Cyanobacteria and Red Macroalgae as Potential Sources of Antioxidants and UV Radiation-Absorbing Compounds for Cosmeceutical Applications



Abstract
1. Introduction
2. Results
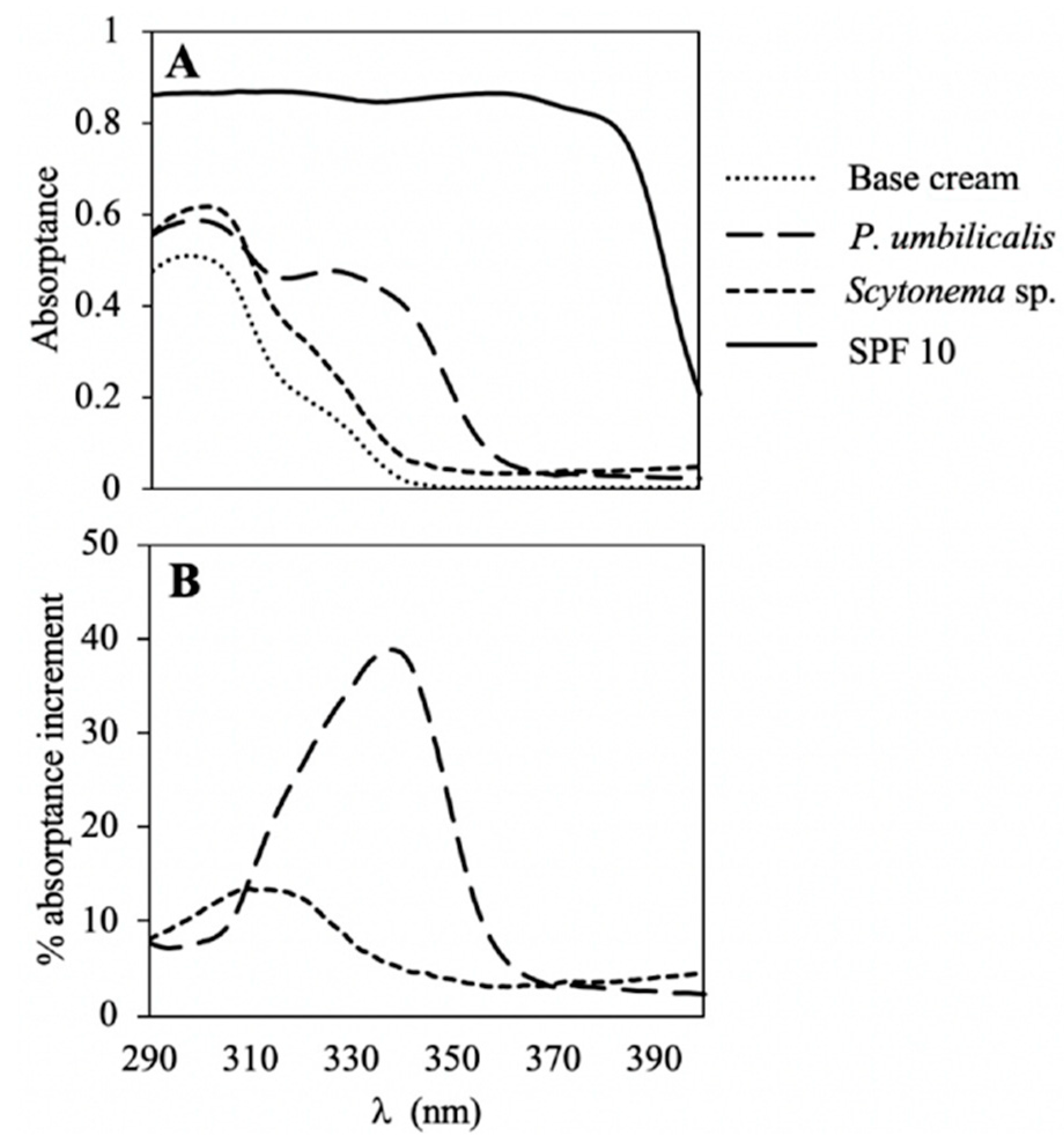
2.1. UV Absorption Spectra
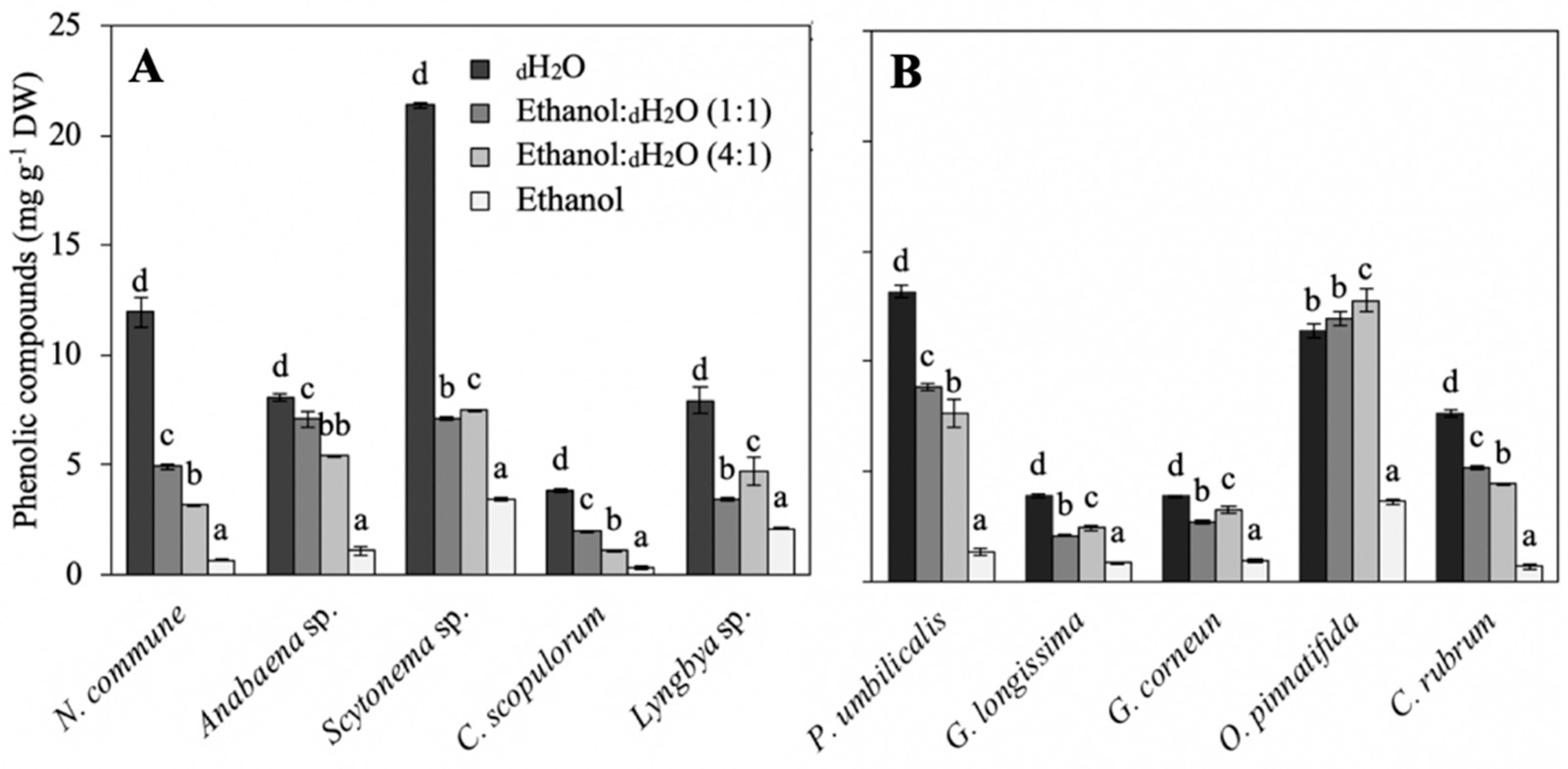
2.2. Biochemical Composition
2.3. Antioxidant Activity
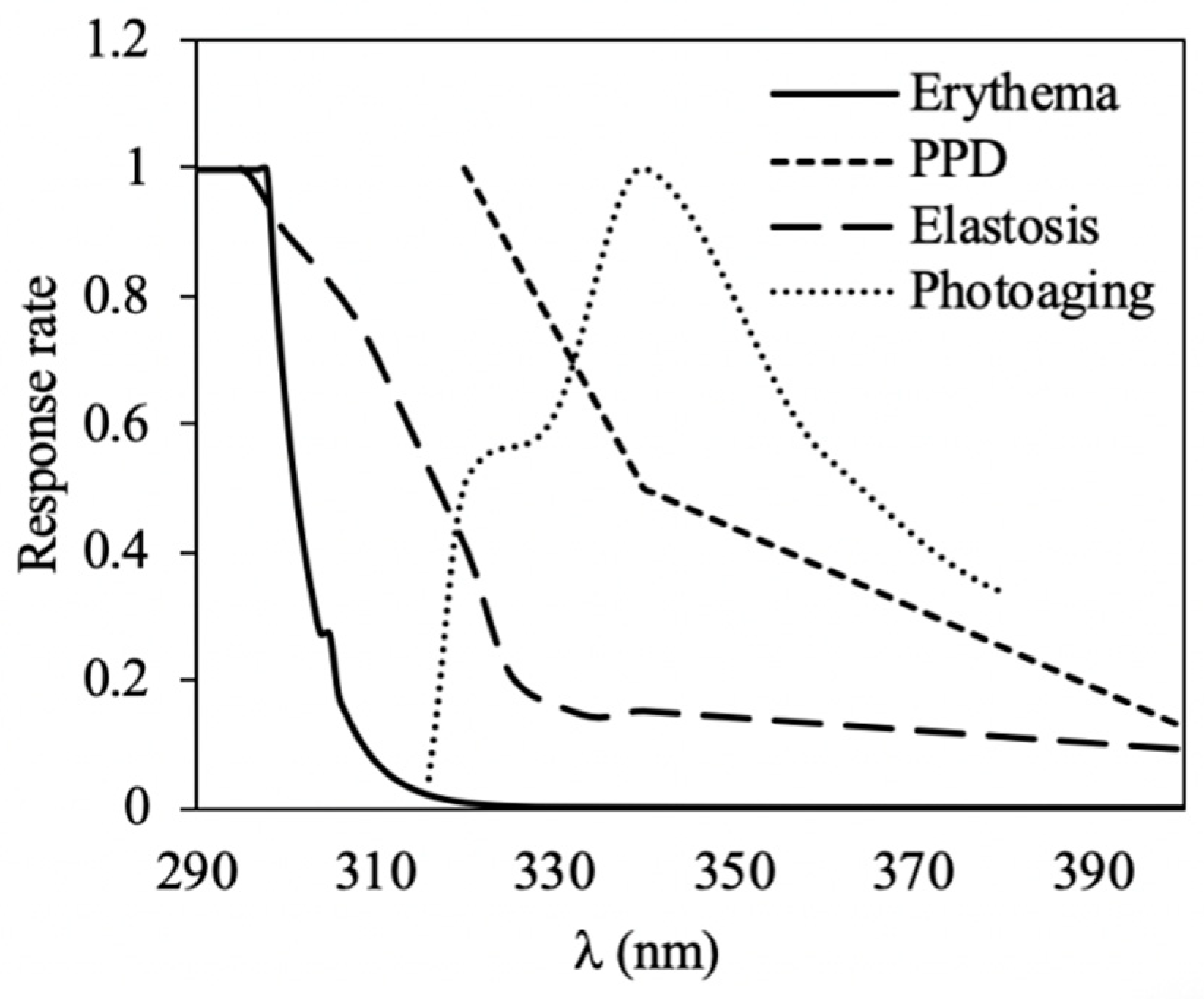
2.4. Photoprotective Capacity
2.5. Correlations and Principal Component Analyses
3. Discussion
3.1. Biochemical Composition
3.2. Bioactivities (Antioxidant and Photoprotective) and its Relation with Biochemical Composition
3.3. Future Perspectives
4. Conclusions
5. Materials and Methods
5.1. Biological Material
5.2. Preparation of Algal Extracts
5.3. Biochemical Composition
5.3.1. Total Carbon and Nitrogen
5.3.2. Proteins
5.3.3. Carbohydrates
5.3.4. Lipids
5.3.5. Phycobiliproteins and Scytonemin
5.3.6. Phenolic Compounds
5.3.7. Mycosporine-Like Amino Acids (MAAs)
5.4. Antioxidant Activity (ABTS and DPPH)
5.5. Photoprotection Capacity
- Act.Sp (λ) = action spectra (Figure 6);
- E (λ) = spectral irradiance of a sunny midday in a summer day (June) in Malaga (W m−2);
- T (λ) = transmittance values at each wavelength (0–1);
- A (λ) = absorptance values at each wavelength (0–1).
5.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ichihashi, M.; Ueda, M.; Budiyanto, A.; Bito, T.; Oka, M.; Fukunaga, M.; Tsuru, K.; Horikawa, T. UV-induced skin damage. Toxicology 2003, 189, 21–39. [Google Scholar] [CrossRef]
- Schuch, A.P.; Moreno, N.C.; Schuch, N.J.; Menck, C.F.M.; Garcia, C.C.M. Sunlight damage to cellular DNA: Focus on oxidatively generated lesions. Free Radic. Biol. Med. 2017, 107, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Harrison, G.I.; Young, A.R. Ultraviolet radiation-induced erythema in human skin. Methods 2002, 28, 14–19. [Google Scholar] [CrossRef]
- Gallagher, R.P.; Lee, T.K. Adverse effects of ultraviolet radiation: A brief review. Prog. Biophys. Mol. Biol. 2006, 92, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y.; Ananthaswamy, H.N. Toxic effects of ultraviolet radiation on the skin. Toxicol. Appl. Pharmacol. 2004, 195, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Sola, Y.; Lorente, J. Contribution of UVA irradiance to the erythema and photoaging effects in solar and sunbed exposures. J. Photochem. Photobiol. B Biol. 2015, 143, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, B.H.; Hexsel, C.L.; Hamzavi, I.H.; Lim, H.W. Effects of visible light on the skin. Photochem. Photobiol. 2008, 84, 450–462. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, B.H.; Ruvolo, E.; Hexsel, C.L.; Liu, Y.; Owen, M.R.; Kollias, N.; Lim, H.W.; Hamzavi, I.H. Impact of long-wavelength UVA and visible light on melanocompetent skin. J. Invest. Dermatol. 2010, 130, 2092–2097. [Google Scholar] [CrossRef]
- Nakashima, Y.; Ohta, S.; Wolf, A.M. Blue light-induced oxidative stress in live skin. Free Radic. Biol. Med. 2017, 108, 300–310. [Google Scholar] [CrossRef]
- Wang, J.; Pan, L.; Wu, S.; Lu, L.; Xu, Y.; Zhu, Y.; Guo, M.; Zhuang, S. Recent advances on endocrine disrupting effects of UV filters. Int. J. Environ. Res. Public Health 2016, 13, 782. [Google Scholar] [CrossRef]
- Tsui, M.M.P.; Leung, H.W.; Wai, T.C.; Yamashita, N.; Taniyasu, S.; Liu, W.; Lam, P.K.S.; Murphy, M.B. Occurrence, distribution and ecological risk assessment of multiple classes of UV filters in surface waters from different countries. Water Res. 2014, 67, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Quiles, D.; Tovar-Sánchez, A. Are sunscreens a new environmental risk associated with coastal tourism? Environ. Int. 2015, 83, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Grether-Beck, S.; Marini, A.; Jaenicke, T.; Krutmann, J. Photoprotection of human skin beyond ultraviolet radiation. Photodermatol. Photoimmunol. Photomed. 2014, 30, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Saewan, N.; Jimtaisong, A. Natural products as photoprotection. J. Cosmet. Dermatol. 2015, 14, 47–63. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Ravichandran, Y.D.; Khan, S.B.; Kim, Y.T. Prospective of the cosmeceuticals derived from marine organisms. Biotechnol. Bioprocess Eng. 2008, 13, 511–523. [Google Scholar] [CrossRef]
- Wada, N.; Sakamoto, T.; Matsugo, S. Multiple roles of photosynthetic and sunscreen pigments in cyanobacteria focusing on the oxidative stress. Metabolites 2013, 3, 463–483. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K. Algae as production systems of bioactive compounds. Eng. Life Sci. 2015, 15, 160–176. [Google Scholar] [CrossRef]
- Kumar, J.; Singh, D.; Tyagi, M.B.; Kumar, A. Cyanobacteria: Applications in biotechnology. In Cyanobacteria: From Basic Science to Applications; Academic Press: Cambridge, MA, USA, 2018; pp. 327–346. [Google Scholar] [CrossRef]
- Whitton, B.A.; Potts, M. Introduction to the cyanobacteria. In Ecology Cyanobacteria II; Springer: Dordrecht, The Netherlands, 2012; pp. 1–13. [Google Scholar]
