Immunoadjuvant Activity of Fucoidans from the Brown Alga Fucus evanescens


 ,
,
Abstract
1. Introduction
2. Results
2.1. The Effect of Fucoidans on the Maturation of Dcs Generated from Mice Bone Marrow
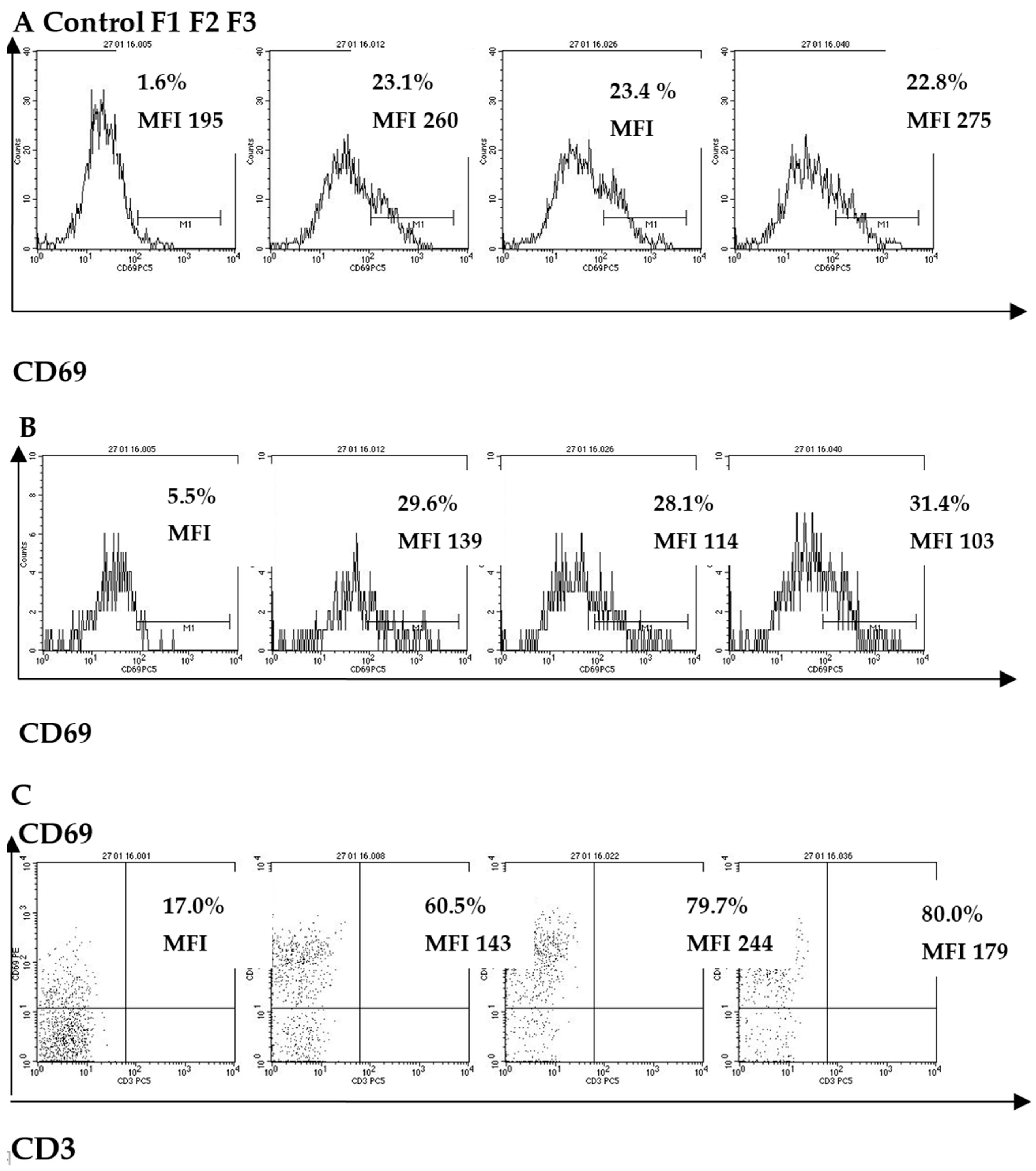
2.2. The Effect of Fucoidans on CD Molecule Expression in Peripheral Blood Human Innate Immunity Cells (Neutrophils, Monocytes, NK-Cells)
2.3. The Effect of Fucoidans on the Expression of Activation Molecules on Peripheral Blood T- and B-Cells
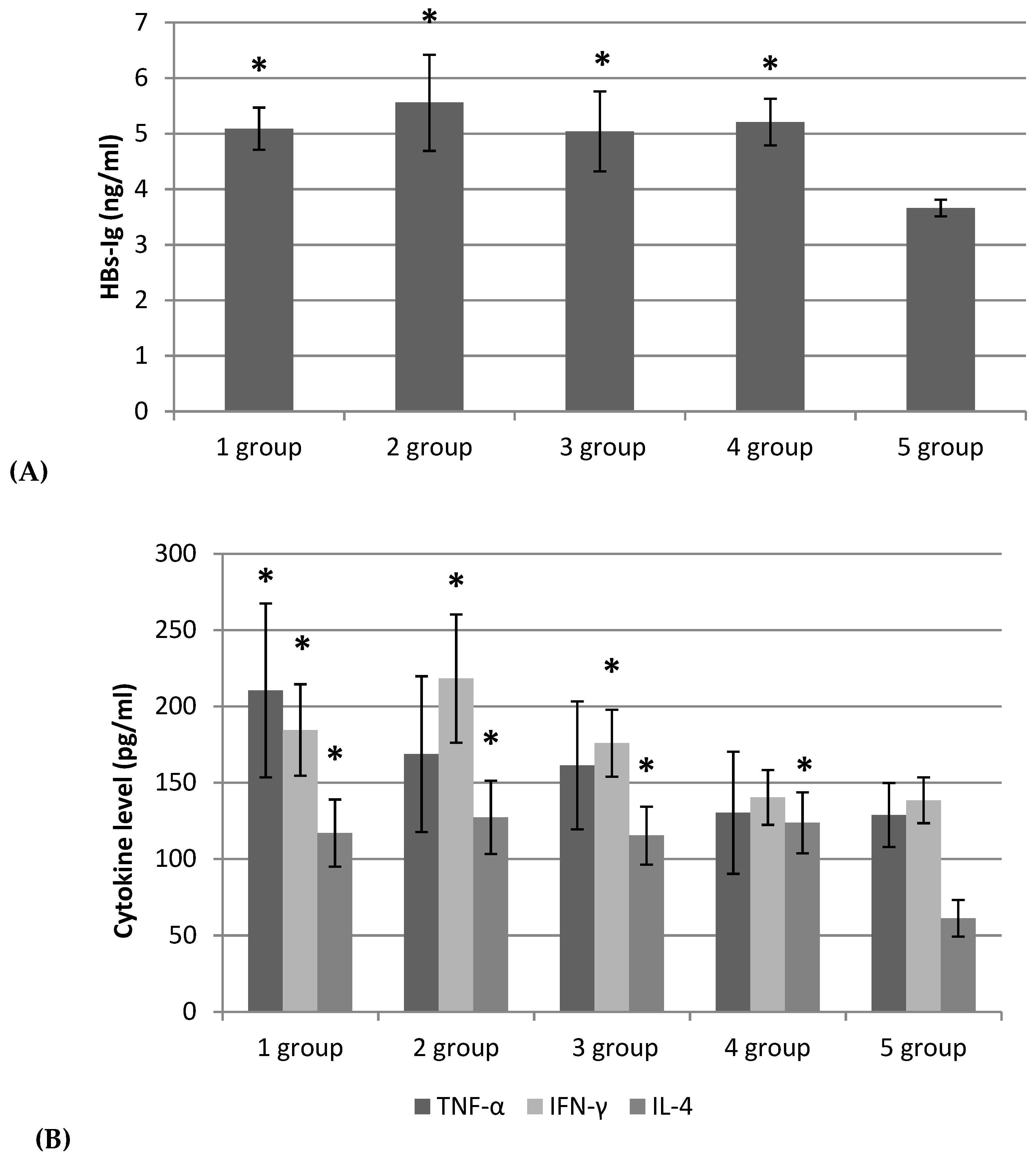
2.4. Adjuvant Effect of Fucoidans on the Hb-Specific Immune Response and Cytokine Production In Vivo
3. Discussion
4. Materials and Methods
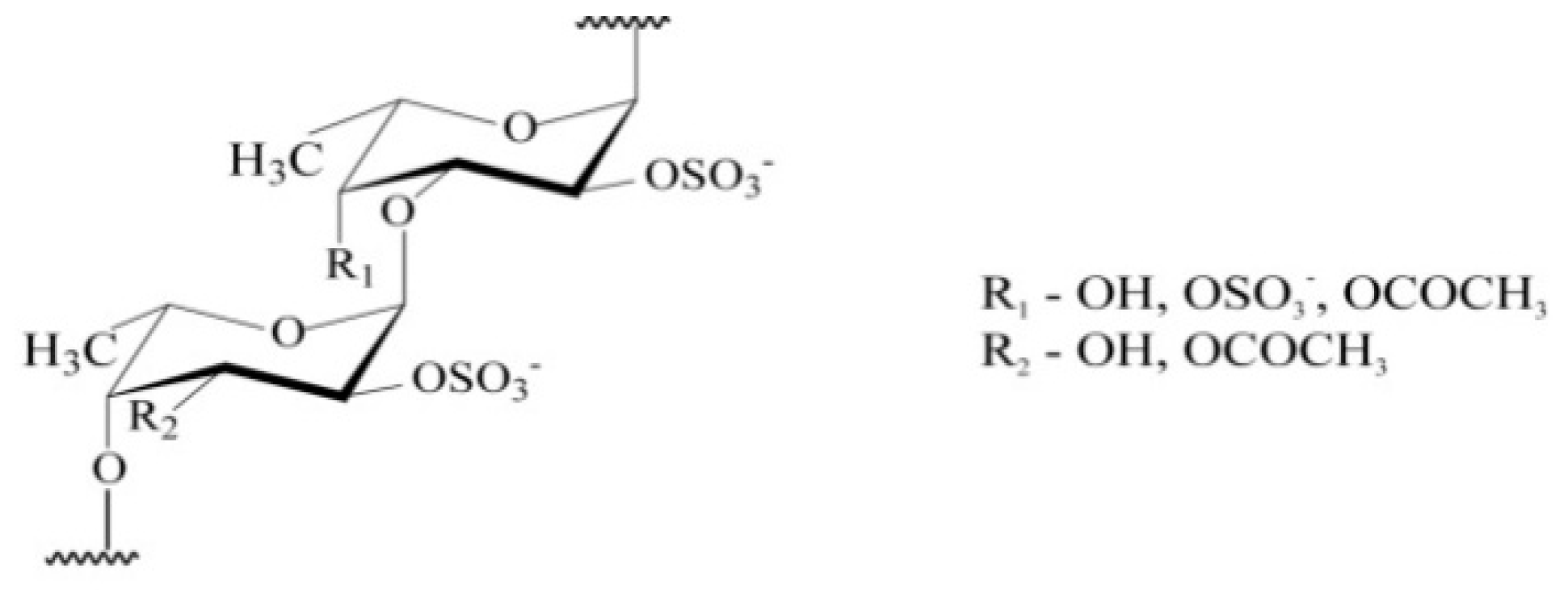
4.1. Chemical Characteristics of the Fucoidans
4.2. Peripheral Blood Cells
4.3. Flow Cytometry Analysis
4.4. Mice and Ethics Statement
4.5. DC Generation
4.6. Immunization of Mice with Recombinant Hepatitis B Virus Surface Antigen (Hbs-AG) (Abcam Limited, United Kingdom) and Blood Sampling
4.7. ELISA, Antibody Titration
4.8. Quantitative Detection of Cytokines in the Serum of Mice
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DC | dendritic cell |
| CD | cluster of differentiation |
| HBs AG | recombinant hepatitis B virus surface antigen |
| IFN-γ | interferon gamma |
| IG | immunoglobulin |
| IL | interleukin |
| MAbs | monoclonal antibodies |
| NK | natural killer cells |
| OVA | ovalbuminum |
| Th1, Th | T-helper1, 2 |
| TNF-α | tumor necrosis factor alpha |
References
- Menshova, R.V.; Shevchenko, N.M.; Imbs, T.I.; Zvyagintseva, T.N.; Maluarenko, O.S.; Zaporoshets, T.S.; Besednova, N.N.; Ermakova, S.P. Fucoidans from Brown Alga Fucus evanescens: Structure and Biological Activity. Front. Mar. Sci. 2016, 3, 129. [Google Scholar] [CrossRef]
- Li, B.; Wei, X.J.; Sun, J.L.; Xu, S.Y. Structural investigation of a fucoidan containing a fucose-free core from the brown seaweed, Hizikia fusiforme. Carbohydr. Res. 2006, 341, 1135–1146. [Google Scholar] [CrossRef] [PubMed]
