
Peptides, Peptidomimetics, and Polypeptides from Marine Sources: A Wealth of Natural Sources for Pharmaceutical Applications


Abstract
:1. Introduction
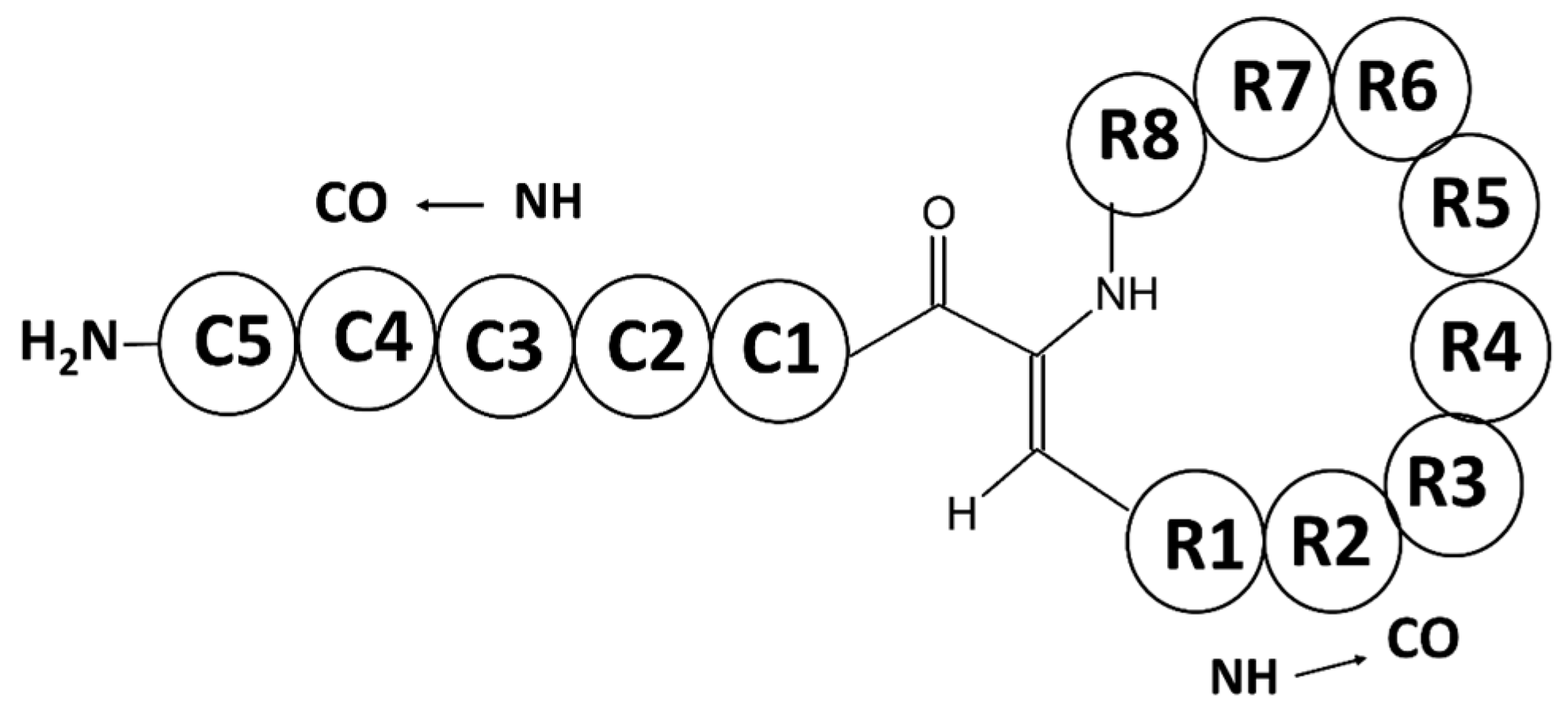
Peptides from Natural Sources
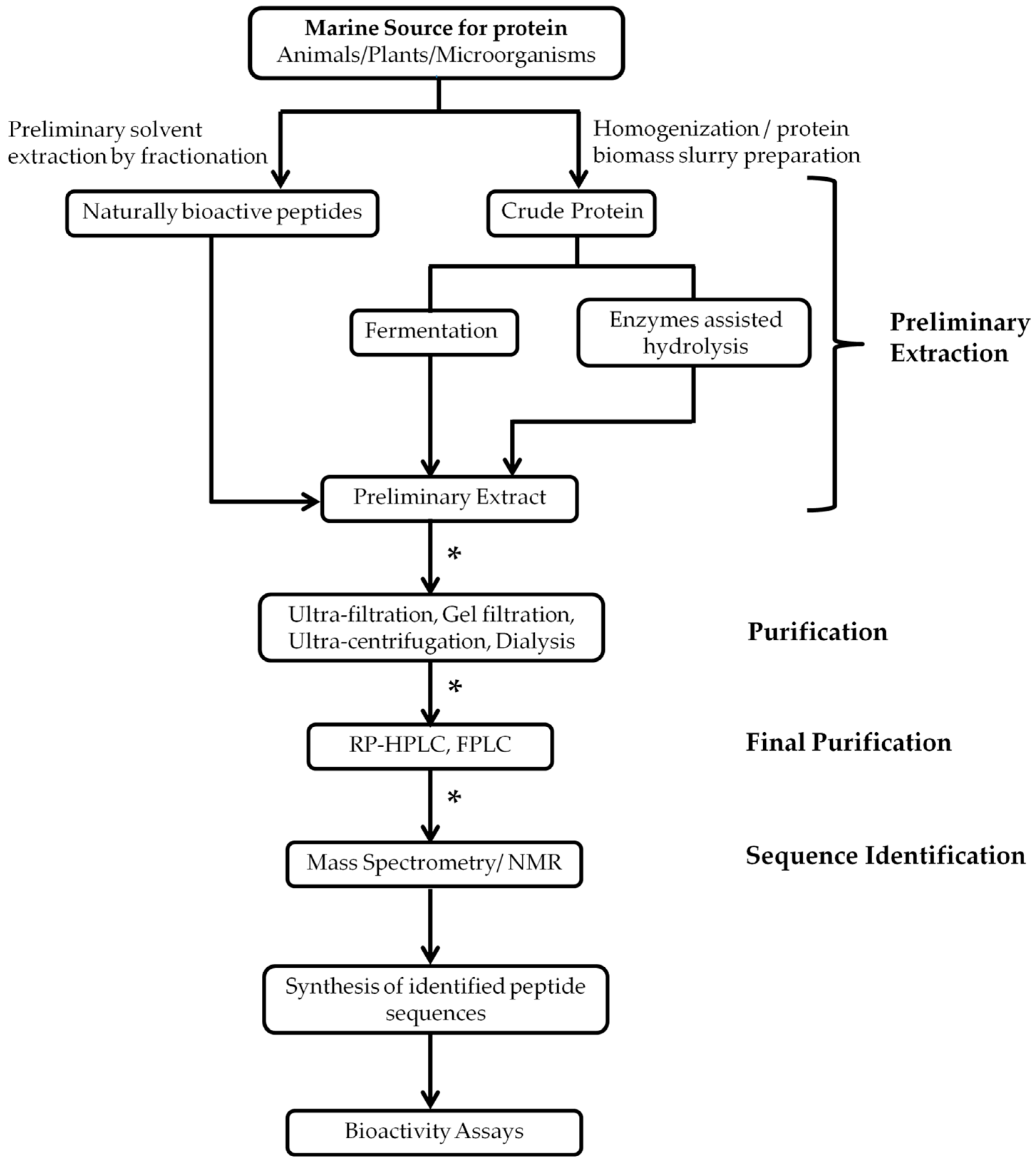
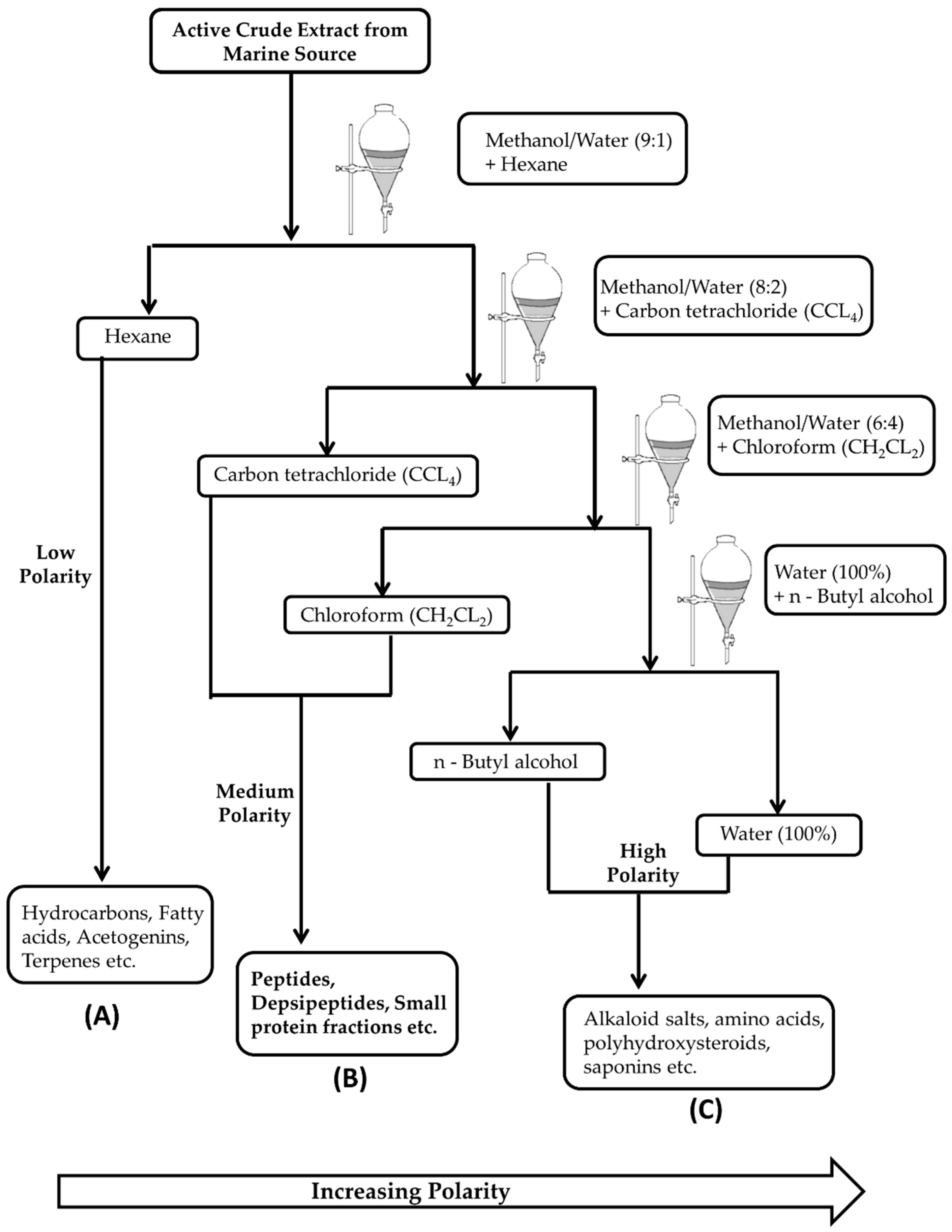
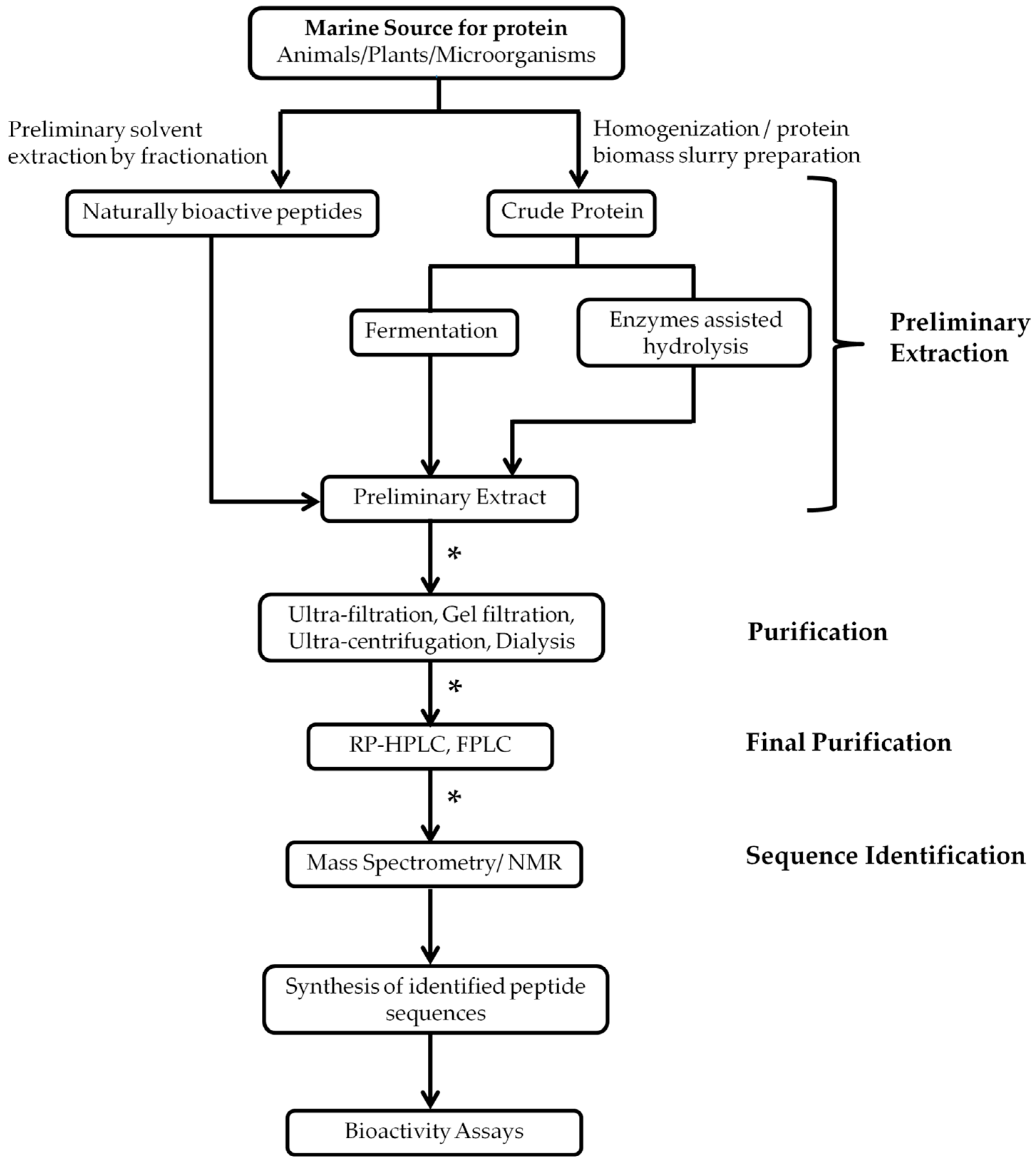
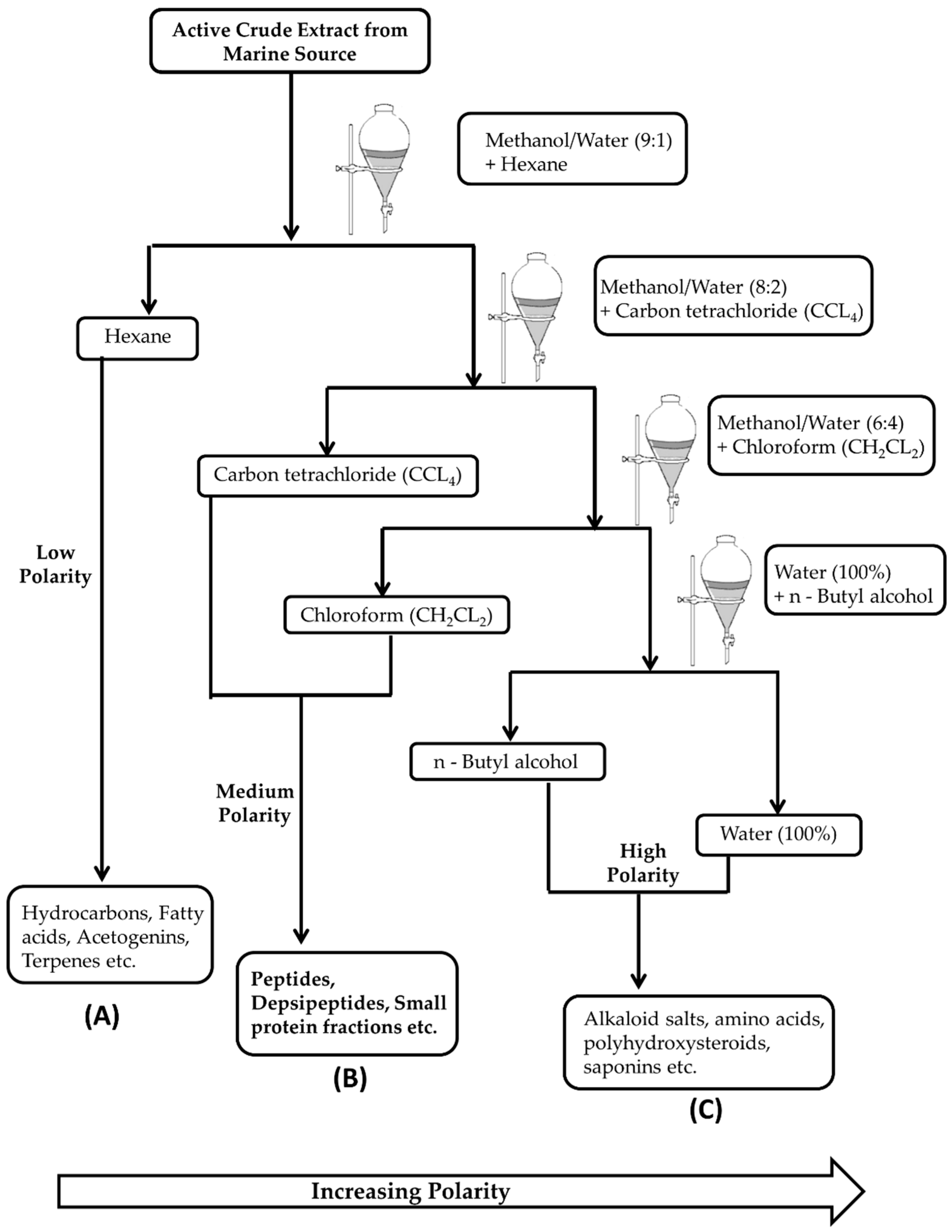
2. Isolation and Extraction of Marine Bioactive Peptides (MBPs)
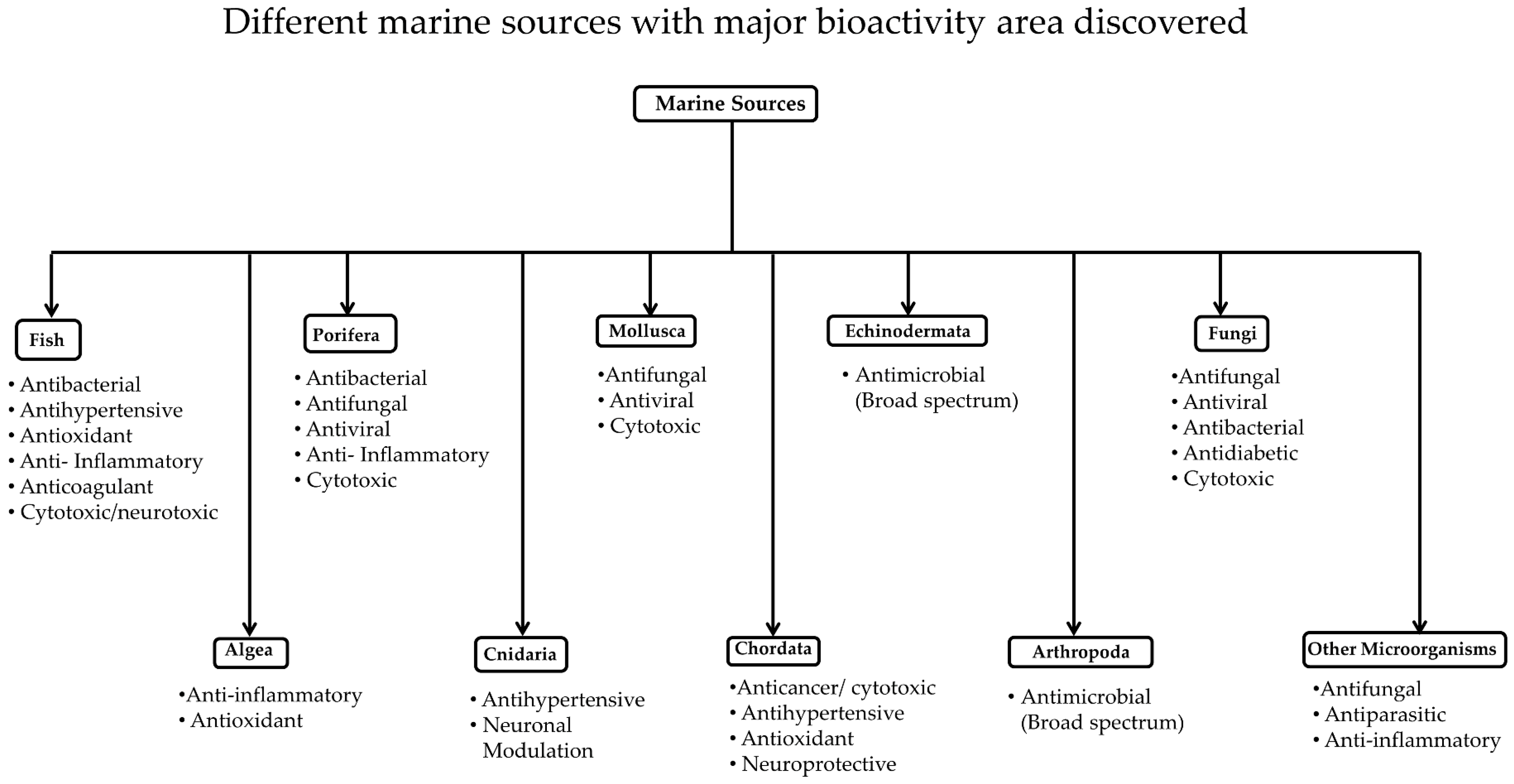
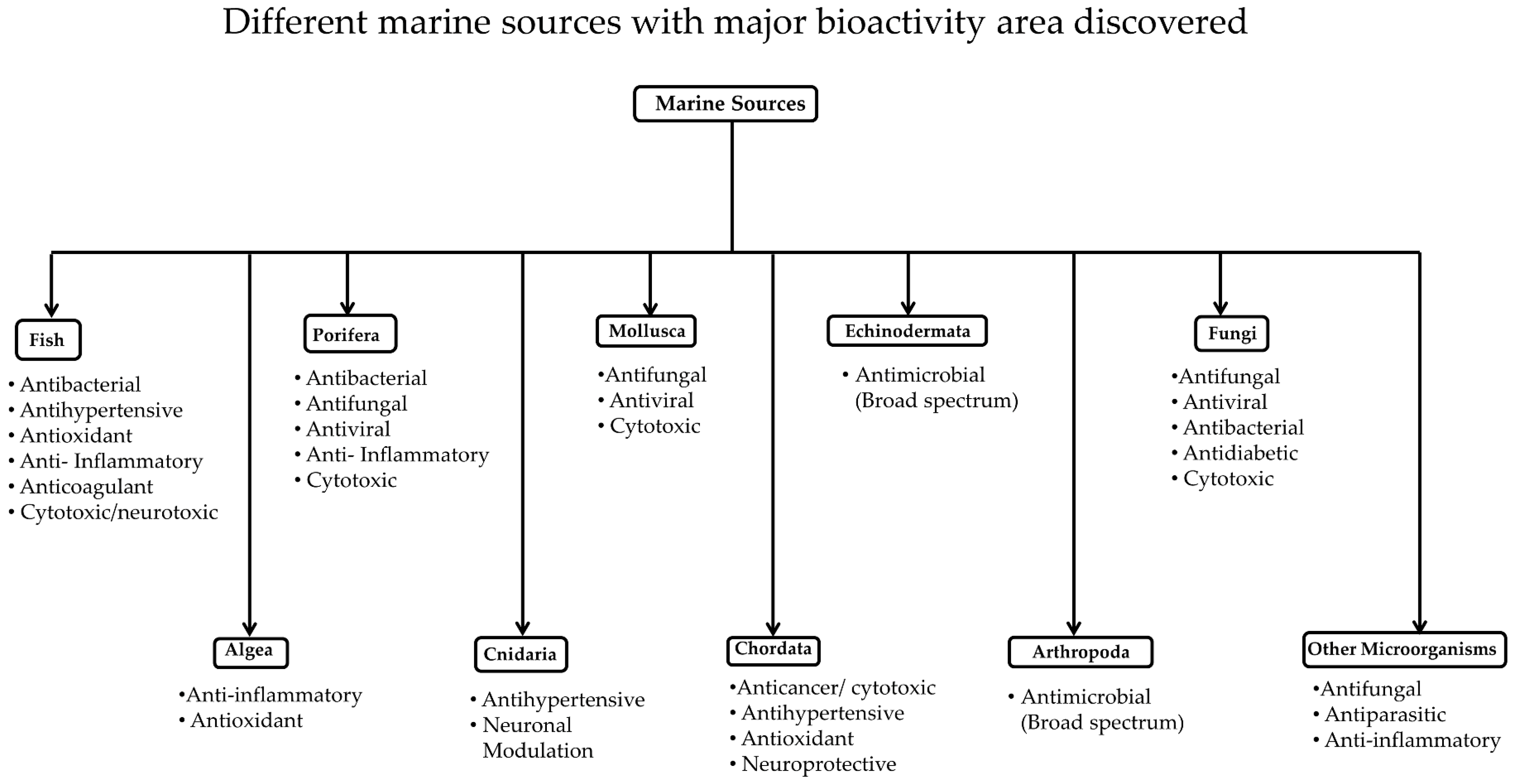
3. Classification of Marine Resources
3.1. Marine Invertebrates
3.1.1. Porifera
3.1.2. Cnidaria
3.1.3. Mollusca
3.1.4. Chordata
3.1.5. Echinodermata
3.1.6. Marine Arthropods
3.1.7. Algae, Fungi and Bacteria
3.2. Proteobacteria
3.3. Actinobacteria
4. Marine-Derived Products That Have In Vivo Efficacy
5. Drugs on the Market and in Clinical Trials
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fosgerau, K.; Hoffmann, T. Peptide therapeutics: Current status and future directions. Drug Discov. Today 2015, 20, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Padhi, A.; Sengupta, M.; Sengupta, S.; Roehm, K.H.; Sonawane, A. Antimicrobial peptides and proteins in mycobacterial therapy: Current status and future prospects. Tuberculosis 2014, 94, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Buchwald, H.; Dorman, R.B.; Rasmus, N.F.; Michalek, V.N.; Landvik, N.M.; Ikramuddin, S. Effects on GLP-1, PYY, and leptin by direct stimulation of terminal ileum and cecum in humans: Implications for ileal transposition. Surg. Obes. Relat. Dis. 2014, 10, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Khazanov, N.A.; Carlson, H.A. Exploring the composition of protein-ligand binding sites on a large scale. PLoS Comput. Biol. 2013, 9, e1003321. [Google Scholar] [CrossRef] [PubMed]
