Cytotoxic Tricycloalternarene Compounds from Endophyte Alternaria sp. W-1 Associated with Laminaria japonica


Abstract
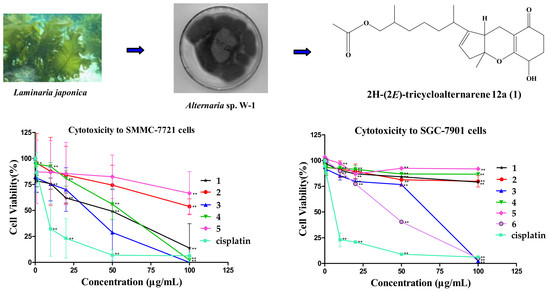
1. Introduction
2. Results and Discussion
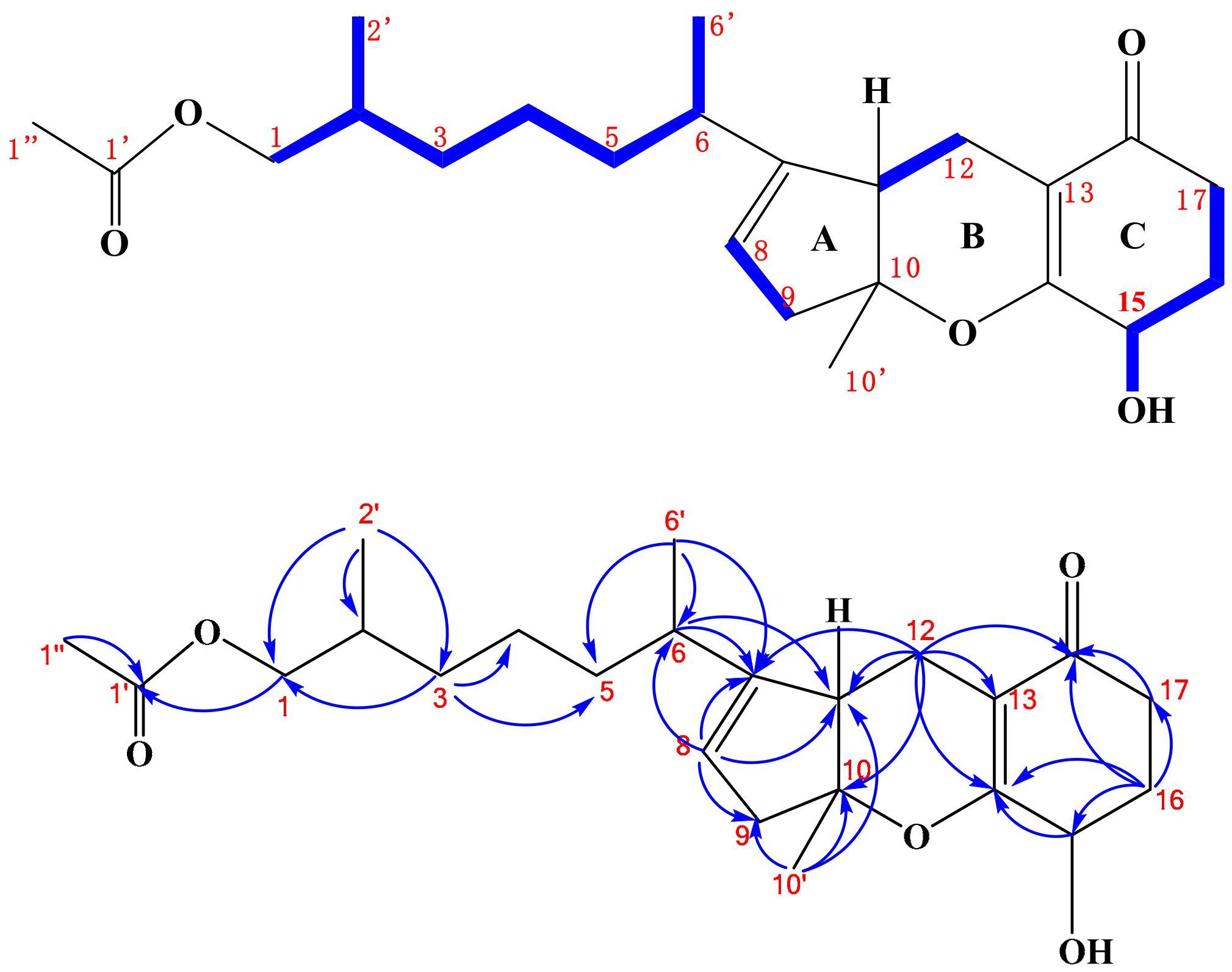
2.1. Structure Elucidation
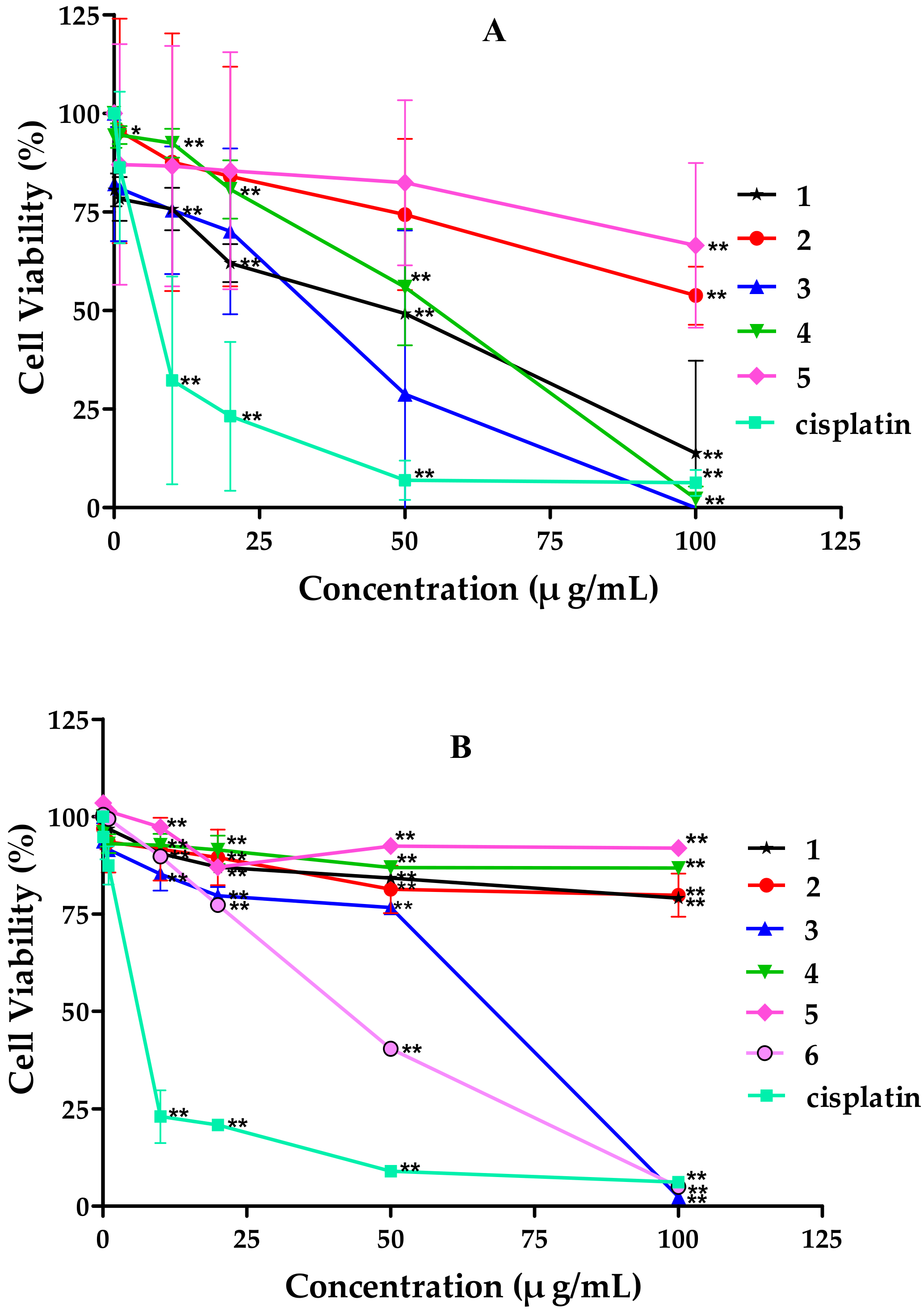
2.2. Cytotoxicity of Tricycloalternarene Compounds
2.3. Tricycloalternarene 3a (3) Induced G1 Phase Cell Cycle Arrest in SMMC-7721 Cells
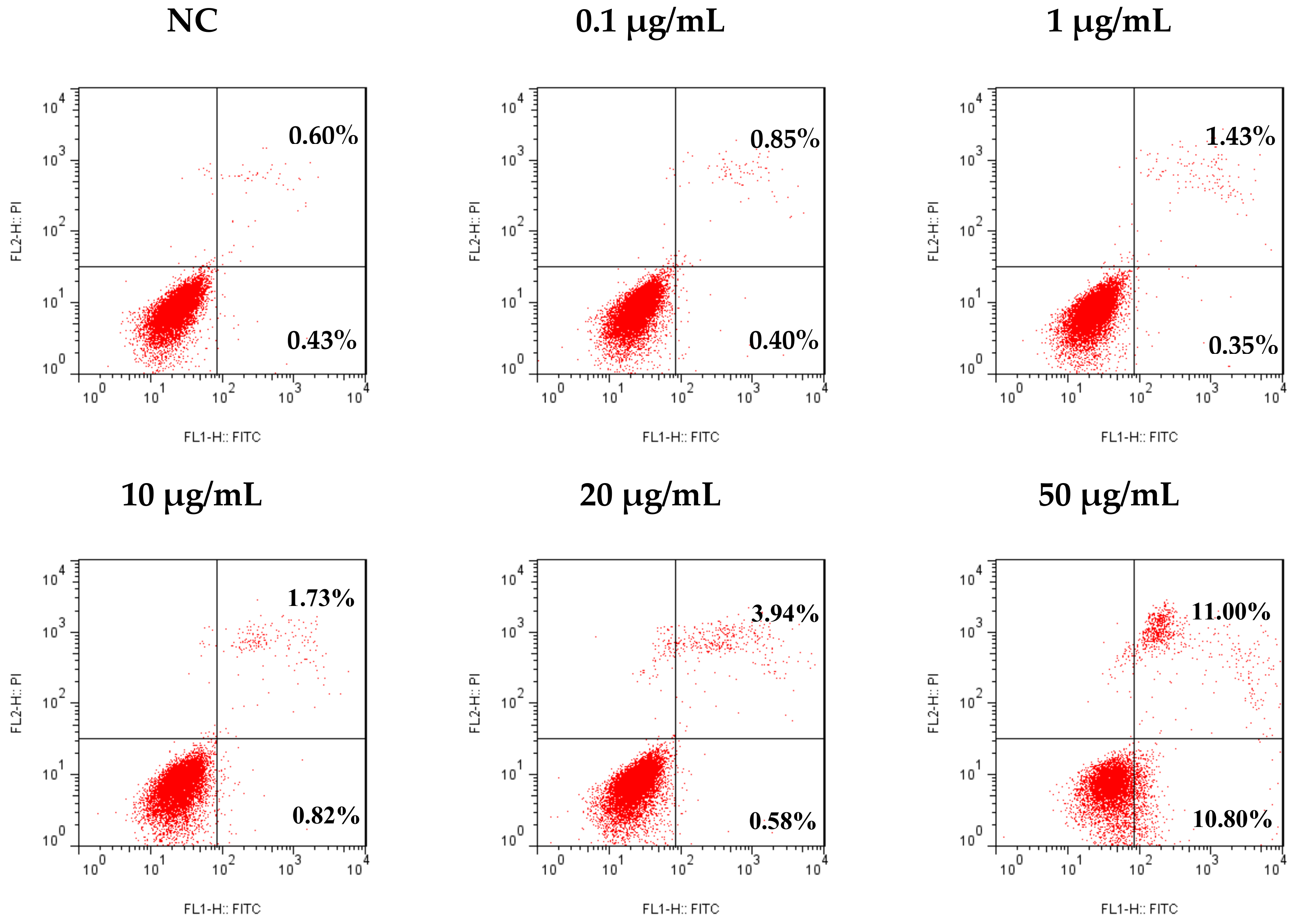
2.4. Tricycloalternarene 3a (3) Induced Apoptosis in SMMC-7721 Cells
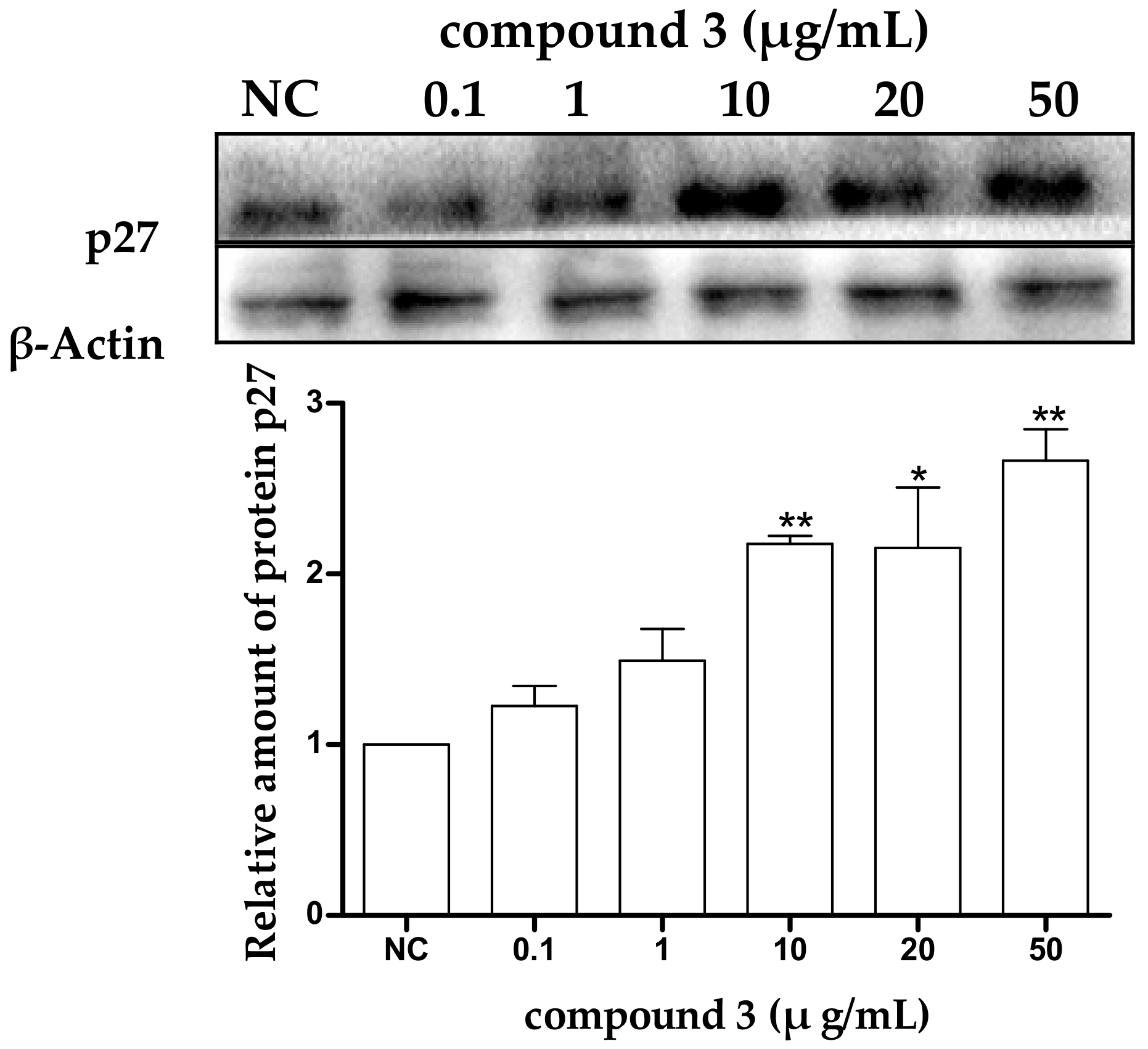
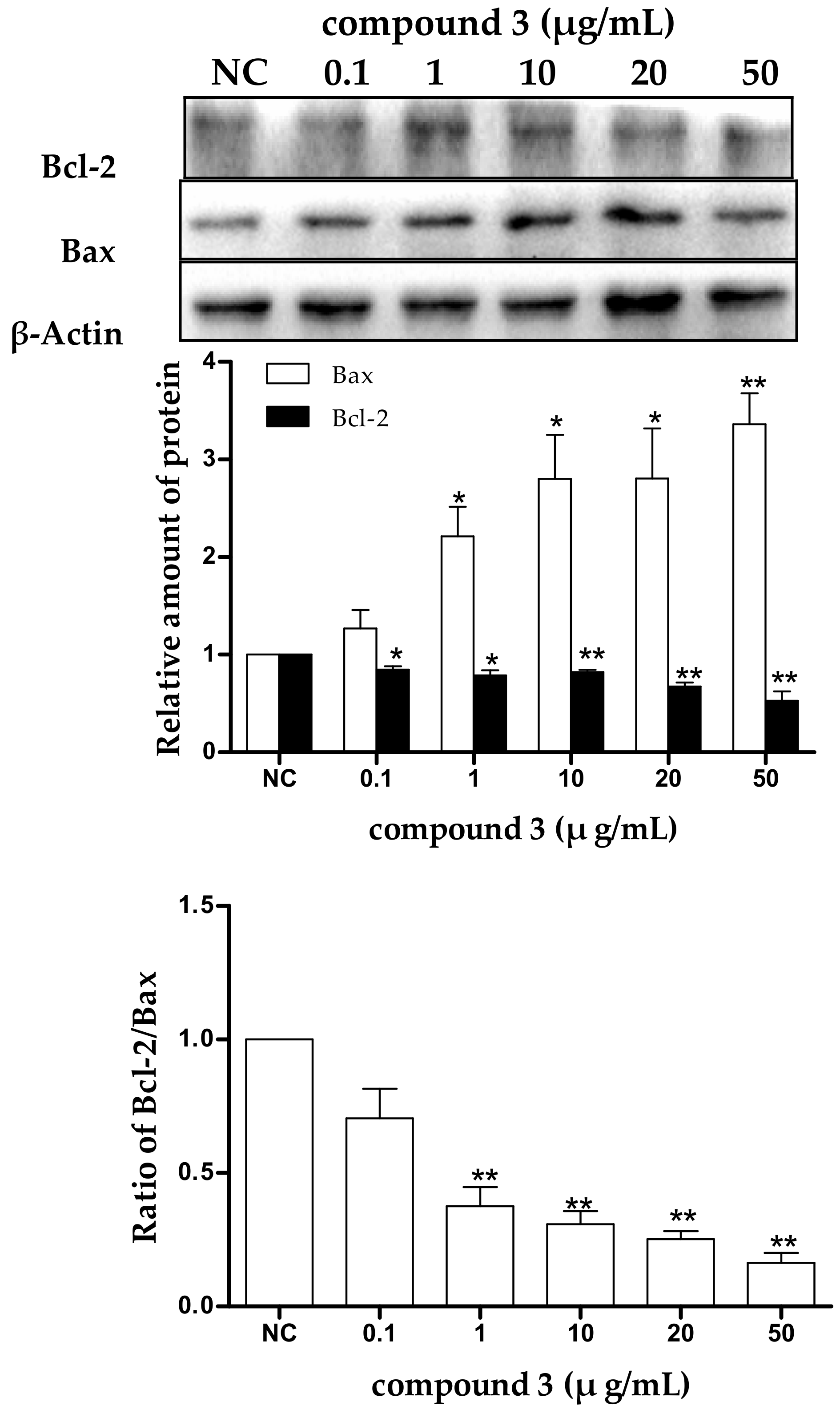
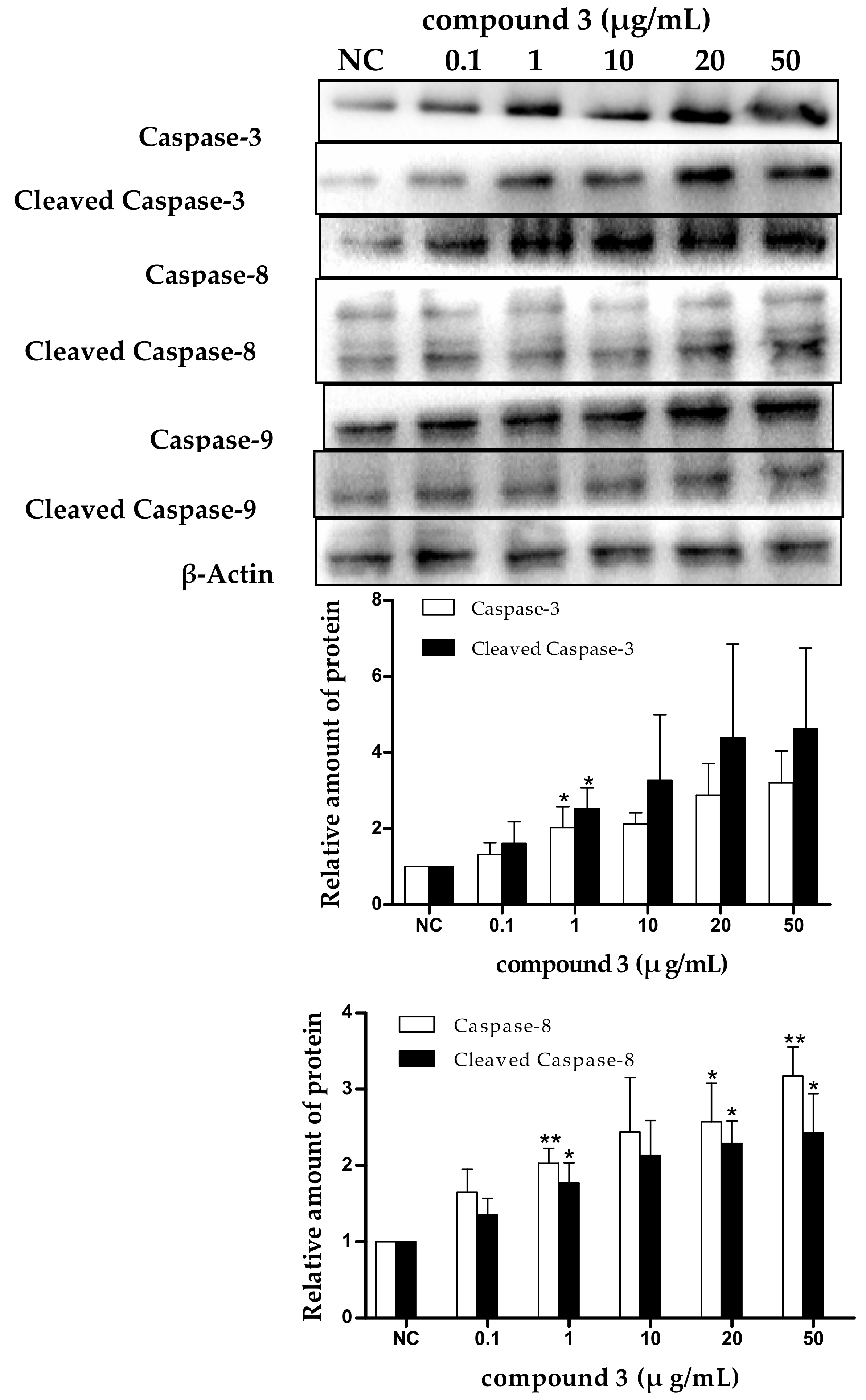
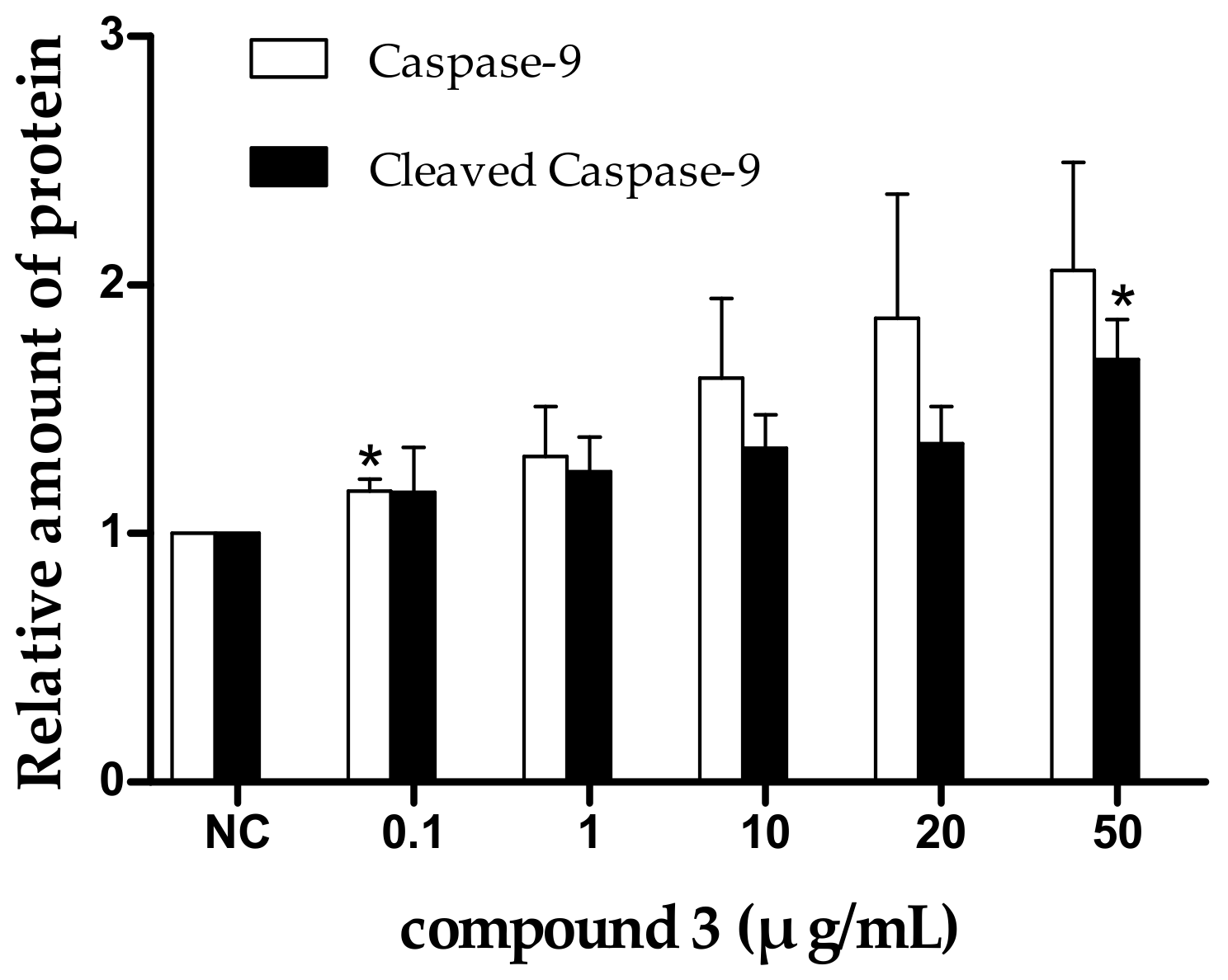
2.5. Tricycloalternarene 3a-Induced Apoptosis of SMMC-7721 Cells Involved in Both the Death Receptor Pathway and the Mitochondrial Pathway
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Strain
3.3. Extraction and Isolation
3.4. Cytotoxicity Assay
3.5. Cell Cycle Analysis
3.6. Detection of Apoptosis
3.7. Western Blot Analysis
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Miao, L.; Zheng, Z.H.; Su, W.J. Research development of bioactive substance of symbiotic and epibiotic marine microorganisms. Mar. Sci. Bull. 2002, 21, 62–68. [Google Scholar]
- John, W.; Brent, R.; Robert, A.; Murray, H.G.; Michele, R. Marine natural products. Nat. Prod. Rep. 2014, 31, 160–258. [Google Scholar]