- Bilan, M.I.; Grachev, A.A.; Ustuzhanina, N.E.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. Structure of a fucoidan from the brown seaweed Fucus evanescens C.Ag. Carbohydr. Res. 2002, 337, 719–730. [Google Scholar] [CrossRef]
- Zvyagintseva, T.N.; Shevchenko, N.M.; Chizhov, A.O.; Krupnova, T.N.; Sundukova, E.V.; Isakov, V.V. Water-soluble polysaccharides of some brown seaweeds. Distribution, structure and dependence on the developmental conditions. J. Exp. Mar. Biol. Ecol. 2003, 294, 1–13. [Google Scholar] [CrossRef]
- Hahn, T.; Lang, S.; Ulber, R.; Muffler, K. Novel procedures for the extraction of fucoidan from brown algae. Process Biochem. 2012, 47, 1691–1698. [Google Scholar] [CrossRef]
- Silchenko, A.S.; Rasin, A.B.; Kusaykin, M.I.; Malyarenko, O.S.; Shevchenko, N.M.; Zueva, A.O.; Kalinovsky, A.I.; Zvyagintseva, T.N.; Ermakova, S.P. Modification of native fucoidan from Fucus evanescens by recombinant fucoidanase from marine bacteria Formosa algae. Carbohydr. Polym. 2018, 193, 189–195. [Google Scholar] [CrossRef]
- Cao, H.T.T.; Mikkelsen, M.D.; Lezyk, M.J.; Bui, L.M.; Tran, V.T.T.; Silchenko, A.S.; Kusaykin, M.I.; Pham, T.D.; Truong, B.H.; Holck, J.; et al. Novel Enzyme Actions for Sulphated Galactofucan Depolymerisation and a New Engineering Strategy for Molecular Stabilisation of Fucoidan Degrading Enzymes. Mar. Drugs 2018, 16, 422. [Google Scholar] [CrossRef]
- Jin, J.O.; Zhang, W.; Du, J.Y.; Wong, K.W.; Oda, T.; Yu, Q. Fucoidan can function as an adjuvant in vivo to enhance dendritic cell maturation and function and promote antigen-specific T cell immune responses. PLoS ONE 2014, 9, e99396. [Google Scholar] [CrossRef]
- Zhang, W.; Oda, T.; Yu, Q.; Jin, J.-O. Fucoidan from Macrocystis pyrifera has powerful immune-modulatory effects compared to three other fucoidans. Mar. Drugs 2015, 13, 1084–1104. [Google Scholar]
