Characteristics of the Copper,Zinc Superoxide Dismutase of a Hadal Sea Cucumber (Paelopatides sp.) from the Mariana Trench

Abstract
:1. Introduction
2. Results
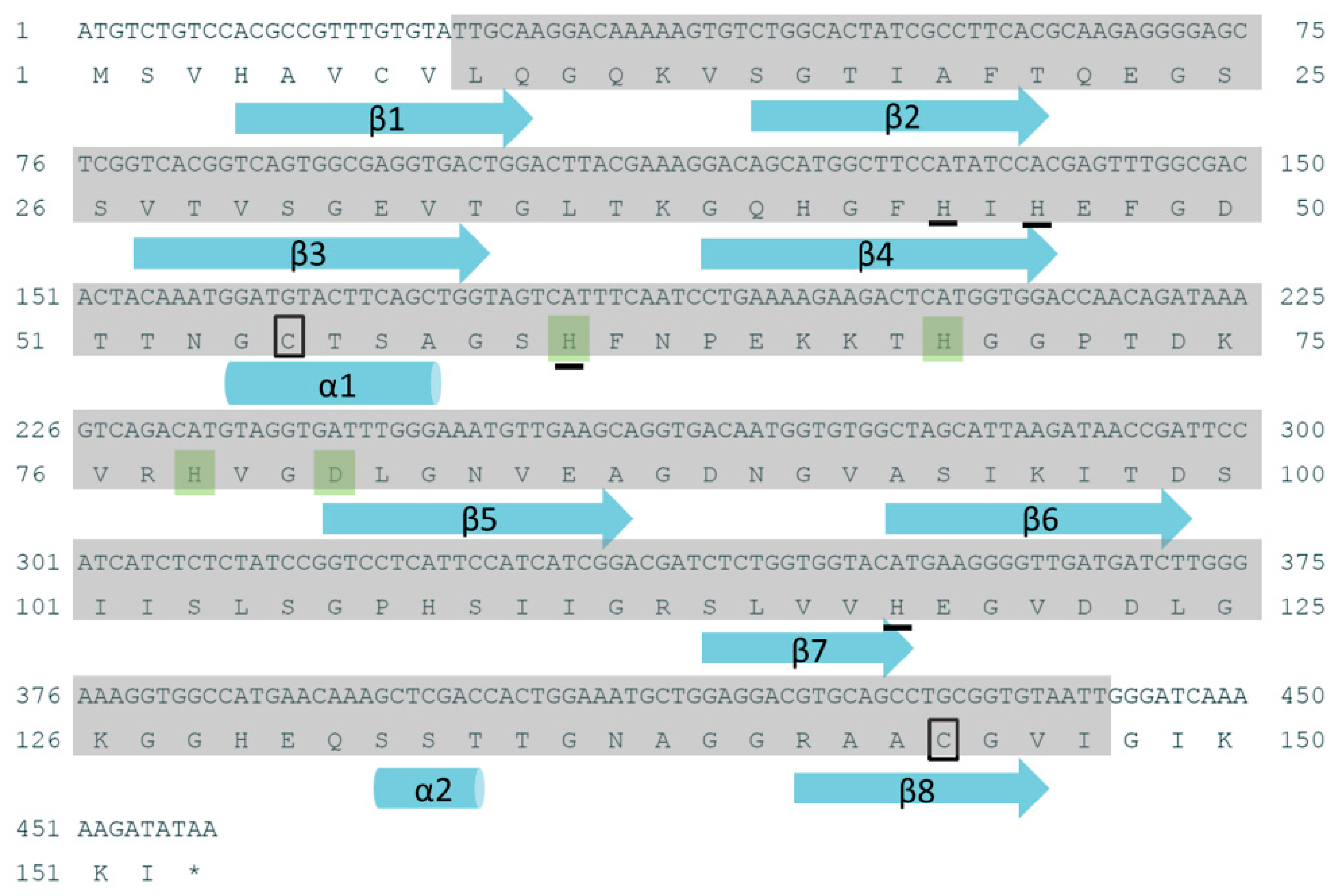
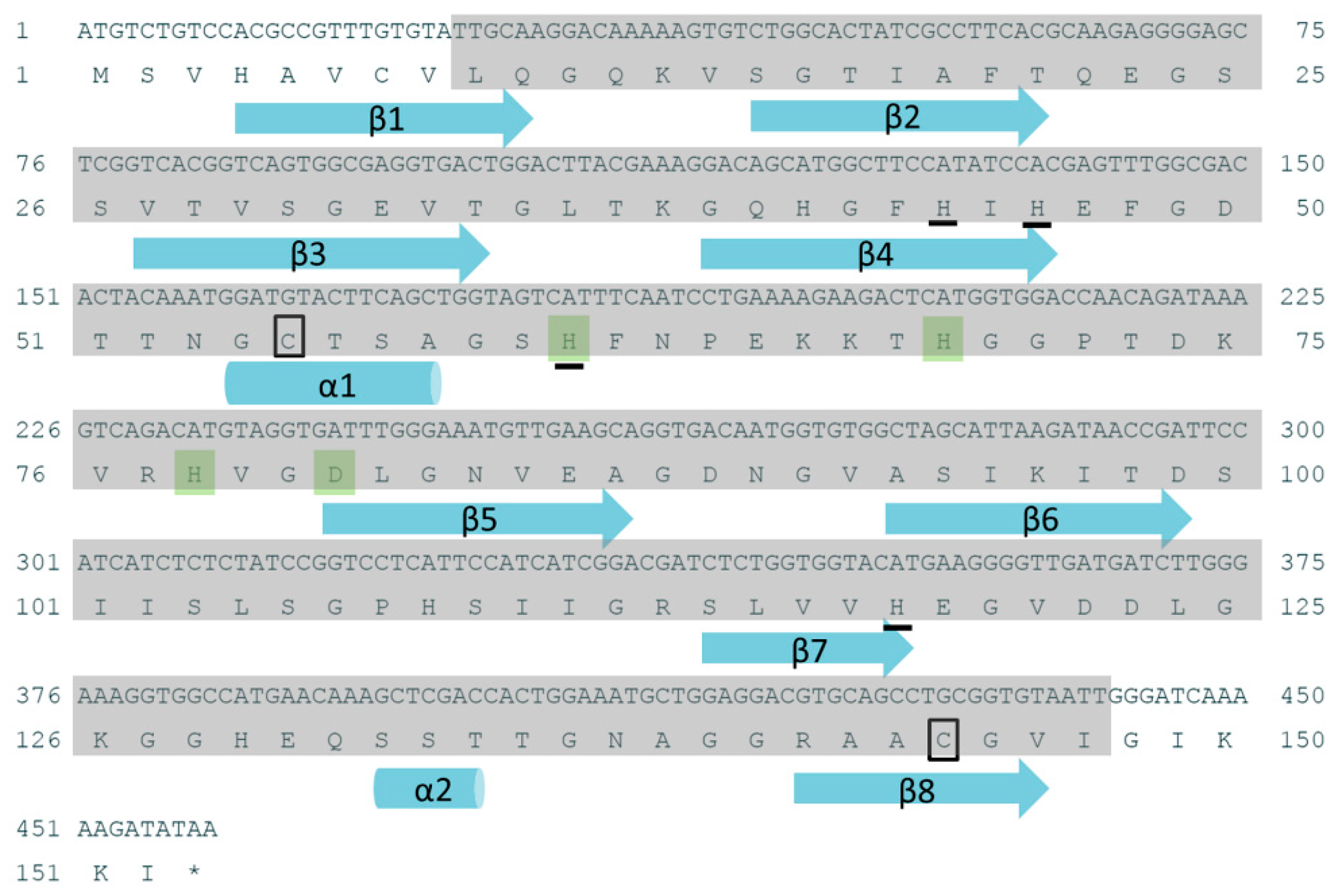
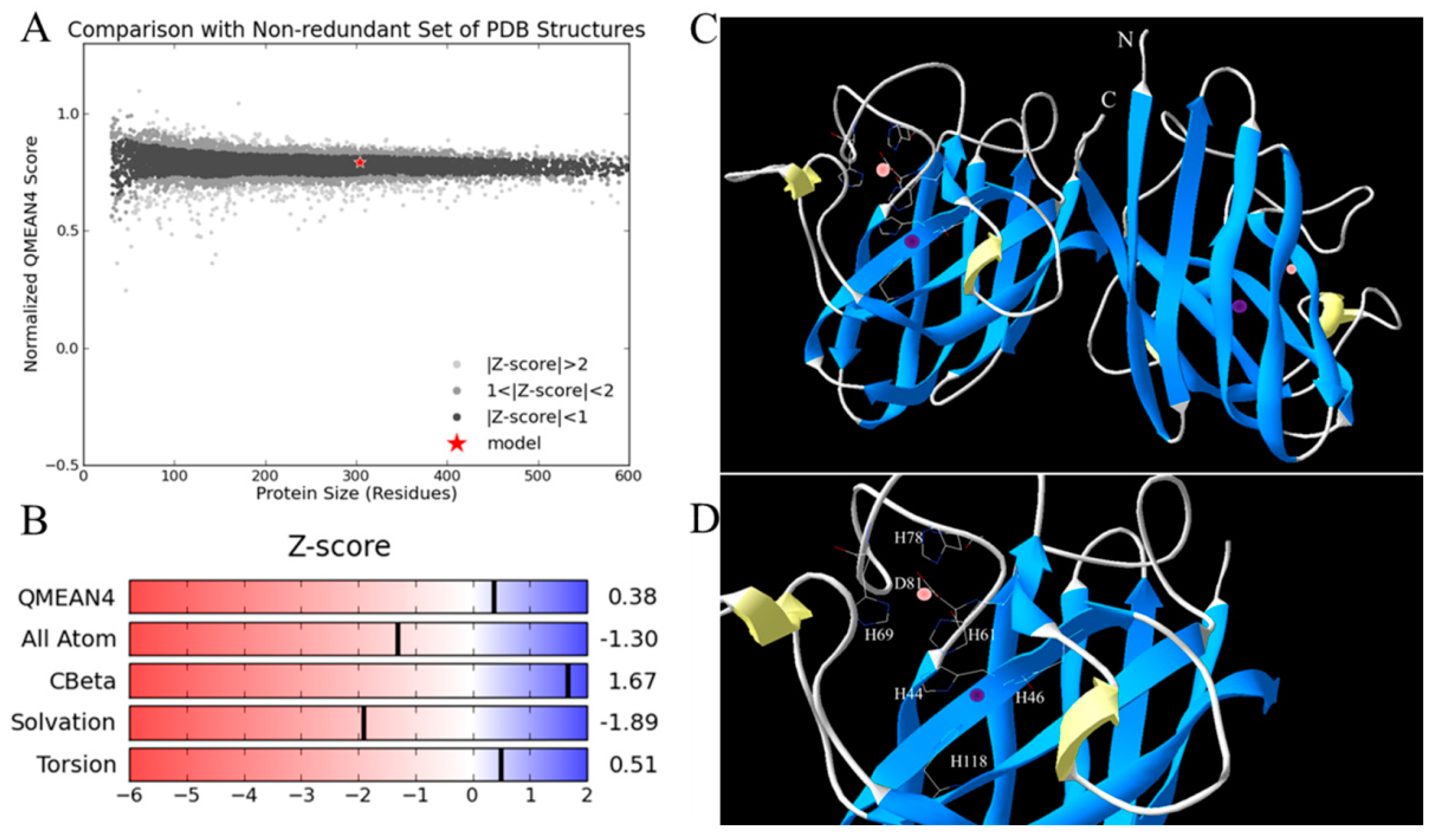
2.1. Sequence Analysis
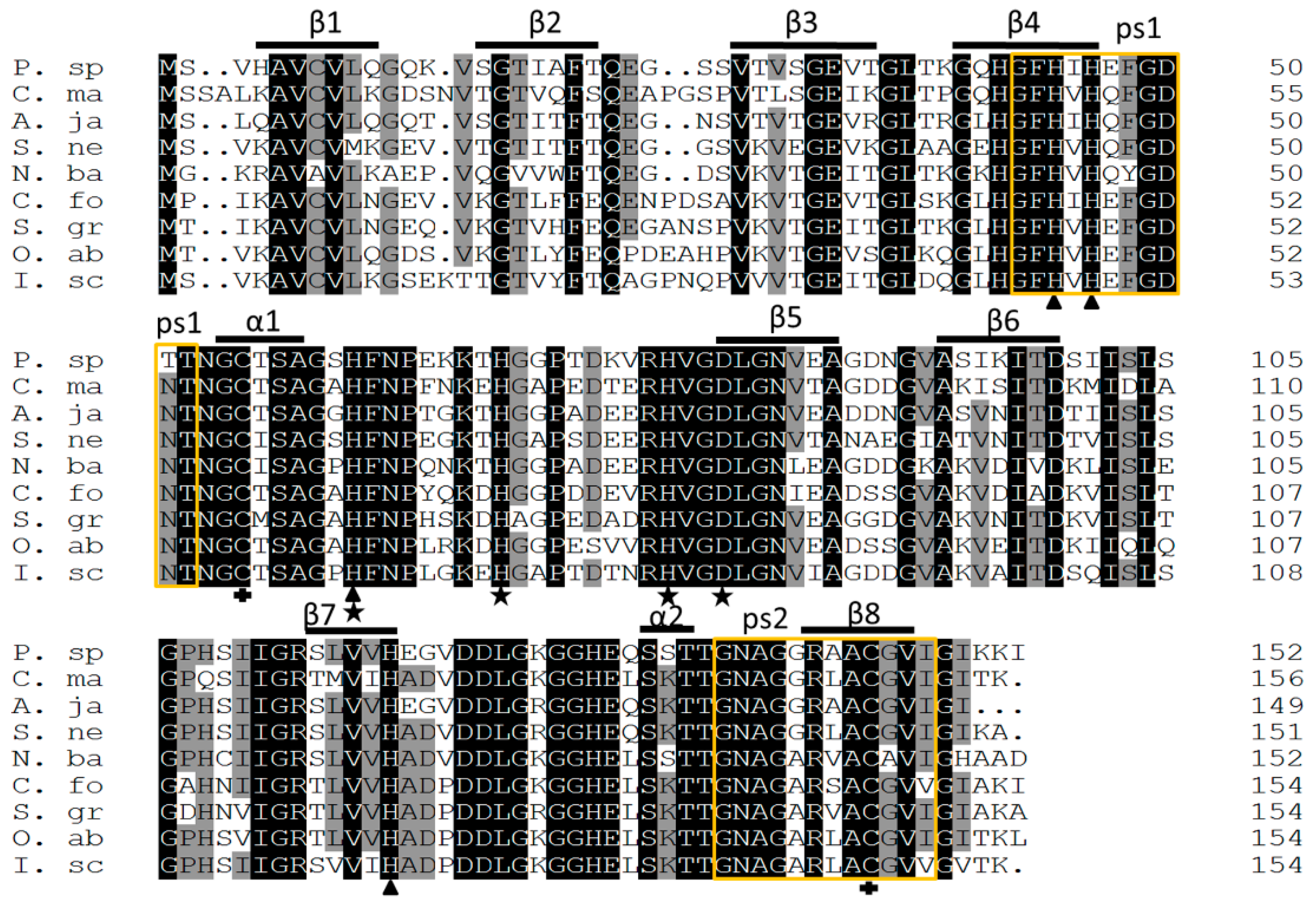
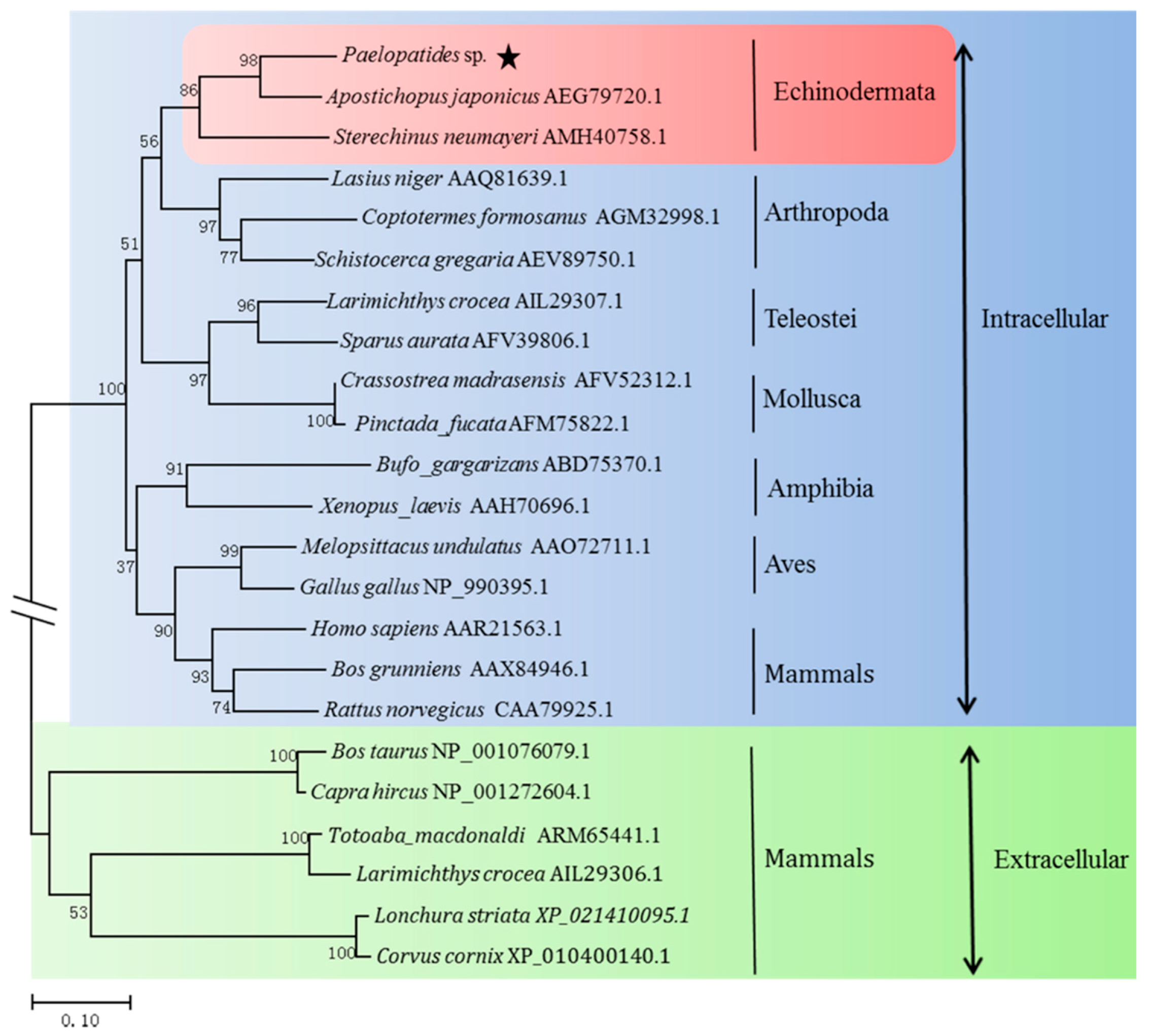
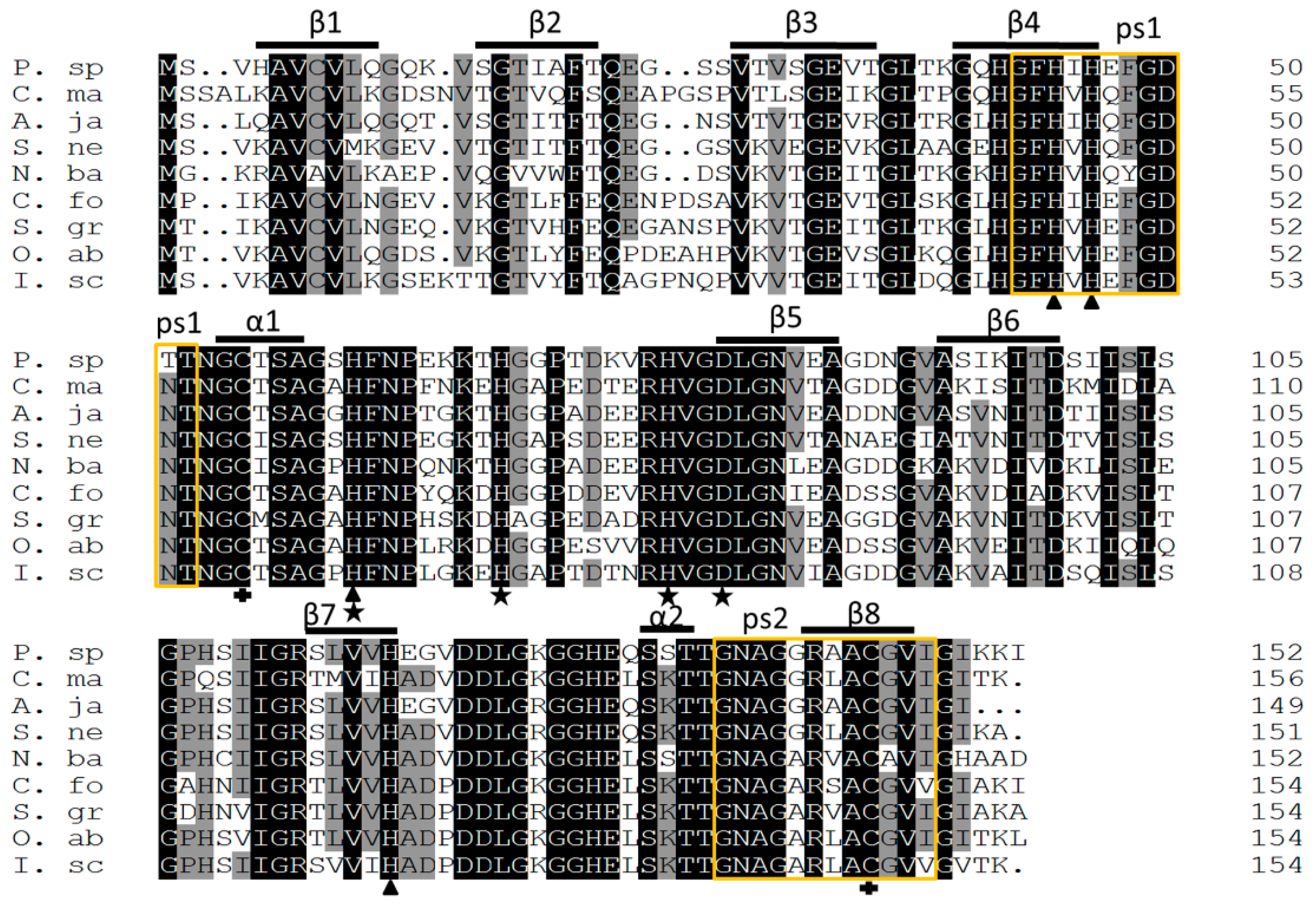
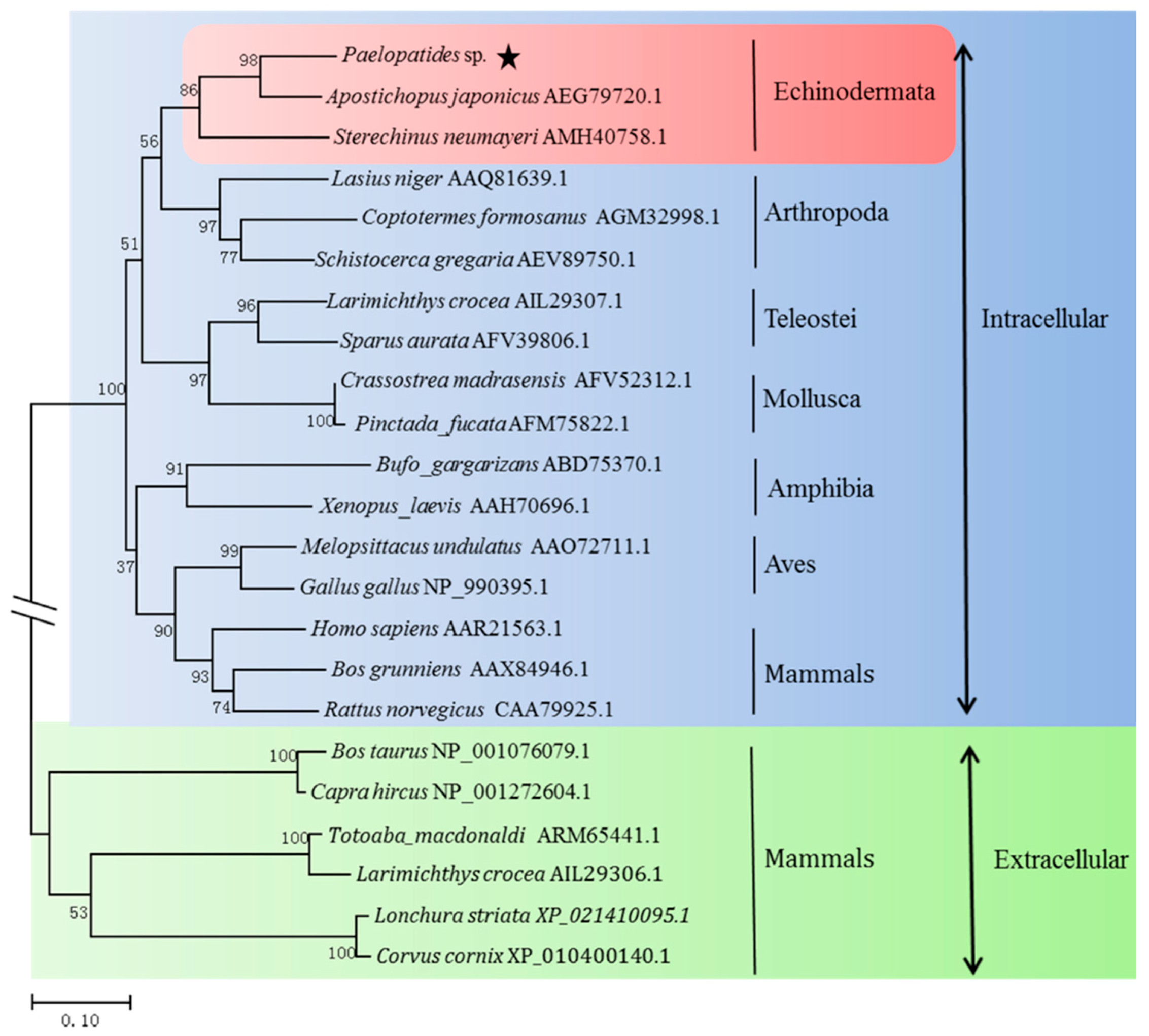
2.2. Homology and Phylogenetic Analysis
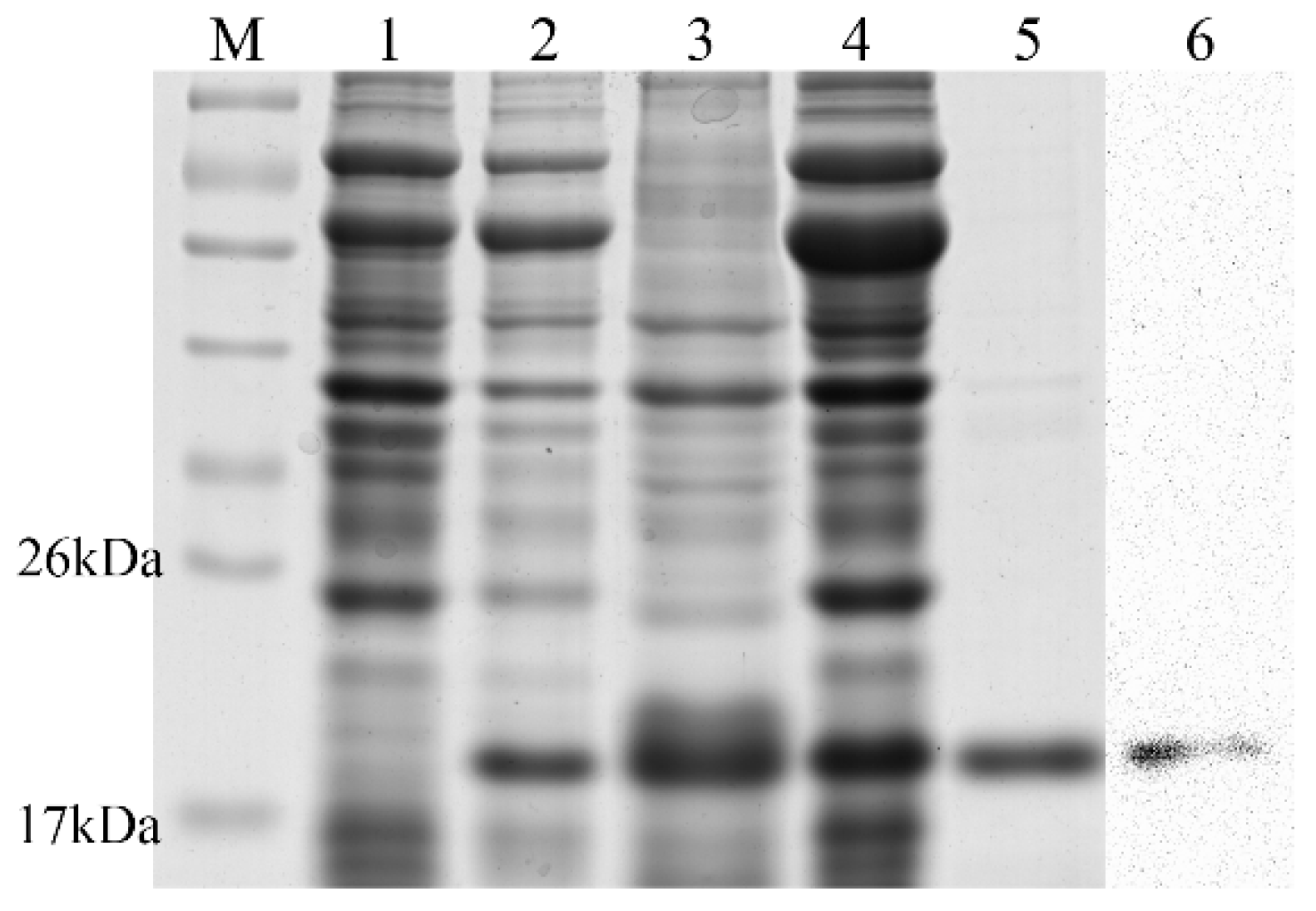
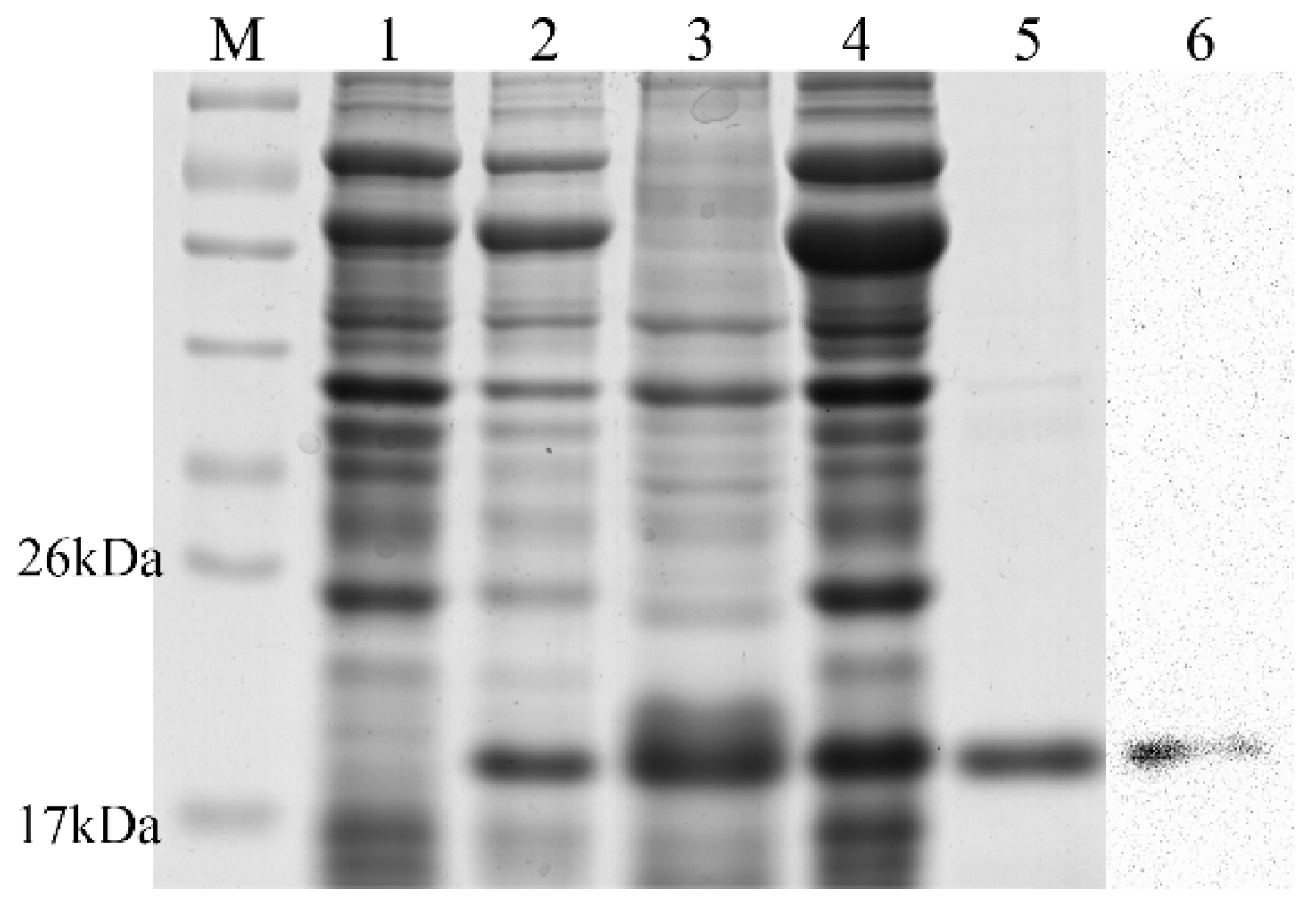
2.3. Expression, Purification, and Validation of the Recombinant Ps-Cu,Zn-SOD
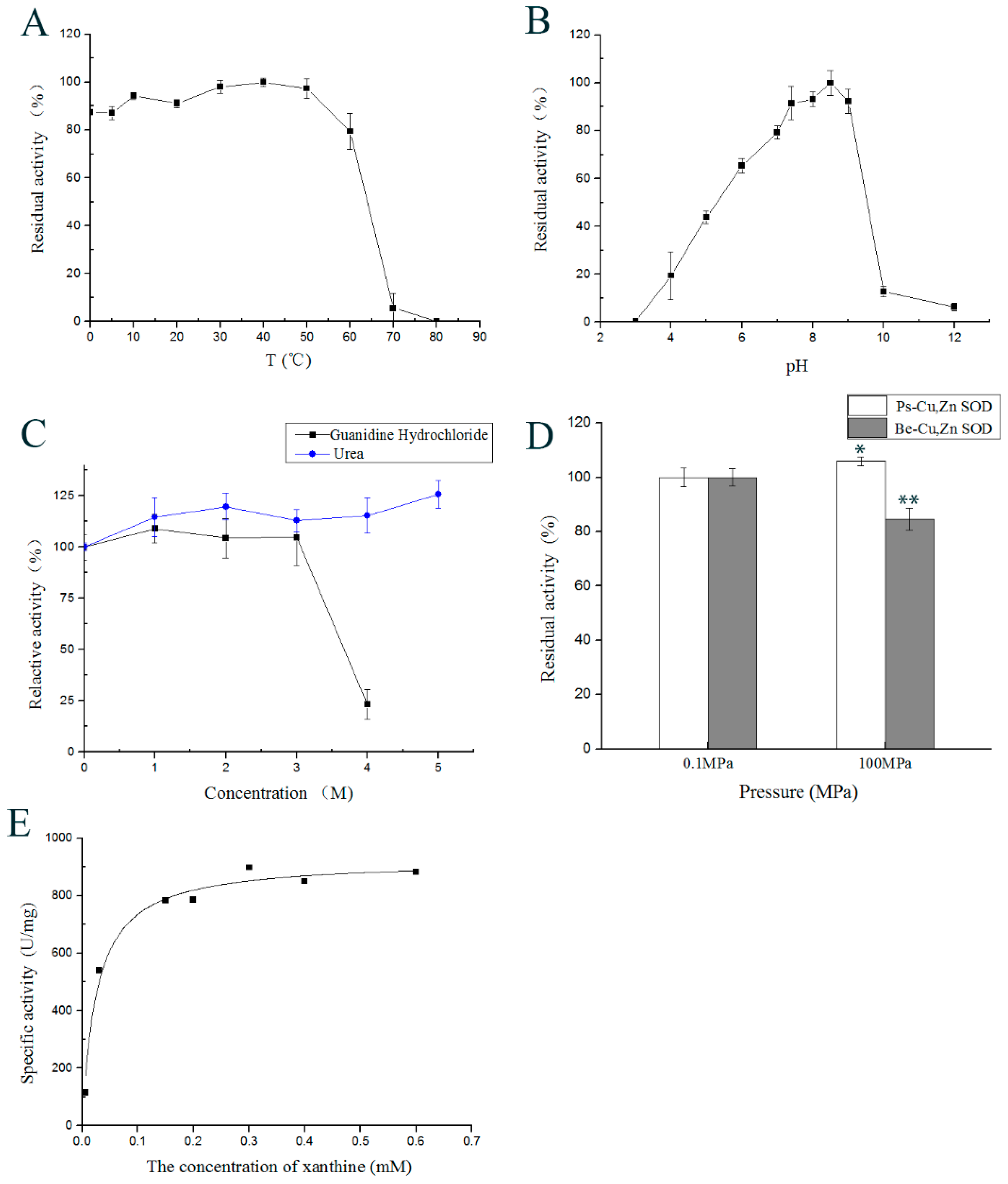
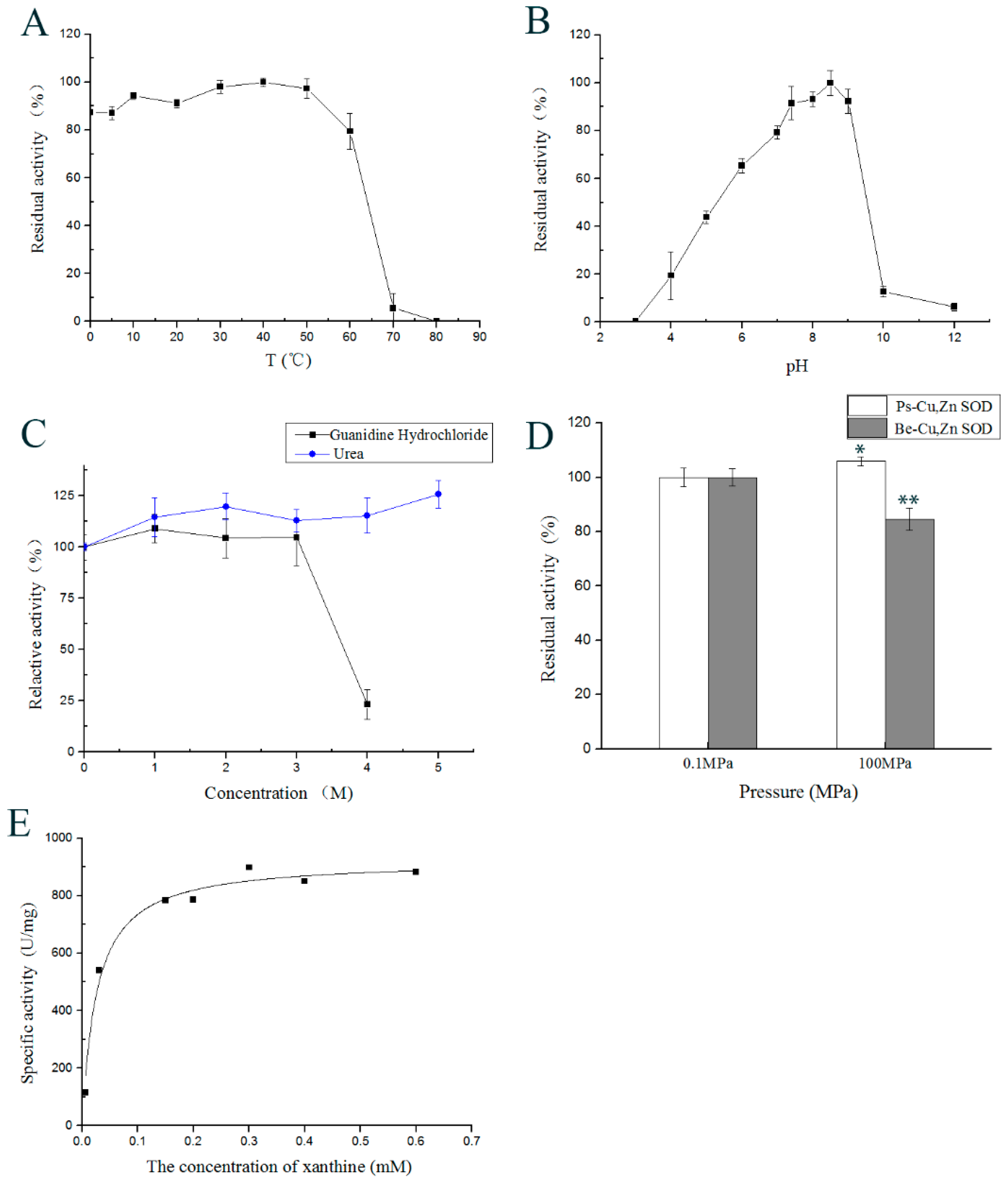
2.4. Effect of Temperature on Enzyme Activity
2.5. Effect of pH on the Enzyme Activity
2.6. Effect of Divalent Metal Ions on Enzyme Activity
2.7. Effects of Inhibitors, Reductants, Detergents, and Denaturants on Enzyme Activity
2.8. Resistance of Ps-Cu,Zn-SOD to Cleavage by Digestive Enzyme
2.9. Effect of High Hydrostatic Pressure on Ps-Cu,Zn-SOD Activity
2.10. Kinetic Parameters of Ps-Cu,Zn-SOD
3. Discussion
4. Materials and Methods
4.1. Material, RNA Extraction, and cDNA Cloning
4.2. Construction of Expression Vector
4.3. Expression and Purification of SOD Protein
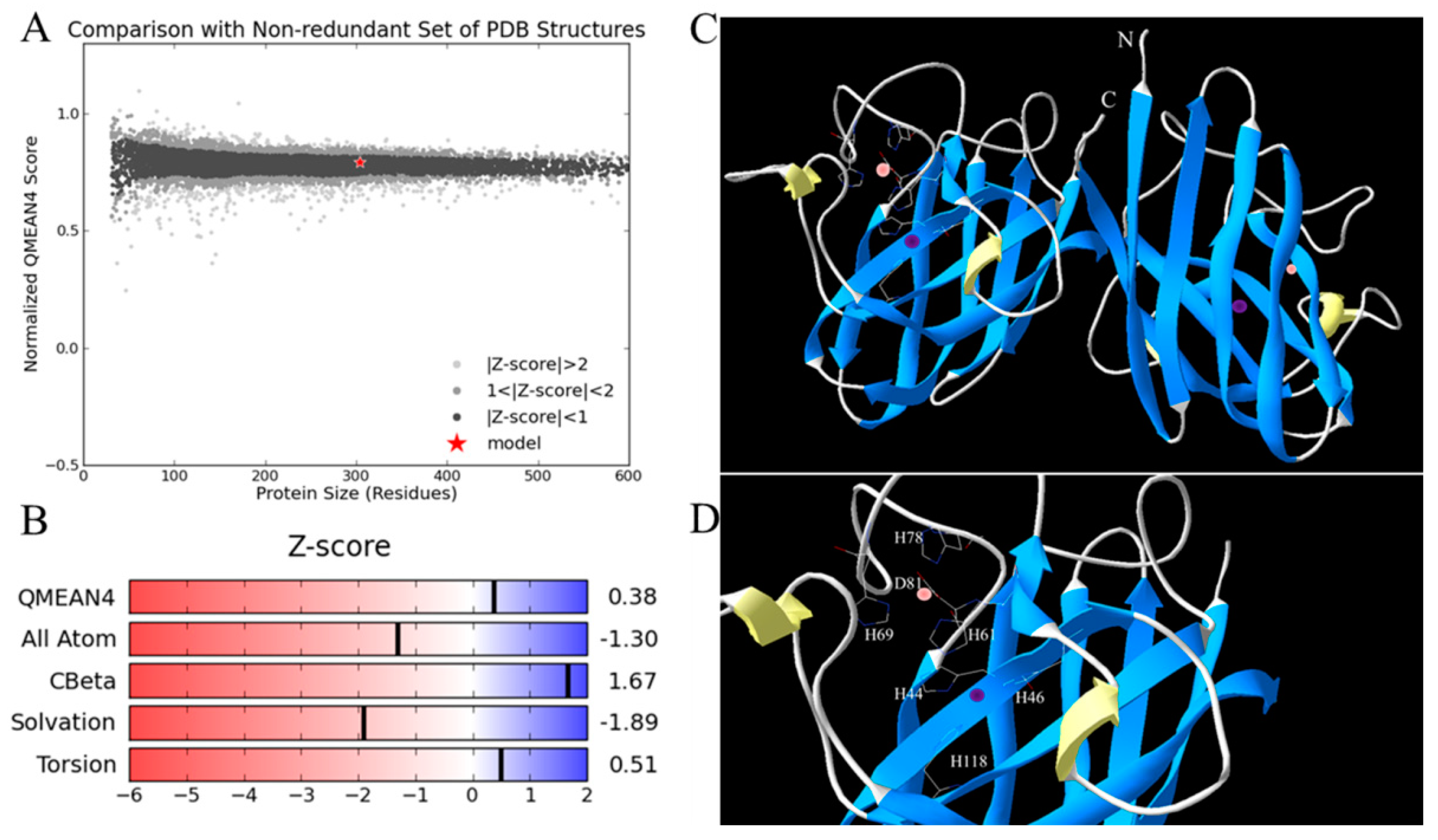
4.4. Sequence Analysis, Structure Modeling and Phylogenetic Analysis
4.5. Western Blot Analysis
4.6. Enzyme Characterization
4.6.1. SOD Assays and Protein Concentration Measurement
4.6.2. Thermostability of the Recombinant Ps-Cu,Zn-SOD
4.6.3. pH Stability of the Recombinant Ps-Cu,Zn-SOD
4.6.4. Effect of Ions on the Activity of the Recombinant Ps-Cu,Zn-SOD