- Craik, D.J.; Fairlie, D.P.; Liras, S.; Price, D. The future of peptide-based drugs. Chem. Biol. Drug Des. 2013, 81, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Hirschmann, R.F.; Nicolaou, K.C.; Angeles, A.R.; Chen, J.S.; Smith, A.B., 3rd. The beta-d-glucose scaffold as a beta-turn mimetic. Acc. Chem. Res. 2009, 42, 1511–1520. [Google Scholar] [CrossRef] [PubMed]
- Jayatunga, M.K.; Thompson, S.; Hamilton, A.D. alpha-Helix mimetics: Outwards and upwards. Bioorg. Med. Chem. Lett. 2014, 24, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, C.L. Advances in peptide pharmaceuticals. Curr. Pharm. Biotechnol. 2009, 10, 122–137. [Google Scholar] [CrossRef] [PubMed]
- Banga, A.K. Therapeutic Peptides and Proteins: Formulation, Processing, and Delivery Systems, 3rd ed.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2015; p. 400. [Google Scholar]
- Pollaro, L.; Heinis, C. Strategies to prolong the plasma residence time of peptide drugs. MedChemComm 2010, 1, 319–324. [Google Scholar] [CrossRef]
- Hess, S.; Ovadia, O.; Shalev, D.E.; Senderovich, H.; Qadri, B.; Yehezkel, T.; Salitra, Y.; Sheynis, T.; Jelinek, R.; Gilon, C.; et al. Effect of structural and conformation modifications, including backbone cyclization, of hydrophilic hexapeptides on their intestinal permeability and enzymatic stability. J. Med. Chem. 2007, 50, 6201–6211. [Google Scholar] [CrossRef] [PubMed]
- Penchala, S.C.; Miller, M.R.; Pal, A.; Dong, J.; Madadi, N.R.; Xie, J.; Joo, H.; Tsai, J.; Batoon, P.; Samoshin, V.; et al. A biomimetic approach for enhancing the in vivo half-life of peptides. Nat. Chem. Biol. 2015, 11, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Pollaro, L.; Diderich, P.; Angelini, A.; Bellotto, S.; Wegner, H.; Heinis, C. Measuring net protease activities in biological samples using selective peptidic inhibitors. Anal. Biochem. 2012, 427, 18–20. [Google Scholar] [CrossRef] [PubMed]