- Song, L.; Chen, X.; Liu, X.; Zhang, F.; Hu, L.; Yue, Y.; Li, K.; Li, P. Characterization and comparison of the structural features, immune-modulatory and anti-Avian influenza virus activities conferred by three algal sulfated polysaccharides. Mar. Drugs 2016, 14, 4. [Google Scholar] [CrossRef]
- Li, L.J.; Li, M.Y.; Li, Y.T.; Feng, J.J.; Hao, F.Q.; Lun, Z. Adjuvant activity of Sargassum pallidum polysaccharides against combined newcastle disease, infectious bronchitis and avian influenza inactivated vaccines. Mar. Drugs 2012, 10, 2648–2660. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Joo, H.G. Evaluation of adjuvant effects of fucoidan for improving vaccine efficacy. J. Vet. Sci. 2015, 16, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Teruya, T.; Takeda, S.; Tamaki, Y.; Tako, M. Fucoidan isolated from Laminaria angustata var. longissima induced macrophage activation. Biosci. Biotechnol. Biochem. 2010, 74, 1960–1962. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, S.; Grenha, A. Activation of macrophages: Establishing a role for polysaccharides in drug delivery strategies envisaging antibacterial therapy. Curr. Pharm. Des. 2015, 21, 4869–4887. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.O.; Yu, Q. Fucoidan delays apoptosis and induces pro-inflammatory cytokine production in human neutrophils. Int. J. Biol. Macromol. 2015, 73, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Du, J.Y.; Jiang, Z.; Okimura, T.; Oda, T.; Yu, Q.; Jin, J.O. Ascophyllan purified from Ascophyllum nodosum induces Th1 and Tc1 immune responses by promoting dendritic cell maturation. Mar. Drugs 2014, 12, 4148–4164. [Google Scholar] [CrossRef]
- Negishi, H.; Mori, M.; Mori, H.; Yamori, Y. Supplementation of elderly japanese men and women with fucoidan from seaweed increases immune responses to seasonal influenza vaccination. J. Nutr. 2013, 143, 1794–1798. [Google Scholar] [CrossRef]