- Zha, C.Y.; Zhu, T.H.; Zhu, W.M. New marine natural products of microbial origin from 2010 to 2013. Chin. J. Org. Chem. 2013, 33, 1195–1234. [Google Scholar] [CrossRef]
- Lou, J.F.; Fu, L.Y.; Peng, Y.L.; Zhou, L.G. Metabolites from Alternaria fungi and their bioactivities. Molecules 2013, 18, 5891–5935. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.Z.; Yin, X.L.; Fang, S.T.; Miao, F.P.; Ji, N.Y. Two new isomeric tricycloalternarenes from the marine alga-epiphytic fungus Alternaria alternata k23-3. Magn. Reson. Chem. 2018, 56, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Liebermann, B.; Ellinger, R.; Gunther, W.; Ihn, W.; Gallander, H. Tricycloalternarenes produced by Alternaria alternata related to ACTG-toxins. Phytochemistry 1997, 46, 297–303. [Google Scholar] [CrossRef]
- Wang, Q.X.; Bao, L.; Yang, X.L.; Guo, H.; Ren, B.; Guo, L.D.; Song, F.H.; Wang, W.Z.; Liu, H.W.; Zhang, L.X. Tricycloalternarenes F-H: Three new mixed terpenoids produced by an endolichenic fungus Ulocladium sp. using OSMAC method. Fitoterapia 2013, 85, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Liang, F.L.; Li, D.L.; Chen, Y.C.; Tao, M.H.; Zhang, W.M.; Zhang, D.Z. Secondary metabolites of endophytic Guignardia mangiferae from Smilax glabra and their antitumor activities. Zhongcaoyao 2012, 43, 856–860. [Google Scholar]
- Kono, Y.; Gardner, J.M.; Suzuki, Y.; Takeuchi, S. Studies on host-selective toxins produced by a pathotype of Alternaria citri causing brown spot disease of Mandarins. Agric. Biol. Chem. 1986, 50, 1597–1606. [Google Scholar] [CrossRef]