4.6.5. Effects of Inhibitors, Reductants, Detergents, and Denaturants on the Activity of the Recombinant Ps-Cu,Zn-SOD
4.6.6. Proteolytic Susceptibility
4.6.7. High-Hydrostatic Pressure Stability
4.6.8. Determination of Kinetic Parameters
4.7. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Kawanishi, S.; Inoue, S. Damage to DNA by reactive oxygen and nitrogen species. Seikagaku J. Jpn. Biochem. Soc. 1997, 69, 1014–1017. [Google Scholar]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012. [Google Scholar] [CrossRef]
- Noor, R.; Mittal, S.; Iqbal, J. Superoxide dismutase—Applications and relevance to human diseases. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2002, 8, RA210–RA215. [Google Scholar]
- Auclair, J.R.; Brodkin, H.R.; D’Aquino, J.A.; Petsko, G.A.; Ringe, D.; Agar, J.N. Structural consequences of cysteinylation of Cu/Zn-superoxide dismutase. Biochemistry 2013, 52, 6145–6150. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Li, L.; Xu, F.; Zhang, G. Intracellular copper/zinc superoxide dismutase from bay scallop Argopecten irradians: Its gene structure, mRNA expression and recombinant protein. Fish Shellfish. Immunol. 2009, 27, 210–220. [Google Scholar] [CrossRef] [PubMed]
- And, V.P.; Siegbahn, P.E.M. Copper–zinc superoxide dismutase: Theoretical insights into the catalytic mechanism. Inorg. Chem. 2005, 44, 3311–3320. [Google Scholar]
- Bafana, A.; Dutt, S.; Kumar, S.; Ahuja, P.S. Superoxide dismutase: An industrial perspective. Crit. Rev. Biotechnol. 2011, 31, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Bafana, A.; Dutt, S.; Kumar, A.; Kumar, S.; Ahuja, P.S. The basic and applied aspects of superoxide dismutase. J. Mol. Catal. B Enzym. 2011, 68, 129–138. [Google Scholar] [CrossRef]
