Types and Distribution of Bioactive Polyunsaturated Aldehydes in a Gradient from Mesotrophic to Oligotrophic Waters in the Alborán Sea (Western Mediterranean)

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
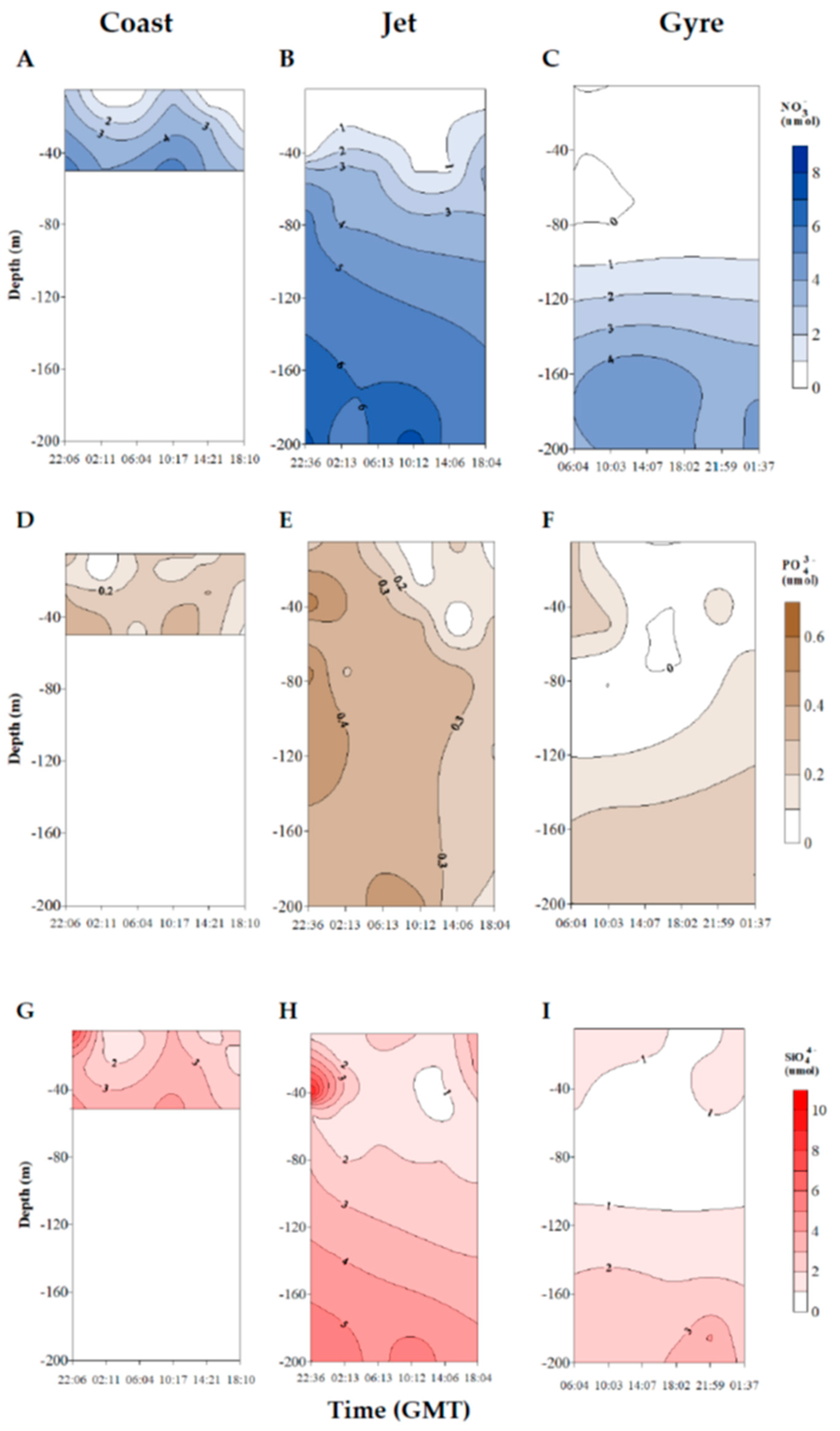
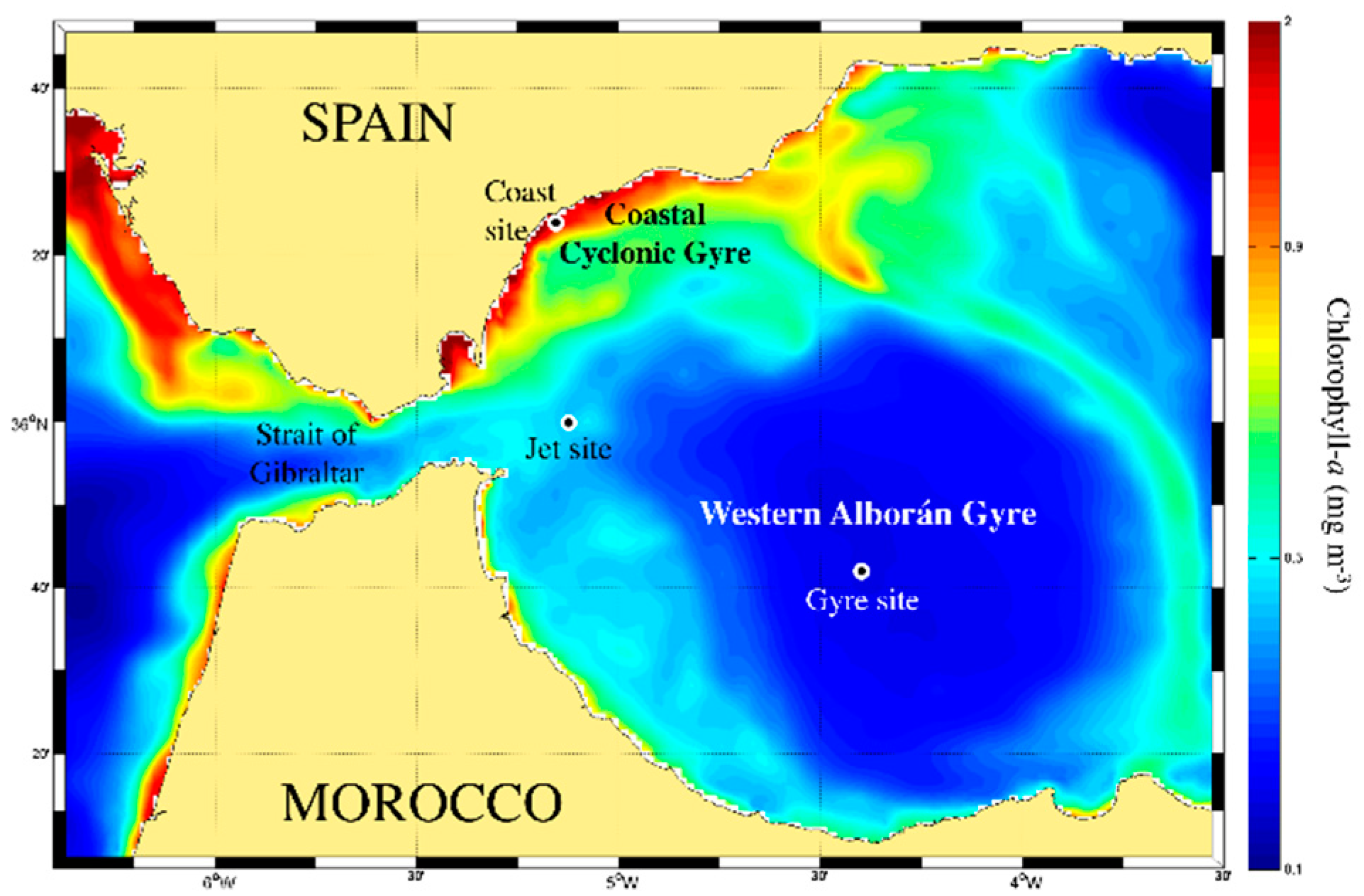
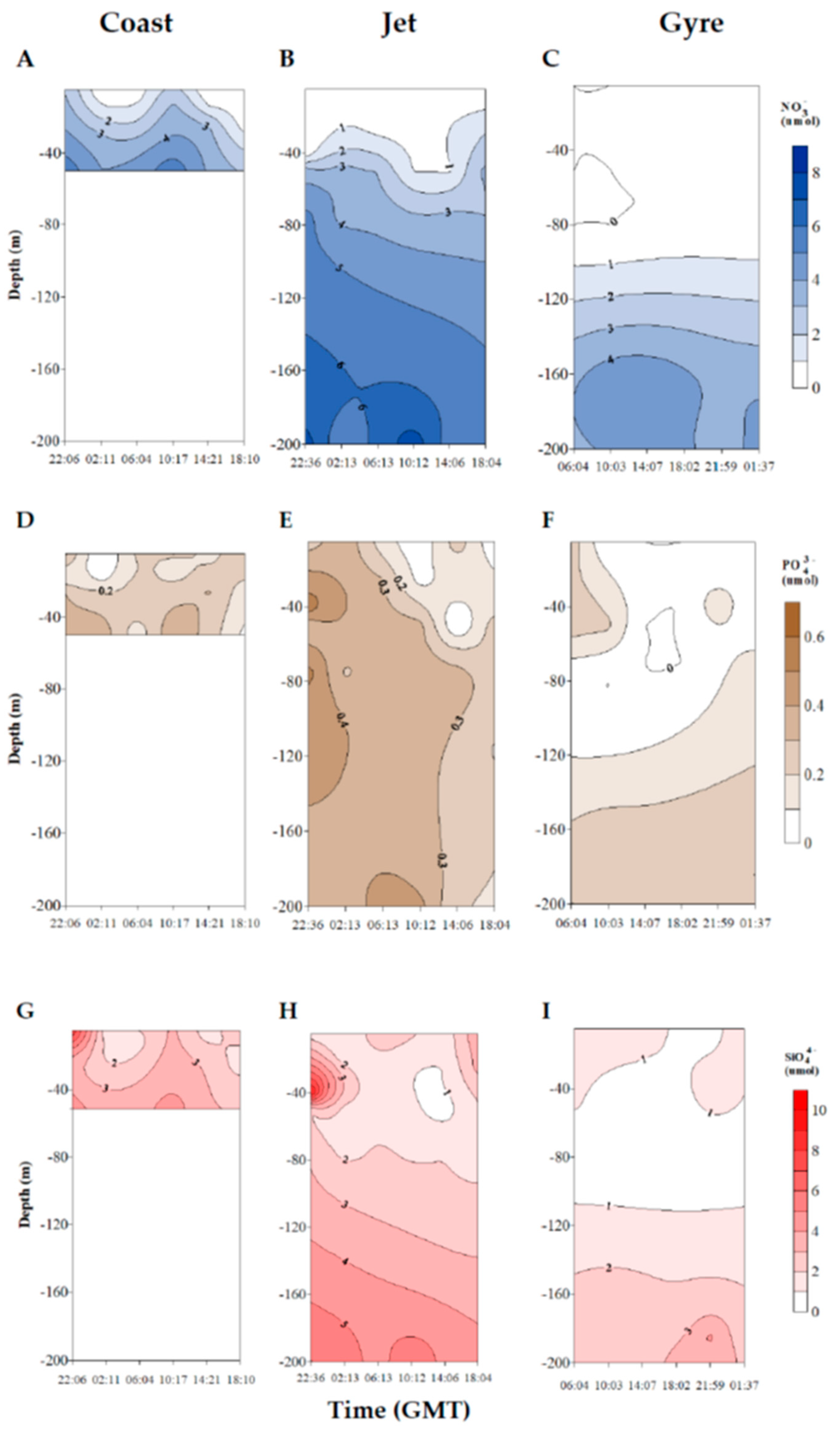
2.1. Trophic Status of the Different Sites and Physicochemical Characterization of the Water Mass
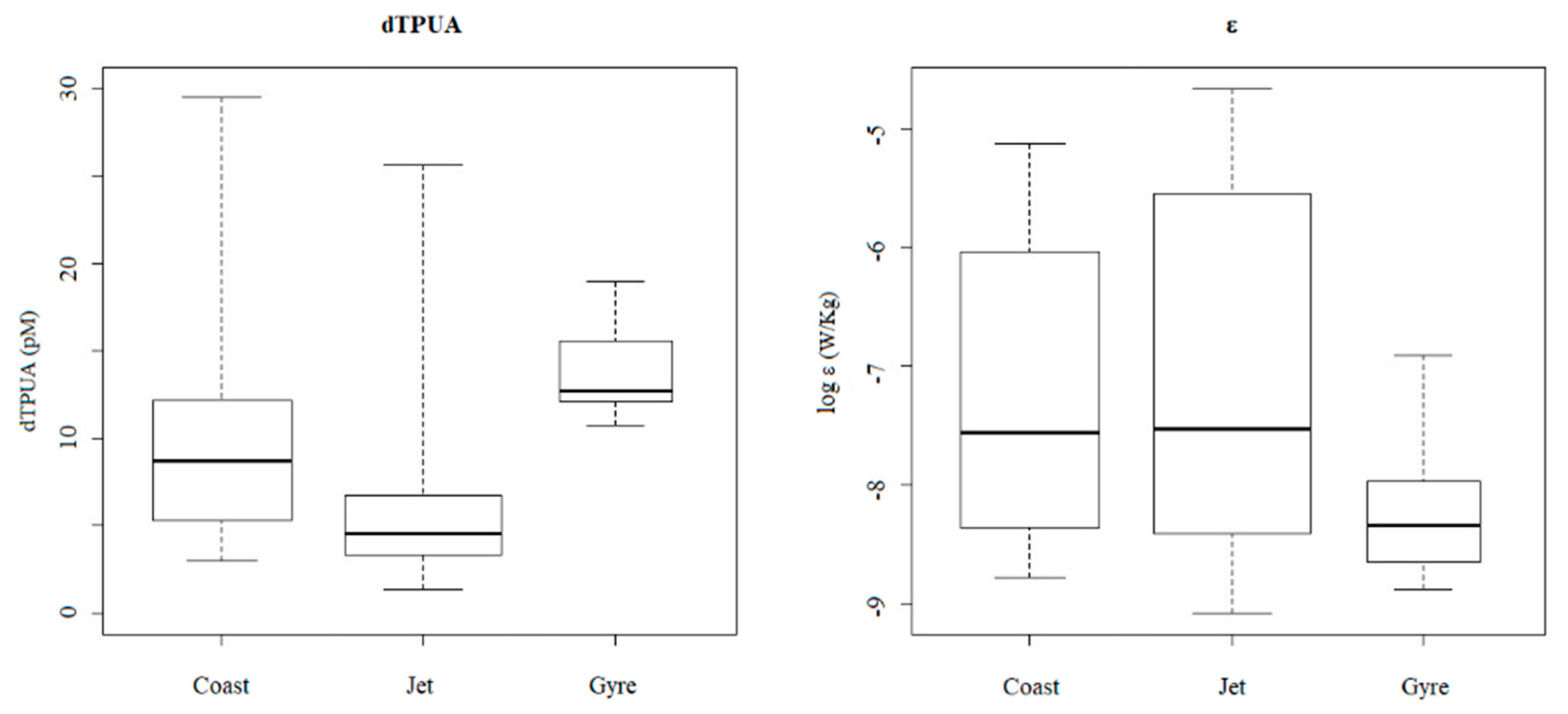
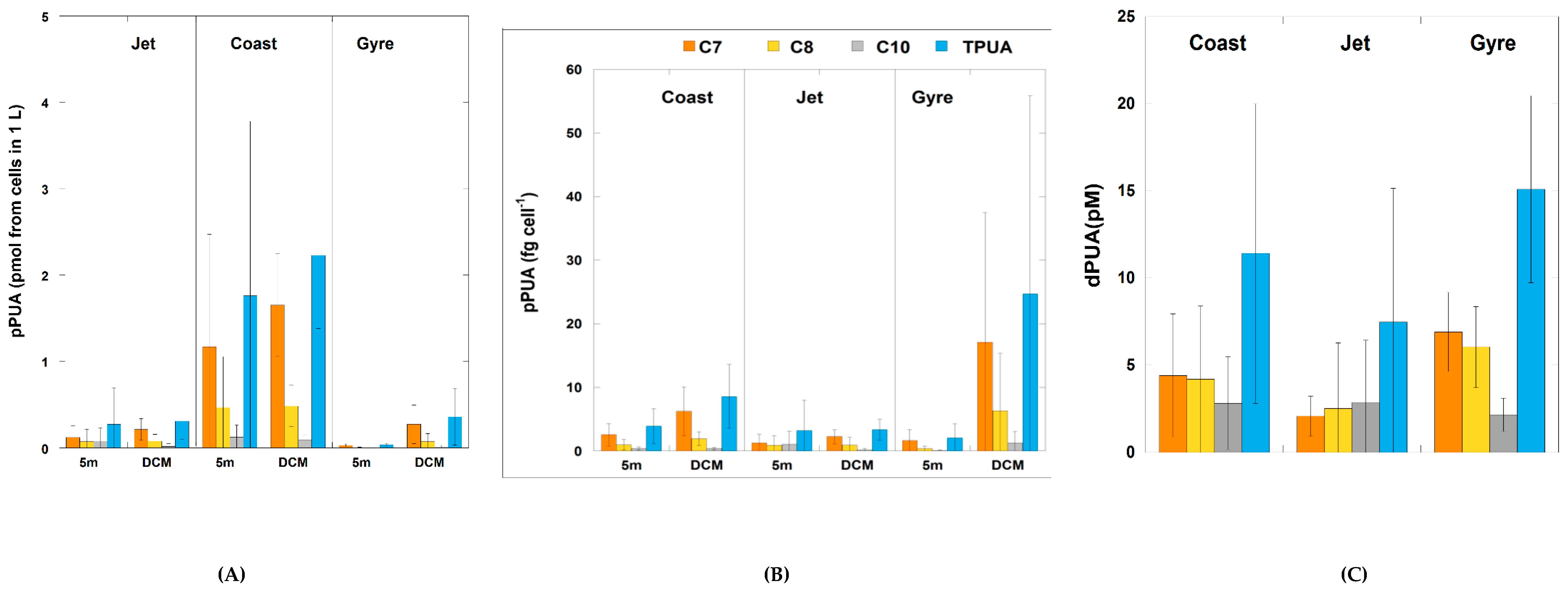
2.2. Polyunsaturated Aldehydes
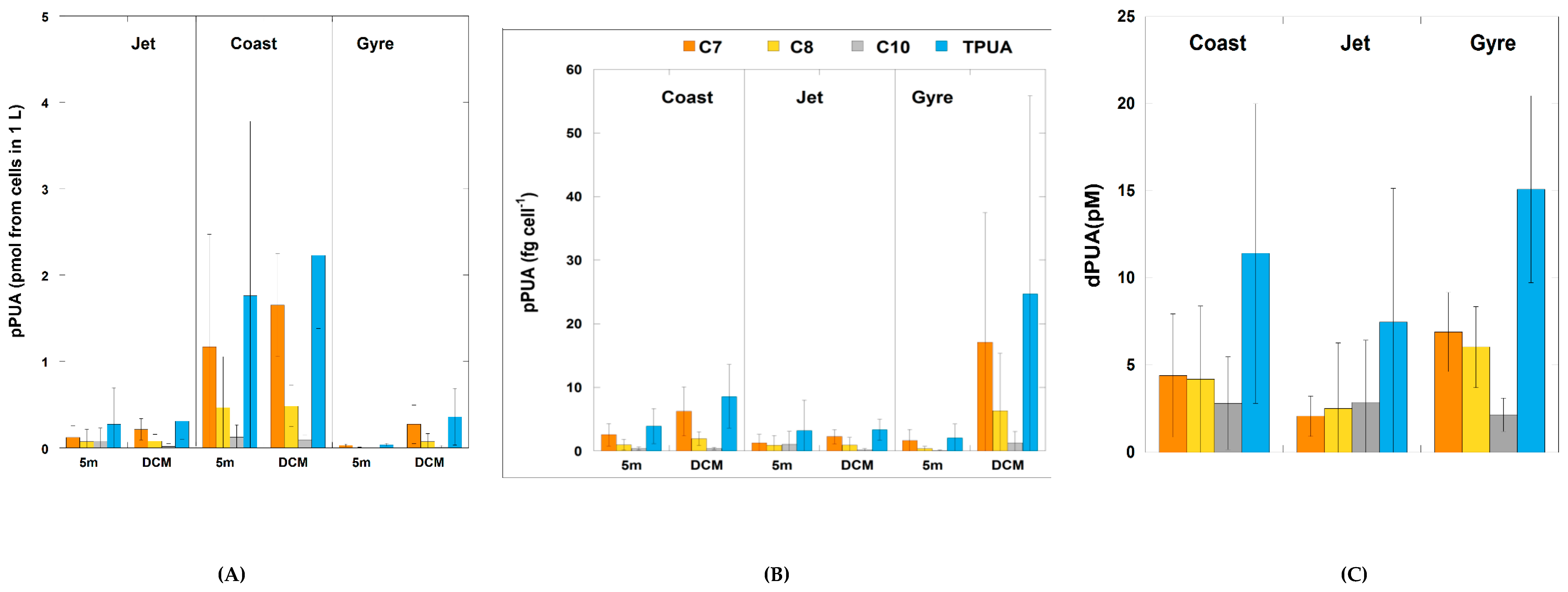
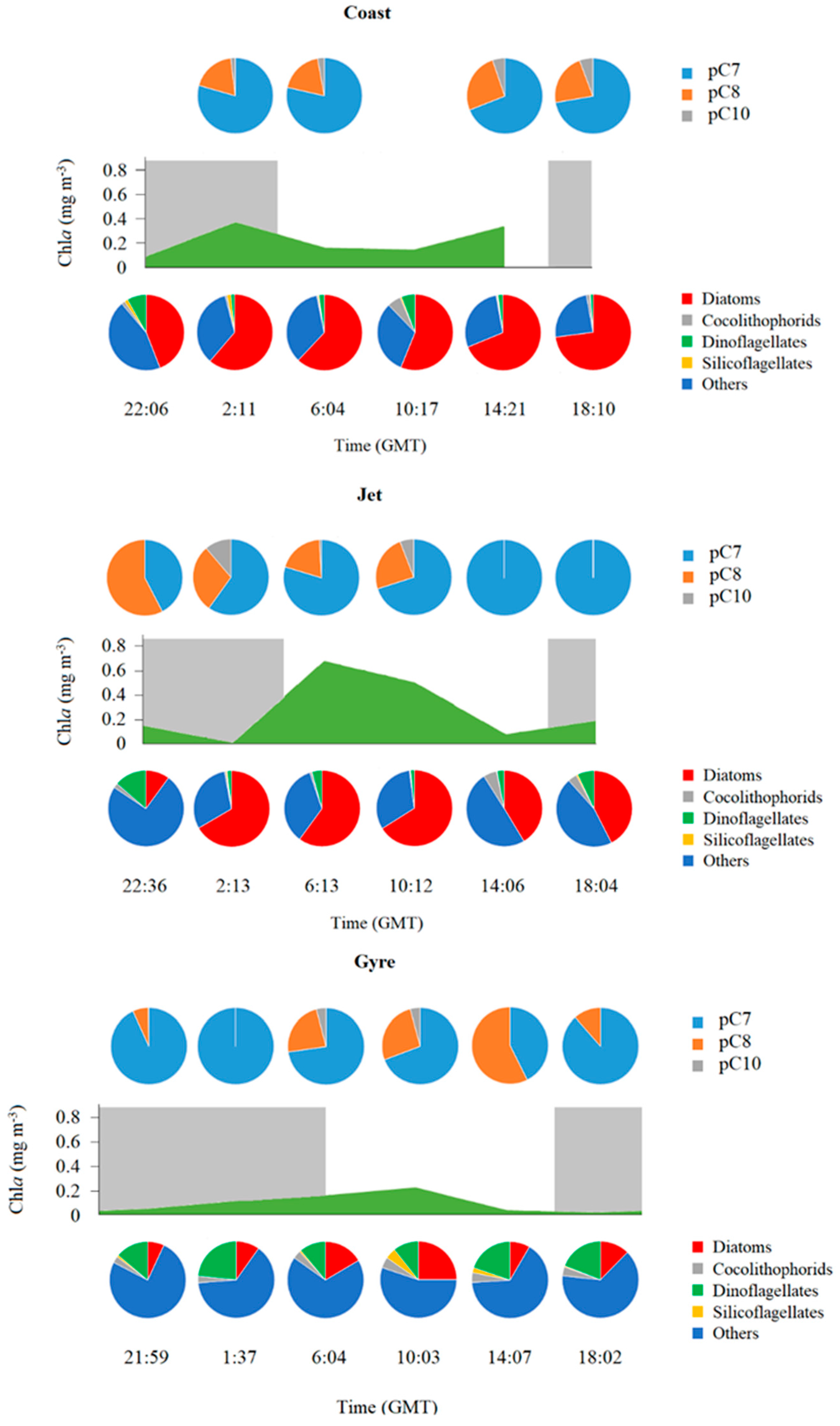
2.2.1. Particulate PUA (pPUA > 10 μm): Type and Distribution
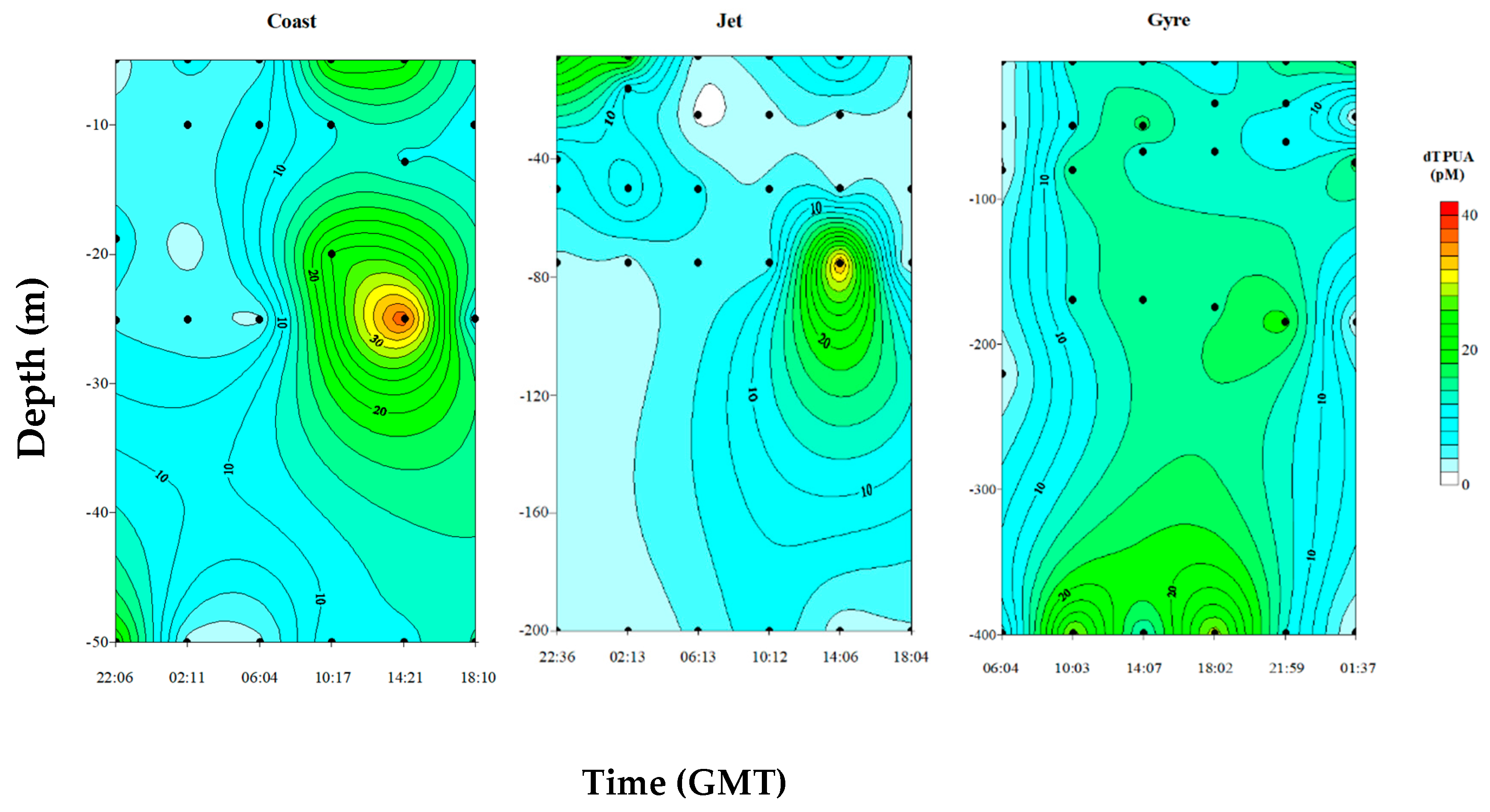
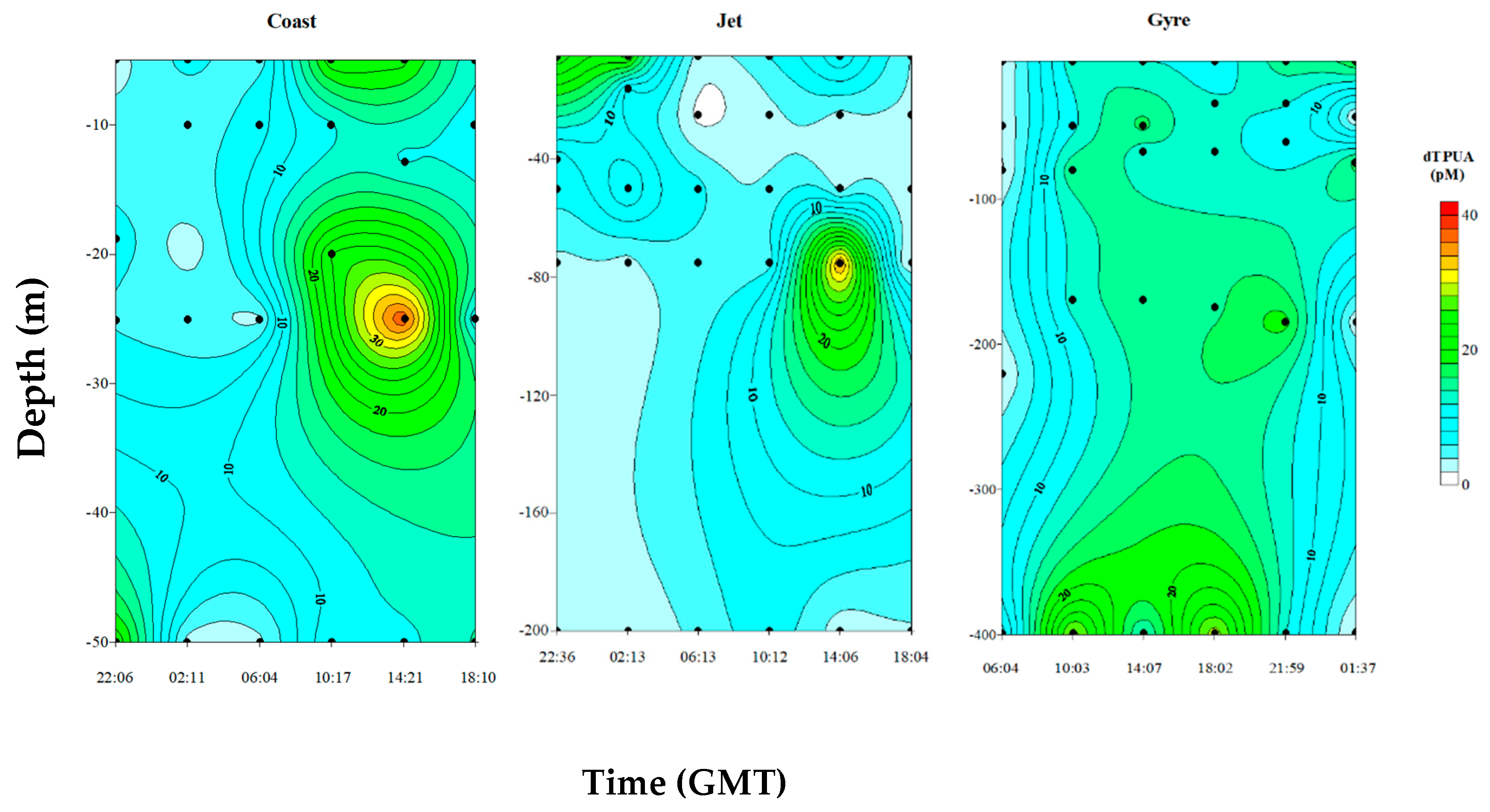
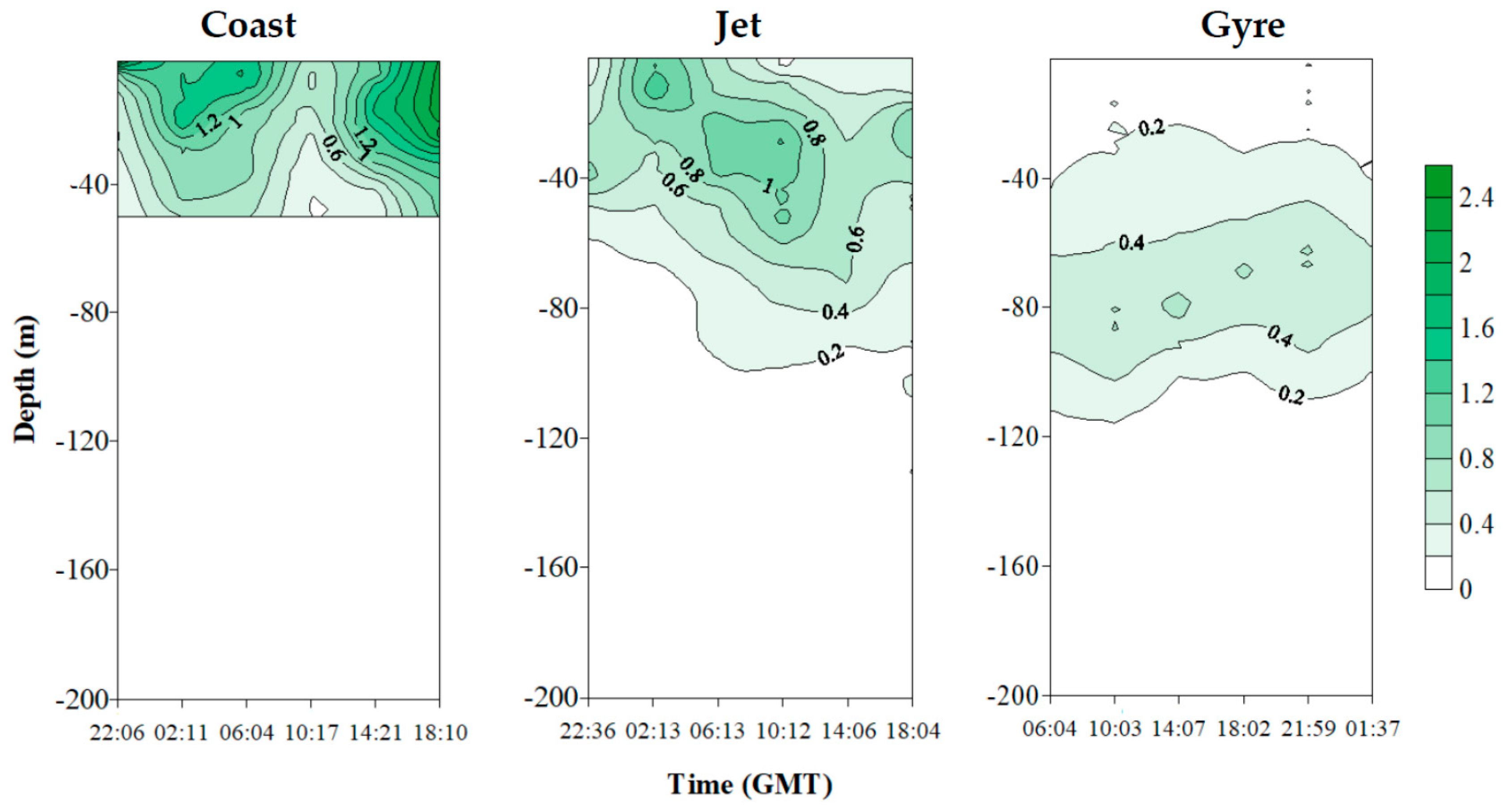
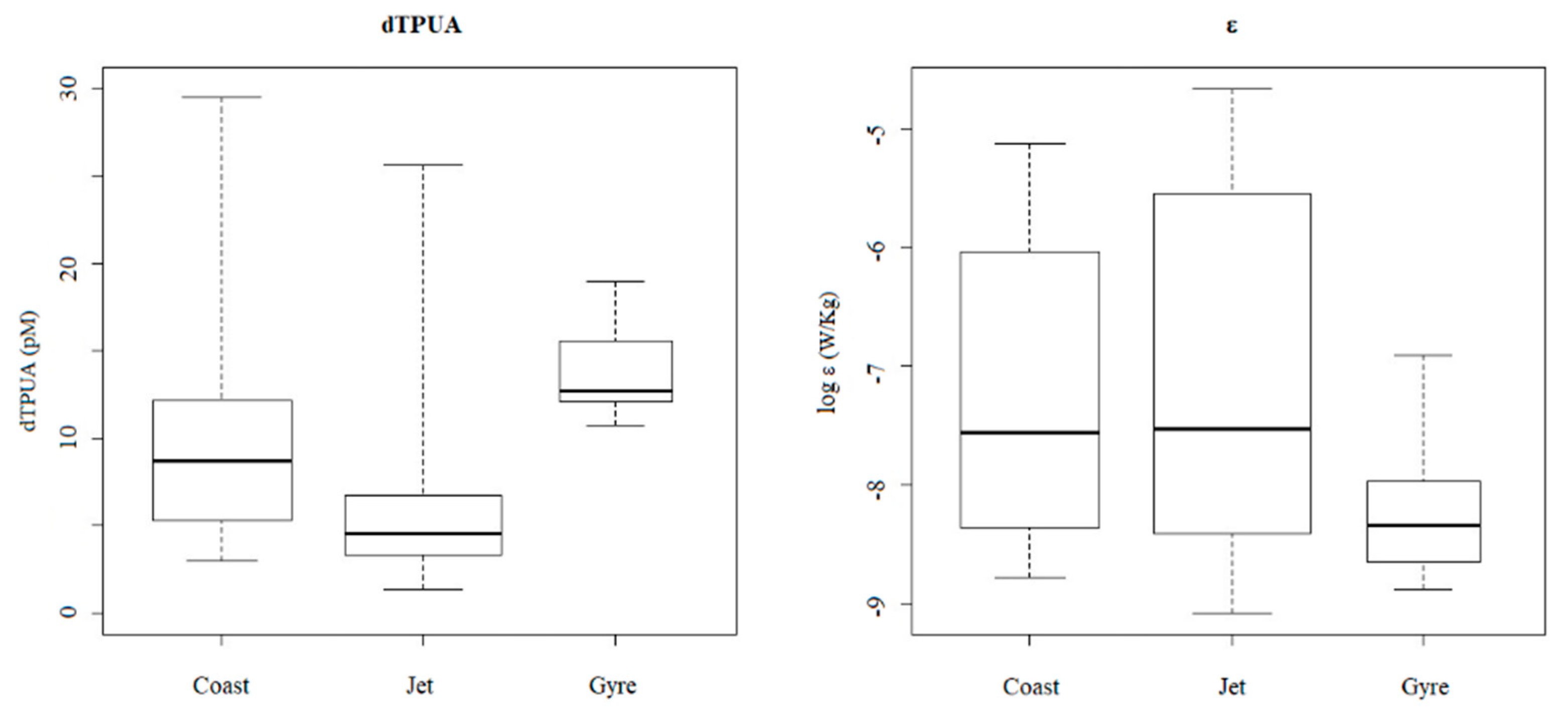
2.2.2. Dissolved PUA (dPUA): Types and Distribution
2.3. Chlorophyll Fluorescence and Pigments Concentrations
2.4. Phytoplankton Composition
2.4.1. Small-Size (<10 μm) Phytoplankton Abundance and Biovolume
2.4.2. Large-Size (10–250 μm) Phytoplankton Abundance and Biovolume
2.4.3. Total Phytoplankton Biovolume Partitioning
2.4.4. Phytoplankton Physiological State: Quantum Yield (Fv/Fm) and Percentage of Active Chlorophyll (%AChla)
2.5. Correlations among Polyunsaturated Aldehydes and Physicochemical and Biological Variables
3. Discussion
4. Materials and Methods
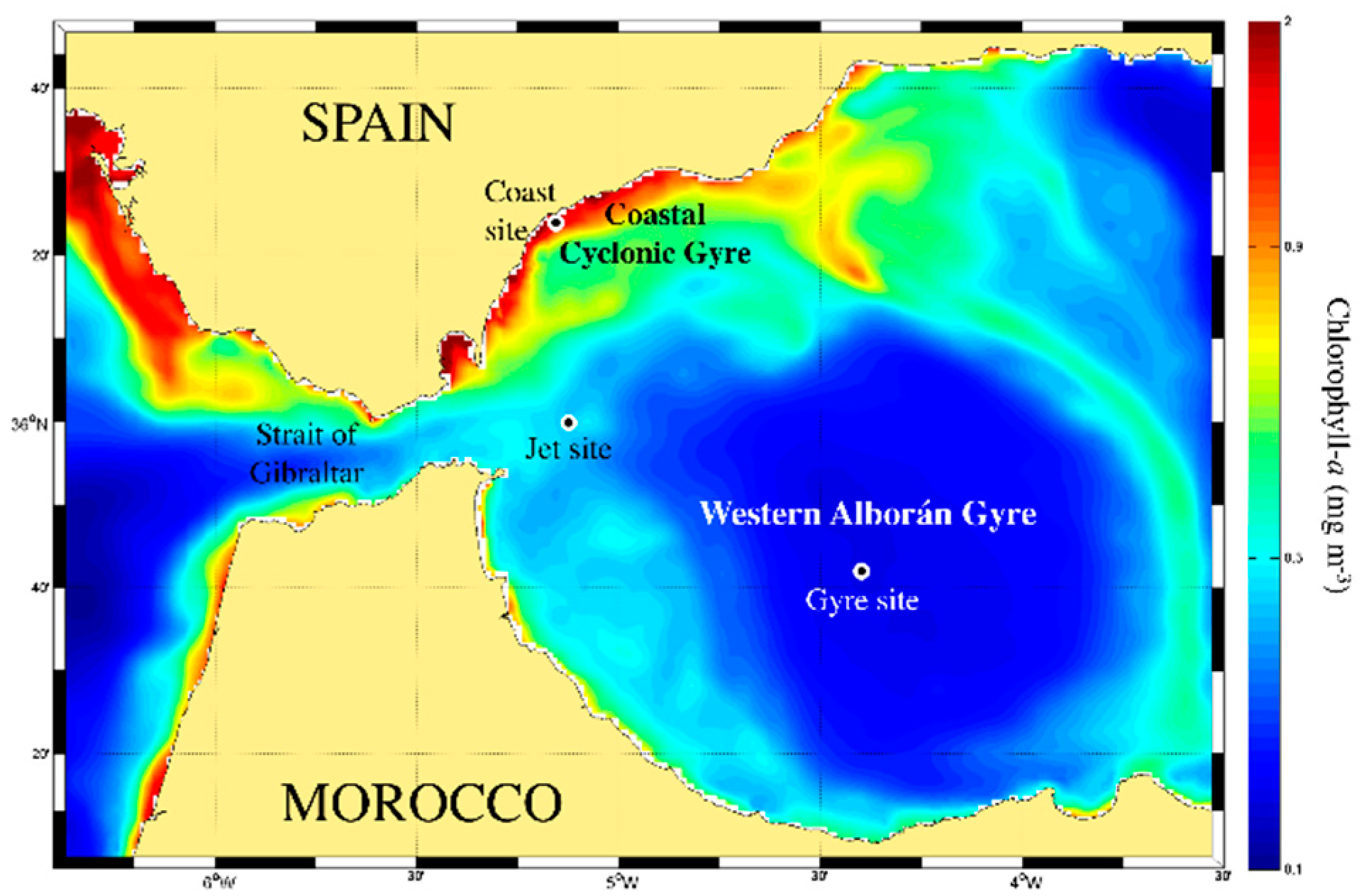
4.1. Study Area and Field Sampling
4.2. Physicochemical Analysis
4.3. Biological Variables
4.3.1. Pigment Quantification by High-Performance Liquid Chromatography (HPLC)
4.3.2. Calculation of Trophic Index (Fp)
4.3.3. Active Chlorophyll and Maximal PSII Quantum Yield
4.3.4. Phytoplankton Community Analyses
Small-Size Phytoplankton Analyses by Flow Cytometry
Large-Size Phytoplankton Analysis (10–250 μm) by Fluid Imaging FlowCAM®
4.3.5. Sampling and Extraction of Polyunsaturated Aldehydes (pPUA and dPUA)
4.3.6. pPUA and dPUA Quantification
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gardner, H.W. Recent investigations into the lypoxigenase pathway of plants. Biochim. Biophys. Acta 1991, 1084, 221–236. [Google Scholar] [CrossRef]
- Hamberg, M.; Ponce de León, I.; Rodríguez, M.J.; Castresana, C. Fatty acid alpha-dioxygenases. Prostaglandins Other Lipid Mediat. 2002, 338, 169–174. [Google Scholar] [CrossRef]
- Andreou, A.; Brodhun, F.; Feussner, I. Biosynthesis of oxilipins in no mammals. Prog. Lipid. Res. 2009, 48, 148–170. [Google Scholar] [CrossRef] [PubMed]
- Mosblech, A.; Fussner, I.; Heilmann, I. Oxylipins: Structurally diverse metabolites from fatty acid oxidation. Plant Physiol. Biochem. 2009, 47, 511–517. [Google Scholar] [CrossRef]
- Fontana, A.; d’Ippolito, G.; Cutignano, A.; Miralto, A.; Ianora, A.; Romano, G.; Cimino, G. Chemistry of oxylipin pathways in marine diatoms. Pure Appl. Chem. 2007, 79, 481–490. [Google Scholar] [CrossRef]
- Rettner, J.; Werner, M.; Meyer, N.; Werz, O.; Pohnert, G. Survey of the C20 and C22 oxylipin family in marine diatoms. Tetrahedron Lett. 2018, 59, 828–831. [Google Scholar] [CrossRef]
- Armbrust, E.V. The life of diatoms in the world’s oceans. Nature 2009, 459, 185–192. [Google Scholar] [CrossRef]
- Miralto, A.; Barone, G.; Romano, G.; Poulet, S.A.; Ianora, A.; Ruso, G.L.; Buttino, I.; Mazzarella, G.; Laabir, M.; Cabrini, M.; et al. The insiduous effect of diatoms on copepod reproduction. Nature 1999, 402, 173–176. [Google Scholar] [CrossRef]
- Van Donk, E.; Ianora, A.; Vos, M. Induced defences in marine and freshwater phytoplankton: A review. Hydrobiologia 2011, 668, 3–19. [Google Scholar] [CrossRef] [Green Version]