- Shi, Z.Z.; Miao, F.P.; Fang, S.T.; Liu, X.H.; Yin, X.L.; Ji, N.Y. Sesteralterin and tricycloalterfurenes A-D: Terpenes with rarely occurring frameworks from the marine-alga-epiphytic fungus Alternaria alternata k21-1. J. Nat. Prod. 2017, 80, 2524–2529. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Zhao, P.J.; Ma, J.; Li, G.H.; Shen, Y.M. Tricycloalternarenes A-E: Five new mixed terpenoids from the endophytic fungal strain Alternaria alternate Ly83. Helv. Chim. Acta 2008, 91, 1588–1594. [Google Scholar] [CrossRef]
- Guimaraes, D.O.; Lopes, N.P.; Pupo, M.T. Meroterpenes isolated from the endophytic fungus Guignardia mangiferae. Phytochem. Lett. 2012, 5, 519–523. [Google Scholar] [CrossRef]
- Yoiprommarat, S.; Srichomthong, K.; Deelai, S.; Suetrong, S.; Sakayaroj, J.; Bunyapaiboonsri, T.; Unagul, P. Secondary metabolites of the marine fungus Paradendryphiella arenariae BCC 17999. Bot. Mar. 2015, 58, 393–399. [Google Scholar] [CrossRef]
- Pegg, K.G. Studies of a strain of Alternaria citri Pierce, the causal organism of brown spot of Emperor mandarin. Qld. J. Agric. Anim. Sci. 1966, 23, 15–28. [Google Scholar]
- Whiteside, J.O. A newly recorded Alternaria-induced brown spot disease on Dancy tangerines in Florida. Plant Dis. Rep. 1976, 60, 326–329. [Google Scholar]