- Suzuki, Y.; Matsumoto, T.; Okamoto, S.; Hibi, T. A lecithinized superoxide dismutase (PC-SOD) improves ulcerative colitis. Colorectal Dis. 2008, 10, 931–934. [Google Scholar] [CrossRef] [PubMed]
- Sanclemente, G.; Garcia, J.; Zuleta, J.; Diehl, C.; Correa, C.; Falabella, R. A double-blind, randomized trial of 0.05% betamethasone vs. topical catalase/dismutase superoxide in vitiligo. J. Eur. Acad. Dermatol. Venereol. 2008, 22, 1359–1364. [Google Scholar] [CrossRef] [PubMed]
- Riedl, C.R.; Sternig, P.; Gallé, G.; Langmann, F.; Vcelar, B.; Vorauer, K.; Wagner, A.; Katinger, H.; Pflüger, H. Liposomal recombinant human superoxide dismutase for the treatment of peyronie’s disease: A randomized placebo-controlled double-blind prospective clinical study. Eur. Urol. 2005, 48, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Homaei, A.; Lavajoo, F.; Sariri, R. Development of marine biotechnology as a resource for novel proteases and their role in modern biotechnology. Int. J. Biol. Macromol. 2016, 88, 542–552. [Google Scholar] [CrossRef] [PubMed]
- Homaei, A.; Ghanbarzadeh, M.; Monsef, F. Biochemical features and kinetic properties of α-amylases from marine organisms. Int. J. Biol. Macromol. 2016, 83, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Sharifian, S.; Homaei, A.; Hemmati, R.; Khajeh, K. Light emission miracle in the sea and preeminent applications of bioluminescence in recent new biotechnology. J. Photochem. Photobiol. B Biol. 2017, 172, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hao, J.; Zhang, Y.Q.; Chen, X.L.; Xie, B.B.; Shi, M.; Zhou, B.C.; Zhang, Y.Z.; Li, P.Y. Identification and characterization of a novel salt-tolerant esterase from the