Thalassosterol, a New Cytotoxic Aromatase Inhibitor Ergosterol Derivative from the Red Sea Seagrass Thalassodendron ciliatum

 ,
,  , , , ,
, , , , 
Abstract
1. Introduction
2. Results and Discussion
2.1. Metabolic Profiling
2.2. Identification of the Isolated Metabolites
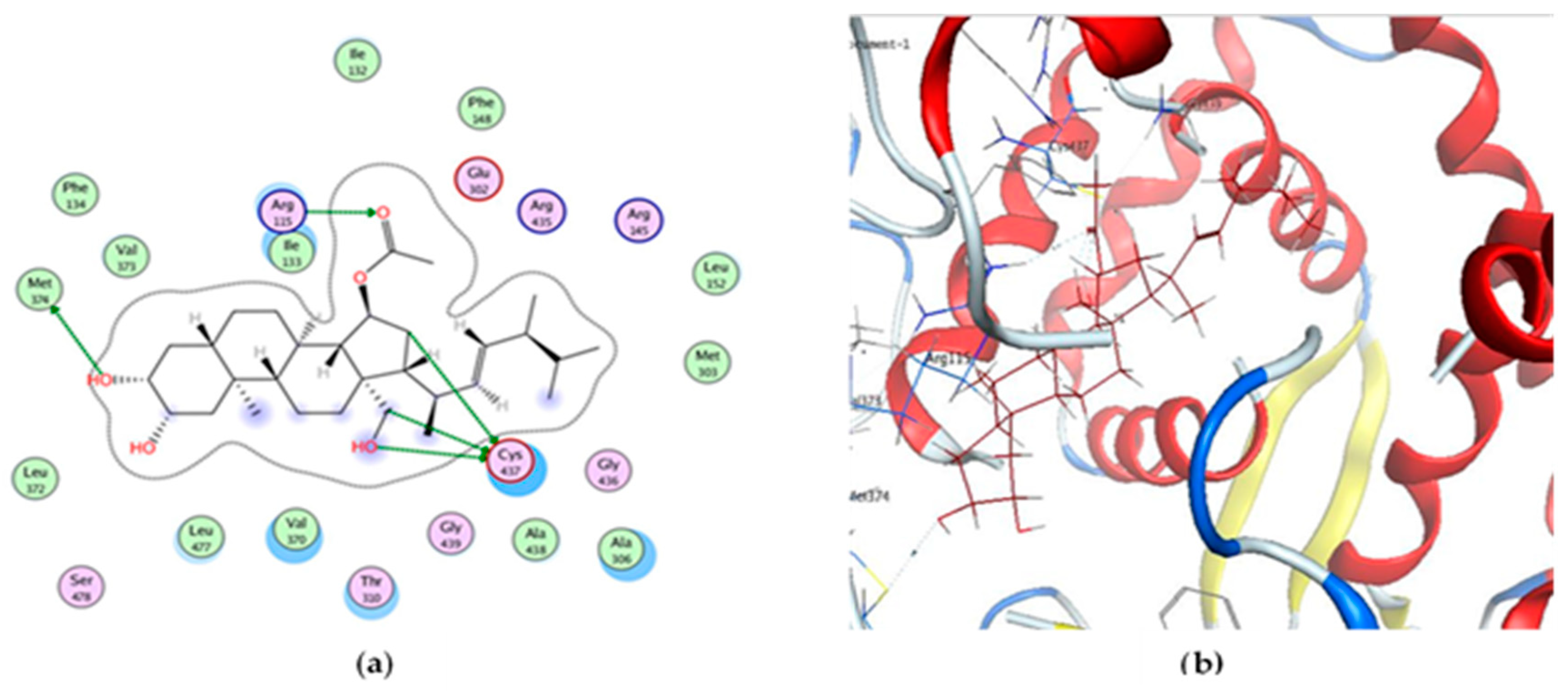
2.3. Docking Study
2.4. In Vitro Antiproliferative Activity
3. Materials and Methods
3.1. Plant Material
3.2. General Experimental Procedures
3.3. Extraction and Isolation
3.4. Metabolic Profiling
3.5. Modeling Study on the Binding Between the New Ergosterol Derivative 1 and the Aromatase Binding Site
3.6. In Vitro Antiproliferative Assay
3.6.1. In Vitro Cell Culture
3.6.2. Sulforhodamine B Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pergent-Martini, C.; Leoni, V.; Pasqualini, V.; Ardizzone, G.D.; Balestri, E.; Bedini, R.; Belluscio, A.; Belsher, T.; Borg, J.; Boudouresque, C.F.; et al. Descriptors of Posidonia oceanica meadows: Use and application. Ecol. Indic. 2005, 5, 213–230. [Google Scholar] [CrossRef]
- Bandeira, S. Dynamics, biomass and total rhizome length of the seagrass Thalassodendron ciliatum at Inhaca Island, Mozambique. Plant Ecol. 1997, 130, 133–141. [Google Scholar] [CrossRef]
- Short, F.; Carruthers, T.; Dennison, W.; Waycott, M. Global seagrass distribution and diversity: A bioregional model. J. Exp. Mar. Biol. Ecol. 2007, 350, 3–20. [Google Scholar] [CrossRef]
- Kamermans, P.; Hemmingaa, M.A.; Marbàa, N.; Mateoa, M.A.; Mtolerab, M.; Stapel, J. Leaf production, shoot demography, and flowering of Thalassodendron ciliatum along the east African coast. Aquat. Bot. 2001, 70, 243–258. [Google Scholar] [CrossRef]
- Lipkin, Y. Thalassodendron ciliatum in Sinai (northern Red Sea) with special reference to quantitative aspects. Aquat. Bot. 1998, 31, 125–139. [Google Scholar] [CrossRef]
- Hamdy, A.A.; Mettwally, W.S.A.; El Fotouh, M.A.; Rodriguez, B.; El-Dewany, A.I.; El-Toumy, S.A.A.; Hussein, A.A. Bioactive Phenolic Compounds from the Egyptian Red Sea Seagrass Thalassodendron ciliatum. Z. Naturforsch. 2012, 67, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.K.; Youssef, A.I.; Arafa, A.S.; Foad, R.; Radwan, M.M.; Ross, S.; Hassanean, H.A.; Ahmed, S.A. Anti-H5N1 virus new diglyceride ester from the Red Sea grass Thallasodendron ciliatum. Nat. Prod. Res. 2013, 27, 1625–1632. [Google Scholar] [CrossRef]
- Ramah, S.; Etwarysing, L.; Auckloo, N.; Gopeechund, A.; Bhagooli, R.; Bahorun, T. Prophylactic antioxidants and phenolics of seagrass and seaweed species: A seasonal variation study in a Southern Indian Ocean Island, Mauritius. IJMU 2014, 9, 27–37. [Google Scholar]
- Mohammed, M.M.D.; Hamdy, A.A.; El-Fiky, N.M.; Mettwally, W.S.A.; El-Beih, A.A.; Kobayashi, N. Anti-influenza A virus activity of a new dihydrochalcone diglycoside isolated from the Egyptian seagrass Thalassodendron ciliatum (Forsk.). Nat. Prod. Res. 2014, 28, 377–382. [Google Scholar] [CrossRef]
- Abdelhameed, R.F.; Ibrahim, A.K.; Yamada, K.; Ahmed, S.A. Cytotoxic and anti-inflammatory compounds from Red Sea grass Thalassodendron ciliatum. Med. Chem. Res. 2018, 27, 1238–1244. [Google Scholar] [CrossRef]
- Goda, M.S.; Eltamany, E.E.; Habib, E.S.; Hassanean, H.A.; Ahmed, A.A.; Abdelhameed, R.F.A.; Ibrahim, A.K. Gas Chromatography-Mass Spectrometry Analysis of Marine Seagrass Thalassodendron ciliatum Collected from Red Sea. Rec. Pharm. Biomed. Sci. 2020, 4, 1–15. [Google Scholar] [CrossRef]
