N-Terminal Acetylation and C-Terminal Amidation of Spirulina platensis-Derived Hexapeptide: Anti-Photoaging Activity and Proteomic Analysis


Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. The Acetylated and Amidated Modification of Hexapeptide
2.3. Cell Viability Using the MTT Method
2.4. Animal Experiment
2.4.1. Animal
2.4.2. UV Irradiation and Peptide Treatment
2.4.3. Histological Examination and Moisture Content Test of Skin
2.4.4. Determination of SOD, CAT, GSH-Px, and MDA in Skin Tissue
2.4.5. Determination of MMP-1 and MMP-3 in Skin Tissue
2.5. Proteomics
2.5.1. Sample Preparation and Labeling
2.5.2. Sample Fractionation and LC–MS/MS Analysis
2.5.3. Data Analysis
2.5.4. Western Blot
2.6. Statistical Analysis
3. Results
3.1. The Toxicity on Hacats and the Protective Effect on the Damaged Hacats
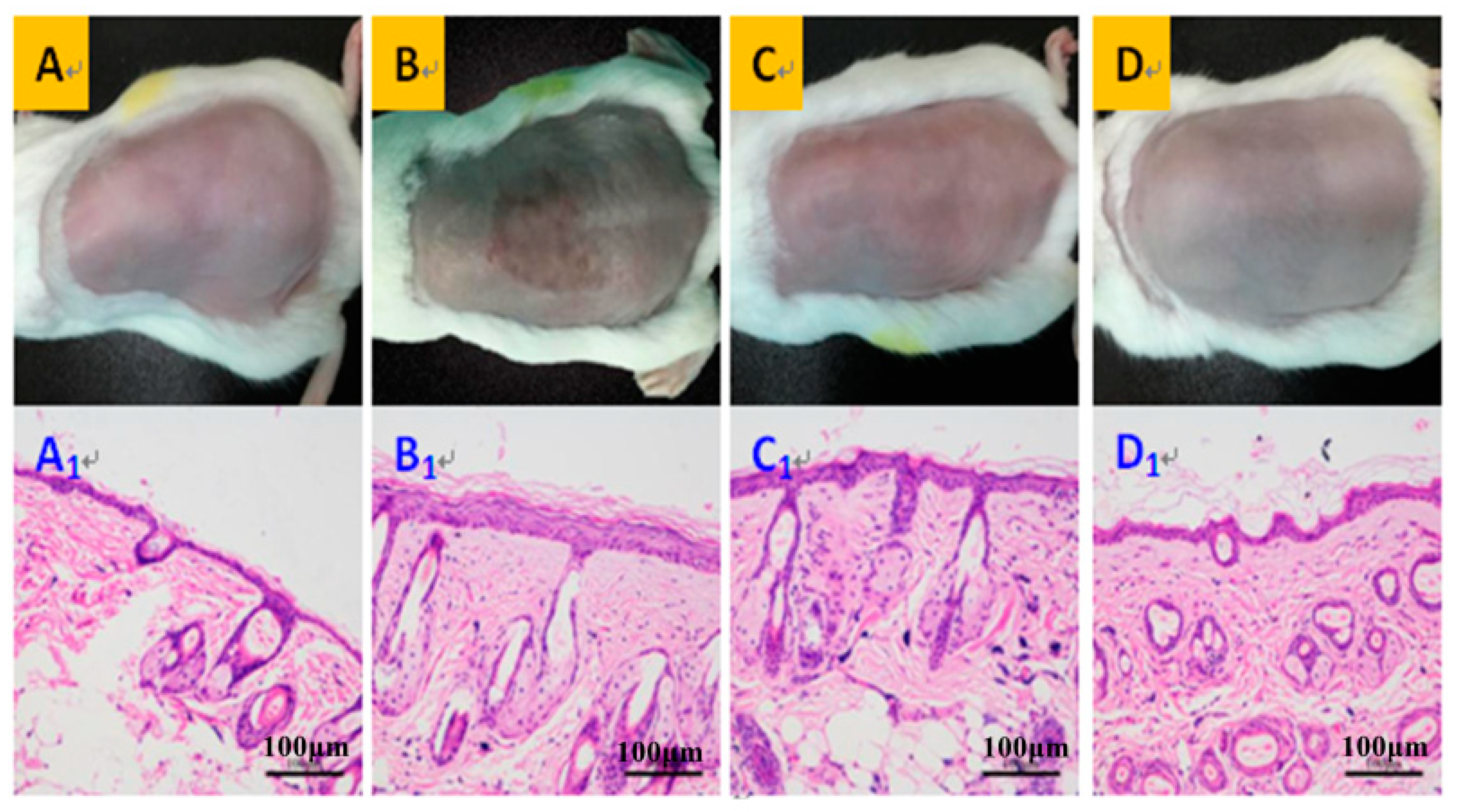
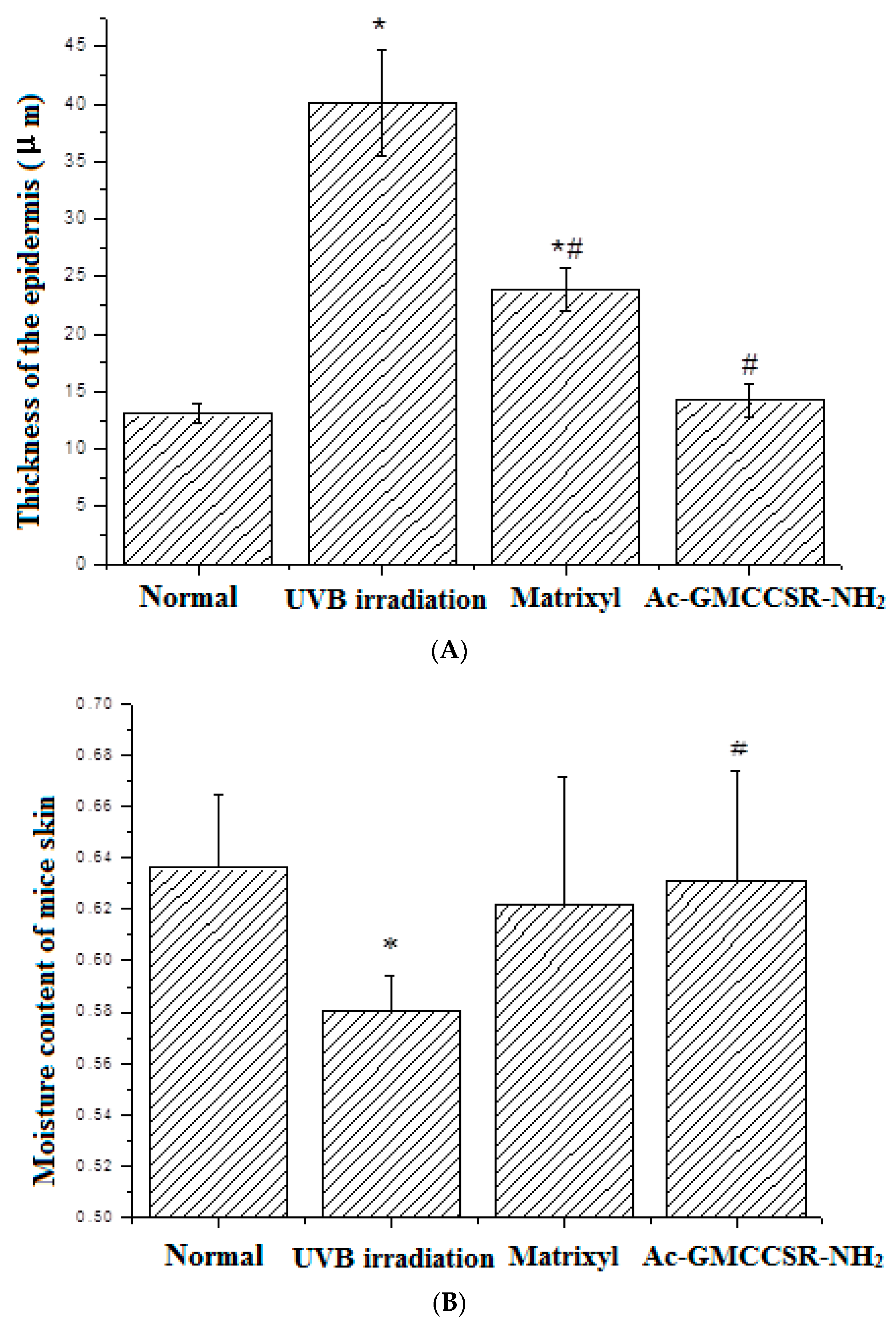
3.2. Morphology, Thickness, and Moisture Content of the Dorsal Skin on Mice
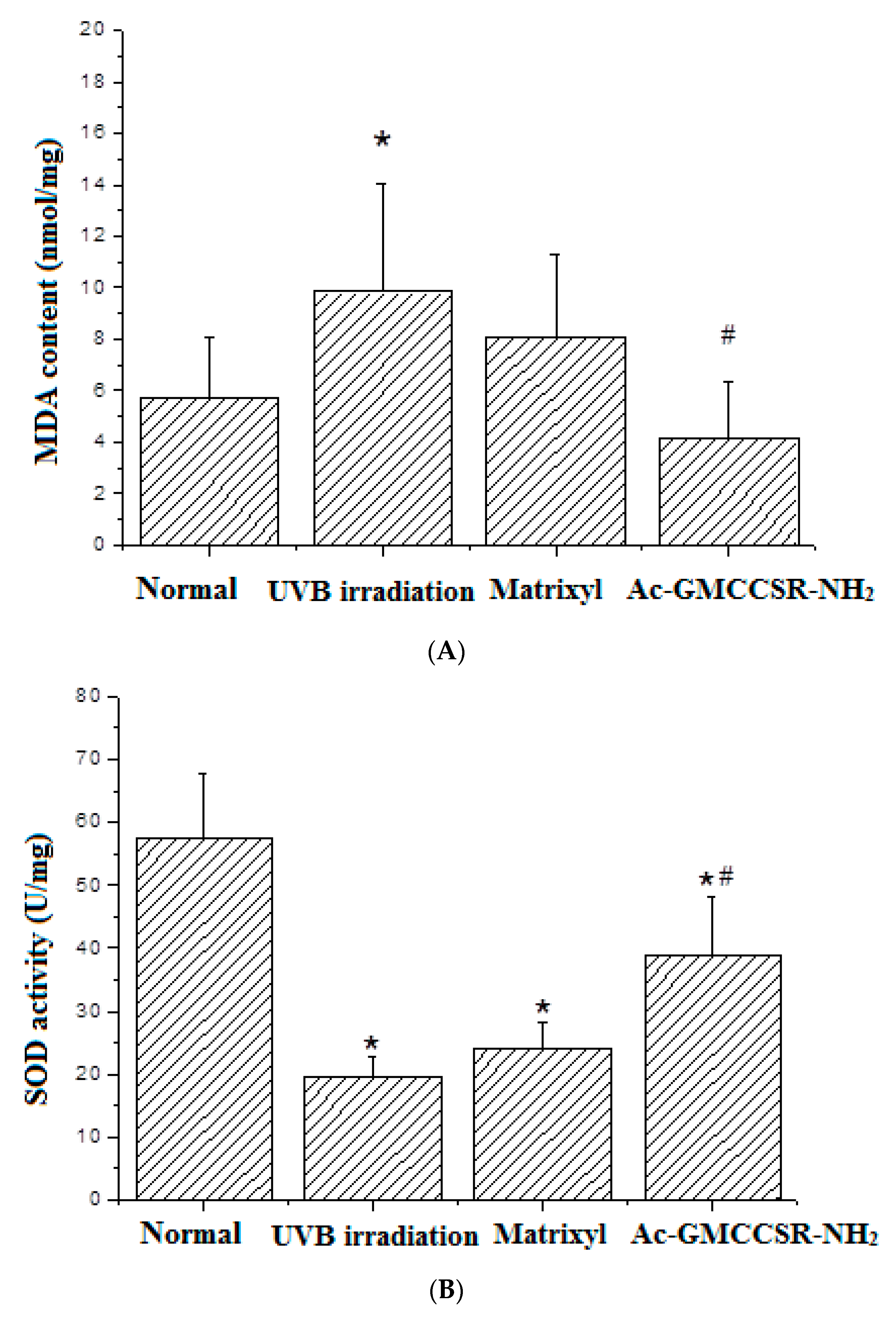
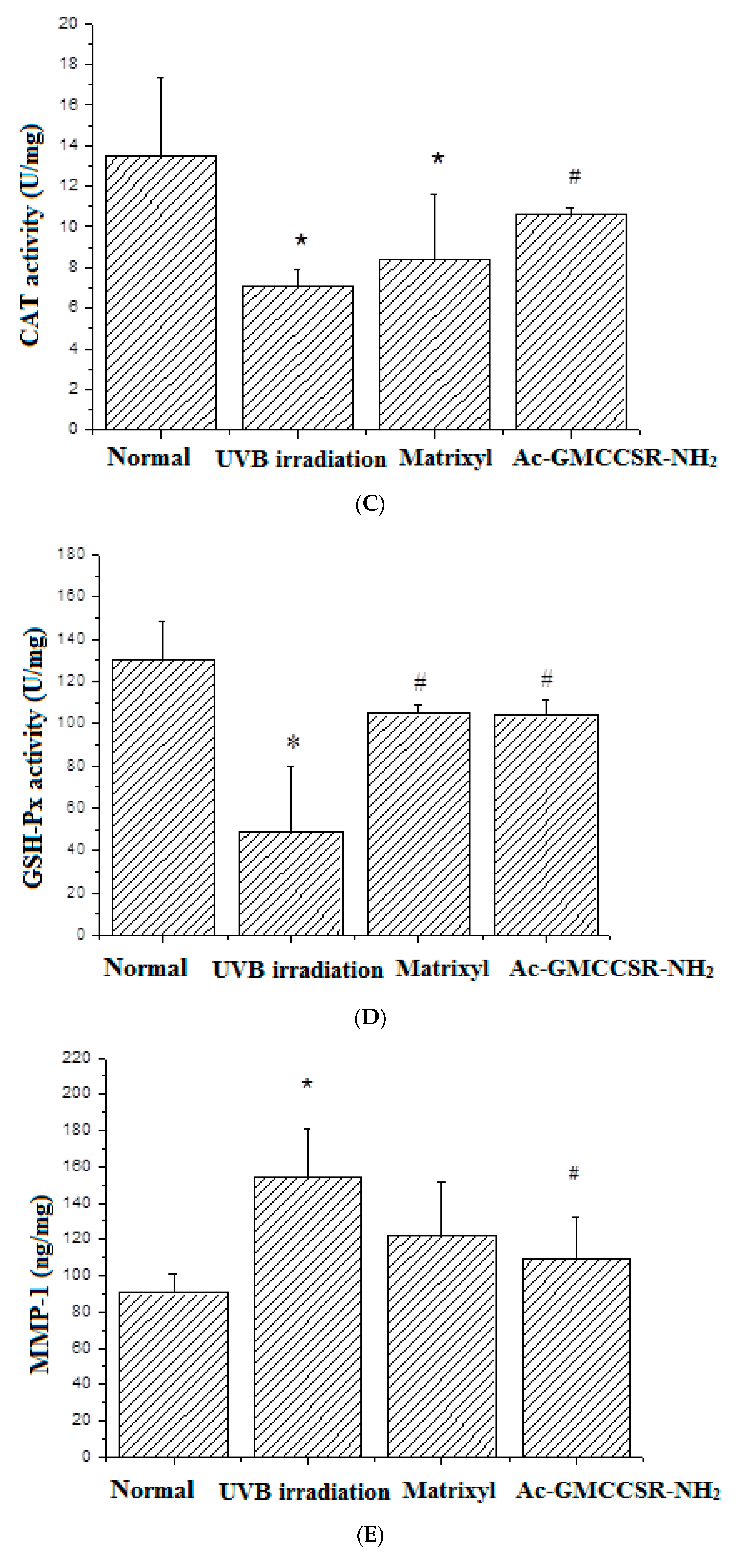
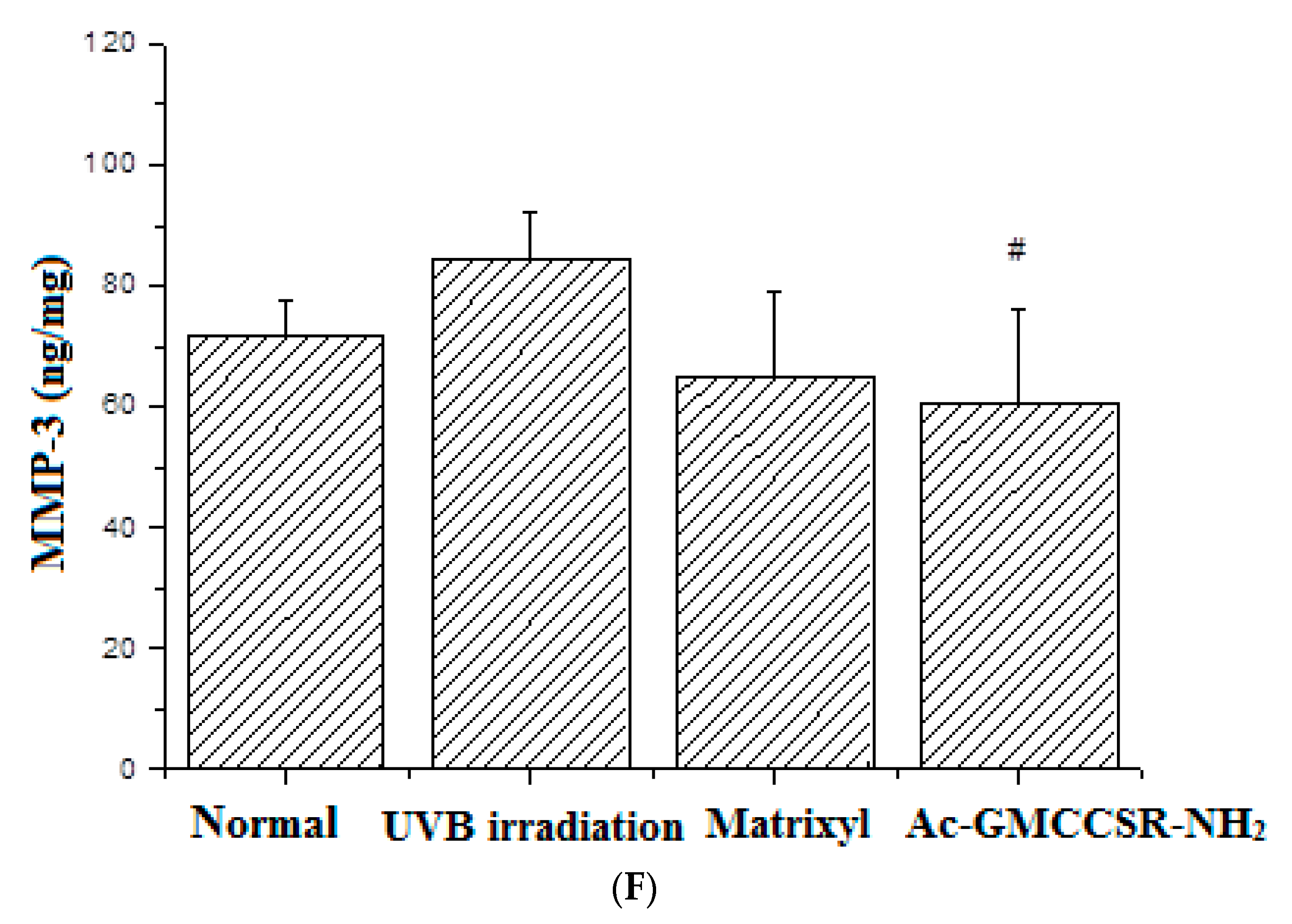
3.3. Effects on MDA, SOD, CAT, GSH-Px, MMP-1, and MMP-3 in Skin Tissue of Mice
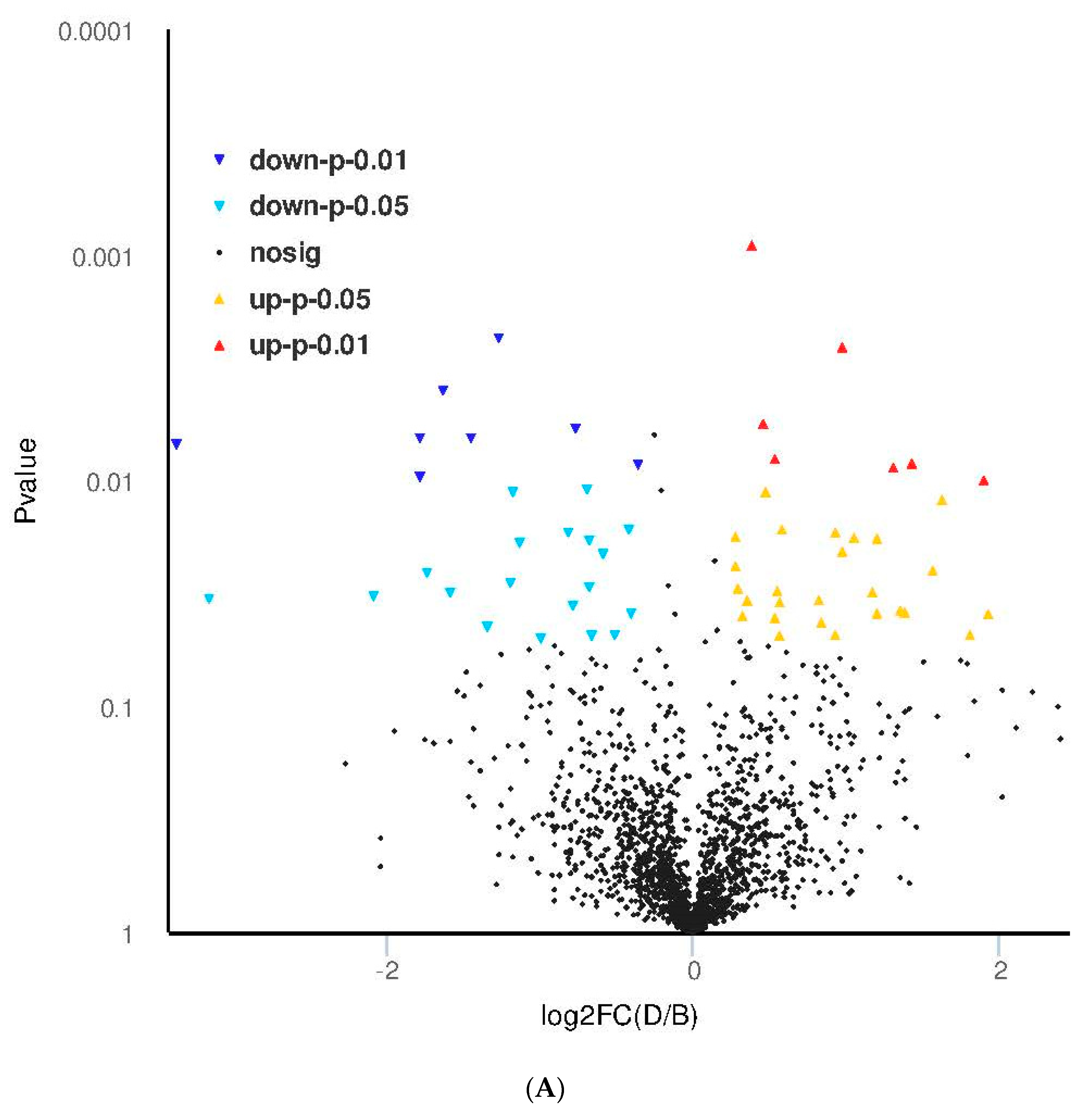
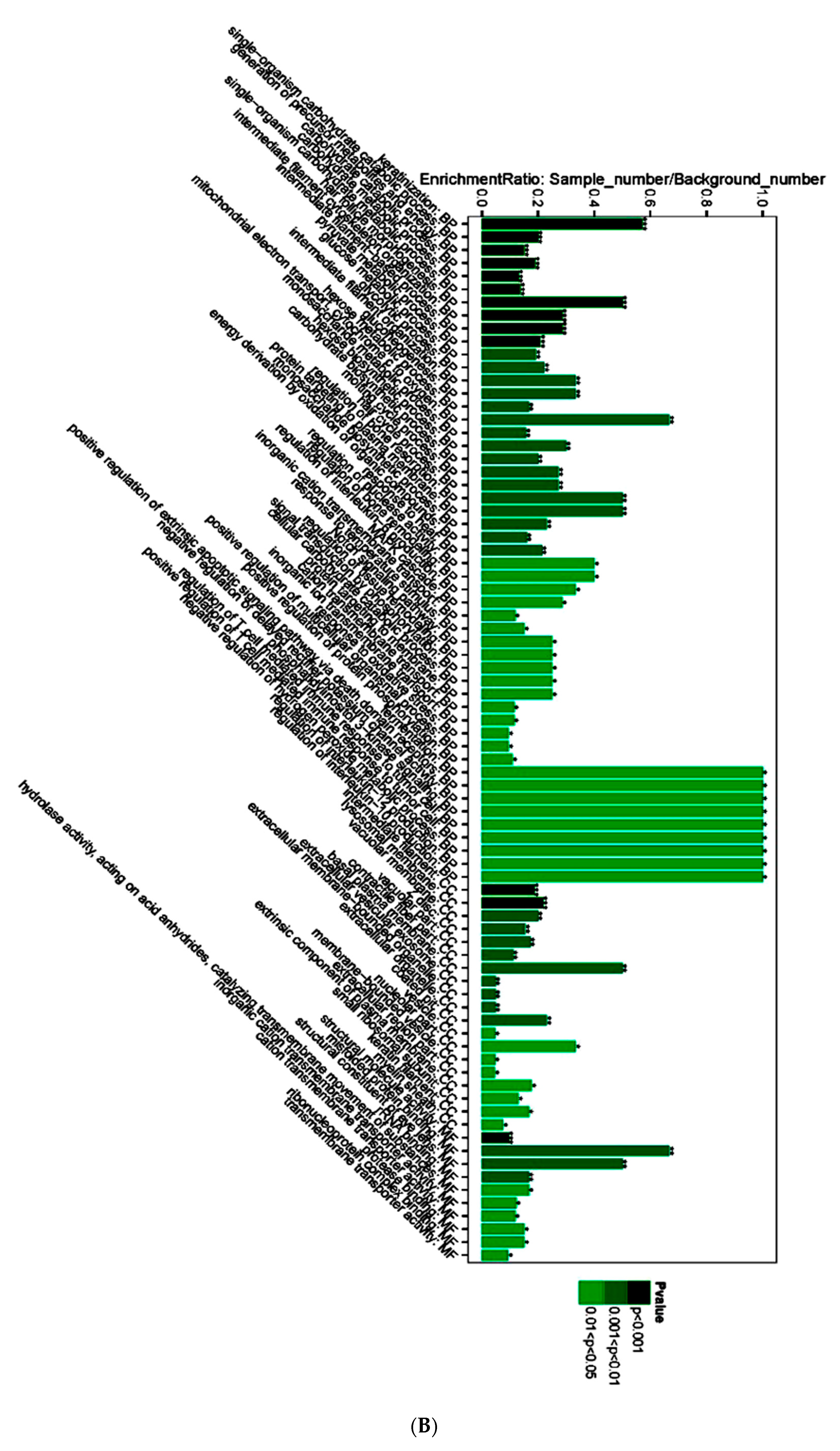
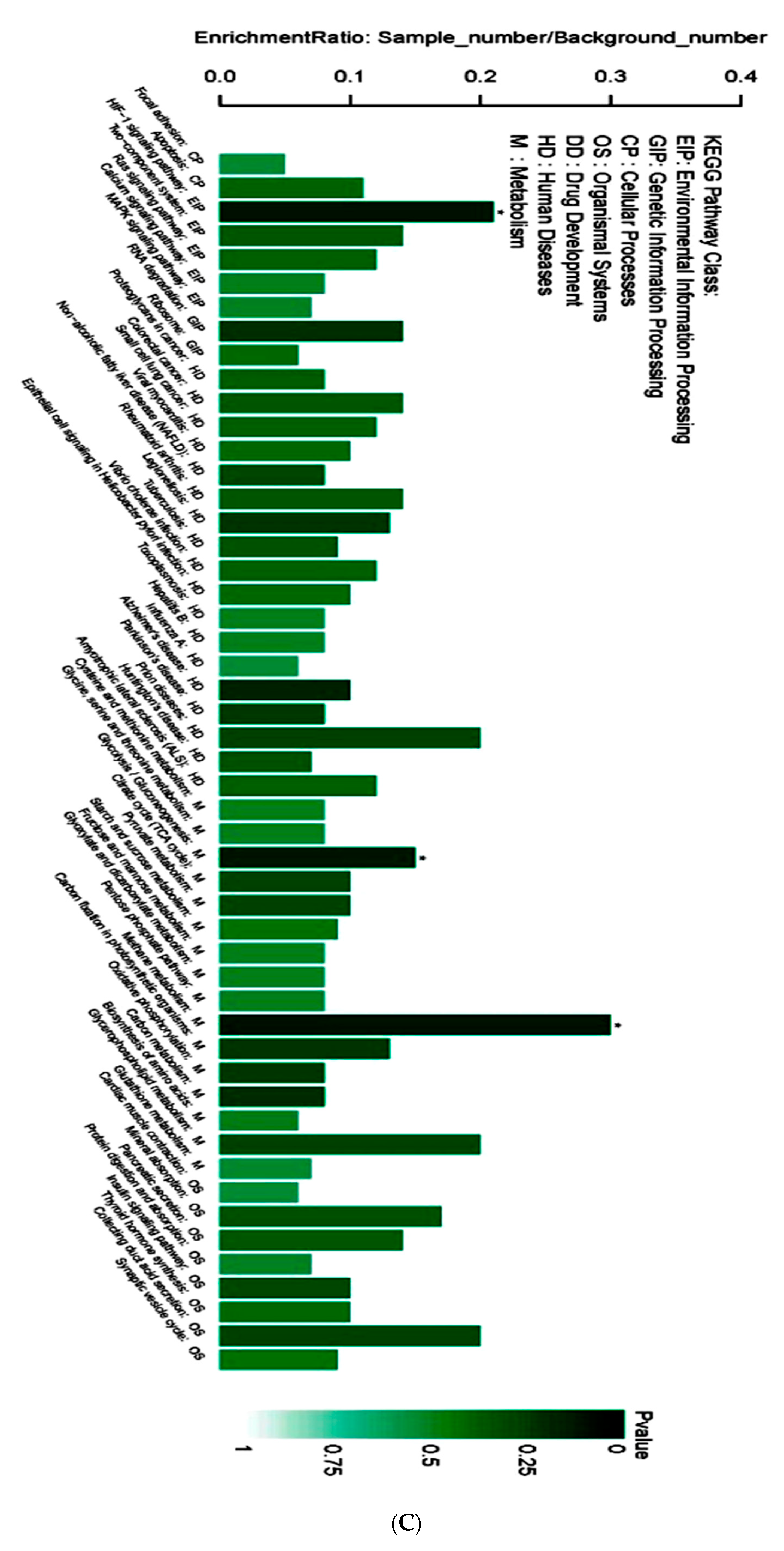
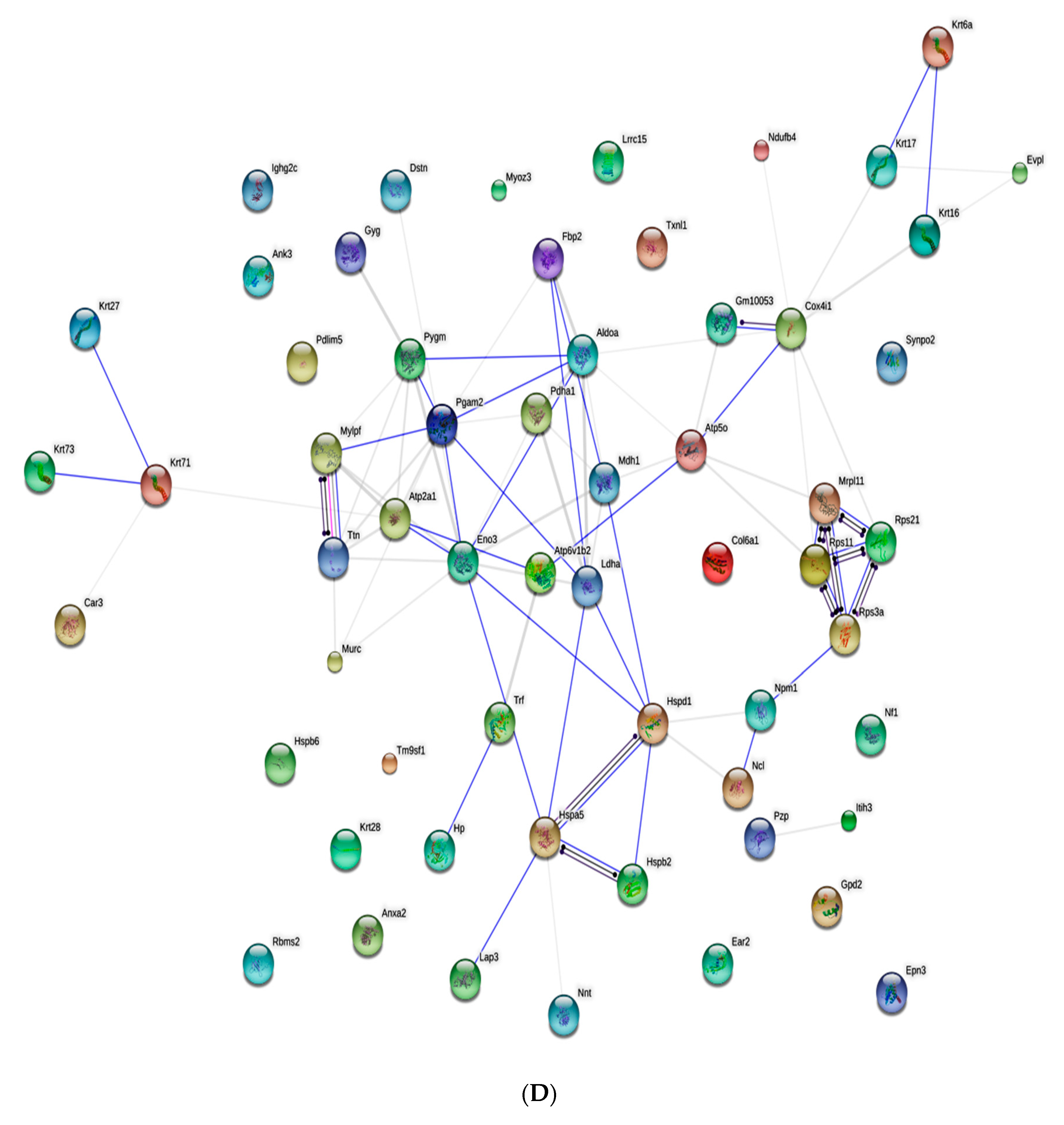
3.4. Proteomic Analysis of Skin Tissue in Mice
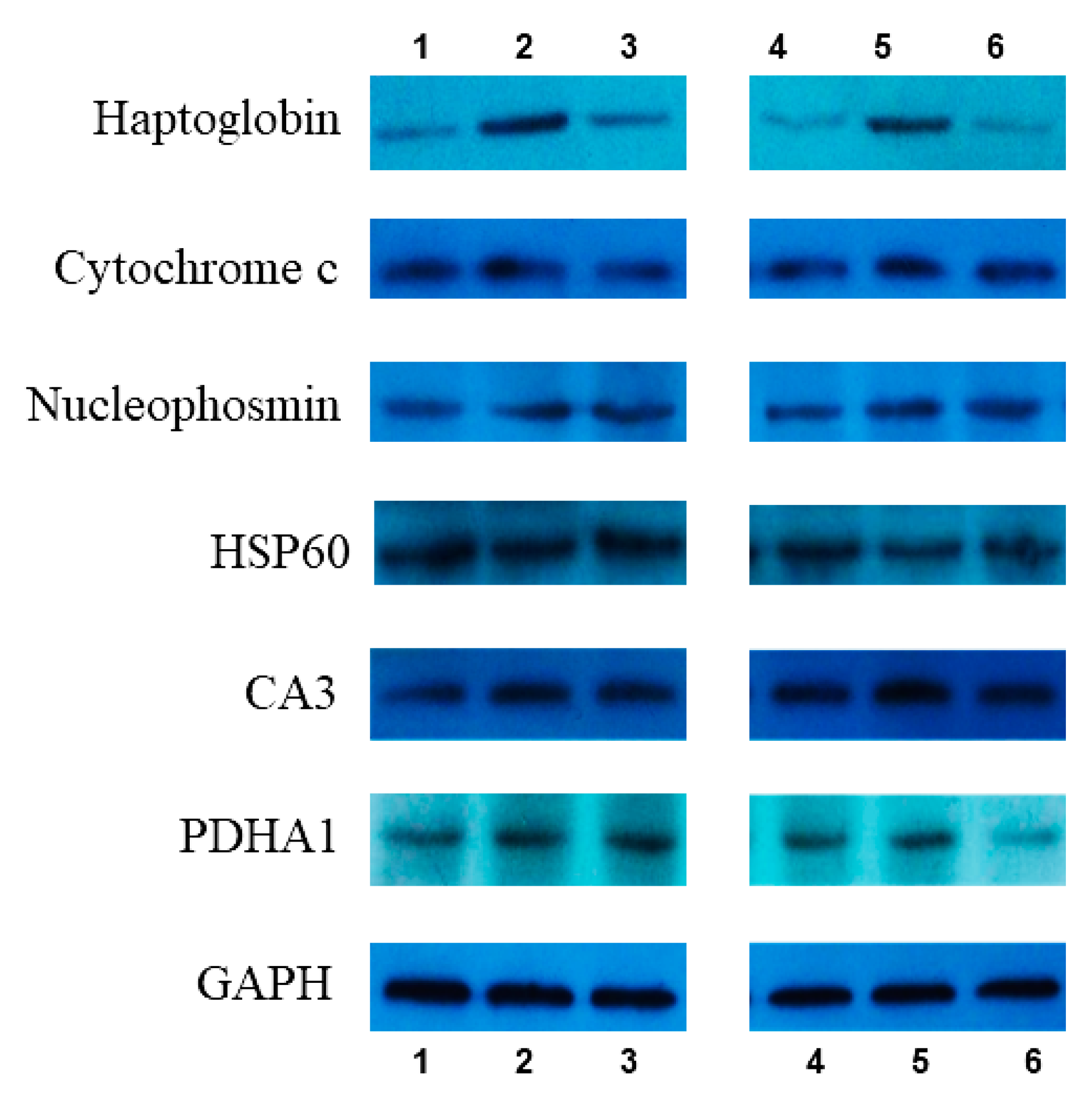
3.5. Western Blot Verification
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Na, C.R.; Wang, S.; Kirsner, R.S.; Federman, D.G. Elderly adults and skin disorders: Common problems for nondermatologists. South Med. J. 2012, 105, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Kong, S.Z.; Shi, X.G.; Feng, X.X.; Li, W.J.; Liu, W.H.; Chen, Z.W.; Xie, J.H.; Lai, X.P.; Zhang, S.X.; Zhang, X.J.; et al. Inhibitory effect of hydroxysafflor yellow a on mouse skin photoaging induced by ultraviolet irradiation. Rejuvenation Res. 2013, 16, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Brenneisen, P.; Sies, H.; Scharffetter-Kochanek, K. Ultraviolet-B irradiation and matrix metalloproteinases: From induction via signaling to initial events. Ann. N. Y. Acad. Sci. 2002, 973, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Debacq-Chainiaux, F.; Borlon, C.; De Hertogh, B.; Remacle, J.; Morvan, P.Y.; Vallée, R.; Toussaint, O. Identification of potential anti-photoageing algal compounds using an in-vitro model of photoageing. J. Pharm Pharmacol. 2006, 58, 1577–1583. [Google Scholar] [CrossRef] [PubMed]
- Neyrinck, A.M.; Taminiau, B.; Walgrave, H.; Daube, G.; Cani, P.D.; Bindels, L.B.; Delzenne, N.M. Spirulina Protects against Hepatic Inflammation in Aging: An Effect Related to the Modulation of the Gut Microbiota? Nutrients 2017, 9, 633. [Google Scholar] [CrossRef] [PubMed]
- Sadek, K.M.; Lebda, M.A.; Nasr, S.M.; Shoukry, M. Spirulina platensis prevents hyperglycemia in rats by modulating gluconeogenesis and apoptosis via modification of oxidative stress and MAPK-pathways. Biomed. Pharmacother. 2017, 92, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Liu, L.; Miron, A.; Klímová, B.; Wan, D.; Kuca, K. The antioxidant, immunomodulatory, and antiinflammatory activities of Spirulina: An overview. Arch. Toxicol. 2016, 90, 1817–1840. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.; Campos, P.M.; Schanzer, S.; Albrecht, S.; Lohan, S.B.; Lademann, J.; Darvin, M.E.; Meinke, M.C. Radical scavenging activity of a sunscreen enriched by antioxidants providing protection in the whole solar spectral range. Eur. J. Pharm Sci. 2017. [Google Scholar] [CrossRef]
- Zeng, Q.H.; Fan, X.D.; Zheng, Q.P.; Wang, J.J.; Zhang, X.W. Anti-oxidant, hemolysis inhibition, and collagen-stimulating activities of a new hexapeptide derived from Arthrospira (Spirulina) platensis. J. Appl. Phycol. 2018, 30, 1655–1665. [Google Scholar] [CrossRef]
- Wallace, R.J. Acetylation of peptides inhibits their degradation by rumen micro-organisms. Br. J. Nutr. 1992, 68, 365–372. [Google Scholar] [CrossRef]