- Lalegerie, F.; Gager, L.; Stiger-Pouvreau, V.; Connan, S. The stressful life of red and brown seaweeds on the temperate intertidal zone: Effect of abiotic and biotic parameters on the physiology of macroalgae and content variability of particular metabolites. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2020; Volume 95, pp. 247–287. [Google Scholar] [CrossRef]
- Dring, M.J. Stress Resistance and disease resistance in seaweeds: The role of reactive oxygen metabolism. Adv. Bot. Res. 2005, 43, 175–207. [Google Scholar] [CrossRef]
- Lesser, M.P. Oxidative stress in marine environments: Biochemistry and physiological ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef]
- Singh, S.P.; Häder, D.-P.; Sinha, R.P. Cyanobacteria and ultraviolet radiation (UVR) stress: Mitigation strategies. Ageing Res. Rev. 2010, 9, 79–90. [Google Scholar] [CrossRef]
- Bischof, K.; Steinhoff, F.S. Impacts of ozone Stratospheric depletion and solar UVB radiation on seaweeds. In Seaweed Biology; Ecological Studies: Watsonville, CA, USA, 2012; pp. 433–448. [Google Scholar] [CrossRef]
- Noyma, N.P.; Silva, T.P.; Chiarini-Garcia, H.; Amado, A.M.; Roland, F.; Melo, R.C.N. Potential effects of UV radiation on photosynthetic structures of the bloom-forming cyanobacterium Cylindrospermopsis raciborskii CYRF-01. Front. Microbiol. 2015, 6, 1202. [Google Scholar] [CrossRef] [PubMed]
- Mallick, N.; Mohn, F.H. Reactive oxygen species: Response of algal cells. J. Plant Physiol. 2000, 157, 183–193. [Google Scholar] [CrossRef]
- McKinlay, A.F. Reference action spectrum for ultraviolet induced erythema in human skin. CIE J. 1987, 6, 17–22. [Google Scholar]
- Diffey, B.L.; Robson, J. A new substrate to measure sunscreen protection factors throughout the ultraviolet spectrum. J. Soc. Cosmet. Chem. 1989, 40, 127–133. [Google Scholar]
- Pissavini, M.; Tricaud, C.; Wiener, G.; Lauer, A.; Contier, M.; Kolbe, L.; Trullás Cabanas, C.; Boyer, F.; Nollent, V.; Meredith, E.; et al. Validation of an in vitro sun protection factor (SPF) method in blinded ring-testing. Int. J. Cosmet. Sci. 2018, 40, 263–268. [Google Scholar] [CrossRef] [PubMed]
- The European Cosmetic and Perfumery Association. In Vitro Method for the Determination of the UVA Protection Factor and “Critical Wavelength” Values of Sunscreen Products; The European Cosmetic and Perfumery Association: Brussels, Belgium, 2011. [Google Scholar]
- Wulf, H.C.; Poulsen, T.; Davies, R.E.; Urbach, F. Narrow-band UV radiation and induction of dermal elastosis and skin cancer. Photodermatology 1989, 6, 44–51. [Google Scholar] [PubMed]
- Bissett, D.L.; Hannon, D.P.; Orr, T.V. Wavelength dependence of histological, physical and visible changes in chronically UV-irradiated hairless mouse skin. Photochem. Photobiol. 1989, 50, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Fabo, E.C.D.; Kripke, M.L. Wavelength dependence and dose-rate independence of UV radiation-induced immunologic unresponsiveness of mice to a UV-induced fibrosarcoma. Photochem. Photobiol. 1980, 32, 183–188. [Google Scholar] [CrossRef]
- De la Coba, F.; Aguilera, J.; Korbee, N.; de Gálvez, M.V.; Herrera-Ceballos, E.; Álvarez-Gómez, F.; Figueroa, F.L. UVA and UVB Photoprotective capabilities of topical formulations containing mycosporine-like amino acids (maas) through different biological effective protection factors (BEPFs). Mar. Drugs 2019, 17, 55. [Google Scholar] [CrossRef]
- Schneider, G.; Figueroa, F.L.; Vega, J.; Chaves, P.; Álvarez-Gómez, F.; Korbee, N.; Bonomi-Barufi, J. Photoprotection properties of marine photosynthetic organisms grown in high ultraviolet exposure areas: Cosmeceutical applications. Algal Res. 2020, 49, 101956. [Google Scholar] [CrossRef]
- Lourenço, S.O.; Barbarino, E.; De-Paula, J.C.; Pereira, L.O.D.S.; Marquez, U.M.L. Amino acid composition, protein content and calculation of nitrogen-to-protein conversion factors for 19 tropical seaweeds. Phycol. Res. 2002, 50, 233–241. [Google Scholar] [CrossRef]
- Vega, J.; Álvarez-Gómez, F.; Güenaga, L.; Figueroa, F.L.; Gómez-Pinchetti, J.L. Antioxidant activity of extracts from marine macroalgae, wild-collected and cultivated, in an integrated multi-trophic aquaculture system. Aquaculture 2020, 522, 735088. [Google Scholar] [CrossRef]
- Lourenço, S.O.; Barbarino, E.; Lavín, P.L.; Marquez, U.M.L.; Aidar, E. Distribution of intracellular nitrogen in marine microalgae: Calculation of new nitrogen-to-protein conversion factors. Eur. J. Phycol. 2004, 39, 17–32. [Google Scholar] [CrossRef]
- Lapointe, B.E.; Ryther, J.H. The effects of nitrogen and seawater flow rate on the growth and biochemical composition of Gracilaria foliifera var. angustissima in mass outdoor cultures. Bot. Mar. 1979, 22, 529–538. [Google Scholar] [CrossRef]
- Duarte, C.M. Nutrient concentration of aquatic plants: Patterns across species. Limnol. Oceanogr. 1992, 37, 882–889. [Google Scholar] [CrossRef]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Usov, A.I. Polysaccharides of the red algae. In Advances in Carbohydrate Chemistry and Biochemistry; Academic Press: Cambridge, MA, USA, 2011; Volume 65, pp. 115–217. [Google Scholar] [CrossRef]