- Sica, D.; Piccialli, V.; Masullo, A. Configuration at C-24 of sterols from the marine phanerogames Posidonia oceanica and Cymodocea nodosa. Phytochemistry 1984, 23, 2609–2611. [Google Scholar] [CrossRef]
- Kontiza, I.; Abatis, D.; Malakate, K.; Vagias, C.; Roussis, V. 3-Keto steroids from the marine organisms Dendrophyllia cornigera and Cymodocea nodosa. Steroids 2006, 71, 177–181. [Google Scholar] [CrossRef]
- Gilian, F.T.; Hogg, R.W.; Drew, E.A. The sterol and fatty acid compositions of seven tropical seagrasses from North Queensland, Australia. Phytochemistry 1984, 23, 2817–2821. [Google Scholar] [CrossRef]
- Lu, W.; Clasquin, M.F.; Melamud, E.; Amador-Noguez, D.; Caudy, A.A.; Rabinowitz, J.D. Metabolomic Analysis via Reversed-Phase Ion-Pairing Liquid Chromatography Coupled to a Stand Alone Orbitrap Mass Spectrometer. Anal. Chem. 2010, 82, 3212–3221. [Google Scholar] [CrossRef]
- Liu, J.; Meng, C.; Liu, S.; Kan, J.; Jin, C. Preparation and characterization of protocatechuic acid grafted chitosan films with antioxidant activity. Food Hydrocoll. 2017, 63, 457–466. [Google Scholar] [CrossRef]
- Cheng, Z.; Kuo, S.; Chan, S.; Ko, F.; Teng, C. Antioxidant properties of butein isolated from Dalbergia odorifera. Biochim. Biophys. Acta 1998, 1392, 291–299. [Google Scholar] [CrossRef]
- Goda, M.S.; Ahmed, S.A.; El Sherif, F.; Hassanean, H.A.; Ibrahim, A.K. Genetically stable plants with boosted flavonoids content after in vitro regeneration of the endangered Capparis spinosa L. Glob. Drugs Ther. 2017, 2, 1–7. [Google Scholar] [CrossRef][Green Version]
- Kirakosyan, A.; Mitchell Seymour, E.; Noon, K.R.; Urcuyo Llanes, D.E.; Kaufman, P.B.; Warber, S.L.; Bolling, S.F. Interactions of antioxidants isolated from tart cherry (Prunus cerasus) fruits. Food Chem. 2010, 122, 78–83. [Google Scholar] [CrossRef]
- Yen, G. Antioxidant activity of anthraquinones and anthrone. Food Chem. 2000, 70, 437–441. [Google Scholar] [CrossRef]
- Wang, Y.; Chan, F.L.; Chen, S.; Leung, L.K. The plant polyphenol butein inhibits testosterone-induced proliferation in breast cancer cells expressing aromatase. Life Sci. 2005, 77, 39–51. [Google Scholar] [CrossRef]
- Asif, M. Health effects of omega- 3,6,9 fatty acids: Perilla frutescens is a good example of plant oils. Orient. Pharm. Exp. Med. 2011, 11, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Kilic, T.; Carikci, S.; Topcu, G.; .Aslan, I.; Goren, A.C. Diterpenoids from Sideritis condensata. Evaluation of chemotaxonomy of Sideritis species and insecticidal activity. Chem. Nat. Compd. 2009, 45, 918–920. [Google Scholar] [CrossRef]
- Smyrniotopoulos, V.; Vagias, C.; Bruyère, C.; Lamoral-Theys, D.; Kiss, R.; Roussis, V. Structure and in vitro antitumor activity evaluation of brominated diterpenes from the red alga Sphaerococcus coronopifolius. Bioorg. Med. Chem. 2010, 18, 1321–1330. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yong, T.; Zhang, Y.; Su, J.; Jiao, C.; Xie, Y. Anti-tumor and anti-angiogenic ergosterols from Ganoderma lucidum. Front. Chem. 2017, 5, 85. [Google Scholar] [CrossRef]
- Osman, M.; Sazalli, N.N.H.; Willium, D.; Golam, F.; Nezhadahmadi, A. Potentiality of Roselle and On-ion (Allium cepa) peel as Raw Materials for Producing Protocatechuic Acid in Tropical Malaysia: A Comparative Study. INDJST 2014, 7, 1847–1851. [Google Scholar]