- Kim, K.H.; Seong, B.L. Peptide Amidation: Production of Peptide Hormones in vivo and in vitro. Biotechnol. Bioprocess. Eng. 2001, 6, 244–251. [Google Scholar] [CrossRef]
- Thomas, A. Towards a Functional Understanding of Protein N-Terminal Acetylation. PLoS Biol. 2011, 9, e1001074. [Google Scholar]
- Song, K.H.; Kim, S.B.; Shim, C.K.; Chung, S.J.; Kim, D.D.; Rhee, S.K.; Choi, G.J.; Kim, C.H.; Kim, K. Paracellular permeation-enhancing effect of AT1002 C-terminal amidation in nasal delivery. Drug Des. Devel Ther. 2015, 9, 1815–1823. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, J.M. Young AR the sunburn cell revisited: An update on mechanistic aspects. Photochem. Photobiol. Sci. 2002, 1, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Tsoyi, K.; Park, H.B.; Kim, Y.M.; Chung, J.I.; Shin, S.C.; Shim, H.J.; Lee, W.S.; Seo, H.G.; Lee, J.H.; Chang, K.C.; et al. Protective effect of anthocyanins from black soybean seed coats on UVB-induced apoptotic cell death in vitro and in vivo. J. Agric. Food Chem. 2008, 56, 10600–10605. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lee, J.Y.; Song, K.C.; Kim, J.; Park, J.H.; Chun, K.H.; Hwang, G.S. Protective Effect of Processed Panax ginseng, Sun Ginseng on UVB-irradiated Human Skin Keratinocyte and Human Dermal Fibroblast. J. Ginseng Res. 2012, 36, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.Y.; Wang, P.W.; Huang, C.H.; Chen, M.H.; Wu, Y.R.; Pan, T.L. Skin aging caused by intrinsic or extrinsic processes characterized with functional proteomics. Proteomics 2016, 16, 2718–2731. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.X.; Yu, X.T.; Li, W.J.; Kong, S.Z.; Liu, Y.H.; Zhang, X.; Xian, Y.F.; Zhang, X.J.; Su, Z.R.; Lin, Z.X. Effects of topical application of patchouli alcohol on the UV-induced skin photoaging in mice. Eur. J. Pharm Sci. 2014, 63, 113–123. [Google Scholar] [CrossRef]