- Craik, D.J.; Swedberg, J.E.; Mylne, J.S.; Cemazar, M. Cyclotides as a basis for drug design. Expert Opin. Drug Discov. 2012, 7, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Poth, A.G.; Chan, L.Y.; Craik, D.J. Cyclotides as grafting frameworks for protein engineering and drug design applications. Biopolymers 2013, 100, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Veber, D.F.; Johnson, S.R.; Cheng, H.Y.; Smith, B.R.; Ward, K.W.; Kopple, K.D. Molecular properties that influence the oral bioavailability of drug candidates. J. Med. Chem. 2002, 45, 2615–2623. [Google Scholar] [CrossRef] [PubMed]
- Vlieghe, P.; Lisowski, V.; Martinez, J.; Khrestchatisky, M. Synthetic therapeutic peptides: Science and market. Drug Discov. Today 2010, 15, 40–56. [Google Scholar] [CrossRef] [PubMed]
- Otvos, L., Jr.; Wade, J.D. Current challenges in peptide-based drug discovery. Front. Chem. 2014, 2, 62. [Google Scholar] [CrossRef] [PubMed]
- Lax, R. The Future of Peptide Development in the Pharmaceutical Industry. PharManuf. Int. Pept. Rev. 2010, 2, 10–15. [Google Scholar]
- Pukala, T.L.; Bowie, J.H.; Maselli, V.M.; Musgrave, I.F.; Tyler, M.J. Host-defence peptides from the glandular secretions of amphibians: Structure and activity. Nat. Prod. Rep. 2006, 23, 368–393. [Google Scholar] [CrossRef] [PubMed]
- Shaw, C. Advancing drug discovery with reptile and amphibian venom peptides—Venom-based medicines. Regul. Tech. 2009, 31, 34–37. [Google Scholar]
- Shaw, C.; Hirst, D.; Chen, T.; O’rourke, M.; Rao, P. Bradykinin B2 Receptor Antagonist Peptide from Amphibian Skin. Google Patents US8003610 B2, 23 August 2011. [Google Scholar]
- Li, J.; Xu, X.; Xu, C.; Zhou, W.; Zhang, K.; Yu, H.; Zhang, Y.; Zheng, Y.; Rees, H.H.; Lai, R.; Yang, D.; Wu, J. Anti-infection peptidomics of amphibian skin. Mol. Cell. Proteom. 2007, 6, 882–894. [Google Scholar] [CrossRef] [PubMed]
- Uhlig, T.; Kyprianou, T.; Martinelli, F.G.; Oppici, C.A.; Heiligers, D.; Hills, D.; Calvo, X.R.; Verhaert, P. The emergence of peptides in the pharmaceutical business: From exploration to exploitation. EuPA Open Proteom. 2014, 4, 58–69. [Google Scholar] [CrossRef]