- Sebolai, O.M.; Kock, J.L.F.; Pohl, C.H.; Botes, P.J.; Nigam, S. Report on the discovery of a novel 3-hydroxy oxylipin cascade in the yeast Saccharomycopsis synnaedendra. Prostaglandins Other Lipid Mediat. 2004, 74, 139–146. [Google Scholar] [CrossRef]
- Mikhailova, M.V.; Bemis, D.L.; Wise, M.L.; Gerwick, W.H.; Norris, J.N.; Jacobs, R.S. Structure and biosynthesis of novel conjugated polyene fatty acids from the marine green alga Anadyomene stellate. Lipids 1995, 30, 583–589. [Google Scholar] [CrossRef]
- Zhang, Y.; Seeram, N.P.; Lee, R.; Feng, L.; Heber, D. The Isolation and Identification of Isolation and Identification of Strawberry Phenolics with Antioxidant and Human Cancer Cell Antiproliferative Properties. J. Agric. Food Chem. 2008, 56, 670–675. [Google Scholar] [CrossRef]
- Wang, S.; Li, X.M.; Teuscher, F.; Li, D.L.; Diesel, A.; Ebel, R.; Proksch, P.; Wang, B.G. Chaetopyranin, a benzaldehyde derivative, and other related metabolites from Chaetomium globosum, an endophytic fungus derived from the marine red alga Polysiphonia urceolata. J. Nat. Prod. 2006, 69, 1622–1625. [Google Scholar] [CrossRef]
- Paranjape, S.R.; Chiang, Y.M.; Sanchez, J.F.; Entwistle, R.; Wang, C.C.; Oakley, B.R.; Gamblin, T.C. Inhibition of Tau aggregation by three Aspergillus nidulans secondary metabolites: 2,ω-dihydroxyemodin, asperthecin, and asperbenzaldehyde. Planta Med. 2014, 80, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Wei, Y.; Ito, Y. Preparative Isolation of Isorhamnetin from Stigma maydis using High Speed Countercurrent Chromatography. J. Liq. Chromatogr. Relat. Technol. 2008, 32, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Smyrniotopoulos, V.; Vagias, C.; Roussis, V. Sphaeroane and Neodolabellane Diterpenes from the Red Alga Sphaerococcus coronopifolius. Mar. Drugs 2009, 7, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Nieto, I.; Avila, I. Determination of fatty acids and triterpenoid compounds from the fruiting body of Suillus luteus. Rev. Colomb. Quim. 2008, 37, 297–304. [Google Scholar]
- Zhang, W.; Che, C. Isomalabaricane-Type Nortriterpenoids and Other Constituents of the Marine Sponge Geodia japonica. J. Nat. Prod. 2001, 64, 1489–1492. [Google Scholar] [CrossRef]
- Liu, Y. Dietary Chinese Herbs, 1st ed.; Springer: Vienna, Austria, 2015; pp. 789–796. [Google Scholar] [CrossRef]
- Goad, L.J.; Akihisa, T. Analysis of Sterols, 1st ed.; Blackie Academic and Professional: London, UK, 1997; pp. 1–437. [Google Scholar] [CrossRef]
- Ványolòs, A.; Béni, Z.; Dékány, M.; Simon, A.; Báthori, M. Novel Ecdysteroids from Serratula wolffii. Sci. World J. 2012, 1–5. [Google Scholar] [CrossRef][Green Version]
- Shady, N.H.; Abdelmohsen, U.R.; Ahmed, S.; Fouad, M.; Kamel, M.S. Phytochemical and biological investigation of the red sea marine sponge Hyrtios sp. J. Pharmacogn. Phytochem. 2017, 6, 241–246. [Google Scholar]
- Ghosh, T.; Maity, T.K.; Singh, J. Evaluation of antitumor activity of stigmasterol, a constituent isolated from Bacopa monnieri Linn. aerial parts against Ehrlich Ascites Carcinoma in mice. Orient. Pharm. Exp. Med. 2011, 11, 41–49. [Google Scholar] [CrossRef]