- Zhang, G.J.; Wu, G.G.; Zhu, T.J.; Kurtan, T.; Mandi, A.; Jiao, J.Y.; Li, J.; Qi, X.; Gu, Q.Q.; Li, D.H. Meroterpenoids with diverse ring systems from the sponge-associated fungus Alternaria sp. JJY-32. J. Nat. Prod. 2013, 76, 1946–1957. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.Y.; Zhang, G.J.; Tang, X.L.; Jiao, J.Y.; Kim, S.Y.; Lee, J.Y.; Zhu, T.J.; Li, D.H.; Yun, Y.G.; Gu, Q.Q. Toll-like receptor 4/nuclear factor-κB signaling pathway is involved in ACTG-toxin H-mediated anti-inflammatory effect. Mol. Cell. Biochem. 2013, 374, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Huang, W.Z.; Zhao, P.J. Mixed terpenoids from an endophyte Alternaria sp. Ly81 cultivated on Maytenus hookeri. Chin. Pharm. J. 2013, 48, 1823–1826. [Google Scholar]
- Sun, H.; Gao, S.S.; Li, X.M.; Li, C.S.; Wang, B.G. Chemical constituents of marine mangrove-derived endophytic fungus Alternaria tenuissima EN-192. Chin. J. Oceanol. Limnol. 2013, 31, 464–470. [Google Scholar] [CrossRef]
- Shi, Z.Z.; Fang, S.T.; Miao, F.P.; Ji, N.Y. Two new tricycloalternarene esters from an alga-epiphytic isolate of Alternaria alternate. Nat. Prod. Res. 2018, 26, 1–6. [Google Scholar]
- Wang, H.F.; Bai, J.; Chen, G.; Liu, Z.Q.; Lu, X.; Pei, Y.H. 1H and 13C NMR assignments of three new tricycloaternarenes from endophytic fungus Alternaria tenuissima HS03-07. Magn. Reson. Chem. 2015, 53, 227–232. [Google Scholar] [CrossRef] [PubMed]
- You, C.C.; Wang, Y.L.; Huang, L.M. Research progress of the function of p27 and its related factors in malignant tumor. Chem. Life 2010, 30, 914–918. [Google Scholar]