- Zhang, Q.; Yu, P.; Li, Z.; Zhang, H.; Xu, Z.; Li, P. Antioxidant activities of sulfated polysaccharide fractions from Porphyra haitanesis. J. Appl. Phycol. 2003, 15, 305–310. [Google Scholar] [CrossRef]
- Abdala-Díaz, R.T.; Chabrillón, M.; Cabello-Pasini, A.; Gómez-Pinchetti, J.L.; Figueroa, F.L. Characterization of polysaccharides from Hypnea spinella (Gigartinales) and Halopithys incurva (Ceramiales) and their effect on RAW 264.7 macrophage activity. J. Appl. Phycol. 2011, 23, 523–528. [Google Scholar] [CrossRef]
- Abdala-Díaz, R.T.; Casas Arrojo, V.; Arrojo Agudo, M.A.; Cárdenas, C.; Dobretsov, S.; Figueroa, F.L. Immunomodulatory and Antioxidant Activities of Sulfated Polysaccharides from Laminaria ochroleuca, Porphyra umbilicalis, and Gelidium corneum. Mar. Biotechnol. 2019, 21, 577–587. [Google Scholar] [CrossRef]
- Singh, S.; Das, S. Screening, production, optimization and characterization of cyanobacterial polysaccharide. World J. Microbiol. Biotechnol. 2011, 27, 1971–1980. [Google Scholar] [CrossRef]
- Wang, H.B.; Wu, S.J.; Liu, D. Preparation of polysaccharides from cyanobacteria Nostoc commune and their antioxidant activities. Carbohydr. Polym. 2014, 99, 553–555. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xu, J.; Liu, Y.; Ai, S.; Qin, F.; Li, Z.; Zhang, H.; Huang, Z. Antioxidant and moisture-retention activities of the polysaccharide from Nostoc commune. Carbohydr. Polym. 2011, 83, 1821–1827. [Google Scholar] [CrossRef]
- Kumari, P.; Kumar, M.; Gupta, V.; Reddy, C.R.K.; Jha, B. Tropical marine macroalgae as potential sources of nutritionally important PUFAs. Food Chem. 2010, 120, 749–757. [Google Scholar] [CrossRef]
- Patel, V.K.; Sundaram, S.; Patel, A.K.; Kalra, A. Characterization of Seven species of cyanobacteria for high-quality biomass production. Arab. J. Sci. Eng. 2018, 43, 109–121. [Google Scholar] [CrossRef]
- Freile-Pelegrín, Y.; Robledo, D. Bioactive phenolic compounds from algae. In Bioactive Compounds from Marine Foods: Plant and Animal Sources; Wiley Blackwell: Hoboken, NJ, USA, 2013; pp. 113–129. ISBN 9781118412893. [Google Scholar]
- Machu, L.; Misurcova, L.; Ambrozova, J.V.; Orsavova, J.; Mlcek, J.; Sochor, J.; Jurikova, T. Phenolic content and antioxidant capacity in algal food products. Molecules 2015, 20, 1118–1133. [Google Scholar] [CrossRef] [PubMed]
- Stevanato, R.; Bertelle, M.; Fabris, S. Photoprotective characteristics of natural antioxidant polyphenols. Regul. Toxicol. Pharmacol. 2014, 69, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Zhang, X.; Chen, F.; Wang, M. Dietary polyphenols as photoprotective agents against UV radiation. J. Funct. Foods 2017, 30, 108–118. [Google Scholar] [CrossRef]
- Gager, L.; Connan, S.; Molla, M.; Couteau, C.; Arbona, J.F.; Coiffard, L.; Cérantola, S.; Stiger-Pouvreau, V. Active phlorotannins from seven brown seaweeds commercially harvested in Brittany (France) detected by 1H NMR and in vitro assays: Temporal variation and potential valorization in cosmetic applications. J. Appl. Phycol. 2020, 32, 2375–2386. [Google Scholar] [CrossRef]
- López, A.; Rico, M.; Rivero, A.; Suárez de Tangil, M. The effects of solvents on the phenolic contents and antioxidant activity of Stypocaulon scoparium algae extracts. Food Chem. 2011, 125, 1104–1109. [Google Scholar] [CrossRef]
- Hwang, E.S.; Do Thi, N. Effects of extraction and processing methods on antioxidant compound contents and radical scavenging activities of laver (Porphyra tenera). Prev. Nutr. Food Sci. 2014, 19, 40–48. [Google Scholar] [CrossRef]
- Zubia, M.; Robledo, D.; Freile-Pelegrin, Y. Antioxidant activities in tropical marine macroalgae from the Yucatan Peninsula, Mexico. J. Appl. Phycol. 2007, 19, 449–458. [Google Scholar] [CrossRef]
- Li, H.B.; Cheng, K.W.; Wong, C.C.; Fan, K.W.; Chen, F.; Jiang, Y. Evaluation of antioxidant capacity and total phenolic content of different fractions of selected microalgae. Food Chem. 2007, 102, 771–776. [Google Scholar] [CrossRef]
- Singh, D.P.; Prabha, R.; Verma, S.; Meena, K.K.; Yandigeri, M. Antioxidant properties and polyphenolic content in terrestrial cyanobacteria. 3 Biotech 2017, 7, 134. [Google Scholar] [CrossRef] [PubMed]
- Samarakoon, K.; Jeon, Y.J. Bio-functionalities of proteins derived from marine algae—A review. Food Res. Int. 2012, 48, 948–960. [Google Scholar] [CrossRef]
- Ferdouse, F.; Holdt, S.L.; Smith, R.; Murúa, P.; Yang, Z. The global status of seaweed production, trade and utilization. FAO Globefish Res. Program. 2018, 124, 120. [Google Scholar]
- Rosales-Loaiza, N.; Aiello-Mazzarri, C.; Gómez, L.; Arredondo, B.; Morales, E. Nutritional quality of biomass from four strains of Nostoc and Anabaena grown in batch cultures. Int. Food Res. J. 2017, 24, 2212–2219. [Google Scholar]
- Hemlata; Fatma, T. Screening of cyanobacteria for phycobiliproteins and effect of different environmental stress on its yield. Bull. Environ. Contam. Toxicol. 2009, 83, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Olvera-Ramírez, R.; Coria-Cedillo, M.; Cañizares-Villanueva, R.O.; Martínez-Jerónimo, F.; Ponce-Noyola, T.; Ríos-Leal, E. Growth evaluation and bioproducts characterization of Calothrix sp. Bioresour. Technol. 2000, 72, 121–124. [Google Scholar] [CrossRef]