- Wu, P.Y.; Huang, C.C.; Chu, Y.; Huang, Y.H.; Lin, P.; Liu, Y.H.; Wen, K.C.; Lin, C.Y.; Hsu, M.C.; Chiang, H.M. Alleviation of Ultraviolet B-Induced Photodamage by Coffea arabica Extract in Human Skin Fibroblasts and Hairless Mouse Skin. Int. J. Mol. Sci. 2017, 18, 782. [Google Scholar] [CrossRef] [PubMed]
- Pillai, S.; Oresajo, C.; Hayward, J. Ultraviolet radiation and skin aging: Roles of reactive oxygen species, inflammation and protease activation, and strategies for prevention of inflammation-induced matrix degradation—A review. Int. J. Cosmet. Sci. 2005, 27, 17–34. [Google Scholar] [CrossRef]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Steinbrenner, H.; Ramos, M.C.; Stuhlmann, D.; Sies, H.; Brenneisen, P. UVA-mediated downregulation of MMP-2 and MMP-9 in human epidermal keratinocytes. Biochem. Biophys. Res. Commun. 2003, 308, 486–491. [Google Scholar] [CrossRef]
- Lee, Y.R.; Noh, E.M.; Jeong, E.Y.; Yun, S.K.; Jeong, Y.J.; Kim, J.H.; Kwon, K.B.; Kim, B.S.; Lee, S.H.; Park, C.S.; et al. Cordycepin inhibits UVB-induced matrix metalloproteinase expression by suppressing the NF-kappaB pathway in human dermal fibroblasts. Exp. Mol. Med. 2009, 41, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Park, Y.G.; Lee, H.J.; Lim, S.J.; Nho, C.W. Youngiasides A and C Isolated from Youngia denticulatum Inhibit UVB-Induced MMP Expression and Promote Type I Procollagen Production via Repression of MAPK/AP-1/NF-κB and Activation of AMPK/Nrf2 in HaCaT Cells and Human Dermal Fibroblasts. J. Agric. Food Chem. 2015, 63, 5428–5438. [Google Scholar] [CrossRef]
- Wu, C.L.; Chou, H.C.; Cheng, C.S.; Li, J.M.; Lin, S.T.; Chen, Y.W.; Chan, H.L. Proteomic analysis of UVB-induced protein expression-and redox-dependent changes in skin fibroblasts using lysine- and cysteine-labeling two-dimensional difference gel electrophoresis. J. Proteom. 2012, 75, 1991–2014. [Google Scholar] [CrossRef]
- Bertrand-Vallery, V.; Belot, N.; Dieu, M.; Delaive, E.; Ninane, N.; Demazy, C.; Raes, M.; Salmon, M.; Poumay, Y.; Debacq-Chainiaux, F.; et al. Proteomic profiling of human keratinocytes undergoing UVB-induced alternative differentiation reveals TRIpartite Motif Protein 29 as a survival factor. PLoS ONE 2010, 5, e10462. [Google Scholar] [CrossRef]