- Catalá, A. Five decades with polyunsaturated fatty acids: Chemical synthesis, enzymatic formation, lipid peroxidation and its biological effects. J. Lipids 2013, 2013, 710290. [Google Scholar] [CrossRef] [Green Version]
- Pohnert, G. Diatom/copepod interactions in plankton: The indirect chemical defence of unicellular algae. Chem.Bio.Chem. 2005, 6, 946–959. [Google Scholar] [CrossRef] [PubMed]
- Romano, G.; Miralto, A.; Ianora, A. Teratogenic effects of diatom metabolites on sea urchin Paracentrotus lividus embryos. Mar. Drugs 2010, 8, 950–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzè, G.; Pierson, J.J.; Stoecker, D.K.; Lavrentyev, P.J. Diatom-produced allelochemicals trigger trophic cascades in the planktonic food web. Limnol. Oceanogr. 2018, 63, 1093–1108. [Google Scholar] [CrossRef] [Green Version]
- Balestra, C.; Alonso-Sáez, L.; Gasol, J.M.; Casotti, R. Group-specific effects on coastal bacterioplankton of polyunsaturated aldehydes produced by diatoms. Aquat. Microb. Ecol. 2011, 63, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Paul, C.; Reunamo, A.; Lindehoff, E.; Bergkvist, J.; Mausz, M.A.; Larsson, H.; Richter, H.; Wängberg, S.-A.; Leskinen, P.; Båmstedt, U.; et al. Diatom derived polyunsaturated aldehydes do not structure the planktonic microbial community in a mesocosm study. Mar. Drugs 2012, 10, 775–792. [Google Scholar] [CrossRef]
- Edwards, B.R.; Bidle, K.D.; Van Mooy, B.A.S. Dose-dependent regulation of microbial activity on sinking particles by polyunsaturated aldehydes: Implications for the carbon cycle. Proc. Natl. Acad. Sci. USA 2015, 112, 5909–5914. [Google Scholar] [CrossRef] [Green Version]
- Lavrentyev, P.J.; Franze, G.; Pierson, J.J.; Stoecker, D.K. The effect of dissolved polyunsaturated aldehydes on microzooplankton growth rates in the Chesapeake Bay and Atlantic Coastal Waters. Mar. Drugs 2015, 13, 2834–2856. [Google Scholar] [CrossRef] [Green Version]
- Pepi, M.; Heipieper, H.J.; Balestra, C.; Borra, M.; Biffali, E.; Casotti, R. Toxicity of diatom polyunsaturated aldehydes to marine bacterial isolates reveals their mode of action. Chemosphere 2017, 177, 258–265. [Google Scholar] [CrossRef]
- Varrella, S.; Romano, G.; Costantini, S.; Ruocco, N.; Ianora, A.; Bentley, M.G.; Costantini, M. Toxic diatom aldehydes affect defence gene networks in sea urchins. PLoS ONE 2016, 11, e0149734. [Google Scholar] [CrossRef] [Green Version]
- Leflaive, J.; Ten-Hage, L. Chemical interactions in diatoms: Role of polyunsaturated aldehydes and precursors. New Phytol. 2009, 184, 794–805. [Google Scholar] [CrossRef]
- Strandberg, U.; Hiltunen, M.; Jelkänen, E.; Taipale, S.J.; Kainz, M.J.; Brett, M.T.; Kankaala, P. Selective transfer of polyunsaturated fatty acids from phytoplankton to planktivorous fish in large boreal lakes. Sci. Total Environ. 2015, 536, 858–865. [Google Scholar] [CrossRef] [PubMed]
- Adolph, S.; Bach, S.; Blondel, M.; Cueff, A.; Moreau, M.; Pohnert, G.; Poulet, S.A.; Wichard, T.; Zuccaro, A. Cytotoxicity of diatom-derived oxylipins in organisms belonging to different phyla. J. Exp. Biol. 2004, 207, 2935–2946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ianora, A.; Poulet, S.A.; Miralto, A. The effects of diatoms on copepod reproduction: A review. Phycologia 2013, 42, 351–363. [Google Scholar] [CrossRef]
- Ianora, A.; Miralto, A. Toxigenic effects of diatoms on grazers, phytoplankton and other microbes: A review. Ecotoxicology 2010, 19, 493–511. [Google Scholar] [CrossRef] [PubMed]
- Vidoudez, C.; Casotti, R.; Basanini, M.; Pohnert, G. Quantification of dissolved and particulate polyunsaturated aldehydes in the Adriatic Sea. Mar. Drugs 2011, 9, 500–513. [Google Scholar] [CrossRef]