- Roleira, F.M.F.; Varela, C.; Amaral, C.; Costa, S.C.; Correia-da-Silva, G.; Moraca, F.; Costa, G.; Alcaro, S.; Teixeira, N.A.A.; Tavares da Silva, E.J. C-6α- vs C-7α-Substituted Steroidal Aromatase Inhibitors: Which Is Better? Synthesis, Biochemical Evaluation, Docking Studies, and Structure–Activity Relationships. J. Med. Chem. 2019, 62, 3636–3657. [Google Scholar] [CrossRef]
- Rampogu, S.; Ravinder, D.; Pawar, S.C.; Lee, K.W. Natural Compound Modulates the Cervical Cancer Microenvironment—A Pharmacophore Guided Molecular Modelling Approaches. J. Clin. Med. 2018, 7, 551. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Bajaj, S.; Bodla, R.B. Preclinical screening methods in cancer. Indian J. Pharmacol. 2016, 48, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, R.S.; Seif El-Din, A.A.; Abu-Serie, M.; Abd El Rahman, N.M.; El-Demellawy, M.; Metwally, A.M. Investigation of In-Vitro Cytotoxic and Potential Anticancer Activities of Flavonoidal Aglycones from Egyptian Propolis. Rec. Pharm. Biomed. Sci. 2017, 2, 13–20. [Google Scholar] [CrossRef]
- Abdelhafez, O.H.; Othman, E.M.; Fahim, J.R.; Desouky, S.Y.; Pimentel-Elardo, S.M.; Nodwell, J.R.; Schirmeister, T.; Tawfike, A.; Abdelmohsen, U.R. Metabolomics analysis and biological investigation of three Malvaceae plants. Phytochem. Anal. 2020, 31, 204–214. [Google Scholar] [CrossRef]
- Gasteiger, J.; Rudolph, C.; Sadowski, J. Automatic generation of 3D-atomic coordinates for organic molecules. Tetrahedron Comput. Methodol. 1990, 3, 537–547. [Google Scholar] [CrossRef]
- Skehan, P.; Strong, R.; Scudiero, D.; Monks, A.; McMahon, J.; Vistica, D.; Warren, J.T.; Bokesch, H.; Kenney, S.; Boyd, M.R. New colorimetric cytotoxicity assay for anticancer drug screening. J. Natl. Cancer Inst. 1990, 82, 1107–1112. [Google Scholar] [CrossRef] [PubMed]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef]
- Abu-Dahab, R.; Afifi, F. Antiproliferative activity of selected medicinal plants of Jordan against a breast adenocarcinoma cell line (MCF7). Sci. Pharm. 2007, 75, 121–136. [Google Scholar] [CrossRef]


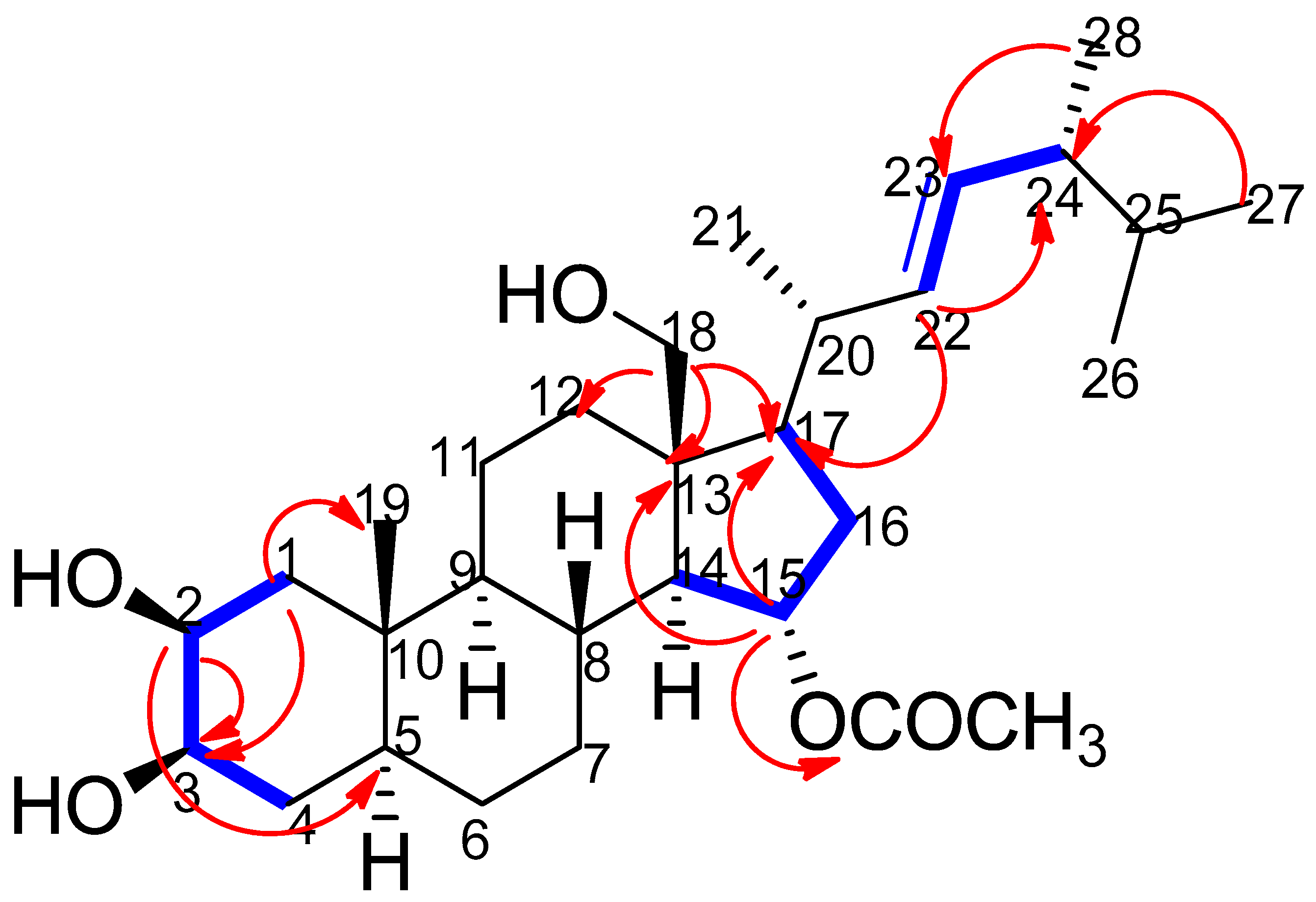
 ) and Heteronuclear Multiple Bond Correlation (HMBC) (
) and Heteronuclear Multiple Bond Correlation (HMBC) ( ) interactions in thalassosterol (1), (2β,18-dihydroxy-15α-acetoxy-5,6,7,8-tetrahydroergosterol). Blue bold line represents 1H-1H COSY correlations between H1/H2, H2/H3, H3/H4, H14/H15, H15/H16, H16/H17, H22/H23 and H23/H24, while red single-head arrow represents HMBC correlations between H1/C19, H1/C3, H2/C3, H2/C5, H15/C13, H15/C17, H15/CO, H18/C12, H18/C17, H18/C13, H22/C17, H22/C24, H27/C24 and H28/C23.
) and Heteronuclear Multiple Bond Correlation (HMBC) () interactions in thalassosterol (1), (2β,18-dihydroxy-15α-acetoxy-5,6,7,8-tetrahydroergosterol). Blue bold line represents 1H-1H COSY correlations between H1/H2, H2/H3, H3/H4, H14/H15, H15/H16, H16/H17, H22/H23 and H23/H24, while red single-head arrow represents HMBC correlations between H1/C19, H1/C3, H2/C3, H2/C5, H15/C13, H15/C17, H15/CO, H18/C12, H18/C17, H18/C13, H22/C17, H22/C24, H27/C24 and H28/C23.
) interactions in thalassosterol (1), (2β,18-dihydroxy-15α-acetoxy-5,6,7,8-tetrahydroergosterol). Blue bold line represents 1H-1H COSY correlations between H1/H2, H2/H3, H3/H4, H14/H15, H15/H16, H16/H17, H22/H23 and H23/H24, while red single-head arrow represents HMBC correlations between H1/C19, H1/C3, H2/C3, H2/C5, H15/C13, H15/C17, H15/CO, H18/C12, H18/C17, H18/C13, H22/C17, H22/C24, H27/C24 and H28/C23.