deep-sea sediment of the south China sea. Front Microbiol 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Hung, V.S.; Hatada, Y.; Goda, S.; Jie, L.; Hidaka, Y.; Li, Z.; Akita, M.; Ohta, Y.; Watanabe, K.; Matsui, H. α-Glucosidase from a strain of deep-sea Geobacillus: A potential enzyme for the biosynthesis of complex carbohydrates. Appl. Microbiol. Biotechnol. 2013, 68, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Teh, A.H.; Kanamasa, S.; Kajiwara, S.; Kumasaka, T. Structure of Cu/Zn superoxide dismutase from the heavy-metal-tolerant yeast Cryptococcus liquefaciens strain N6. Biochem. Biophys. Res. Commun. 2008, 374, 475–478. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Jiang, Y.H.; Miao, J.L.; Wang, Q.F.; Zhang, B.T.; Li, G.Y. Purification and characterization of a cold-active iron superoxide dismutase from a psychrophilic bacterium, Marinomonas sp. NJ522. Biotechnol. Lett. 2006, 28, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yin, M.; Zhu, H.; Lu, J.; Cui, Z. Purification and characterization of a hyperthermostable Mn-superoxide dismutase from Thermus thermophilus HB27. Extremophiles 2011, 15, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.N.; He, Y.X.; Li, W.F.; Teng, Y.B.; Yu, J.; Chen, Y.; Zhou, C.Z. Crystal structures of holo and Cu-deficient Cu/Zn-SOD from the silkworm Bombyx mori and the implications in amyotrophic lateral sclerosis. Proteins Struct. Funct. Bioinform. 2010, 78, 1999–2004. [Google Scholar]
- And, M.M.; Colón, W. Structural basis of protein kinetic stability: Resistance to sodium dodecyl sulfate suggests a central role for rigidity and a bias toward β-sheet structure. Biochemistry 2004, 43, 11248–11254. [Google Scholar]