- Vidoudez, C.; Nejstgaard, J.C.; Jakobsen, H.H.; Pohnert, G. Dynamics of dissolved and particulate polyunsaturated aldehydes in mesocosms inoculated with different densities of the diatom Skeletonema marinoi. Mar. Drugs 2011, 9, 345–358. [Google Scholar] [CrossRef] [Green Version]
- Morillo-García, S.; Valcárcel-Pérez, N.; Cózar, A.; Ortega, M.J.; Macías, D.; Ramírez-Romero, E.; García, C.M.; Echevarría, F.; Bartual, A. Potential polyunsaturated aldehydes in the Strait of Gibraltar under two tidal regimes. Mar. Drugs 2014, 12, 1438–1459. [Google Scholar] [CrossRef] [Green Version]
- Bartual, A.; Morillo-García, S.; Ortega, M.J.; Cózar, A. First report on vertical distribution of dissolved polyunsaturated aldehydes in marine coastal waters. Mar. Chem. 2018, 204, 1–10. [Google Scholar] [CrossRef]
- Vardi, A.; Formiggini, F.; Casotti, R.; De Martino, A.; Ribalet, F.; Miralto, A.; Bowler, C. A stress surveillance system based on calcium and nitric oxide in marine diatoms. PLoS Biol. 2006, 4, 411–419. [Google Scholar] [CrossRef] [Green Version]
- Cózar, A.; Morillo-García, S.; Ortega, M.J.; Li, Q.P.; Bartual, A. Macroecological patterns of the phytoplankton production of polyunsaturated aldehydes. Sci. Rep. 2018, 8, 12282. [Google Scholar] [CrossRef]
- Powley, H.R.; Krom, M.D.; Van Cappellen, P. Understanding the unique biogeochemistry of the Mediterranean Sea: Insights from a coupled phosphorus and nitrogen model. Glob. Biogeochem. Cycles 2017, 31, 1010–1031. [Google Scholar] [CrossRef]
- Macías, D.; García-Gorriz, E.; Piroddi, C.; Stips, A. Biogeochemical control of marine productivity in the Mediterranean Sea during the last 50 years. Glob. Biogeochem. Cycles 2014, 28, 897–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macías, D.; García-Gorriz, E.; Stips, A. Major fertilization sources and mechanisms for Mediterranean Sea coastal ecosystems. Limnol. Oceanogr. 2018, 63, 897–914. [Google Scholar] [CrossRef] [Green Version]
- Echevarría, F.; García Lafuente, J.; Bruno, M.; Gorsky, G.; Goutx, M.; González, N.; García, C.M.; Gómez, F.; Vargas, J.M.; Picheral, M.; et al. Physical-biological coupling in the Strait of Gibraltar. Deep-Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 4115–4130. [Google Scholar] [CrossRef]
- Claustre, H. The trophic status of various oceanic provinces as revealed by phytoplankton pigment signatures. Limnol. Oceanogr. 1994, 39, 1206–1210. [Google Scholar] [CrossRef]
- Adolph, S.; Poulte, S.; Pohnert, G. Synthesis and biological activity of α,β,γ,δ-unsaturated aldehydes from diatoms. Tetrahedron 2003, 59, 3003–3008. [Google Scholar] [CrossRef]
- Jeffrey, S.W.; Vest, M. Introduction to marine phytoplankton and their pigment signatures. In Phytoplankton Pigments in Oceanography; Jeffrey, S.W., Mantoura, R.F.C., Wright, S.W., Eds.; UNESCO: Paris, France, 1997; pp. 37–84. [Google Scholar]
- Kheireddine, M.; Ouhssain, M.; Claustre, H.; Uitz, J.; Gentili, B.; Jones, B.H. Assessing pigment-based phytoplankton community distributions in the Red Sea. Front. Mar. Sci. 2017, 4, 132. [Google Scholar] [CrossRef] [Green Version]
- Uitz, J.; Claustre, H.; Morel, A.; Hooker, S.B. Vertical distribution of phytoplankton communities in open-ocean: An assessment based on surface chlorophyll. J. Geophys. Res. 2006, 111, C08005. [Google Scholar] [CrossRef]
- Raateoja, M.; SeppäläJukka, J.; Ylöstalo, P. Fast repetition rate fluorometry is not applicable to studies of filamentous cyanobacteria from the Baltic Sea. Limnol. Oceanogr. 2004, 49, 1006–1012. [Google Scholar] [CrossRef]