) and Heteronuclear Multiple Bond Correlation (HMBC) () interactions in thalassosterol (1), (2β,18-dihydroxy-15α-acetoxy-5,6,7,8-tetrahydroergosterol). Blue bold line represents 1H-1H COSY correlations between H1/H2, H2/H3, H3/H4, H14/H15, H15/H16, H16/H17, H22/H23 and H23/H24, while red single-head arrow represents HMBC correlations between H1/C19, H1/C3, H2/C3, H2/C5, H15/C13, H15/C17, H15/CO, H18/C12, H18/C17, H18/C13, H22/C17, H22/C24, H27/C24 and H28/C23.
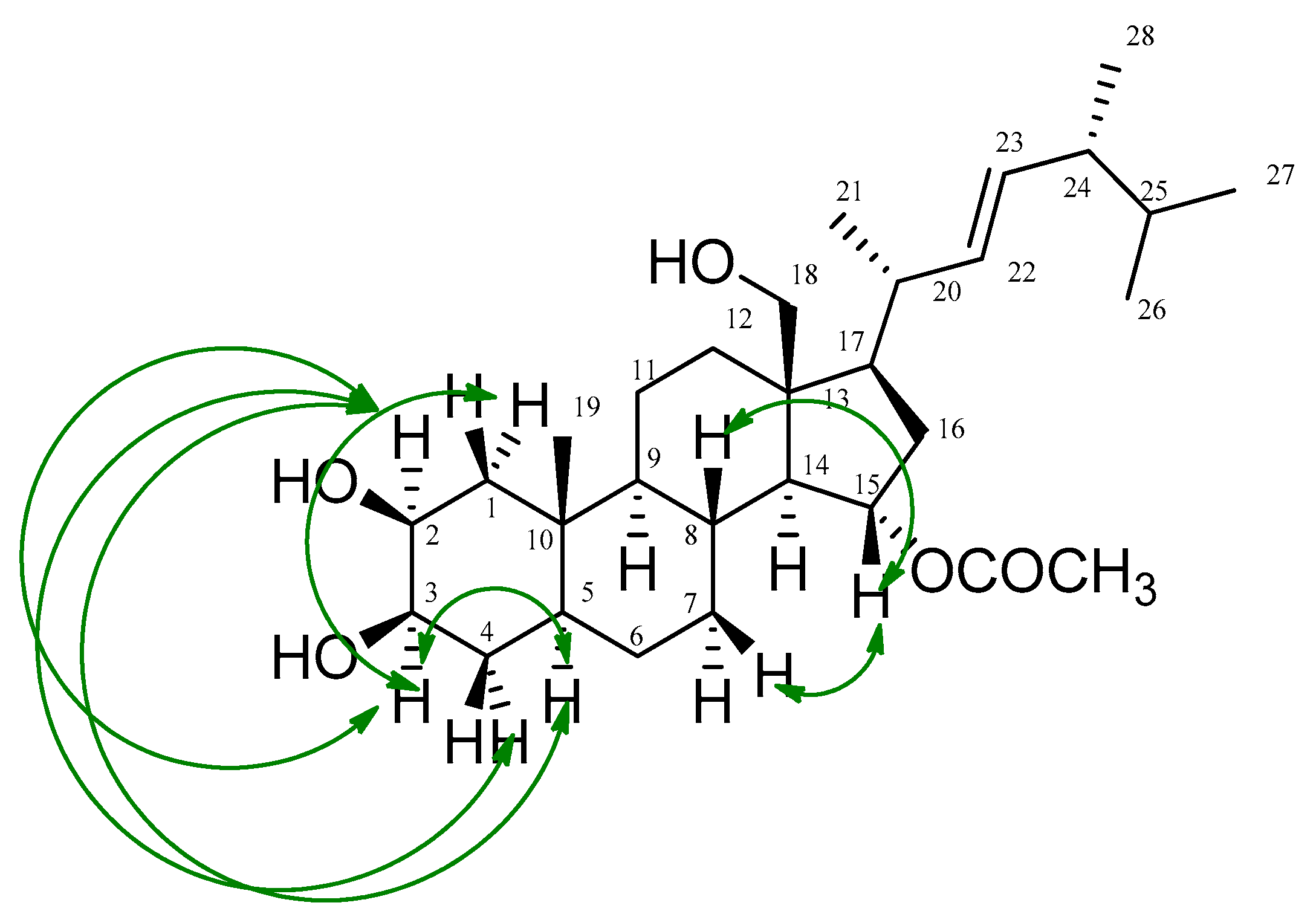
 ) correlations of thalassosterol (1), (2β,18-dihydroxy-15α-acetoxy-5,6,7,8-tetrahydroergosterol). Green double-head arrow represents NOESY correlations between H2/H3, H2/H5, H2/H4a, H3/H1a, H3/H2, H3/H5, H15/H7b and H15/H8.
) correlations of thalassosterol (1), (2β,18-dihydroxy-15α-acetoxy-5,6,7,8-tetrahydroergosterol). Green double-head arrow represents NOESY correlations between H2/H3, H2/H5, H2/H4a, H3/H1a, H3/H2, H3/H5, H15/H7b and H15/H8.
) correlations of thalassosterol (1), (2β,18-dihydroxy-15α-acetoxy-5,6,7,8-tetrahydroergosterol). Green double-head arrow represents NOESY correlations between H2/H3, H2/H5, H2/H4a, H3/H1a, H3/H2, H3/H5, H15/H7b and H15/H8.