- Johnson, E.M.; Kumar, K.; Das, D. Physicochemical parameters optimization, and purification of phycobiliproteins from the isolated Nostoc sp. Bioresour. Technol. 2014, 166, 541–547. [Google Scholar] [CrossRef]
- Sampath-Wiley, P.; Neefus, C.D.; Jahnke, L.S. Seasonal effects of sun exposure and emersion on intertidal seaweed physiology: Fluctuations in antioxidant contents, photosynthetic pigments and photosynthetic efficiency in the red alga Porphyra umbilicalis Kützing (Rhodophyta, Bangiales). J. Exp. Mar. Bio. Ecol. 2008, 361, 83–91. [Google Scholar] [CrossRef]
- Sfriso, A.A.; Gallo, M.; Baldi, F. Phycoerythrin productivity and diversity from five red macroalgae. J. Appl. Phycol. 2018, 30, 2523–2531. [Google Scholar] [CrossRef]
- Korbee, N.; Figueroa, F.L.; Aguilera, J. Effect of light quality on the accumulation of photosynthetic pigments, proteins and mycosporine-like amino acids in the red alga Porphyra leucosticta (Bangiales, Rhodophyta). J. Photochem. Photobiol. B Biol. 2005, 80, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Lu, F.; Bi, Y.; Hu, Z. Effects of light intensity and quality on phycobiliprotein accumulation in the cyanobacterium Nostoc sphaeroides Kützing. Biotechnol. Lett. 2015, 37, 1663–1669. [Google Scholar] [CrossRef] [PubMed]
- Pagels, F.; Bonomi-Barufi, J.; Vega, J.; Abdala-Díaz, R.; Vasconcelos, V.; Guedes, A.C.; Figueroa, F.L. Light quality triggers biochemical modulation of Cyanobium sp.—Photobiology as tool for biotechnological optimization. J. Appl. Phycol. 2020, 32, 2851–2861. [Google Scholar] [CrossRef]
- Niu, J.F.; Wang, G.C.; Tseng, C.K. Method for large-scale isolation and purification of R-phycoerythrin from red alga Polysiphonia urceolata Grev. Protein Expr. Purif. 2006, 49, 23–31. [Google Scholar] [CrossRef]
- Proteau, P.J.; Gerwick, W.H.; Garcia-Pichel, F.; Castenholz, R. The structure of scytonemin, an ultraviolet sunscreen pigment from the sheaths of cyanobacteria. Experientia 1993, 49, 825–829. [Google Scholar] [CrossRef]
- Garcia-Pichel, F.; Castenholz, R.W. Characterization and biological implications of scytonemin, a cyanobacterial sheath pigment. J. Phycol. 1991, 27, 395–409. [Google Scholar] [CrossRef]
- Sinha, R.P.; Häder, D.P. UV-protectants in cyanobacteria. Plant Sci. 2008, 174, 278–289. [Google Scholar] [CrossRef]
- Mushir, S.; Deep, S.; Fatma, T.; Student, P.G. Screening of cyanobacterial strains for UV screening compound scytonemin-environmental perspectives. IJIRSET 2014, 3. [Google Scholar]
- Rath, J.; Mandal, S.; Adhikary, S.P. Salinity induced synthesis of UV-screening compound scytonemin in the cyanobacterium Lyngbya aestuarii. J. Photochem. Photobiol. B Biol. 2012, 115, 5–8. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Incharoensakdi, A. Characterization of UV-screening compounds, mycosporine-like amino acids, and scytonemin in the cyanobacterium Lyngbya sp. CU2555. FEMS Microbiol. Ecol. 2014, 87, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhao, L.; Xu, J.; Yang, R.; He, S.; Yan, X. Determination of oxidized scytonemin in Nostoc commune Vauch cultured on different conditions by high performance liquid chromatography coupled with triple quadrupole mass spectrometry. J. Appl. Phycol. 2013, 25, 1001–1007. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Sonani, R.R.; Madamwar, D. The high-energy radiation protectant extracellular sheath pigment scytonemin and its reduced counterpart in the cyanobacterium Scytonema sp. R77DM. Bioresour. Technol. 2014, 171, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Mushir, S.; Fatma, T. Monitoring stress responses in cyanobacterial scytonemin—Screening and characterization. Environ. Technol. 2012, 33, 153–157. [Google Scholar] [CrossRef]
- Fleming, E.D.; Castenholz, R.W. Effects of nitrogen source on the synthesis of the UV-screening compound, scytonemin, in the cyanobacterium Nostoc punctiforme PCC 73102. FEMS Microbiol. Ecol. 2008, 63, 301–308. [Google Scholar] [CrossRef]
- Sinha, R.P.; Singh, S.P.; Häder, D.P. Database on mycosporines and mycosporine-like amino acids (MAAs) in fungi, cyanobacteria, macroalgae, phytoplankton and animals. J. Photochem. Photobiol. B Biol. 2007, 89, 29–35. [Google Scholar] [CrossRef]
- Carreto, J.I.; Carignan, M.O. Mycosporine-like amino acids: Relevant secondary metabolites. Chemical and ecological aspects. Mar. Drugs 2011, 9, 387–446. [Google Scholar] [CrossRef]
- Garcia-Pichel, F.; Castenholz, R.W. Occurrence of UV-absorbing, mycosporine-like compounds among cyanobacterial isolates and an estimate of their screening capacity. Appl. Environ. Microbiol. 1993, 59, 163–169. [Google Scholar] [CrossRef]
- Singh, S.P.; Klisch, M.; Sinha, R.P.; Häder, D.P. Effects of abiotic stressors on synthesis of the mycosporine-like amino acid shinorine in the cyanobacterium Anabaena variabilis PCC 7937. Photochem. Photobiol. 2008, 84, 1500–1505. [Google Scholar] [CrossRef]