- Kumar, A.; Kaachra, A.; Bhardwaj, S.; Kumar, S. Copper, zinc superoxide dismutase of Curcuma aromatica is a kinetically stable protein. Process Biochem. 2014, 49, 1288–1296. [Google Scholar] [CrossRef]
- Zhu, Y.B.; Li, H.B.; Ni, H.; Liu, J.W.; Xiao, A.F.; Cai, H.N. Purification and biochemical characterization of manganesecontaining superoxide dismutase from deep-sea thermophile Geobacillus sp. EPT3. Acta Oceanol. Sin. 2014, 33, 163–169. [Google Scholar] [CrossRef]
- Schäfer, G.; Kardinahl, S. Iron superoxide dismutases: Structure and function of an archaic enzyme. Biochem. Soc. Trans. 2003, 31, 1330–1334. [Google Scholar] [CrossRef] [PubMed]
- Sujiwattanarat, P.; Pongsanarakul, P.; Temsiripong, Y.; Temsiripong, T.; Thawornkuno, C.; Uno, Y.; Unajak, S.; Matsuda, Y.; Choowongkomon, K.; Srikulnath, K. Molecular cloning and characterization of Siamese crocodile (Crocodylus siamensis) copper, zinc superoxide dismutase (CSI-Cu, Zn-SOD) gene. Comp. Biochem. Physiol. Part A 2016, 191, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-L.; Li, Y.-L.; Han, Z.; Chen, X.; Chen, Q.-J.; Wang, Y.; He, L.-S. Molecular Characterization of a Novel N-Acetylneuraminate Lyase from a Deep-Sea Symbiotic Mycoplasma. Mar. Drugs 2018, 16. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.B.; Wang, G.H.; Ni, H.; Xiao, A.F.; Cai, H.N. Cloning and characterization of a new manganese superoxide dismutase from deep-sea thermophile Geobacillus sp. EPT3. World J. Microbiol. Biotechnol. 2014, 30, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.S.; Didonato, M.; Barondeau, D.P.; Hura, G.L.; Hitomi, C.; Berglund, J.A.; Getzoff, E.D.; Cary, S.C.; Tainer, J.A. Superoxide dismutase from the eukaryotic thermophile alvinella pompejana: Structures, stability, mechanism, and insights into amyotrophic lateral sclerosis. J. Mol. Biol. 2009, 385, 1534–1555. [Google Scholar] [CrossRef] [PubMed]