- Maruyama, H.; Tamauchi, H.; Iizuka, M.; Nakano, T. The role of NK cells in antitumor activity of dietary fucoidan from Undaria pinnatifida sporophylls (Mekabu). Planta Med. 2006, 72, 1415–1417. [Google Scholar] [CrossRef]
- Kim, M.-H.; Joo, H.-G. Immunostimulatory effects of fucoidan on bone marrow-derived dendritic cells. Immunol. Lett. 2008, 115, 138–143. [Google Scholar] [CrossRef]
- Yang, M.; Ma, C.; Sun, J.; Shao, Q.; Gao, W.; Zhang, Y.; Li, Z.; Xie, Q.; Dong, Z.; Qu, X. Fucoidan stimulation induces a functional maturation of human monocyte-derived dendritic cells. Int. Immunopharmacol. 2008, 8, 1754–1760. [Google Scholar] [CrossRef]
- Hochrein, H.; O’Keeffe, M. Dendritic cell subsets and toll-like receptors. Handb. Exp. Pharmacol. 2008, 183, 153–179. [Google Scholar]
- Blander, J.M.; Medzhitov, R. Toll-dependent selection of microbial antigens for presentation by dendritic cells. Nature 2006, 440, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Joffre, O.; Nolte, M.A.; Sporri, R.; Reis, E.; Sousa, C. Inflammatory signals in dendritic cell activation and the induction of adaptive immunity. Immunol. Rev. 2009, 227, 234–247. [Google Scholar] [CrossRef] [PubMed]
- Delirezh, N.; Majedi, L.; Rezaei, A.S.; Ranjkeshzadeh, H. Generation of Mature Monocyte-Derived Dendritic Cells in the Presence of Heparin and Monocyte Conditioned Medium: Phenotypic and Functional Comparison. Iran. Biomed. J. 2011, 15, 79–84. [Google Scholar]
- Klein, E.; Koch, S.; Borm, B.; Neumann, J.; Herzog, V.; Koch, N.; Bieber, T. CD83 localization in a recycling compartment of immature human monocyte-derived dendritic cells. Int. Immunol. 2005, 17, 477–487. [Google Scholar] [CrossRef]
- Clausen, J.; Vergeiner, B.; Enk, M.; Petzer, A.L.; Gastl, G.; Gunsilius, E. Functional significance of the activation-associated receptor CD25 and CD69 on human NK-cells and NK-like T-cells. Immunobiology 2003, 207, 85–93. [Google Scholar] [CrossRef]