- Sinha, R.P. Biochemical characterization of sunscreening mycosporine-like amino acids from two Nostoc species inhabiting diverse habitats. Protoplasma 2014, 252, 199–208. [Google Scholar] [CrossRef]
- Sinha, R.P.; Klisch, M.; Walter Helbling, E.; Häder, D.P. Induction of mycosporine-like amino acids (MAAs) in cyanobacteria by solar ultraviolet-B radiation. J. Photochem. Photobiol. B Biol. 2001, 60, 129–135. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, N.; Zhou, J.; Dong, S.; Zhang, X.; Guo, L.; Guo, G. Distribution, contents, and types of mycosporine-like amino acids (MAAs) in marine macroalgae and a database for MAAs based on these characteristics. Mar. Drugs 2020, 18, 43. [Google Scholar] [CrossRef] [PubMed]
- Korbee, N. Fotorregulación y Efecto del Nitrógeno Inorgánico en la Acumulación de Aminoácidos Tipo Micosporina en Algas Rojas. Ph.D. Thesis, Universidad de Málaga, Málaga, Spain, 2003. [Google Scholar]
- Huovinen, P.; Gómez, I.; Figueroa, F.L.; Ulloa, N.; Morales, V.; Lovengreen, C. Ultraviolet-absorbing mycosporine-like amino acids in red macroalgae from Chile. Bot. Mar. 2004, 47, 21–29. [Google Scholar] [CrossRef]
- Briani, B.; Sissini, M.N.; Lucena, L.A.; Batista, M.B.; Costa, I.O.; Nunes, J.M.C.; Schmitz, C.; Ramlov, F.; Maraschin, M.; Korbee, N.; et al. The influence of environmental features in the content of mycosporine-like amino acids in red marine algae along the Brazilian coast. J. Phycol. 2018, 54, 380–390. [Google Scholar] [CrossRef] [PubMed]
- Navarro, N.; Figueroa, F.L.; Korbee, N.; Bonomi, J.; Gómez, F.Á.; De la Coba, F. Mycosporine-like amino acids from red algae to develop natural UV sunscreens. In Sunscreens: Source, Formulations, Efficacy and Recommendations; Ministry of Environment, Forest and Climate Change, Research in Environment (RE) Division, Indira Paryavaran Bhavan: New Delhi, India, 2018; pp. 99–129. ISBN 9781536132953. [Google Scholar]
- Torres, P.; Santos, J.P.; Chow, F.; Dos Santos, D.Y.A.C. A comprehensive review of traditional uses, bioactivity potential, and chemical diversity of the genus Gracilaria (Gracilariales, Rhodophyta). Algal Res. 2019, 37, 288–306. [Google Scholar] [CrossRef]
- Chrapusta, E.; Kaminski, A.; Duchnik, K.; Bober, B.; Adamski, M.; Bialczyk, J. Mycosporine-like amino acids: Potential health and beauty ingredients. Mar. Drugs 2017, 15, 326. [Google Scholar] [CrossRef]
- Chaves-Peña, P.; De La Coba, F.; Figueroa, F.L.; Korbee, N. Quantitative and qualitative HPLC analysis of mycosporine-like amino acids extracted in distilled water for cosmetical uses in four rhodophyta. Mar. Drugs 2020, 18, 27. [Google Scholar] [CrossRef]
- Álvarez-Gómez, F.; Korbee, N.; Figueroa, F.L. Analysis of antioxidant capacity and bioactive compounds in marine macroalgal and lichenic extracts using different solvents and evaluation methods. Ciencias Mar. 2016, 42, 271–288. [Google Scholar] [CrossRef]
- Álvarez-Gómez, F.; Korbee, N.; Casas-Arrojo, V.; Abdala-Díaz, R.T.; Figueroa, F.L. UV photoprotection, cytotoxicity and immunology capacity of red algae extracts. Molecules 2019, 24, 341. [Google Scholar] [CrossRef]
- Lalegerie, F.; Lajili, S.; Bedoux, G.; Taupin, L.; Stiger-Pouvreau, V.; Connan, S. Photo-protective compounds in red macroalgae from Brittany: Considerable diversity in mycosporine-like amino acids (MAAs). Mar. Environ. Res. 2019, 147, 37–48. [Google Scholar] [CrossRef]
- Torres, P.B.; Chow, F.; Ferreira, M.J.P.; Dos Santos, D.Y.A.C. Mycosporine-like amino acids from Gracilariopsis tenuifrons (Gracilariales, Rhodophyta) and its variation under high light. J. Appl. Phycol. 2016, 28, 2035–2040. [Google Scholar] [CrossRef]
- De La Coba, F.; Aguilera, J.; Figueroa, F.L.; De Gálvez, M.V.; Herrera, E. Antioxidant activity of mycosporine-like amino acids isolated from three red macroalgae and one marine lichen. J. Appl. Phycol. 2009, 21, 161–169. [Google Scholar] [CrossRef]
- Wada, N.; Sakamoto, T.; Matsugo, S. Mycosporine-like amino acids and their derivatives as natural antioxidants. Antioxidants 2015, 4, 603–646. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, K.P.; Gacesa, R.; Long, P.F.; Young, A.R. Molecular photoprotection of human keratinocytes in vitro by the naturally occurring mycosporine-like amino acid palythine. Br. J. Dermatol. 2018, 178, 1353–1363. [Google Scholar] [CrossRef] [PubMed]
- Guedes, A.; Gião, M.; Seabra, R.; Ferreira, A.; Tamagnini, P.; Moradas-Ferreira, P.; Malcata, F. Evaluation of the antioxidant activity of cell extracts from microalgae. Mar. Drugs 2013, 11, 1256–1270. [Google Scholar] [CrossRef]
- Jerez-Martel, I.; García-Poza, S.; Rodríguez-Martel, G.; Rico, M.; Afonso-Olivares, C.; Gómez-Pinchetti, J.L. Phenolic profile and antioxidant activity of crude extracts from microalgae and cyanobacteria strains. J. Food Qual. 2017, 2017, 2924508. [Google Scholar] [CrossRef]
- Assunção, M.F.G.; Amaral, R.; Martins, C.B.; Ferreira, J.D.; Ressurreição, S.; Santos, S.D.; Varejão, J.M.T.B.; Santos, L.M.A. Screening microalgae as potential sources of antioxidants. J. Appl. Phycol. 2017, 29, 865–877. [Google Scholar] [CrossRef]