- Nourse, J.; Firpo, E.; Flanagan, W.M.; Coats, S.; Polyak, K.; Lee, M.H.; Massaque, J.; Crabtree, G.R.; Roberts, J.M. Interleukin-2-mediated elimination of the p27 Kip1 cyclin-dependent kinase inhibitor prevented by rapamycin. Nature 1994, 372, 570–573. [Google Scholar] [CrossRef] [PubMed]
- Peng, D.; Fan, Z.; Lu, Y.; DeBlasio, T.; Scher, H.; Mendelsohn, J. Anti-epidermal growth factor receptor monooclonal antibody 225 up-regulates p27 KIP1 and induces G1 arrest in prostatic cancer cell line DU145. Cancer Res. 1996, 56, 3666–3669. [Google Scholar] [PubMed]
- Slingerland, J.; Pagano, M. Regulation of the cdk inhibitor p27 and its deregulation in cancer. J. Cell. Physiol. 2000, 183, 10–17. [Google Scholar] [CrossRef]
- Evan, G.; Littlewood, T. A matter of life and cell death. Science 1998, 281, 1317–1322. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.T. Programmed cell death: Apoptosis and oncogenesis. Cell 1991, 65, 1097–1098. [Google Scholar] [CrossRef]
- Ouyang, L.; Shi, Z.; Zhao, S.; Wang, F.T.; Zhou, T.T.; Liu, B.; Bao, J.K. Programmed cell death pathways in cancer: A review of apoptosis, autophagy and programmed necrosis. Cell Prolif. 2012, 45, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Cohen-Saidon, C.; Carmi, I.; Keren, A.; Razin, E. Antiapoptotic function of Bcl-2 in mast cells is dependent on its association with heat shock protein 90β. Blood 2006, 107, 1413–1420. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.S.; Robertson, A.A.B.; Cooper, M.A. Natural product and natural product derived drugs. Nat. Prod. Rep. 2014, 31, 1612–1661. [Google Scholar] [CrossRef] [PubMed]
- Baggstrom, M.Q.; Qi, Y.; Koczywas, M.; Argiris, A.; Johnson, E.A.; Millward, M.J.; Murphy, S.C.; Erlichman, C.; Rudin, C.M.; Govindan, R. A phase II study of AT-101 (gossypol) in chemotherapy-sensitive recurrent extensive stage small cell lung cancer (ES-SCLC). J. Thorac. Oncol. 2011, 6, 1757–1760. [Google Scholar] [CrossRef] [PubMed]
- Gross, A.; McDonnell, J.M.; Korsmeyer, S.J. BCL-2 family members and the mitochondria in apoptosis. Genes Dev. 1999, 13, 1899–1911. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Dassonneville, L.; Lansiaux, A.; Wattelet, A.; Wattez, N.; Mahieu, C.; Miert, S.V.; Pieters, L.; Bailly, C. Cytotoxicity and cell cycle effects of the plant alkaloids cryptolepine and neocryptolepine: Relation to drug-induced apoptosis. Eur. J. Pharmacol. 2000, 409, 9–18. [Google Scholar] [CrossRef]
- Degterev, A.; Boyce, M.; Yuan, J. A decade of caspases. Oncogene 2003, 22, 8543–8567. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Slingerland, J.M. Multiple roles of the PI3K/PKB (Akt) pathway in cell cycle progression. Cell Cycle 2003, 2, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Hausler, P.; Papoff, G.; Eramo, A.; Reif, K.; Cantrell, D.A.; Ruberti, G. Protection of CD95-mediated apoptosis by activation of phosphatidylinositide 3-kinase and protein kinase B. Eur. J. Immunol. 1998, 28, 57–69. [Google Scholar] [CrossRef]