) correlations of thalassosterol (1), (2β,18-dihydroxy-15α-acetoxy-5,6,7,8-tetrahydroergosterol). Green double-head arrow represents NOESY correlations between H2/H3, H2/H5, H2/H4a, H3/H1a, H3/H2, H3/H5, H15/H7b and H15/H8.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
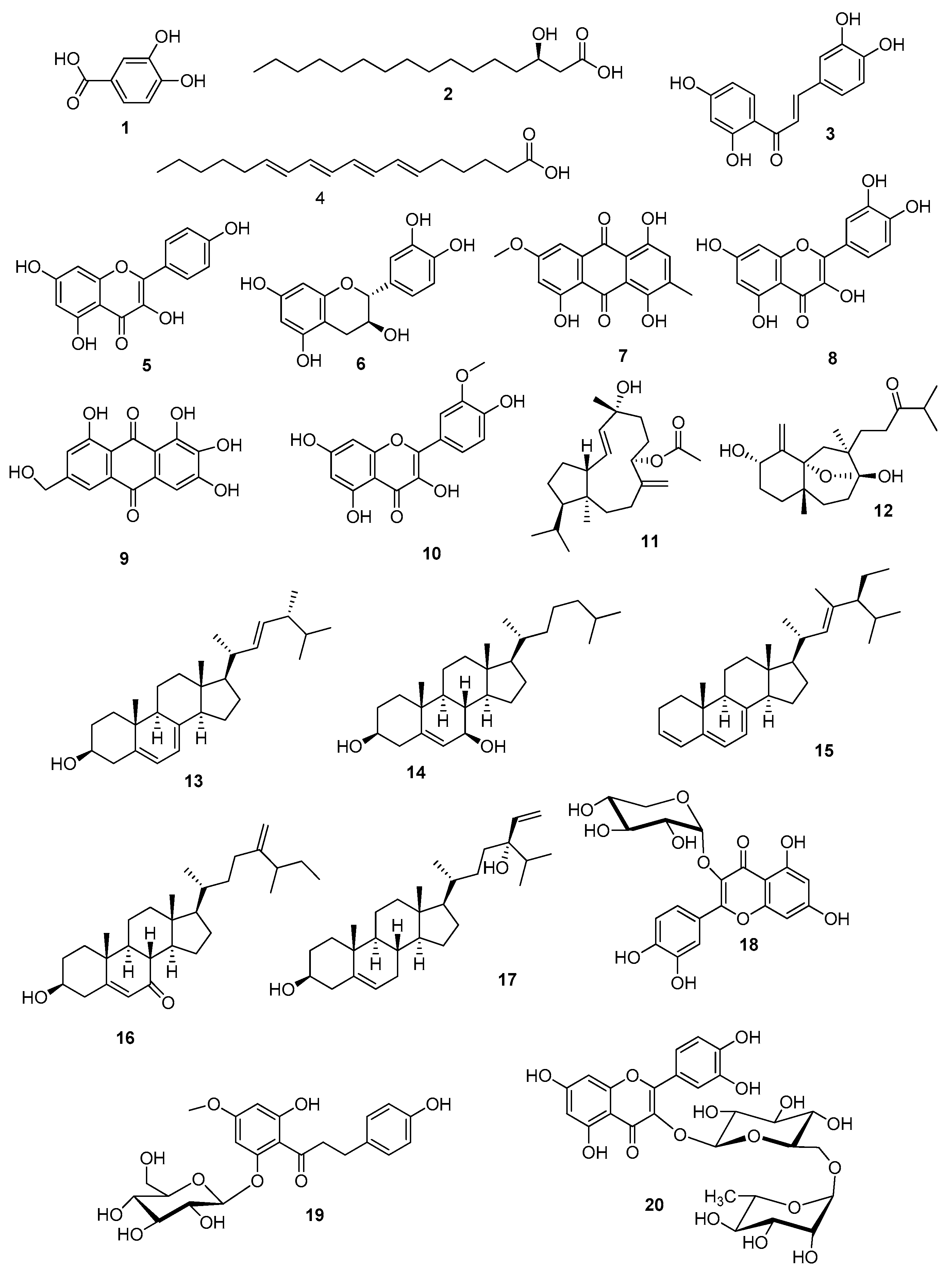
| Polarity Mode | Retention Time (min) | MZmine ID | m/z * | Measured Mass | Expected Mass | Mass Error (ppm) | Name | Molecular Formula ** | Source | Ref. | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | positive | 1.51 | 9930 | 155.0347 | 154.0274 | 154.0266 | 5.2 | Protocatechuic acid | C7H6O4 | Allium cepa | [26] |
| 2 | negative | 11.29 | 382 | 271.2269 | 272.2342 | 272.2351 | −3.3 | 3R-Hydroxypalmitic acid | C16H32O3 | Saccharomycopsis sp. | [27] |
| 3 | positive | 5.07 | 9019 | 273.0748 | 272.0675 | 272.0685 | −3.7 | Butein | C15H12O5 | Dalbergia odorifera | [17] |
| 4 | positive | 5.63 | 9699 | 277.2153 | 276.2080 | 276.2089 | −3.3 | 6E,8E,10E,12E-Octadecatetraenoic acid | C18H28O2 | Anadyomene stellata | [28] |
| 5 | negative | 4.46 | 3937 | 285.0393 | 286.0466 | 286.0477 | −3.8 | Kaempferol | C15H10O6 | Fragaria chiloensis | [29] |
| 6 | negative | 2.18 | 3954 | 289.0705 | 290.0778 | 290.0790 | −4.1 | Catechin | C15H14O6 | T. ciliatum | [6] |
| 7 | negative | 5.43 | 4767 | 299.0551 | 300.0623 | 300.0634 | −3.7 | 1,4,5-Trihydroxy-7-methoxy-3-methylanthraquinone | C16H12O6 | Chaetomium globosum | [30] |
| 8 | negative | 5.07 | 4942 | 301.0341 | 302.0414 | 302.0427 | −4.3 | Quercetin | C15H10O7 | Fragaria chiloensis | [29] |
| 9 | positive | 3.37 | 9298 | 303.0503 | 302.0430 | 302.0427 | 0.9 | 2-ω-Dihydroxyemodin | C15H10O7 | Aspergillus nidulans | [31] |
| 10 | negative | 4.76 | 4129 | 315.0497 | 316.0570 | 316.0583 | −4.1 | Isorhamnetin | C16H12O7 | Stigma maydis | [32] |
| 11 | negative | 13.90 | 4382 | 347.2581 | 348.2654 | 348.2664 | −2.9 | Sphaerollane I | C22H36O3 | Sphaerococs coronopifolis | [33] |