- Bond, A. Exenatide (Byetta) as a novel treatment option for type 2 diabetes mellitus. Proc. (Bayl. Univ. Med. Cent.) 2006, 19, 281–284. [Google Scholar] [PubMed]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Wilmes, M.; Cammue, B.P.; Sahl, H.G.; Thevissen, K. Antibiotic activities of host defense peptides: More to it than lipid bilayer perturbation. Nat. Prod. Rep. 2011, 28, 1350–1358. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.V.; Yedery, R.D.; Aranha, C. Antimicrobial peptides: Premises and promises. Int. J. Antimicrob. Agents 2004, 24, 536–547. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jung, J.H.; Liu, Y. Antimicrobial Compounds from Marine Invertebrates-Derived Microorganisms. Curr. Med. Chem. 2016, 23, 2892–2905. [Google Scholar] [CrossRef] [PubMed]
- Anjum, K.; Abbas, S.Q.; Akhter, N.; Shagufta, B.I.; Shah, S.A.; Hassan, S.S. Emerging biopharmaceuticals from bioactive peptides derived from marine organisms. Chem. Biol. Drug Des. 2016. [Google Scholar] [CrossRef] [PubMed]
- Arnison, P.G.; Bibb, M.J.; Bierbaum, G.; Bowers, A.A.; Bugni, T.S.; Bulaj, G.; Camarero, J.A.; Campopiano, D.J.; Challis, G.L.; Clardy, J.; et al. Ribosomally synthesized and post-translationally modified peptide natural products: Overview and recommendations for a universal nomenclature. Nat. Prod. Rep. 2013, 30, 108–160. [Google Scholar] [CrossRef] [PubMed]
- Payne, J.A.; Schoppet, M.; Hansen, M.H.; Cryle, M.J. Diversity of nature’s assembly lines—Recent discoveries in non-ribosomal peptide synthesis. Mol. Biosyst. 2016, 13, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Desriac, F.; Jegou, C.; Balnois, E.; Brillet, B.; Le Chevalier, P.; Fleury, Y. Antimicrobial peptides from marine proteobacteria. Mar. Drugs 2013, 11, 3632–3660. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, S.; Adholeya, A.; Deshmukh, S.K. The Pharmacological Potential of Non-ribosomal Peptides from Marine Sponge and Tunicates. Front. Pharmacol. 2016, 7, 333. [Google Scholar] [CrossRef] [PubMed]