- Evans, J.H.; Horowitz, A.; Mehrabi, M.; Wise, E.L.; Pease, J.E.; Riley, E.M.; Davis, D.M. A distinct subset of human NK cells expressing HLA-DR expand in response to IL-2 and can aid immune responses to BCG. Eur. J. Immunol. 2011, 41, 1924–1933. [Google Scholar] [CrossRef]
- Romagnani, S. T-cell subsets (Th1 versus Th2). Ann. Allergy Asthma Immunol. 2000, 85, 9–21. [Google Scholar] [CrossRef]
- Seder, R.A.; Hill, A.V. Vaccines against intracellular infections requiring cellular immunity. Nature 2000, 406, 793–798. [Google Scholar] [CrossRef]
- McKee, A.S.; Munks, M.W.; Marrack, P. How do adjuvants work? Important considerations for new generation adjuvants. Immunity 2007, 27, 687–690. [Google Scholar] [CrossRef]
- Zamorano, J.; Riva, M.D.; Pérez, G.M. Interleukin-4: A multifunctional cytokine. Immunología 2003, 22, 215–224. [Google Scholar]
- Mathers, A.R.; Cuff, C.F. Role of Interleukin-4 (IL-4) and IL-10 in Serum Immunoglobulin G Antibody Responses following Mucosal or Systemic Reovirus. Infect. J. Virol. 2004, 78, 3352–3360. [Google Scholar] [CrossRef] [PubMed]
- Jafarzadeh, A.; Shokri, F. The antibody response to HBs antigen is regulated by coordinated Th1 and Th2 cytokine production in healthy neonates. Clin. Exp. Immunol. 2003, 131, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Gherardi, R.K.; Eidi, H.; Crepeaux, G.; Authier, F.J.; Cadusseau, J. Biopersistence and brain translocation of aluminum adjuvants of vaccines. Front. Neurol. 2015, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Zvyagintseva, T.N.; Shevchenko, N.M.; Popivnich, I.B.; Isakov, V.V.; Scobun, A.S.; Sundukova, E.V.; Elyakova, L.A. A new procedure for the separation of water-soluble polysaccharides from brown seaweeds. Carbohydr. Res. 1999, 322, 32–39. [Google Scholar] [CrossRef]