| 12 | positive | 8.66 | 12289 | 353.2706 | 352.2634 | 352.2614 | 5.7 | Linearol | C21H36O4 | Sideritis condensata | [23] |
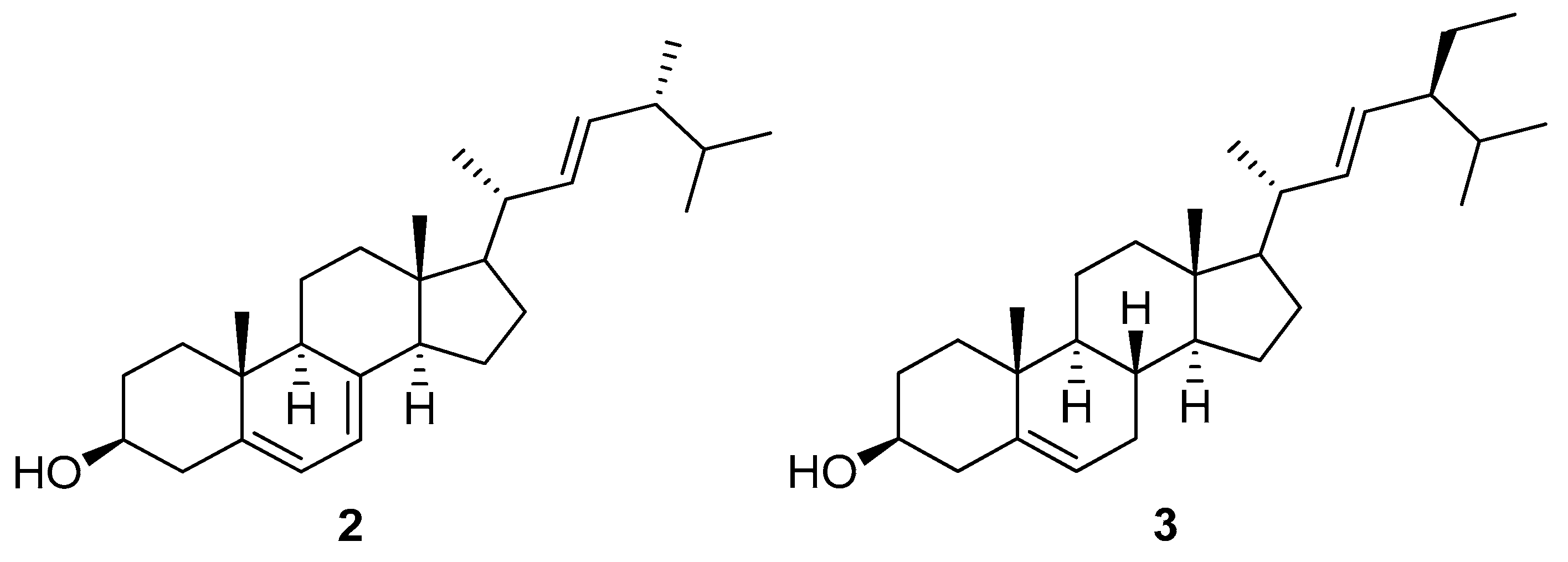
| 13 | positive | 12.71 | 9842 | 397.3452 | 396.3379 | 396.3392 | −3.3 | Ergosterol (2) | C28H44O | Ganoderma lucidum | [25] |
| 14 | positive | 13.60 | 10050 | 403.3528 | 402.3456 | 402.3498 | −10.4 | 7β-Hydroxycholesterol | C27H46O2 | T. ciliatum | [10] |
| 15 | positive | 12.49 | 8715 | 407.3677 | 406.3604 | 406.3600 | 0.9 | 23-Methylstigmasta-3Z,5Z,7Z,22E-tetraene | C30H46 | Suillus luteus | [34] |
| 16 | positive | 10.46 | 8941 | 427.3572 | 426.3499 | 426.3498 | 0.2 | 26-Methylergosta-5,24(28)-diene-7-one-3-ol | C29H46O2 | Geodia japonica | [35] |
| 17 | positive | 15.43 | 320 | 429.3732 | 428.3659 | 428.3654 | 1.2 | 24R-Stigmasta-5,28-diene-3β,24 -diol | C29H48O2 | Sargassum fusiforme | [36] |
| 18 | negative | 3.37 | 4110 | 433.0765 | 434.0838 | 434.0849 | −2.5 | Quercetin-3-O-β-D-xylopyranoside | C20H18O11 | T. ciliatum | [6] |
| 19 | positive | 6.10 | 705 | 451.1642 | 450.1570 | 450.1526 | 9.8 | Asebotin | C22H26O10 | T. ciliatum | [6] |
| 20 | negative | 2.33 | 5311 | 609.1466 | 610.1539 | 610.1534 | 0.8 | Rutin | C27H30O16 | T. ciliatum | [6] |
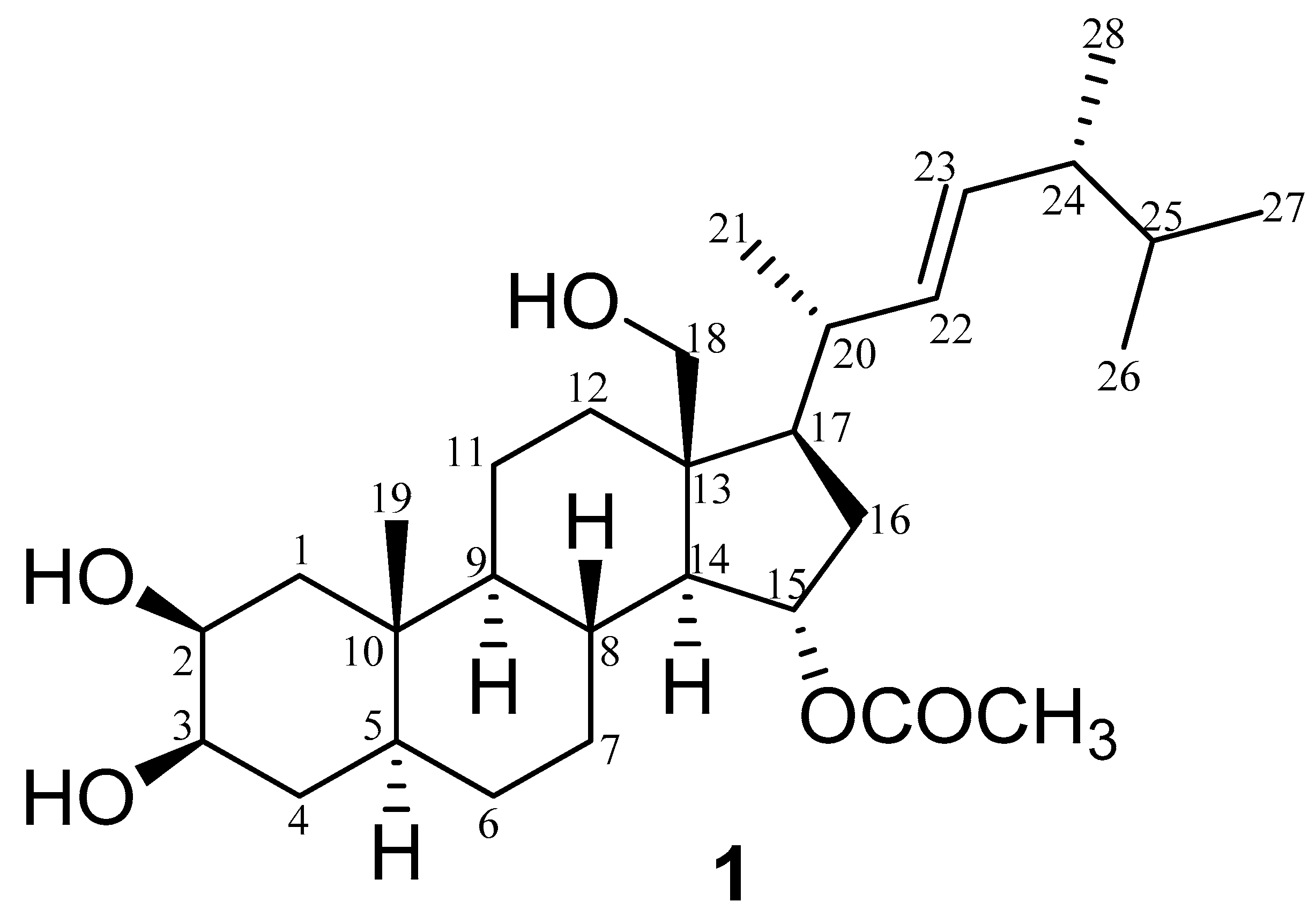
| No. | δH | δC |
|---|---|---|
| 1 | 1.42, 2.09, m | 39.1 |
| 2 | 4.72, br. d (J = 2.2) | 76.0 |
| 3 | 4.75, br. d (J = 2.2) | 76.3 |
| 4 | 1.60, 1.80, m | 30.4 |
| 5 | 1.59, m | 40.2 |
| 6 | 1.25, 1.50, m | 28.9 |
| 7 | 0.96, 1.59, m | 32.3 |
| 8 | 1.74, m | 32.4 |
| 9 | 0.83, m | 56.8 |
| 10 | ---- | 36.5 |
| 11 | 1.37, 1.56, m | 21.8 |
| 12 | 0.96, 2.62, m | 36.2 |
| 13 | --- | 48.8 |
| 14 | 1.20, m | 59.8 |
| 15 | 4.99, m | 74.4 |
| 16 | 1.30, 2.46, m | 39.9 |
| 17 | 1.14, m | 58.1 |
| 18 | 3.65, d (J = 12.2) 3.75 d (J = 12.2) | 60.6 |
| 19 | 1.03, br. s | 14.2 |
| 20 | 1.72, m | 36.6 |
| 21 | 1.06, d (J = 6.4) | 19.9 |
| 22 | 5.15, dd (J = 15.2, 6.5) | 137.5 |
| 23 | 5.21, dd (J = 15.2, 7.2) | 133.2 |
| 24 | 1.84, m | 44.4 |