- Kelman, D.; Kromkowski Posner, E.; Mcdermid, K.J.; Tabandera, N.K.; Wright, P.R.; Wright, A.D. Antioxidant activity of Hawaiian marine algae. Mar. Drugs 2012, 10, 403–416. [Google Scholar] [CrossRef]
- Hajimahmoodi, M.; Faramarzi, M.A.; Mohammadi, N.; Soltani, N.; Oveisi, M.R.; Nafissi-Varcheh, N. Evaluation of antioxidant properties and total phenolic contents of some strains of microalgae. J. Appl. Phycol. 2010, 22, 43–50. [Google Scholar] [CrossRef]
- Monteiro, M.; Santos, R.A.; Iglesias, P.; Couto, A.; Serra, C.R.; Gouvinhas, I.; Barros, A.; Oliva-Teles, A.; Enes, P.; Díaz-Rosales, P. Effect of extraction method and solvent system on the phenolic content and antioxidant activity of selected macro-and microalgae extracts. J. Appl. Phycol. 2020, 32, 349–362. [Google Scholar] [CrossRef]
- Guedes, A.C.; Amaro, H.M.; Gião, M.S.; Malcata, F.X. Optimization of ABTS radical cation assay specifically for determination of antioxidant capacity of intracellular extracts of microalgae and cyanobacteria. Food Chem. 2013, 138, 638–643. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B. Some methodological problems in the determination of antioxidant activity using chromogen radicals: A practical case. Trends Food Sci. Technol. 2000, 11, 419–421. [Google Scholar] [CrossRef]
- Kim, D.O.; Lee, K.W.; Lee, H.J.; Lee, C.Y. Vitamin C equivalent antioxidant capacity (VCEAC) of phenolic phytochemicals. J. Agric. Food Chem. 2002, 50, 3713–3717. [Google Scholar] [CrossRef] [PubMed]
- Sekar, S.; Chandramohan, M. Phycobiliproteins as a commodity: Trends in applied research, patents and commercialization. J. Appl. Phycol. 2008, 20, 113–136. [Google Scholar] [CrossRef]
- Pagels, F.; Guedes, A.C.; Amaro, H.M.; Kijjoa, A.; Vasconcelos, V. Phycobiliproteins from cyanobacteria: Chemistry and biotechnological applications. Biotechnol. Adv. 2019, 37, 422–443. [Google Scholar] [CrossRef]
- Matsui, K.; Nazifi, E.; Hirai, Y.; Wada, N.; Matsugo, S.; Sakamoto, T. The cyanobacterial UV-absorbing pigment scytonemin displays radical-scavenging activity. J. Gen. Appl. Microbiol. 2012, 58, 137–144. [Google Scholar] [CrossRef]
- Connan, S.; Delisle, F.; Deslandes, E.; Ar Gall, E. Intra-thallus phlorotannin content and antioxidant activity in Phaeophyceae of temperate waters. Bot. Mar. 2006, 49, 39–46. [Google Scholar] [CrossRef]
- Cornish, M.L.; Garbary, D.J. Antioxidants from macroalgae: Potential applications in human health and nutrition. Algae 2010, 25, 155–171. [Google Scholar] [CrossRef]
- Dunlap, W.C.; Yamamoto, Y. Small-molecule antioxidants in marine organisms: Antioxidant activity of mycosporine-glycine. Comp. Biochem. Physiol. Part B Biochem. 1995, 112, 105–114. [Google Scholar] [CrossRef]
- Yang, E.C.; Boo, S.M.; Bhattacharya, D.; Saunders, G.W.; Knoll, A.H.; Fredericq, S.; Graf, L.; Yoon, H.S. Divergence time estimates and the evolution of major lineages in the florideophyte red algae. Sci. Rep. 2016, 6, 21361. [Google Scholar] [CrossRef] [PubMed]
- De la Coba, F.; Aguilera, J.; De Gálvez, M.V.; Álvarez, M.; Gallego, E.; Figueroa, F.L.; Herrera, E. Prevention of the ultraviolet effects on clinical and histopathological changes, as well as the heat shock protein-70 expression in mouse skin by topical application of algal UV-absorbing compounds. J. Dermatol. Sci. 2009, 55, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Mercurio, D.G.; Wagemaker, T.A.L.; Alves, V.M.; Benevenuto, C.G.; Gaspar, L.R.; Maia Campos, P.M.B.G. In vivo photoprotective effects of cosmetic formulations containing UV filters, vitamins, ginkgo biloba and red algae extracts. J. Photochem. Photobiol. B Biol. 2015, 153, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Conde, F.R.; Churio, M.S.; Previtali, C.M. The photoprotector mechanism of mycosporine-like amino acids. Excited-state properties and photostability of porphyra-334 in aqueous solution. J. Photochem. Photobiol. B Biol. 2000, 56, 139–144. [Google Scholar] [CrossRef]
- Hartmann, A.; Gostner, J.; Fuchs, J.E.; Chaita, E.; Aligiannis, N.; Skaltsounis, L.; Ganzera, M. Inhibition of collagenase by mycosporine-like amino acids from marine sources. Planta Med. 2015, 81, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Hartmann, A.; Ganzera, M.; Fuchs, D.; Gostner, J. Immunomodulatory effects of the mycosporine-like amino acids shinorine and porphyra-334. Mar. Drugs 2016, 14, 119. [Google Scholar] [CrossRef] [PubMed]
- González, S.; Pathak, M.A.; Cuevas, J.; Villarrubia, V.G.; Fitzpatrick, T.B. Topical or oral administration with an extract of Polypodium leucotomos prevents acute sunburn and psoralen-induced phototoxic reactions as well as depletion of Langerhans cells in human skin. Photodermatol. Photoimmunol. Photomed. 1997, 13, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Parrado, C.; Nicolas, J.; Juarranz, A.; Gonzalez, S. The role of the aqueous extract: Polypodium leucotomos in photoprotection. Photochem. Photobiol. Sci. 2020, 19, 831–843. [Google Scholar] [CrossRef]
- Aziz, A. “Scytonemin” pigment in Lyngbya notarisii (Meneghini) wille and possibility of using it in preparing skin protecting cream. J. Drug Res. Dev. 2018, 4. [Google Scholar] [CrossRef]