- Sugget, D.J.; Moore, M.; Hickman, A.E.; Geider, R.J. Interpretation of fast repetition rate (FRR) fluorescence: Signatures of phytoplankton community structure versus physiological state. Mar. Ecol. Prog. Ser. 2009, 376, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Romero-Martínez, L.; Van Slooten, C.; Nebot, E.; Acevedo-Merino, A.; Peperzak, L. Assessment of imaging-in-flow system (FlowCAM) for systematic ballast water management. Sci. Total Environ. 2017, 603–604, 550–561. [Google Scholar] [CrossRef] [PubMed]
- Franklin, D.J.; Choi, C.J.; Hughes, C.; Malin, G.; Berges, J.A. Effect of dead phytoplankton cells on the apparent efficiency of photosystem II. Mar. Ecol. Prog. Ser. 2009, 382, 35–40. [Google Scholar] [CrossRef]
- Franklin, D.J.; Brussaard, C.P.D.; Berges, J.A. What is the role and nature of programmed cell death in phytoplankton ecology? Eur. J. Phycol. 2006, 41, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.C.; Gong, G.C.; Chang, J. Lateral water exchange between shelf-margin upwelling and Kuroshio waters influences phosphorus stress in microphytoplankton. Mar. Ecol. Prog. Ser. 2010, 409, 121–130. [Google Scholar] [CrossRef]
- Qi, H.; Wang, J.; Wang, Z. A comparative study of the sensitivity of Fv/Fm to phosphorus limitation on four marine algae. J. Ocean Univ. China. 2013, 12, 77–84. [Google Scholar] [CrossRef]
- Wu, Z.; Li, Q. Spatial distributions of polyunsaturated aldehydes and their biogeochemical implications in the Pearl River Estuary and the adjacent northern South China Sea. Prog. Oceanogr. 2016, 147, 1–9. [Google Scholar] [CrossRef]
- Bartual, A.; Arandia-Gorostidi, N.; Cozar, A.; Morillo-Garcia, S.; Ortega, M.J.; Vidal, M.; Cabello, A.M.; Gonzalez-Gordillo, J.I.; Echevarría, F. Polyunsaturated aldehydes from large phytoplankton of the Atlantic Ocean surface (42°N to 33°S). Mar. Drugs 2014, 12, 682–699. [Google Scholar] [CrossRef] [Green Version]
- Ribalet, F.; Wichard, T.; Pohnert, G.; Ianora, A.; Miralto, A.; Casotti, R. Age and nutrient limitation enhance polyunsaturated aldehyde production in marine diatoms. Phytochemistry 2007, 68, 2059–2067. [Google Scholar] [CrossRef]
- Brembu, T.; Mühlroth, A.; Alipanah, L.; Bones, A.M. The effects of phosphorus limitation on carbon metabolism in diatoms. Phil. Trans. R. Soc. 2017, 372, 20160406. [Google Scholar] [CrossRef]
- Reitan, K.I.; Rainuzzo, J.R.; Olsen, Y. Effect of nutrient limitation on fatty acid and lipid content of marine microalgae. J. Phycol. 1994, 30, 972–979. [Google Scholar] [CrossRef]
- Wichard, T.; Poulet, S.A.; Pohnert, G. Determination and quantification of α,β,γ,δ-unsaturated aldehydes as pentafluorobenzyl-oxime derivates in diatom cultures and natural phytoplankton populations: Application in marine field studies. J. Chromatogr. B 2005, 814, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Hansen, E.; Eilertsen, H.C. Do the polyunsaturated aldehydes produced by Phaeocystis pouchetii (Hariot) Lagerheim influence diatom growth during the spring bloom in Northern Norway? J. Plank. Res. 2007, 29, 87–96. [Google Scholar] [CrossRef]
- Agustí, S.; Duarte, C. Phytoplankton lysis predicts dissolved organic carbon release in marine plankton communities. Biogeosciences 2013, 10, 1259–1264. [Google Scholar] [CrossRef] [Green Version]
- Ribalet, F.; Bastianini, M.; Vidoudez, C.; Acri, F.; Berges, J.; Ianora, A.; Miralto, A.; Pohnert, G.; Romano, G.; Wichard, T.; et al. Phytoplankton cell lysis associated with polyunsaturated aldehyde release in the Northern Adriatic Sea. PLoS ONE 2014, 9, e98727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartual, A.; Ortega, M.J. Temperature differentially affects the persistence of polyunsaturated aldehydes in seawater. Environ. Chem. 2013, 10, 403–408. [Google Scholar] [CrossRef]