- Janssens, B.J.; Childress, J.J.; Baguet, F.; Rees, J.F. Reduced enzymatic antioxidative defense in deep-sea fish. J. Exp. Biol. 2000, 203, 3717–3725. [Google Scholar] [PubMed]
- Chilukuri, L.N.; Bartlett, D.H. Isolation and characterization of the gene encoding single-stranded-DNA-binding protein (SSB) from four marine Shewanella strains that differ in their temperature and pressure optima for growth. Microbiology 1997, 143, 1163–1174. [Google Scholar] [CrossRef] [PubMed]
- Kawano, H.; Nakasone, K.; Matsumoto, M.; Yoshida, Y.; Usami, R.; Kato, C.; Abe, F. Differential pressure resistance in the activity of RNA polymerase isolated from Shewanella violacea and Escherichia coli. Extremophiles 2004, 8, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Benkert, P.; Biasini, M.; Schwede, T. Toward the estimation of the absolute quality of individual protein structure models. Bioinformatics 2011, 27, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Oyanagui, Y. Reevaluation of assay methods and establishment of kit for superoxide dismutase activity. Anal. Biochem. 1984, 142, 290–296. [Google Scholar] [CrossRef]
- Zheng, L.; Wu, B.; Liu, Z.; Tian, J.; Yu, T.; Zhou, L.; Sun, X.; Yang, A.G. A manganese superoxide dismutase (MnSOD) from ark shell, Scapharca broughtonii: Molecular characterization, expression and immune activity analysis. Fish Shellfish. Immunol. 2015, 45, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Ken, C.-F.; Hsiung, T.-M.; Huang, Z.-X.; Juang, R.-H.; Lin, C.T. Characterization of Fe/Mn−superoxide dismutase from diatom Thallassiosira weissflogii: Cloning, expression, and property. J. Agric. Food Chem. 2005, 53, 1470–1474. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Genbank No. | Abbreviation | Size | Similarity % | Identity % | Gap % |