- Wei, Y.H.; Wu, S.B.; Ma, Y.S.; Lee, H.C. Respiratory function decline and DNA mutation in mitochondria, oxidative stress and altered gene expression during aging. Chang Gung Med. J. 2009, 32, 113–132. [Google Scholar]
- Hiranandani, N.; Bupha-Intr, T.; Janssen, P.M. SERCA overexpression reduces hydroxyl radical injury in murine myocardium. Am. J. Physiol Heart Circ. Physiol. 2006, 291, H3130–H3135. [Google Scholar] [CrossRef]
- Lisi, S.; Gamucci, O.; Vottari, T.; Scabia, G.; Funicello, M.; Marchi, M.; Galli, G.; Arisi, I.; Brandi, R.; D’Onofrio, M.; et al. Obesity-associated hepatosteatosis and impairment of glucose homeostasis are attenuated by haptoglobin deficiency. Diabetes 2011, 60, 2496–2505. [Google Scholar] [CrossRef]
- Kinn, P.M.; Holdren, G.O.; Westermeyer, B.A.; Abuissa, M.; Fischer, C.L.; Fairley, J.A.; Brogden, K.A.; Brogden, N.K. Age-dependent variation in cytokines, chemokines, and biologic analytes rinsed from the surface of healthy human skin. Sci. Rep. 2015, 5, 10472. [Google Scholar] [CrossRef]
- Depianto, D.; Kerns, M.L.; Dlugosz, A.A.; Coulombe, P.A. Keratin 17 promotes epithelial proliferation and tumor growth by polarizing the immune response in skin. Nat. Genet. 2010, 42, 910–914. [Google Scholar] [CrossRef] [PubMed]
- Wawersik, M.J.; Mazzalupo, S.; Nguyen, D.; Coulombe, P.A. Increased levels of keratin 16 alter epithelialization potential of mouse skin keratinocytes in vivo and ex vivo. Mol. Biol. Cell. 2001, 12, 3439–3450. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | B | D | FC (D/B) | log2FC (D/B) | p-Value | Significant | Regulate |
|---|---|---|---|---|---|---|---|
| ANK3_MOUSE | 0.91623 | 1.196753 | 1.306171 | 0.385343 | 0.000904 | yes | up |
| CO6A1_MOUSE | 0.827951 | 0.345235 | 0.416976 | −1.26197 | 0.002326 | yes | down |
| HSPB6_MOUSE | 0.625411 | 1.224668 | 1.95818 | 0.969513 | 0.002556 | yes | up |