- Satchwill, T.; Watson, S.B.; Dixon, E. Odorous algal-derived alkenes: Differences in stability and treatment responses in drinking water. Water Sci. Tech. 2007, 55, 95–102. [Google Scholar] [CrossRef]
- Romano, C.; Fiorito, D.; Mazet, C. Remote functionalization of a,b-unsaturated carbonyls by multimetallic sequential catalysis. J. Am. Chem. Soc. 2019, 141, 16983–16990. [Google Scholar] [CrossRef]
- Verhulst, A.; Janssen, G.; Parmentier, G.; Eyssen, H. Isomerization of polyunsaturated long chain fatty acids by propionibacteria. Syst. Appl. Microbiol. 1987, 9, 12–15. [Google Scholar] [CrossRef]
- Heipieper, H.J.; Meinhardt, F.; Segura, A. The cis-trans isomerase of unsaturated fatty acids in Pseudomonas and Vibrio: Biochemistry, molecular biology and physiological function of a unique stress adaptive mechanism. FEMS Microbiol. Lett. 2003, 229, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Arnosti, C.; Bell, C.; Moorhead, D.L.; Sinsabaugh, R.L.; Steen, A.D.; Stromberger, M.; Wallenstein, M.; Weintraub, M.N. Extracellular enzymes in terrestrial, freshwater and marine environments: Perspectives on system variability and common research needs. Biogeochemistry 2014, 117, 5–21. [Google Scholar] [CrossRef]
- Armi, L.; Farmer, D. The flow of Mediterranean water through the Strait of Gibraltar. Prog. Oceanog. 1988, 21, 3–41, 43–47, 49, 51, 53, 55, 57–63, 65–67, 69, 71, 73, 75–77, 79, 81–89, 91–93, 95, 97–101, 103. [Google Scholar] [CrossRef]
- Gascard, J.C.; Richez, C. Water masses and circulation in the western Alboran Sea and in the Strait of Gibraltar. Prog. Oceanog. 1985, 15, 157–216. [Google Scholar] [CrossRef]
- Gómez, F.; Gorsky, G.; Striby, L.; Vargas, J.M.; Gonzalez, N.; Picheral, M.; García-Lafuente, J.; Varela, M.; Goutx, M. Small-scale temporal variations in biogeochemical features in the Strait of Gibraltar, Mediterranean side-the role of NACW and the interface oscillation. J. Mar. Sys. 2001, 30, 207–220. [Google Scholar] [CrossRef]
- Bormans, M.; Garrett, C. A simple criterion for gyre formation by the surface outflow from a strait, with application to the Alboran Sea. J. Geophys. Res. 1989, 94, 12637–12644. [Google Scholar] [CrossRef]
- Renault, L.; Oguz, T.; Pascual, A.; Vizoso, G.; Tintore, J. Surface circulation in the Alboran Sea (western Mediterranean) inferred from remotely sensed data. J. Geophys. Res. 2012, 117, C08009. [Google Scholar] [CrossRef] [Green Version]
- Viúdez, A.; Haney, R.L.; Tintoré, J. Circulation in the Alboran Sea as determined by quasi-synoptic hydrographic observations. Part II: Mesoscale ageostrophic motion diagnosed through density dynamical assimilation. J. Phys. Oceanogr. 1996, 26, 706–724. [Google Scholar] [CrossRef] [Green Version]
- Sarhan, T.; García-Lafuente, J.; Vargas, M.; Vargas, J.M.; Plaza, F. Upwelling mechanisms in the northwestern Alboran Sea. J. Mar. Syst. 2000, 23, 317–331. [Google Scholar] [CrossRef]
- Macías, D.; Bruno, M.; Echevarría, F.; Vázquez, A.; García, C.M. Metereologically-induced mesoscale variability of the north-western Alboran Sea (southern Spain) and related biological patterns. Estuar. Coast. Shelf. Sci. 2008, 78, 250–266. [Google Scholar] [CrossRef] [Green Version]
- Bruno, M.; Chioua, J.; Romero, V.A.; Macías, D.; Dastis, C.; Ramírez-Romero, E.; Echevarria, F.; Reyes, J.; García, C.M. The importance of sub-mesoscale processes for the exchange of properties through the Strait of Gibraltar. Prog. Oceanog. 2013, 116, 66–79. [Google Scholar] [CrossRef] [Green Version]