|---|---|---|---|---|---|---|
| Apostichopus japonicus | AEG79720.1 | A. ja | 149 aa | 88.8 | 82.9 | 2.0 |
| Sterechinus neumayeri | AMH40758.1 | S. ne | 151 aa | 83.6 | 73.7 | 0.7 |
| Ixodes scapularis | AAY66847.1 | I. sc | 154 aa | 79.4 | 69.7 | 2.6 |
| Coptotermes formosanus | AGM32998.1 | C. fo | 154 aa | 80.5 | 68.8 | 1.3 |
| Orussus abietinus | XP_012280251.1 | O. ab | 155 aa | 77.4 | 68.4 | 1.9 |
| Schistocerca gregaria | AEV89750.1 | S. gr | 154 aa | 78.6 | 67.5 | 1.3 |
| Crassostrea madrasensis | AFV52312.1 | C. ma | 156 aa | 78.3 | 67.5 | 3.8 |
| Neoseiulus barkeri | AON96404.1 | N. ba | 153 aa | 77.1 | 67.3 | 0.7 |
| Divalent Metal Ions | Concentration/mmol·L−1 | Relative Activity/% |
|---|---|---|
| Control | - | 100 ± 3.94 |
| Mn2+ | 0.1 | 96.86 ± 1.25 ** |
| 1 | 13.59 ± 2.04 ** | |
| Co2+ | 0.1 | 93.54 ± 3.08 ** |
| 1 | 42.33 ± 2.50 ** | |
| Ni2+ | 0.1 | 90.64 ± 4.28 ** |
| 1 | 65.92 ± 3.78 ** | |
| Cu2+ | 0.1 | 103.13 ± 1.21 |
| 1 | 13.97 ± 4.68 ** | |
| Ba2+ | 0.1 | 101.42 ± 1.59 |
| 1 | 98.16 ± 2.76 * | |
| Mg2+ | 0.1 | 100.21 ± 4.78 |
| 1 | 99.42 ± 4.12 | |
| Ca2+ | 0.1 | 100.91 ± 3.29 |
| 1 | 101.06 ± 2.08 | |
| Zn2+ | 0.1 | 114.07 ± 4.94 ** |
| 1 | 111.87 ± 4.42 ** |
| Inhibitors, Detergents, and Denaturants | Concentration | Relative Activity/% |
|---|---|---|
| Control | - | 100 ± 3.71 |
| EDTA | 1 mmol·L−1 | 80.86 ± 4.60 ** |
| 10 mmol·L−1 | 71.86 ± 4.86 ** | |
| DTT | 1 mmol·L−1 | 98.93 ± 0.81 * |
| 10 mmol·L−1 | 56.62 ± 4.27 ** | |