- Imbs, T.I.; Skriptsova, A.V.; Zvyagintseva, T.N. Antioxidant activity of fucoise-containing sulfated polysaccharides obtained from Fucus evanescens by different extraction methods. J. Appl. Phycol. 2015, 27, 545–553. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Percentage (%) of Cells Expressing Markers | Negative Control (Immature DC-GM-CSF and IL-4) | Positive Control (TNF-α) | Fucoidans | ||
|---|---|---|---|---|---|
| F1 | F2 | F3 | |||
| CD34 | 42.9 ± 4.5 | 6.9 ± 2.1 ** | 10.2 ± 0.9 ** | 14.3 ± 5.7 * | 12.2 ± 5.6 ** |
| CD83 | 8.1 ± 1.9 | 68.1 ± 4.8 ** | 53.9 ± 7.5 ** | 48.5 ± 6.5 ** | 64.7 ± 5.6 ** |
| CD14 | 57.2 ± 6.2 | 34.7 ± 4.6 * | 40.1 ± 0.7 * | 34.8 ± 7.6 * | 42.1 ± 5.4 * |
| CD11c | 10.5 ± 3.7 | 38.5 ± 1.4 ** | 21.9 ± 5.9 * | 27.1 ± 4.9 * | 32.1 ± 4.1 * |
| CD38 | 20.2 ± 3.6 | 35.8 ± 5.3 * | 27.9 ± 2.5 * | 38.8 ± 8.8 * | 37.6 ± 3.2 * |
| CD80 | 14.3 ± 4.3 | 51.1 ± 5.6 ** | 28.8 ± 4.2 * | 35.7 ± 5.9 * | 49.6 ± 8.6 * |
| CD86 | 15.6 ± 2.4 | 57.9 ± 6.7 ** | 41.4 ± 3.9 ** | 30.2 ± 7.6 * | 36.1 ± 5.6 ** |
| MHC-II | 20.9 ± 1.7 | 78.1 ± 2.8 ** | 70.7 ± 7.1 ** | 63.4 ± 2.5 ** | 71.9 ± 6.3 ** |
| Cell Subpopulation | CD Molecule | Results | Fucoidans | Control | ||
|---|---|---|---|---|---|---|
| F1 | F2 | F3 | ||||
| Neutrophils | CD69 | MFI | 74.3 * (48–87) | 82.0 ** (51–104) | 87.3 ** (49–123) | 31.2 (22–49) |
| CD14 | MFI | 52.0 * (43–82) | 78.6 ** (62–101) | 75.3 ** (66–94) | 25.3 (18–30) | |
| CD11b | MFI | 2621.3 * (1738–3670) | 2706 * (1839–3631) | 2981 * (2341–3894) | 1023 (802–1264) | |
| CD62L | MFI | 62 ** (45–78) | 37.3 ** (30–45) | 34.8 ** (29–44) | 174 (142–211) | |
| Monocytes | CD69 | % | 30 ** (22–44) | 29 ** (24–45) | 28 ** (23–41) | 5 (3–15) |
| MFI | 139 * (95–146) | 114 * (96–143) | 103 * (89–125) | 38 (23–45) | ||
| HLA-DR | % | 68 * (55–80) | 77 * (69–85) | 75 * (68–81) | 47 (44–51) | |