- Hetrick, K.J.; van der Donk, W.A. Ribosomally synthesized and post-translationally modified peptide natural product discovery in the genomic era. Curr. Opin. Chem. Biol. 2017, 38, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Cheung, R.C.; Ng, T.B.; Wong, J.H. Marine Peptides: Bioactivities and Applications. Mar. Drugs 2015, 13, 4006–4043. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.K.; Seo, C.H.; Park, Y. Marine peptides and their anti-infective activities. Mar. Drugs 2015, 13, 618–654. [Google Scholar] [CrossRef] [PubMed]
- Jo, C.; Khan, F.F.; Khan, M.I.; Iqbal, J. Marine bioactive peptides: Types, structures, and physiological functions. Food Rev. Int. 2017, 33, 44–61. [Google Scholar] [CrossRef]
- Bonar, D.B.; Weiner, R.M.; Colwell, R.R. Microbial-invertebrate interactions and potential for biotechnology. Microb. Ecol. 1986, 12, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Okami, Y. Marine microorganisms as a source of bioactive agents. Microb. Ecol. 1986, 12, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Hentschel, U.; Usher, K.M.; Taylor, M.W. Marine sponges as microbial fermenters. FEMS Microbiol. Ecol. 2006, 55, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Wahl, M.; Goecke, F.; Labes, A.; Dobretsov, S.; Weinberger, F. The second skin: Ecological role of epibiotic biofilms on marine organisms. Front. Microbiol. 2012, 3, 292. [Google Scholar] [CrossRef] [PubMed]
- Dubilier, N.; Bergin, C.; Lott, C. Symbiotic diversity in marine animals: The art of harnessing chemosynthesis. Nat. Rev. Microbiol. 2008, 6, 725–740. [Google Scholar] [CrossRef] [PubMed]
- Forest, R.; Victor, S.; Farooq, A.; Nancy, K. Diversity and distribution of coral-associated bacteria. Mar. Ecol. Prog. Ser. 2002, 243, 1–10. [Google Scholar]
- Kelman, D.; Kashman, Y.; Hill, R.T.; Rosenberg, E.; Loya, Y. Chemical warfare in the sea: The search for antibiotics from Red Sea corals and sponges. Pure. Appl. Chem. 2009, 81, 1113–1121. [Google Scholar] [CrossRef]
