 ± 0.9) | >356.1 (>128) |
| 4 | 267.9 ± 4.1 (107.8 ± 1.5) | >318.1 (>128) |
| 5 | 232.5 ± 3.7 (96.8 ± 1.4) | >307.4 (>128) |
| 6 | 216.4 ± 2.5 (93.1 ± 1.1) | >297.4 (>128) |
| 7 | 237.1 ± 3.8 (102.1 ± 1.2) | >297.4 (>128) |
| 8 | 235.1 ± 2.6 (83.8 ± 0.9) | >359.2 (>128) |
| Berberine chloride | 85.9 ± 1.2 (31.9 ± 0.4) | >332.2 (>128) |
| pHMB | 112.5 ± 1.7 (33.1 ± 0.5) | ND 2 |
| Ampicillin | ND | 0.4 (0.1) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.C.; Chung, B.; Lee, J.; Cho, E.; Hwang, J.-Y.; Oh, D.-C.; Shin, J.; Oh, K.-B. Sortase A-Inhibitory Metabolites from a Marine-Derived Fungus Aspergillus sp. Mar. Drugs 2020, 18, 359. https://doi.org/10.3390/md18070359
Park SC, Chung B, Lee J, Cho E, Hwang J-Y, Oh D-C, Shin J, Oh K-B. Sortase A-Inhibitory Metabolites from a Marine-Derived Fungus Aspergillus sp. Marine Drugs. 2020; 18(7):359. https://doi.org/10.3390/md18070359
Chicago/Turabian StylePark, Sung Chul, Beomkoo Chung, Jayho Lee, Eunji Cho, Ji-Yeon Hwang, Dong-Chan Oh, Jongheon Shin, and Ki-Bong Oh. 2020. "Sortase A-Inhibitory Metabolites from a Marine-Derived Fungus Aspergillus sp." Marine Drugs 18, no. 7: 359. https://doi.org/10.3390/md18070359
APA StylePark, S. C., Chung, B., Lee, J., Cho, E., Hwang, J.-Y., Oh, D.-C., Shin, J., & Oh, K.-B. (2020). Sortase A-Inhibitory Metabolites from a Marine-Derived Fungus Aspergillus sp. Marine Drugs, 18(7), 359. https://doi.org/10.3390/md18070359