- Strickland, J.D.H.; Parsons, T.R. A Practical Handbook of Seawater Analysis, 2nd ed.; Fisheries Research Board of Canada: Ottawa, ON, Canada, 1972; pp. 1–310. [Google Scholar]
- Zapata, M.; Rodriguez, F.; Garrido, J.L. Separation of chlorophylls and carotenoids from marine phytoplankton: A new HPLC method using a reversed phase C8 column and pyridine containing mobile phases. Mar. Ecol. Prog. Ser. 2000, 195, 29–45. [Google Scholar] [CrossRef] [Green Version]
- Jeffrey, S.W. Cultivating unicellular marine plants. CSIRO (Hobart, Tasmania). Div. Fish Oceanogr. Res. Rep. 1980, 1977–1979, 22–43. [Google Scholar]
- Wright, S.W.; Jeffrey, S.W. Fucoxanthin pigment markers of marine phytoplankton analysed by HPLC and HPTLC. Mar. Ecol. Prog. Ser. 1987, 38, 259–266. [Google Scholar] [CrossRef]
- Gieskes, W.W.C.; Kraay, G.W. Dominance of Cryptophyceae during the phytoplankton spring bloom in the central North Sea in 1983. Mar. Biol. 1983, 75, 179–185. [Google Scholar] [CrossRef]
- Jakob, T.; Schreiber, U.; Kirchesch, V.; Wilhelm, C. Estimation of chlorophyll content and daily primary production of the major algal groups by means of multiwavelength-excitation PAM chlorophyll fluorometry: Performance and methodological limits. Phot. Res. 2005, 83, 343–361. [Google Scholar] [CrossRef] [PubMed]
- Gasol, J.M.; Morán, X.A.G. Effects of filtration on bacterial activity and picoplankton community structure as assessed by flow cytometry. Aquatic. Microb. Ecol. 1999, 16, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, E.; Moyano, M.; López-Urrutia, A.; Nogueira, E.; Scharek, R. Routine determination of planktoncommunity composition and size structure: A comparison between FlowCAM and light microscopy. J. Plankton Res. 2014, 36, 170–184. [Google Scholar] [CrossRef] [Green Version]
- Mender-Deuer, S.; Lessard, E.; Satterberg, J. Effect of preservation on dinoflagellate and diatom cell volume and consequences for carbon biomass predictions. Mar. Ecol. Progress Ser. 2001, 222, 41–50. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
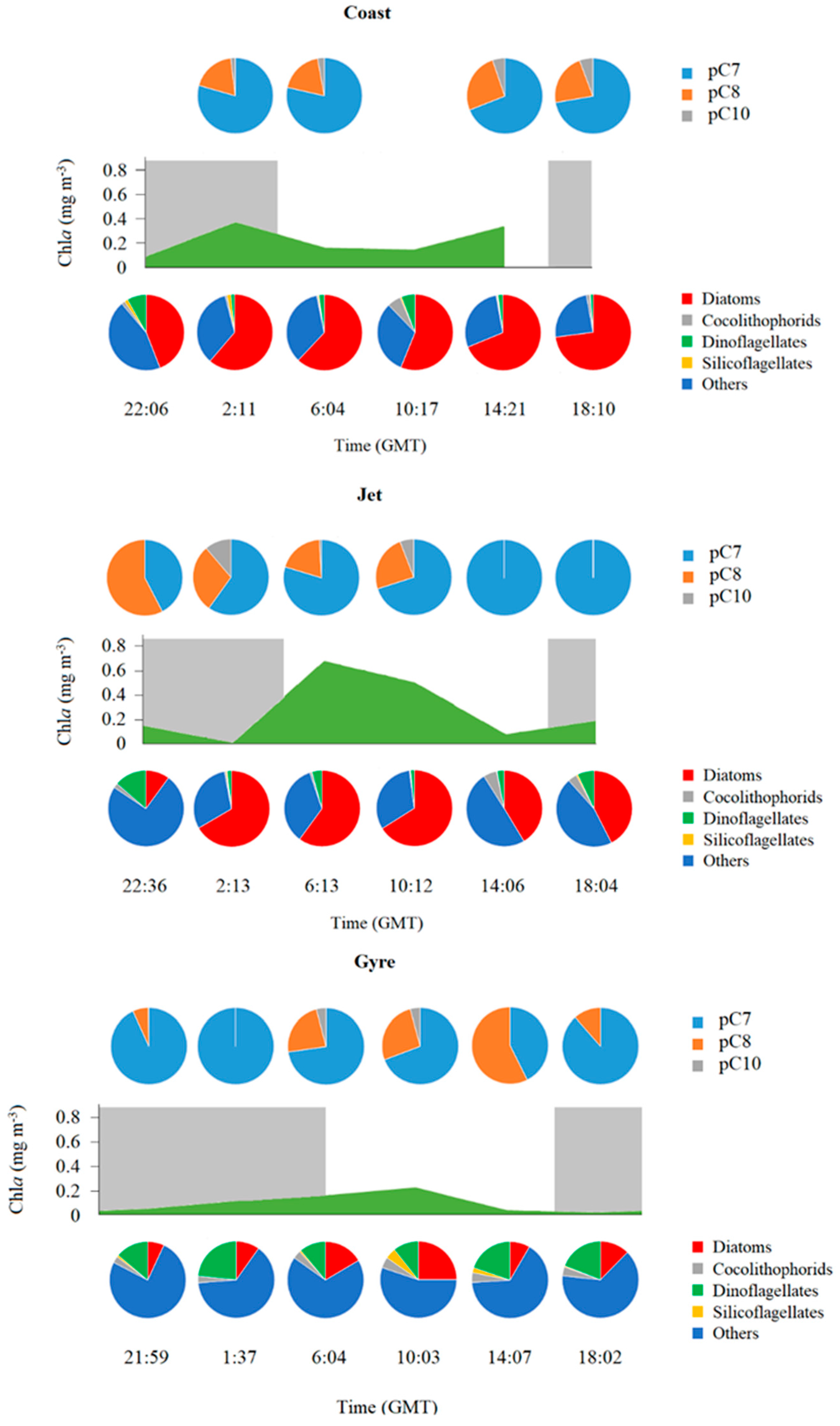{kind=link}
| Coast (0–50 m) | Jet (0–200 m) | Gyre (0–400 m) | |
|---|---|---|---|
| Trophic index (Fp) | 0.77 ± 0.07 [0.9–0.63] | 0.073 ± 0.095 [0.28–0.01] | 0.13 ± 0.14 [0.44–0] |
| Temperature (°C) | 16.31 ± 1.17 [14.08–17.74] | 16.56 ± 2.18 [13.33–20.75] | 18.75 ± 4.16 [13.37–23.27] |
| Salinity | 37.06 ± 0.33 [36.56–37.77] | 36.69 ± 0.75 [36.22–38.27] | 37.11±0.79[36.43–38.5] |
| NOx (μM) | 2.78 ± 1.79 [6.1–0.3] | 2.94 ± 2.32 [7.83–0.17] | 2.74 ± 3.52 [9.89–0.05] |
| PO43− (μM) | 0.22 ± 0.11 [0.34–0.01] | 0.35 ± 0.43 [2.42–0] | 0.16 ± 0.16 [0.45–0] |
| SiO4 (μM) | 2.99 ± 1.39 [7.9–1.4] | 2.68 ± 1.97 [9.39–0.62] | 2.27 ± 2.26 [7.02–0.15] |
| NOx/PO43− | 17.8 ± 16.2 [56.8–1.69] | 10.71 ± 7.57 [24.96–1.02] | 25.51 ± 39.45 [175–0.6] |
| SiO4/NOx | 1.76 ± 1.57 [7.18–0.72] | 2.3 ± 3.7 [16.80–0.45] | 3.99 ± 6.94 [37.7–0.43] |
| Epsilon, ε (m2 s−3) | Coast | Jet | Gyre | |
|---|---|---|---|---|
| Depth averaged (5–200 m) | 1.06 10−6 ± 2.07 10−6 [1.63 10−9–7.44 10−6] | 3.25 10−6–6.05 10−6 [8.24 10−10 ± 2.18 10−5] | 1.71 10−8 ± 3.43 10−8 [1.32 10−9–1.22 10−7] | |
| Daily averaged (5 m) | 3.16 10−6 ± 2.6 10−6 | 1.21 10−5 ± 5.8 10−6 | - | |
| Daily averaged (25 m) | 1.12 10−8 ± 1.25 10−8 | 1.84 10−6 ± 3.65 10−6 | 6.032 10−9 ± 6.54 10−9 | |
| Source of variation | df | MS | F | p |
| site | 2 | 6.37 10−11 | 4.835 | 0.017 |
| Depth | 1 | 3.33 10−10 | 25.27 | 0.00039 |
| Site × depth | 1 | 2.89 10−10 | 21.99 | 0.000091 |
| pC7 | pC8 | pC10 | |
| pC7 | 1 | ||
| pC8 | 0.69 * | 1 | |
| pC10 | 0.49 | 0.76 * | 1 |
| dC7 | dC8 | dC10 | |
| dC7 | 1 | ||
| dC8 | 0.85 * | 1 | |
| dC10 | 0.25 | 0.39 * | 1 |