| MFI | 153 ** (118–186) | 200 ** (157–241) | 149 ** (113–183) | 61 (48–74) | ||
| CD83 | % | 4.3 * (2.0–6.6) | 4.8 * (2.3–7.3) | 7.2 * (4.6–9.7) | 0.8 (0.6–0.9) | |
| MFI | 3.7 * (2.2–4.2) | 5.3 * (3.8–6.7) | 4.9 * (3.1–4.7) | 1.5 (1.1–1.8) | ||
| NK-cells | CD69 | % | 57.9 ** (53.6–61.9) | 78.1 ** (73.2–81) | 78.9 ** (74–81.2) | 12.7 (8.5–20.7) |
| MFI | 140.4 ** (105.4–174) | 220 ** (154–265) | 170 ** (121–202) | 42.8 (31.2–64.7) | ||
| CD25 | % | 7.0 * (5.2–9.3) | 6.7 * (4.5–8.7) | 5.6 * (4.1–6.9) | 2.8 (1.3–3.7) | |
| MFI | 6.3 (4.8–7.5) | 5.2 (3.6–6.8) | 6.4 (4.9–7.3) | 5.9 (3.5–6.8) | ||
| HLA-DR | % | 4.7 (3.0–6.4) | 8.4 * (6.2–10.5) | 4.4 (2.8–6.9) | 4.2 (2.1–6.7) | |
| MFI | 2.8 (1.3–3.9) | 6.1 * (4.5–8.9) | 2.2 (1.2–4.0) | 2.0 (1.1–3.4) | ||
| CD8 | % | 57.6 (53.4–61.2) | 56.7 (51.6–59.7) | 54.2 (49.3–58.9) | 55.5 (49.0–58.1) | |
| MFI | 193 * (179–207) | 183 * (174–191) | 163 (152–173) | 159 (145–162) | ||
| CD107a | % | 2.6 * (1.8–3.9) | 2.1 (1.6–3.8) | 4.0 * (2.5–5.7) | 1.4 (0.7–1.8) | |
| Cell Subpopu Lation | CD Molecule | Results of Measure Ment | Fucoidans | Control | ||
|---|---|---|---|---|---|---|
| F1 | F2 | F3 | ||||
| T-cells | CD69 | % | 5.8 * (3.6–7.2) | 6.2 * (3.8–8.9) | 6.5 * (4.1–8.8) | 3.0 (1.6–5.4) |
| CD25 | % | 9.1 * (6.2–14.1) | 6.4 (4.1–8.8) | 5.3 (3.1–7.8) | 5.9 (3.7–7.9) | |
| HLA-DR | % | 5.1 * (3.4–8.7) | 3.8 (2.2–6.1) | 3.6 (2.9–6.2) | 3.4 (1.9–5.6) | |
| CD71 | % | 6.4 * (4.2–8.9) | 5.4 (3.2–9.0) | 4.9 (3.4–8.1 | 4.1 (2.1–6.9) | |
| B-cells | CD69 | % | 25.1 * (18.5–28.4) | 28.7 * (16.4–32.6) | 24.4 * (17.9–28.6) | 11.3 (6.2–14.2) |
| CD25 | % | 6.6 ** (4.9–9.5) | 5.8 * (3.3–8.2) | 6.9 * (4.1–10.1) | 2.3 (1.2–3.5) | |
| CD86 | % | 15.9 ** (11.8–18.6) | 12.2 * (8.2–17.5) | 14.4 ** (10.5–18.9) | 6.5 (3.2–9.4) | |
| CD71 | % | 30.8 * (24.6–35.7) | 34.5 * (22.7–39.4) | 41.6 * (20.3–44.8) | 24.5 (15.4–27.5) | |
| TLR2 | % | 9.1 ** (6.7–10.3) | 8.8 ** (5.3–13.4) | 8.5 ** (5.9–10.8) | 3.0 (0.9–5.1) | |