- Kang, M.R.; Jo, S.A.; Lee, H.; Yoon, Y.D.; Kwon, J.H.; Yang, J.W.; Choi, B.J.; Park, K.H.; Lee, M.Y.; Lee, C.W.; et al. Inhibition of skin inflammation by scytonemin, an ultraviolet sunscreen pigment. Mar. Drugs 2020, 18, 300. [Google Scholar] [CrossRef]
- Barceló-Villalobos, M.; Figueroa, F.L.; Korbee, N.; Álvarez-Gómez, F.; Abreu, M.H. Production of mycosporine-like amino acids from Gracilaria vermiculophylla (Rhodophyta) cultured through one year in an integrated multi-trophic aquaculture (IMTA) system. Mar. Biotechnol. 2017, 19, 246–254. [Google Scholar] [CrossRef]
- Abreu, M.H.; Pereira, R.; Yarish, C.; Buschmann, A.H.; Sousa-Pinto, I. IMTA with Gracilaria vermiculophylla: Productivity and nutrient removal performance of the seaweed in a land-based pilot scale system. Aquaculture 2011, 312, 77–87. [Google Scholar] [CrossRef]
- Guedes, A.C.; Katkam, N.G.; Varela, J.; Malcata, F.X. Photobioreactors for cyanobacterial culturing. In Cyanobacteria: An Economic Perspective; Wiley Blackwell: Hoboken, NJ, USA, 2013; pp. 270–292. ISBN 9781118402238. [Google Scholar]
- Bandaranayake, W.M. Mycosporines: Are they nature’s sunscreens? Nat. Prod. Rep. 1998, 15, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Cardozo, K.H.M.; Guaratini, T.; Barros, M.P.; Falcão, V.R.; Tonon, A.P.; Lopes, N.P.; Campos, S.; Torres, M.A.; Souza, A.O.; Colepicolo, P.; et al. Metabolites from algae with economical impact. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 146, 60–78. [Google Scholar] [CrossRef] [PubMed]
- Morabito, K.; Shapley, N.C.; Steeley, K.G.; Tripathi, A. Review of sunscreen and the emergence of non-conventional absorbers and their applications in ultraviolet protection. Int. J. Cosmet. Sci. 2011, 33, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Mishra, S.K.; Suh, W.I.; Farooq, W.; Moon, M.; Shrivastav, A.; Park, M.S.; Yang, J.W. Rapid quantification of microalgal lipids in aqueous medium by a simple colorimetric method. Bioresour. Technol. 2014, 155, 330–333. [Google Scholar] [CrossRef]
- Beer, S.; Eshel, A. Determining phycoerythrin and phycocyanin concentrations in aqueous crude extracts of red algae. Mar. Freshw. Res. 1985, 36, 785. [Google Scholar] [CrossRef]
- Bennett, A.; Bogobad, L. Complementary chromatic adaptation in a filamentous blue-green alga. J. Cell Biol. 1973, 58, 419–435. [Google Scholar] [CrossRef]
- Garcia-Pichel, F.; Sherry, N.D.; Castenholz, R.W. Evidence for an ultraviolet sunscreen role of the extracellular pigment scytonemin in the terrestrial cyanobacterium Chlorogloeopsis sp. Photochem. Photobiol. 1992, 56, 17–23. [Google Scholar] [CrossRef]
- Singleton, V.; Rossi, J. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Korbee-Peinado, N.; Abdala Díaz, R.T.; Figueroa, F.L.; Helbling, E.W. Ammonium and UV radiation stimulate the accumulation of mycosporine-like amino acids in Porphyra columbina (Rhodophyta) from Patagonia, Argentina. J. Phycol. 2004, 40, 248–259. [Google Scholar] [CrossRef]
- Karsten, U.; Sawall, T.; Hanelt, D.; Bischof, K.; Figueroa, F.L.; Flores-Moya, A.; Wiencke, C. An inventory of UV-absorbing mycosporine-like amino acids in macroalgae from polar to warm-temperate regions. Bot. Mar. 1998, 41, 443–453. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | C | N | C:N | Total Proteins | Soluble Proteins | Carbohydrates | Lipids |
|---|---|---|---|---|---|---|---|
| N. commune. | 420.1 ± 0.2 d | 59.3 ± 0.3 c | 7.09 ± 0.04 c | 283.4 ± 1.3 c | 70.6 ± 1.7 e | 407.8 ± 64.8 d | 21.1 ± 0.3 c |
| Anabaena sp. | 406.8 ± 0.6 c | 85.1 ± 0.2 e | 4.78 ± 002 a | 406.9 ± 1.0 e | 40.4 ± 0.2 b | 185.4 ± 18.9 a | 15.5 ± 0.8 a |
| Scytonema sp. | 426.9 ± 0.8 e | 61.8 ± 0.2 d | 6.91 ± 0.02 b | 295.3 ± 1.1 d | 62.0 ± 0.5 d | 196.4 ± 63.6 a | 17.9 ± 0.2 b |
| C. scopulorum | 313.3 ± 1.2 a | 34.6 ± 0.3 a | 9.06 ± 0.02 e | 165.4 ±1.2 a | 8.3 ± 0.1 a | 285.6 ± 21.3 b | 19.0 ± 1.1 b |
| Lyngbya sp. | 352.5 ± 0.9 b | 42.8 ± 0.3 b | 8.23 ± 0.03 d | 204.8 ± 1.5 b | 44.3 ± 2.0 c | 334.1 ± 40.3 c | 18.2 ± 0.8 b |
| P. umbilicalis | 340.7 ± 11.5 c | 44.5 ± 2.9 d | 7.65 ± 0.24 b | 219.1 ± 14.3 d | 33.9 ±1.8 e | 318.5 ± 19.2 d | 5.7 ± 0.1 b |
| G. longissima | 255.8 ± 5.5 b | 29.7 ± 3.0 b | 8.66 ± 1.06 d | 146.3 ± 14.8 b | 7.8 ± 0.2 b | 206.5 ± 12.0 b | 4.8 ± 0.1 a |
| G. corneum | 351.2 ± 19.4 c | 33.2 ± 3.0 c | 10.58 ± 0.38 c | 163.6 ± 14.8 c | 11.2 ± 0.2 c | 274.8 ± 44.9 c | 4.8 ± 0.3 a |
| O. pinnatifida | 231.2 ± 5.6 a | 25.1 ± 1.7 a | 9.23 ± 0.40 c | 123.4 ± 8.4 a | 5.4 ± 0.5 a | 192.1 ± 27.9 b | 7.5 ± 0.3 c |
| C. rubrum | 239.4 ± 13.4 a | 36.0 ± 2.4 c | 6.65 ± 0.07 a | 177.1 ± 11.8 c | 19.4 ± 1.2 d | 139.5 ± 29.8 a | 5.5 ± 0.1 b |
| Species | PE | PC | Scyt. |
|---|---|---|---|
| N. commune | 14.01 ± 0.14 e | 24.82 ± 0.07 e | 0.31 ± 0.02 a |
| Anabaena sp. | 1.86 ± 0.01 c | 1.77 ± 0.01 b | 0.09 ± 0.04 a |
| Scytonema sp. | 3.77 ± 0.01 d | 19.58 ± 0.28 d | 2.22 ± 0.26 c |
| C. scopulorum | 0.78 ± 0.08 a | 0.75 ± 0.16 a | 0.14 ± 0.02 a |
| Lyngbya sp. | 1.21 ± 0.09 b | 12.35 ± 0.4 c | 1.24 ± 0.18 b |