- Egan, S.; Harder, T.; Burke, C.; Steinberg, P.; Kjelleberg, S.; Thomas, T. The seaweed holobiont: Understanding seaweed-bacteria interactions. FEMS Microbiol. Rev. 2013, 37, 462–476. [Google Scholar] [CrossRef] [PubMed]
- Piel, J. Metabolites from symbiotic bacteria. Nat. Prod. Rep. 2004, 21, 519–538. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, E.W. Trading molecules and tracking targets in symbiotic interactions. Nat. Chem. Biol. 2008, 4, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Han, X.; Chen, H.; Lin, W.; Yan, X. Marine bacteria associated with marine macroorganisms: The potential antimicrobial resources. Ann. Microbiol. 2005, 55, 119–124. [Google Scholar]
- Wu, R.; Wu, C.; Liu, D.; Yang, X.; Huang, J.; Zhang, J.; Liao, B.; He, H.; Li, H. Overview of Antioxidant Peptides Derived from Marine Resources: The Sources, Characteristic, Purification, and Evaluation Methods. Appl. Biochem. Biotechnol. 2015, 176, 1815–1833. [Google Scholar] [CrossRef] [PubMed]
- Aneiros, A.; Garateix, A. Bioactive peptides from marine sources: Pharmacological properties and isolation procedures. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2004, 803, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Simmons, T.L.; Andrianasolo, E.; McPhail, K.; Flatt, P.; Gerwick, W.H. Marine natural products as anticancer drugs. Mol. Cancer Ther. 2005, 4, 333–342. [Google Scholar] [PubMed]
- Duggan, P.J.; Tuck, K.L. Bioactive Mimetics of Conotoxins and other Venom Peptides. Toxins (Basel) 2015, 7, 4175–4198. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Q. Genome-based studies of marine microorganisms to maximize the diversity of natural products discovery for medical treatments. Evid. Based Complement. Altern. Med. 2011, 2011, 3845–3872. [Google Scholar] [CrossRef] [PubMed]
- Trindade, M.; van Zyl, L.J.; Navarro-Fernandez, J.; Abd Elrazak, A. Targeted metagenomics as a tool to tap into marine natural product diversity for the discovery and production of drug candidates. Front. Microbiol. 2015, 6, 890. [Google Scholar] [CrossRef] [PubMed]