| RS21_MOUSE | 2.443534 | 0.794362 | 0.325087 | −1.6211 | 0.003971 | yes | down |
| GPDM_MOUSE | 0.82043 | 1.132509 | 1.380384 | 0.46507 | 0.00559 | yes | up |
| RS3A_MOUSE | 1.185781 | 0.698351 | 0.588937 | −0.76381 | 0.00586 | yes | down |
| CH60_MOUSE | 1.343677 | 0.492071 | 0.366212 | −1.44925 | 0.00646 | yes | down |
| K1C17_MOUSE | 1.31876 | 0.385973 | 0.292679 | −1.77261 | 0.006467 | yes | down |
| HPT_MOUSE | 3.841142 | 0.372877 | 0.097075 | −3.36476 | 0.006876 | yes | down |
| NF1_MOUSE | 0.887495 | 1.288372 | 1.451696 | 0.537739 | 0.007991 | yes | up |
| ENOB_MOUSE | 0.373393 | 1.004882 | 2.69122 | 1.42826 | 0.008388 | yes | up |
| A2M_MOUSE | 1.374173 | 1.076648 | 0.783488 | −0.35202 | 0.008462 | yes | down |
| CAH3_MOUSE | 0.402064 | 0.995416 | 2.475768 | 1.307876 | 0.008729 | yes | up |
| K1C28_MOUSE | 2.180505 | 0.634469 | 0.290974 | −1.78104 | 0.009607 | yes | down |
| PYGM_MOUSE | 0.245837 | 0.912978 | 3.713754 | 1.892878 | 0.00997 | yes | up |
| NPM_MOUSE | 1.63725 | 1.019672 | 0.622795 | -0.68317 | 0.010896 | yes | down |
| LDHA_MOUSE | 0.724713 | 1.004712 | 1.386359 | 0.471301 | 0.011192 | yes | up |
| TRFE_MOUSE | 1.653381 | 0.731186 | 0.442237 | −1.17711 | 0.011198 | yes | down |
| ODPA_MOUSE | 0.34843 | 1.081721 | 3.104561 | 1.634389 | 0.012074 | yes | up |
| MURC_MOUSE | 0.539877 | 0.805455 | 1.491924 | 0.577174 | 0.016378 | yes | up |
| TXNL1_MOUSE | 1.253474 | 0.942139 | 0.751623 | −0.41192 | 0.016452 | yes | down |
| EPN3_MOUSE | 0.822867 | 1.572029 | 1.910429 | 0.933897 | 0.016926 | yes | up |
| ANXA2_MOUSE | 0.78758 | 0.451204 | 0.572899 | −0.80365 | 0.01695 | yes | down |
| MDHC_MOUSE | 0.737936 | 0.899583 | 1.219054 | 0.285762 | 0.017653 | yes | up |
| SYNP2_MOUSE | 0.546814 | 1.127627 | 2.062178 | 1.044169 | 0.01788 | yes | up |
| ATPO_MOUSE | 0.899583 | 2.080248 | 2.312457 | 1.209427 | 0.017984 | yes | up |
| DEST_MOUSE | 1.349091 | 0.843344 | 0.62512 | −0.6778 | 0.018374 | yes | down |