| P. umbilicalis | 3.36 ± 0.04 e | 0.25 ± 0.06 cd | - |
| G. longissima | 0.51 ± 0.02 b | 0.01 ± 0.01 a | - |
| G. corneum | 1.25± 0.08 d | 0.33 ± 0.10 d | - |
| O. pinnatifida | 0.07 ± 0.05 a | 0.18 ± 0.02 bc | - |
| C. rubrum | 0.91 ± 0.01 c | 0.08 ± 0.01 ab | - |
| Species | Total MAAs Content (mg g−1 DW) | Type of MAA | % Relative | Molecular Formula | λmax (nm) | Error (ppm) | Calculated m/z [M + H]+ | Observed m/z [M + H]+ |
|---|---|---|---|---|---|---|---|---|
| Cyanobacteria | ||||||||
| Scytonema sp. | 0.62 ±0.12 a | Myc-glutaminol | ≥98 | C13H22N2O6 | 310 | 3.06 | 303.15506 | 303.15414 |
| Lyngbya sp. | 1.83 ± 0.15 b | Palythine | 2.4 ± 0.8 | C10H16N2O5 | 320 | 2.29 | 245.11320 | 245.11264 |
| Asterina-330 | 96.4 ± 3.2 | C2H20N2O6 | 330 | 2.66 | 289.13941 | 289.13864 | ||
| Rhodophyta | ||||||||
| P. umbilicalis | 5.2 ± 0.40 d | Myc-glutamine | ≤1 | C13H20N2O7 | 310 | 1.23 | 317.13433 | 317.13394 |
| Palythine | ≤1 | C10H16N2O5 | 320 | 1.14 | 245.11320 | 245.11292 | ||
| Palythinol | ≤1 | C13H22N2O6 | 332 | 1.50 | 303.15506 | 303.15460 | ||
| Asterina-330 | ≤1 | C12H20N2O6 | 330 | 1.28 | 289.13941 | 289.13904 | ||
| Shinorine | 8.4 ± 1.2 | C13H20N2O8 | 333 | 1.65 | 333.12924 | 333.12869 | ||
| Porphyra-334 | 88.2 ± 2.4 | C14H22N2O8 | 334 | 2.27 | 347.14489 | 347.14410 | ||
| G. longissima | 0.42 ± 0.10 a | Shinorine | ≥98 | C13H20N2O8 | 333–334 | 3.48 | 333.12924 | 333.12808 |
| G. corneum | 1.84 ± 0.23 c | Palythine | 14.5 ± 1.1 | C10H16N2O5 | 320 | 2.53 | 245.11320 | 245.11258 |
| Asterina-330 | 62.9 ± 3.8 | C12H20N2O6 | 330 | 3.18 | 289.13941 | 289.13849 | ||
| Shinorine | 18.7 ± 1.6 | C13H20N2O8 | 333–334 | 2.85 | 333.12924 | 333.12830 | ||
| O. pinnatifida | 1.51 ± 0.20 bc | Palythine | 28.8 ± 3.3 | C10H16N2O5 | 320 | 1.35 | 245.11320 | 245.11287 |
| Palythinol | 4.3 ± 1.2 | C13H22N2O6 | 332 | 1.32 | 303.15506 | 303.15466 | ||
| Asterina-330 | 36.1 ± 2.4 | C12H20N2O6 | 330 | 1.62 | 289.13941 | 289.13895 | ||
| Shinorine | 3.8 ± 1.0 | C13H20N2O8 | 333–334 | 1.83 | 333.12924 | 333.12863 | ||
| Porphyra-334 | 25.5 ± 1.5 | C14H22N2O8 | 334 | 1.93 | 347.14489 | 347.14423 | ||
| C. rubrum | 0.97 ± 0.17 b | Palythine | 23.3 ± 4.1 | C10H16N2O5 | 320 | 3.38 | 245.11320 | 245.11237 |
| Shinorine | 75.8 ± 3.7 | C13H20N2O8 | 333–334 | 3.57 | 333.12924 | 333.12805 | ||
| Action Spectra | Index | Base Cream | P. umbilicalis | Scytonema sp. | Positive Control (SPF 10) |
|---|---|---|---|---|---|
| Erythema (SPF) | BEPFs | 1.51 ± 0.02 a | 2.12 ± 0.11 c | 1.96 ± 0.06 bc | 8.30 ± 0.40 |
| % ESAR | 33.40 ± 0.02 a | 50.86 ± 2.31 c | 47.69 ± 1.37 bc | 86.19 ± 4.02 | |
| PPD (UVAPF) | BEPFs | 1.05 ± 0.01 a | 1.45 ± 0.02 c | 1.12 ± 0.01 b | 7.17 ± 0.20 |
| % ESAR | 3.94 ± 0.16 a | 23.69 ± 0.78 c | 8.50 ± 0.28 b | 81.33 ± 3.67 | |
| Elastosis | BEPFs | 1.12 ± 0.04 a | 1.54 ± 0.03 c | 1.26 ± 0.01 b | 7.38 ± 0.20 |
| % ESAR | 7.49 ± 0.12 a | 24.44 ± 0.96 c | 13.26 ± 0.43 b | 79.92 ± 2.43 | |
| Photoaging | BEPFs | 1.03 ± 0.01 a | 1.35 ± 0.01 c | 1.07 ± 0.01 b | 7.15 ± 0.21 |
| % ESAR | 1.05 ± 0.11 a | 17.04 ± 0.65 c | 4.01 ± 0.12 b | 85.27 ± 3.88 |
| Cyanobacteria | Rhodophyta | ||
|---|---|---|---|
| Species | Code | Species | Place and Date of Collection |
| Nostoc commune Vaucher ex Bornet & Flahault | BEA 0024B | Porphyra umbilicalis Kützing | “La Caleta” beach A Tarifa (Cadiz). February 2018 |
| Anabaena sp. | BEA 0871B | Gracilariopsis longissima (S.G.Gmelin) Steentorft, L.M. Irvine & Farnham | “La Esperanza” saline B Cadiz. March 2019 |
| Scytonema sp. | BEA 1603B | Gelidium corneum (Hudson) J.V. Lamouroux | “Las Palomas” island C Tarifa (Cadiz). October 2019 |
| Calothrix scopulorum C. Agardh ex Bornet & Flahault | BEA 0796B | Osmundea pinnatifida (Hudson) Stackhouse | “La Araña” beach D Malaga. February 2018 |
| Lyngbya sp. | BEA 1328B | Ceramium rubrum C.Agardh | “La Araña” beach D Malaga. February 2018 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vega, J.; Bonomi-Barufi, J.; Gómez-Pinchetti, J.L.; Figueroa, F.L. Cyanobacteria and Red Macroalgae as Potential Sources of Antioxidants and UV Radiation-Absorbing Compounds for Cosmeceutical Applications. Mar. Drugs 2020, 18, 659. https://doi.org/10.3390/md18120659
Vega J, Bonomi-Barufi J, Gómez-Pinchetti JL, Figueroa FL. Cyanobacteria and Red Macroalgae as Potential Sources of Antioxidants and UV Radiation-Absorbing Compounds for Cosmeceutical Applications. Marine Drugs. 2020; 18(12):659. https://doi.org/10.3390/md18120659
Chicago/Turabian StyleVega, Julia, José Bonomi-Barufi, Juan Luis Gómez-Pinchetti, and Félix L. Figueroa. 2020. "Cyanobacteria and Red Macroalgae as Potential Sources of Antioxidants and UV Radiation-Absorbing Compounds for Cosmeceutical Applications" Marine Drugs 18, no. 12: 659. https://doi.org/10.3390/md18120659
APA StyleVega, J., Bonomi-Barufi, J., Gómez-Pinchetti, J. L., & Figueroa, F. L. (2020). Cyanobacteria and Red Macroalgae as Potential Sources of Antioxidants and UV Radiation-Absorbing Compounds for Cosmeceutical Applications. Marine Drugs, 18(12), 659. https://doi.org/10.3390/md18120659