- Datta, S.R.; Dudek, H.; Tao, X.; Masters, S.; Fu, H.; Gotoh, Y.; Greenberg, M.E. Akt phosphorylation of BAD couples survival signals to the cell-intrinsic death machinery. Cell 1997, 91, 231–241. [Google Scholar] [CrossRef]
- Song, G.; Ouyang, G.; Bao, S. The activation of Akt/PKB signaling pathway and cell survival. J. Cell. Mol. Med. 2005, 9, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.Y.; Zubovitz, J.; Petrocelli, T.; Kotchetkov, R.; Connor, M.K.; Han, K.; Lee, J.H.; Ciarallo, S.; Catzavelos, C.; Beniston, R.; et al. PKB/Akt phosphorylates p27, impairs nuclear import of p27 and opposes p27-mediated G1 arrest. Nat. Med. 2002, 8, 1153–1160. [Google Scholar] [CrossRef] [PubMed]

 ) and HMBC (
) and HMBC ( ) correlations of compound 1.
) correlations of compound 1.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | δC | δH, Mult. (J in Hz) |
|---|---|---|
| 1 | 68.4 | 3.83, dd (10.8, 6.0); 3.75, dd (10.8, 6.6) |
| 2 | 31.7 | 1.64, m |
| 2′ | 16.5 | 0.81, d (7.2) |
| 3 | 32.6 | 1.23, m; 1.00, m |
| 4 | 23.9 | 1.15 *; 1.15 * |
| 5 | 34.4 | 1.36, m; 1.15 * |
| 6 | 31.9 | 1.89 * |
| 6′ | 20.1 | 0.90, d (6.6) |
| 7 | 149.5 | - |
| 8 | 119.7 | 5.31, s |
| 9 | 44.3 | 2.52, m; 2.43 * |
| 10 | 87.3 | - |
| 10′ | 23.0 | 1.40, s |
| 11 | 46.0 | 2.76, br d (4.8) |