| β-ME | 1 mmol·L−1 | 122.70 ± 1.40 ** |
| 10 mmol·L−1 | 118.78 ± 2.99 ** | |
| Triton X-100 | 0.1% | 106.66 ± 1.63 ** |
| 1% | 100.91 ± 1.70 | |
| SDS | 0.1% | 63.59 ± 4.30 ** |
| 1% | 37.01 ± 3.02 ** | |
| Chaps | 0.1% | 110.37 ± 3.13 ** |
| 1% | 108.32 ± 3.99 ** | |
| Tween 20 | 0.1% | 107.19 ± 1.40 ** |
| 1% | 98.20 ± 1.58 * |
| Name of Enzyme | No. of Cleavages | Positions of Cleavage Sites |
|---|---|---|
| Chymotrypsin-high specificity (C-term to [FYW], not before P) | 4 | 20, 43, 48, and 62 |
| Trypsin | 12 | 13, 38, 66, 67, 75, 77, 96, 113, 126, 141, 150, and 151 |
| Time (h) | Relative Activity (%) |
|---|---|
| 0 | 100 ± 4.30 |
| 1 | 101.09 ± 8.58 |
| 2 | 96.89 ± 3.98 |
| 3 | 93.36 ± 5.62 ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Kong, X.; Chen, J.; Liu, H.; Zhang, H. Characteristics of the Copper,Zinc Superoxide Dismutase of a Hadal Sea Cucumber (Paelopatides sp.) from the Mariana Trench. Mar. Drugs 2018, 16, 169. https://doi.org/10.3390/md16050169
Li Y, Kong X, Chen J, Liu H, Zhang H. Characteristics of the Copper,Zinc Superoxide Dismutase of a Hadal Sea Cucumber (Paelopatides sp.) from the Mariana Trench. Marine Drugs. 2018; 16(5):169. https://doi.org/10.3390/md16050169
Chicago/Turabian StyleLi, Yanan, Xue Kong, Jiawei Chen, Helu Liu, and Haibin Zhang. 2018. "Characteristics of the Copper,Zinc Superoxide Dismutase of a Hadal Sea Cucumber (Paelopatides sp.) from the Mariana Trench" Marine Drugs 16, no. 5: 169. https://doi.org/10.3390/md16050169
APA StyleLi, Y., Kong, X., Chen, J., Liu, H., & Zhang, H. (2018). Characteristics of the Copper,Zinc Superoxide Dismutase of a Hadal Sea Cucumber (Paelopatides sp.) from the Mariana Trench. Marine Drugs, 16(5), 169. https://doi.org/10.3390/md16050169