| pC7-Type I | pC7-Type II | pC8-Type I | pC8-Type II | pC10-Type I | pC10-Type II | |
| Coast 5 m | 98.21 ± 1.11 | 1.79 ± 1.11 | 55.83 ± 11.82 | 44.17 ± 11.82 | 44.73 ± 17.45 | 55.27 ± 17.45 |
| Coast DCM | 98.72 ± 0.73 | 1.28 ± 0.73 | 63.45 ± 9.21 | 36.55 ± 9.21 | 32.91 ± 4.73 | 67.09 ± 4.73 |
| Jet 5 m | 99.20 ± 0.65 | 0.8 ± 0.65 | 17.59 ± 11.57 | 82.41 ± 11.57 | 73.69 ± 22.72 | 26.31 ± 22.72 |
| Jet DCM | 85.86 ± 31.40 | 1.11 ± 1.03 | 21.55 ± 11.73 | 80.83 ± 11.55 | 53.34 ± 28.31 | 46.66 ± 28.31 |
| Gyre 5 m | 92.27 ± 6.73 | 7.73 ± 6.73 | 25.51 ± 20.25 | 74.49 ± 20.25 | 58.64 ± 15 | 41.36 ± 15 |
| Gyre DCM | 98.57 ± 1.23 | 1.43 ± 1.23 | 14.24 ± 24.60 | 85.76 ± 24.60 | 65.51 ± 20.32 | 34.49 ± 20.32 |
| dC7-Type I | dC7-Type II | dC8-Type I | dC8-Type II | dC10-Type I | dC10-Type II | |
| Coast | 8.66 ± 4.92 | 91.34 ± 4.92 | 4.28 ± 2.14 | 95.72 ± 2.14 | 2.08 ± 4.42 | 97.92 ± 4.42 |
| Jet | 7.82 ± 4.13 | 92.18 ± 4.13 | 3.59 ± 1.47 | 96.41 ± 1.47 | 1.56 ± 0.89 | 98.44 ± 0.89 |
| Gyre | 3.32 ± 2.19 | 96.68 ± 2.19 | 5.41 ± 1.98 | 94.59 ± 1.98 | 0.90 ± 0.35 | 99.10 ± 0.35 |
| Coast | Jet | Gyre | ||||
|---|---|---|---|---|---|---|
| Depth | 5 m | DCM | 5 m | DCM | 5 m | DCM |
| Chlorophyll a | 0.311 ± 0.173 | 0.209 ± 0.125 | 0.220 ± 0.128 | 0.312 ± 0.260 | 0.047 ± 0.039 | 0.087 ± 0.079 |
| Chlorphyll b | 0.021 ± 0.005 | 0.011 ± 0.004 | 0.015 ± 0.007 | 0.032 ± 0.009 | 0.003 ± 0.002 | 0.029 ± 0.014 |
| Chlorophyll c | 0.054 ± 0.049 | 0.042 ± 0.049 | 0.017 ± 0.018 | 0.026 ± 0.027 | 0 | 0.002 ± 0.004 |
| Peridinine | 0.007 ± 0.013 | 0.002 ± 0.002 | 0.005 ± 0.006 | 0.002 ± 0.003 | 0.003 ± 0.003 | 0.003 ± 0.004 |
| 19’hex + 19’but | 0.0980 ± 0.0495 | 0.055 ± 0.062 | 0.087 ± 0.037 | 0.166 ± 0.050 | 0.038 ± 0.020 | 0.121 ± 0.060 |
| Ratio DT/Chla | 1.680 ± 0.209 | 1.870 ± 0.645 | 0.951 ± 0.376 | 0.859 ± 0.163 | 2.287 ± 2.289 | 0.687 ± 0.527 |
| Ratio 19’hex/Chla | 0.262 ± 0.065 | 0.224 ± 0.138 | 0.394 ± 0.194 | 0.587 ± 0.323 | 1.769 ± 1.669 | 1.948 ± 2.406 |
| Coast | Jet | Gyre | ||||
|---|---|---|---|---|---|---|
| Depth | 5 m | DCM | 5 m | DCM | 5 m | DCM |
| SPhA | 2493 × 103 ± 602.5 × 103 | 2105 × 103 ± 841.15 × 103 | 2571 × 103 ± 863 × 103 | 3068 × 103 ± 1649 × 103 | 710 × 103 ± 198.2 × 103 | 4451 × 103 ± 1180 × 103 |
| Synechococcus | 538 × 103 ± 206 × 103 | 438 × 103 ± 186 × 103 | 1170 × 103 ± 349 × 103 | 575 × 103 ± 174 × 103 | 501 × 103 ± 139 × 103 | 393 × 103 ± 188 × 103 |
| Prochlorococcus | 414 × 103 ± 62.4 × 103 | 447 × 103 ± 55.7 × 103 | 555 × 103 ± 184 × 103 | 1670 × 103 ± 177 × 103 | 113 × 103 ± 59 × 103 | 3880 × 103 ± 989 × 103 |
| Picoeukaryotes | 1361 × 103 ± 472 × 103 | 1065 × 103 ± 602.1 × 103 | 773 × 103 ± 568 × 103 | 736 × 103 ± 446 103 | 80 × 103 ± 18 × 103 | 37.2 × 103 ± 22.5 × 103 |
| Nanoeukaryotes | 180.2 × 103 ±129 × 103 | 155.3 × 103 ± 149.2 × 103 | 73.5 × 103 ± 22.8 × 103 | 82.9 × 103 ± 4402 × 103 | 14.9 × 103 ±10.1 × 103 | 22.49 × 103 ± 12.32 × 103 |
| SPhB | 1.38 107 ± 2.78 106 | 7.34 × 106 ± 6.87 × 106 | 1.27 × 107± 4.52 × 106 | 8.18 × 106 ± 4.41 × 106 | 2.57 × 106 ± 6.73 × 105 | 1.06 × 107 ± 3.64 × 105 |
| LPhB | 2.7 × 1012 ± 2.64 × 1012 | 1.53 × 108 ± 6.1 × 107 | 1.66 × 1012 ± 1.94 × 10 12 | 4.4 × 109 ± 3.32 × 109 | 1.58 × 1011 ± 1.12 × 1011 | 1.0 × 109 ± 6.76 × 108 |
| LPhA | 46.6 × 103 ± 36.7 × 103 | 29.8 × 103 ± 34.3 × 103 | 11.1 × 103 ± 7.1 × 103 | 12.9 × 103 ± 9.78 × 103 | 4.86 × 103 ± 3.99 × 103 | 3.03 × 103 ± 2.03 × 103 |
| Diatoms | Others | Coccolitophorids | Silicoflagellates | Dinoflagellates | |
|---|---|---|---|---|---|
| Coast 5 m | 62.12 ± 9.73 | 33.12 ± 5.15 | 1.07 ± 0.88 | 0.54 ± 0.57 | 3.13 ± 4.20 |