| 12 | 14.8 | 2.43 *; 2.09, m |
| 13 | 106.7 | - |
| 14 | 170.8 | - |
| 15 | 64.6 | 4.01, t (4.8) |
| 16 | 29.6 | 1.91 *; 1.80, m |
| 17 | 32.2 | 2.47, m; 2.08, m |
| 18 | 195.7 | - |
| 1′ | 170.3 | - |
| 1″ | 20.6 | 2.00, s |
| 15-OH | - | 5.38, s |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, L.; Tian, S.-J.; Song, H.-L.; Chen, X.; Guo, H.; Wan, D.; Wang, Y.-R.; Wang, F.-W.; Liu, L.-J. Cytotoxic Tricycloalternarene Compounds from Endophyte Alternaria sp. W-1 Associated with Laminaria japonica. Mar. Drugs 2018, 16, 402. https://doi.org/10.3390/md16110402
Shen L, Tian S-J, Song H-L, Chen X, Guo H, Wan D, Wang Y-R, Wang F-W, Liu L-J. Cytotoxic Tricycloalternarene Compounds from Endophyte Alternaria sp. W-1 Associated with Laminaria japonica. Marine Drugs. 2018; 16(11):402. https://doi.org/10.3390/md16110402
Chicago/Turabian StyleShen, Li, Shu-Juan Tian, Hui-Liang Song, Xi Chen, Hao Guo, Dan Wan, Yu-Rou Wang, Feng-Wu Wang, and Li-Jun Liu. 2018. "Cytotoxic Tricycloalternarene Compounds from Endophyte Alternaria sp. W-1 Associated with Laminaria japonica" Marine Drugs 16, no. 11: 402. https://doi.org/10.3390/md16110402
APA StyleShen, L., Tian, S.-J., Song, H.-L., Chen, X., Guo, H., Wan, D., Wang, Y.-R., Wang, F.-W., & Liu, L.-J. (2018). Cytotoxic Tricycloalternarene Compounds from Endophyte Alternaria sp. W-1 Associated with Laminaria japonica. Marine Drugs, 16(11), 402. https://doi.org/10.3390/md16110402