| VATB2_MOUSE | 1.616824 | 0.737951 | 0.45642 | −1.13156 | 0.018838 | yes | down |
| MLRS_MOUSE | 0.644024 | 1.264857 | 1.963991 | 0.973788 | 0.02064 | yes | up |
| AMPL_MOUSE | 1.507006 | 1.005905 | 0.667486 | −0.58319 | 0.021085 | yes | down |
| GLYG_MOUSE | 0.676148 | 0.824278 | 1.21908 | 0.285793 | 0.023871 | yes | up |
| AT2A1_MOUSE | 2.670282 | 7.878805 | 2.950552 | 1.560985 | 0.025035 | yes | up |
| K1C27_MOUSE | 1.387947 | 0.415728 | 0.299527 | −1.73924 | 0.025627 | yes | down |
| ITIH3_MOUSE | 1.978311 | 0.867782 | 0.438648 | −1.18887 | 0.028266 | yes | down |
| RS11_MOUSE | 0.805592 | 0.505846 | 0.627919 | −0.67135 | 0.029542 | yes | down |
| RM11_MOUSE | 0.883314 | 1.086471 | 1.229995 | 0.298652 | 0.029977 | yes | up |
| RBMS2_MOUSE | 0.739031 | 1.081721 | 1.463701 | 0.549621 | 0.030714 | yes | up |
| CYC_MOUSE | 0.594349 | 1.33729 | 2.250008 | 1.16993 | 0.031063 | yes | up |
| K1C16_MOUSE | 1.231848 | 0.41048 | 0.333223 | −1.58544 | 0.031209 | yes | down |
| K2C71_MOUSE | 1.331865 | 0.313541 | 0.235415 | −2.08672 | 0.032436 | yes | down |
| K2C6A_MOUSE | 3.102133 | 0.347336 | 0.111967 | −3.15885 | 0.033321 | yes | down |
| HSPB2_MOUSE | 0.563349 | 1.001528 | 1.777809 | 0.8301 | 0.033797 | yes | up |
| NNTM_MOUSE | 0.973119 | 1.247436 | 1.281895 | 0.358278 | 0.033941 | yes | up |
| COX41_MOUSE | 1.271059 | 1.889273 | 1.486377 | 0.5718 | 0.034524 | yes | up |
| GCAB_MOUSE | 0.542972 | 0.317315 | 0.584403 | −0.77496 | 0.035724 | yes | down |
| ALDOA_MOUSE | 0.217516 | 0.558516 | 2.567704 | 1.360479 | 0.03765 | yes | up |
| PDLI5_MOUSE | 0.507804 | 1.323852 | 2.607016 | 1.382399 | 0.038285 | yes | up |
| PGAM2_MOUSE | 0.544537 | 1.247436 | 2.290818 | 1.195863 | 0.038803 | yes | up |
| NUCL_MOUSE | 1.214212 | 0.917008 | 0.755229 | −0.40501 | 0.038826 | yes | down |
| TITIN_MOUSE | 0.525364 | 1.998309 | 3.803666 | 1.927391 | 0.0389 | yes | up |
| NDUB4_MOUSE | 0.711696 | 0.887495 | 1.247013 | 0.318477 | 0.039585 | yes | up |
| TM9S1_MOUSE | 0.661394 | 0.956005 | 1.44544 | 0.531508 | 0.040421 | yes | up |