| Coast DCM | 60.88 ± 10.13 | 33.07 ± 6.99 | 1.73 ± 2.04 | 0.84 ± 0.57 | 3.48 ± 2.85 |
| Jet 5 m | 45.03 ±16.02 | 46.15 ± 11.84 | 1.83 ± 1.57 | 0.59 ± 0.53 | 6.41 ± 3.85 |
| Jet DCM | 47.77 ± 21.58 | 44.49 ± 16.52 | 2.22 ± 2.15 | 0.32 ± 0.26 | 5.21 ± 4.59 |
| Gyre 5 m | 21.59 ± 11.19 | 59.11 ± 11.84 | 3.69 ± 2.12 | 0.52 ± 0.77 | 15.09 ± 4.48 |
| Gyre DCM | 13.18 ± 6.68 | 65.46 ± 6.65 | 3.58 ± 0.73 | 1.45 ± 1.68 | 16.33 ± 5.23 |
| Diatoms | Coccolitophorids | Silicoflagellates | Dinoflagellates | Prochlorococcus | Synechococcus | Picoeukaryotes | Nanoeukaryotes | Others | |
|---|---|---|---|---|---|---|---|---|---|
| Coast 5 m | 83.37 ± 2.98 | 0.49 ± 0.28 | 0.12 ± 0.15 | 0.97 ± 0.60 | 0.097 ± 0.12 | 0.17 ± 0.22 | 0.21 ± 0.18 | 1.34 ± 1.61 | 13.6 ± 0.43 |
| Coast DCM | 86.61 ± 10.58 | 0.09 ± 0.02 | 0.05 ± 0.04 | 0.35 ± 0.24 | 11.16 ± 8.77 | 0.43 ± 0.70 | 0.12 ± 0.16 | 0.43 ± 0.70 | 0.08 ± 0.06 |
| Jet 5 m | 88.30 ±1.58 | 0.39 ± 0.32 | 0.03 ± 0.02 | 1.75 ± 0.86 | 0.12 ± 0.08 | 0.48 ± 0.36 | 0.26 ± 0.29 | 0.48 ± 0.20 | 8.30 ± 2.28 |
| Jet DCM | 80.41 ± 10.30 | 0.27 ± 0.34 | 0.01 ± 0.01 | 1.74 ± 2.56 | 14.45 ± 7.67 | 0.22 ± 0.32 | 1.19 ± 2.30 | 0.22 ± 0.32 | 0.17 ± 0.15 |
| Gyre 5 m | 84.88 ± 0.81 | 0.14 ± 0.01 | 0.20 ± 0.28 | 3.01 ± 0.22 | 0.12 ± 0.01 | 0.7 8 ± 0.28 | 0.13 ± 0.08 | 0.61 ± 0.05 | 9.91 ± 0.23 |
| Gyre DCM | 58.65 ± 13.69 | 1.19 ± 0.98 | 0.48 ± 0.03 | 6.23 ± 4.99 | 21.61 ± 11.71 | 1.28 ± 0.45 | 8.39 ± 3.12 | 1.28 ± 0.47 | 0.37± 0.14 |
| Coast | Jet | Gyre | |
|---|---|---|---|
| %AChla | 15.35 ± 7.80 [31.25**] | 17.92 ± 19.22 [91.07 **] | 6.05 ± 4.89 [13.06*] |
| Fv/Fm | 0.23 ± 0.14 [0.46*] | 0.11 ± 0.14 [0.48*] | 0.036 ± 0.08 [0.26**] |
| Sites | Coordinates | Surface | DCM | AMI | Complementary | Floor |
|---|---|---|---|---|---|---|
| Coast | 36.398° N–5.156° W | 5 | 10, 12, 19 | - | 20, 25 | 50 |
| Jet | 35.998° N–5.129° W | 5 | 17, 25, 40 | 50 | 75 | 200 |
| Gyre | 35.705° N–4.405° W | 5 | 50 | 65, 68, 70, 75, 80 | 175, 220 | 400 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartual, A.; Hernanz-Torrijos, M.; Sala, I.; Ortega, M.J.; González-García, C.; Bolado-Penagos, M.; López-Urrutia, A.; Romero-Martínez, L.; Lubián, L.M.; Bruno, M.; et al. Types and Distribution of Bioactive Polyunsaturated Aldehydes in a Gradient from Mesotrophic to Oligotrophic Waters in the Alborán Sea (Western Mediterranean). Mar. Drugs 2020, 18, 159. https://doi.org/10.3390/md18030159
Bartual A, Hernanz-Torrijos M, Sala I, Ortega MJ, González-García C, Bolado-Penagos M, López-Urrutia A, Romero-Martínez L, Lubián LM, Bruno M, et al. Types and Distribution of Bioactive Polyunsaturated Aldehydes in a Gradient from Mesotrophic to Oligotrophic Waters in the Alborán Sea (Western Mediterranean). Marine Drugs. 2020; 18(3):159. https://doi.org/10.3390/md18030159
Chicago/Turabian StyleBartual, Ana, María Hernanz-Torrijos, Iria Sala, María J. Ortega, Cristina González-García, Marina Bolado-Penagos, Angel López-Urrutia, Leonardo Romero-Martínez, Luís M. Lubián, Miguel Bruno, and et al. 2020. "Types and Distribution of Bioactive Polyunsaturated Aldehydes in a Gradient from Mesotrophic to Oligotrophic Waters in the Alborán Sea (Western Mediterranean)" Marine Drugs 18, no. 3: 159. https://doi.org/10.3390/md18030159
APA StyleBartual, A., Hernanz-Torrijos, M., Sala, I., Ortega, M. J., González-García, C., Bolado-Penagos, M., López-Urrutia, A., Romero-Martínez, L., Lubián, L. M., Bruno, M., Echevarría, F., & García, C. M. (2020). Types and Distribution of Bioactive Polyunsaturated Aldehydes in a Gradient from Mesotrophic to Oligotrophic Waters in the Alborán Sea (Western Mediterranean). Marine Drugs, 18(3), 159. https://doi.org/10.3390/md18030159