| № | Characteristic of Fucoidans | Molecular Weight (kDa) | SO3Na* (%) | Monosaccharide Composition (% mol) | ||||
|---|---|---|---|---|---|---|---|---|
| Fuc | Gal | Xyl | Man | Glc | ||||
| F1 | Native fucoidan [35] | 130–430 | 27.0 | 94.1 | 3.8 | 2.1 | 0 | 0 |
| F2 | Highly purified fucoidan [36] | 130–400 | 28.1 | 94.4 | 3.5 | 2.1 | 0 | 0 |
| F3 | Product of enzymatic hydrolysis of fucoidan [6] | 50.8 | 29.7 | 97.8 | 2.2 | 0 | 0 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuznetsova, T.A.; Smolina, T.P.; Makarenkova, I.D.; Ivanushko, L.A.; Persiyanova, E.V.; Ermakova, S.P.; Silchenko, A.S.; Zaporozhets, T.S.; Besednova, N.N.; Fedyanina, L.N.; et al. Immunoadjuvant Activity of Fucoidans from the Brown Alga Fucus evanescens. Mar. Drugs 2020, 18, 155. https://doi.org/10.3390/md18030155
Kuznetsova TA, Smolina TP, Makarenkova ID, Ivanushko LA, Persiyanova EV, Ermakova SP, Silchenko AS, Zaporozhets TS, Besednova NN, Fedyanina LN, et al. Immunoadjuvant Activity of Fucoidans from the Brown Alga Fucus evanescens. Marine Drugs. 2020; 18(3):155. https://doi.org/10.3390/md18030155
Chicago/Turabian StyleKuznetsova, Tatyana A., Tatyana P. Smolina, Ilona D. Makarenkova, Lydmila A. Ivanushko, Elena V. Persiyanova, Svetlana P. Ermakova, Artem S. Silchenko, Tatyana S. Zaporozhets, Natalya N. Besednova, Lydmila N. Fedyanina, and et al. 2020. "Immunoadjuvant Activity of Fucoidans from the Brown Alga Fucus evanescens" Marine Drugs 18, no. 3: 155. https://doi.org/10.3390/md18030155
APA StyleKuznetsova, T. A., Smolina, T. P., Makarenkova, I. D., Ivanushko, L. A., Persiyanova, E. V., Ermakova, S. P., Silchenko, A. S., Zaporozhets, T. S., Besednova, N. N., Fedyanina, L. N., & Kryzhanovsky, S. P. (2020). Immunoadjuvant Activity of Fucoidans from the Brown Alga Fucus evanescens. Marine Drugs, 18(3), 155. https://doi.org/10.3390/md18030155