| LRC15_MOUSE | 0.769529 | 1.380911 | 1.794488 | 0.843572 | 0.042306 | yes | up |
| GRP78_MOUSE | 1.316223 | 0.519738 | 0.39487 | −1.34055 | 0.044264 | yes | down |
| MYOZ3_MOUSE | 0.498299 | 0.948807 | 1.90409 | 0.929101 | 0.0481 | yes | up |
| EVPL_MOUSE | 1.009253 | 0.708314 | 0.70182 | −0.51083 | 0.048194 | yes | down |
| ECP3_MOUSE | 0.732769 | 2.571728 | 3.509603 | 1.811308 | 0.048229 | yes | up |
| KR151_MOUSE | 0.748812 | 0.476557 | 0.636418 | −0.65195 | 0.048479 | yes | down |
| F16P2_MOUSE | 0.676664 | 1.004627 | 1.484676 | 0.570148 | 0.04869 | yes | up |
| K2C73_MOUSE | 1.303273 | 0.655997 | 0.503346 | −0.99038 | 0.049896 | yes | down |
| Protein Name | D/B | |
|---|---|---|
| iTRAQ | WB | |
| Haptoglobin | −3.36 | −2.33 |
| Cytochrome c | 1.17 | 1.23 |
| Nucleophosmin | −0.68 | −1.22 |
| HSP60 | −1.45 | −1.16 |
| CA3 | 1.31 | 0.75 |
| PDHA1 | 1.63 | 1.04 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, Q.; Jiang, J.; Wang, J.; Zhou, Q.; Zhang, X. N-Terminal Acetylation and C-Terminal Amidation of Spirulina platensis-Derived Hexapeptide: Anti-Photoaging Activity and Proteomic Analysis. Mar. Drugs 2019, 17, 520. https://doi.org/10.3390/md17090520
Zeng Q, Jiang J, Wang J, Zhou Q, Zhang X. N-Terminal Acetylation and C-Terminal Amidation of Spirulina platensis-Derived Hexapeptide: Anti-Photoaging Activity and Proteomic Analysis. Marine Drugs. 2019; 17(9):520. https://doi.org/10.3390/md17090520
Chicago/Turabian StyleZeng, Qiaohui, Jianguo Jiang, Jingjing Wang, Qiuchan Zhou, and Xuewu Zhang. 2019. "N-Terminal Acetylation and C-Terminal Amidation of Spirulina platensis-Derived Hexapeptide: Anti-Photoaging Activity and Proteomic Analysis" Marine Drugs 17, no. 9: 520. https://doi.org/10.3390/md17090520
APA StyleZeng, Q., Jiang, J., Wang, J., Zhou, Q., & Zhang, X. (2019). N-Terminal Acetylation and C-Terminal Amidation of Spirulina platensis-Derived Hexapeptide: Anti-Photoaging Activity and Proteomic Analysis. Marine Drugs, 17(9), 520. https://doi.org/10.3390/md17090520