| 25 | 1.51, m | 29.2 |
| 26 | 0.873, d (J = 6.64) | 23.2 |
| 27 | 0.877, d (J = 6.64) | 23.0 |
| 28 | 0.92, d (J = 6.8) | 18.4 |
| -CO-CH3 | 2.00, s | 21.2 |
| -CO-CH3 | ---- | 172.6 |
| Human Cancer Cell Lines | |||
|---|---|---|---|
| HeLa IC50 (µM) | HepG2 IC50 (µM) | MCF-7 IC50 (µM) | |
| Thalassosterol (1) | 8.13 ± 0.21 * | 48.64 ± 0.22 * | 14.26 ± 0.40 * |
| Doxorubicin | 6.77 ± 0.07 | 9.02 ± 0.06 | 8.65 ± 0.03 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelhameed, R.F.A.; Habib, E.S.; Goda, M.S.; Fahim, J.R.; Hassanean, H.A.; Eltamany, E.E.; Ibrahim, A.K.; AboulMagd, A.M.; Fayez, S.; El-kader, A.M.A.; et al. Thalassosterol, a New Cytotoxic Aromatase Inhibitor Ergosterol Derivative from the Red Sea Seagrass Thalassodendron ciliatum. Mar. Drugs 2020, 18, 354. https://doi.org/10.3390/md18070354
Abdelhameed RFA, Habib ES, Goda MS, Fahim JR, Hassanean HA, Eltamany EE, Ibrahim AK, AboulMagd AM, Fayez S, El-kader AMA, et al. Thalassosterol, a New Cytotoxic Aromatase Inhibitor Ergosterol Derivative from the Red Sea Seagrass Thalassodendron ciliatum. Marine Drugs. 2020; 18(7):354. https://doi.org/10.3390/md18070354
Chicago/Turabian StyleAbdelhameed, Reda F. A., Eman S. Habib, Marwa S. Goda, John Refaat Fahim, Hashem A. Hassanean, Enas E. Eltamany, Amany K. Ibrahim, Asmaa M. AboulMagd, Shaimaa Fayez, Adel M. Abd El-kader, and et al. 2020. "Thalassosterol, a New Cytotoxic Aromatase Inhibitor Ergosterol Derivative from the Red Sea Seagrass Thalassodendron ciliatum" Marine Drugs 18, no. 7: 354. https://doi.org/10.3390/md18070354
APA StyleAbdelhameed, R. F. A., Habib, E. S., Goda, M. S., Fahim, J. R., Hassanean, H. A., Eltamany, E. E., Ibrahim, A. K., AboulMagd, A. M., Fayez, S., El-kader, A. M. A., Al-Warhi, T., Bringmann, G., Ahmed, S. A., & Abdelmohsen, U. R. (2020). Thalassosterol, a New Cytotoxic Aromatase Inhibitor Ergosterol Derivative from the Red Sea Seagrass Thalassodendron ciliatum. Marine Drugs, 18(7), 354. https://doi.org/10.3390/md18070354