- Lluisma, A.O.; Milash, B.A.; Moore, B.; Olivera, B.M.; Bandyopadhyay, P.K. Novel venom peptides from the cone snail Conus pulicarius discovered through next-generation sequencing of its venom duct transcriptome. Mar. Genom. 2012, 5, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Jin, A.H.; Kaas, Q.; Jones, A.; Alewood, P.F.; Lewis, R.J. Deep venomics reveals the mechanism for expanded peptide diversity in cone snail venom. Mol. Cell. Proteom. 2013, 12, 312–329. [Google Scholar] [CrossRef] [PubMed]
- Cheung, R.C.; Wong, J.H.; Pan, W.L.; Chan, Y.S.; Yin, C.M.; Dan, X.L.; Wang, H.X.; Fang, E.F.; Lam, S.K.; Ngai, P.H.; et al. Antifungal and antiviral products of marine organisms. Appl. Microbiol. Biotechnol. 2014, 98, 3475–3494. [Google Scholar] [CrossRef] [PubMed]
- Jensen, I.J.; Maehre, H.K. Preclinical and Clinical Studies on Antioxidative, Antihypertensive and Cardioprotective Effect of Marine Proteins and Peptides-A Review. Mar. Drugs 2016, 14, 211. [Google Scholar] [CrossRef] [PubMed]
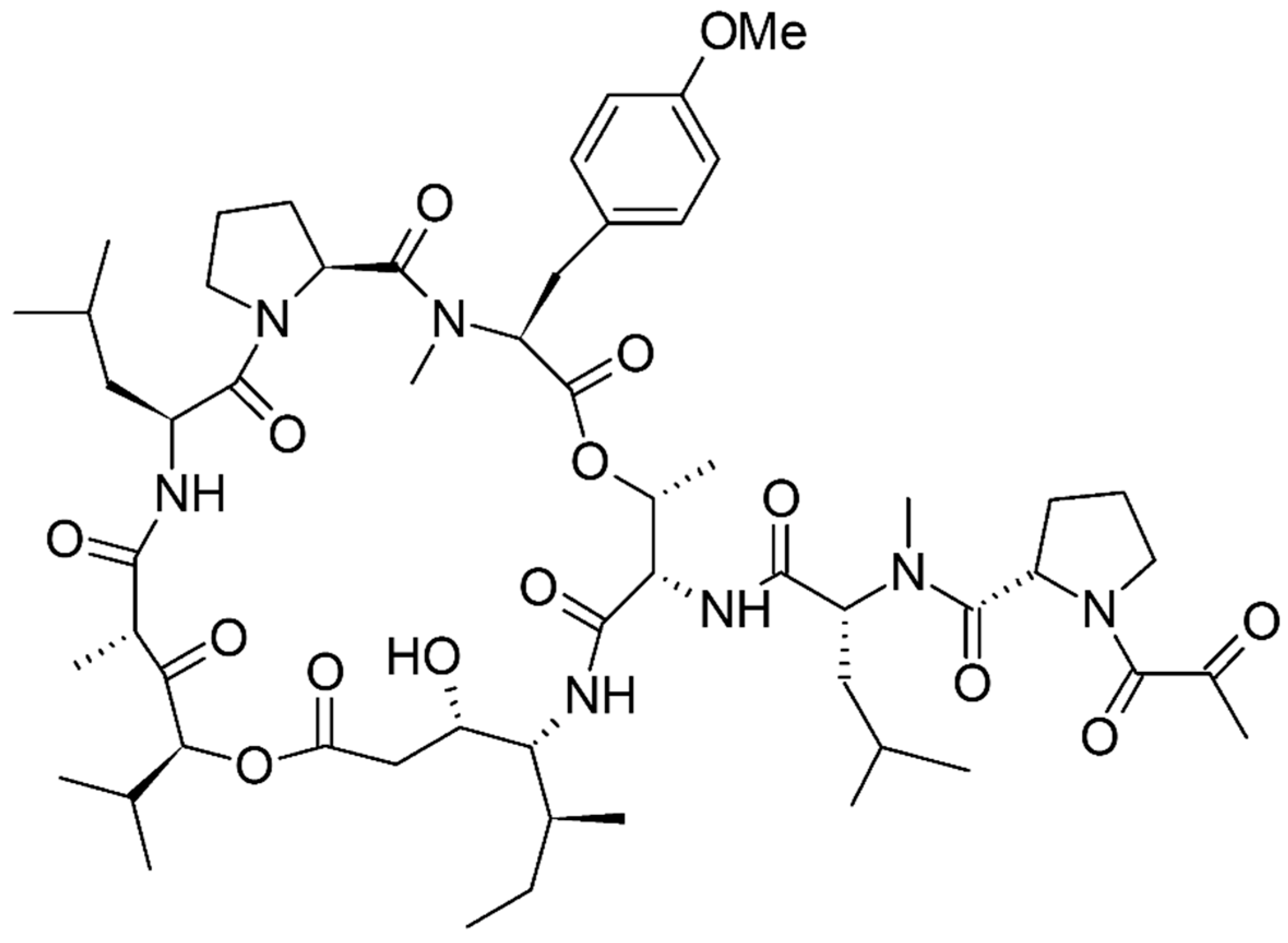
- Rangel, M.; de Santana, C.J.; Pinheiro, A.; Dos Anjos, L.; Barth, T.; Pires, O.R.; Fontes, W.; Castro, M.S. Marine Depsipeptides as Promising Pharmacotherapeutic Agents. Curr. Protein Pept. Sci. 2017, 18, 72–91. [Google Scholar] [CrossRef] [PubMed]
- Andavan, G.S.; Lemmens-Gruber, R. Cyclodepsipeptides from marine sponges: Natural agents for drug research. Mar. Drugs 2010, 8, 810–834. [Google Scholar] [CrossRef] [PubMed]
- Plaza, A.; Gustchina, E.; Baker, H.L.; Kelly, M.; Bewley, C.A. Mirabamides A-D, depsipeptides from the sponge Siliquariaspongia mirabilis that inhibit HIV-1 fusion. J. Nat. Prod. 2007, 70, 1753–1760. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Van Wagoner, R.M.; Harper, M.K.; Baker, H.L.; Hooper, J.N.; Bewley, C.A.; Ireland, C.M. Mirabamides E-H, HIV-inhibitory depsipeptides from the sponge Stelletta clavosa. J. Nat. Prod. 2011, 74, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Ford, P.W.; Gustafson, K.R.; McKee, T.C.; Shigematsu, N.; Maurizi, L.K.; Pannell, L.K.; Williams, D.E.; Dilip de Silva, E.; Lassota, P.; Allen, T.M.; Van Soest, R.; Andersen, R.J.; Boyd, M.R. Papuamides A–D, HIV-Inhibitory and Cytotoxic Depsipeptides from the Sponges Theonella mirabilis and Theonella swinhoei Collected in Papua New Guinea. J. Am. Chem. Soc. 1999, 121, 5899–5909. [Google Scholar] [CrossRef]
- Prasad, P.; Aalbersberg, W.; Feussner, K.D.; Van Wagoner, R.M. Papuamides E and F, Cytotoxic Depsipeptides from the Marine Sponge Melophlus sp. Tetrahedron 2011, 67, 8529–8531. [Google Scholar] [CrossRef] [PubMed]
- Plaza, A.; Bifulco, G.; Keffer, J.L.; Lloyd, J.R.; Baker, H.L.; Bewley, C.A. Celebesides A–C and theopapuamides B-D, depsipeptides from an Indonesian sponge that inhibit HIV-1 entry. J. Org. Chem. 2009, 74, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Zampella, A.; D’Auria, M.V.; Paloma, L.G.; Casapullo, A.; Minale, L.; Debitus, C.; Henin, Y. Callipeltin A, an Anti-HIV Cyclic Depsipeptide from the New Caledonian Lithistida Sponge Callipelta sp. J. Am. Chem. Soc. 1996, 118, 6202–6209. [Google Scholar] [CrossRef]
- Kikuchi, M.; Konno, H. Cytotoxic evaluation of natural and synthetic callipeltins: A revision of cytotoxicity of callipeltin B. Biosci. Biotechnol. Biochem. 2016, 80, 1066–1069. [Google Scholar] [CrossRef] [PubMed]
- Daletos, G.; Kalscheuer, R.; Koliwer-Brandl, H.; Hartmann, R.; de Voogd, N.J.; Wray, V.; Lin, W.; Proksch, P. Callyaerins from the Marine Sponge Callyspongia aerizusa: Cyclic Peptides with Antitubercular Activity. J. Nat. Prod. 2015, 78, 1910–1925. [Google Scholar] [CrossRef] [PubMed]
- Otero-Gonzalez, A.J.; Magalhaes, B.S.; Garcia-Villarino, M.; Lopez-Abarrategui, C.; Sousa, D.A.; Dias, S.C.; Franco, O.L. Antimicrobial peptides from marine invertebrates as a new frontier for microbial infection control. FASEB J. 2010, 24, 1320–1334. [Google Scholar] [CrossRef] [PubMed]
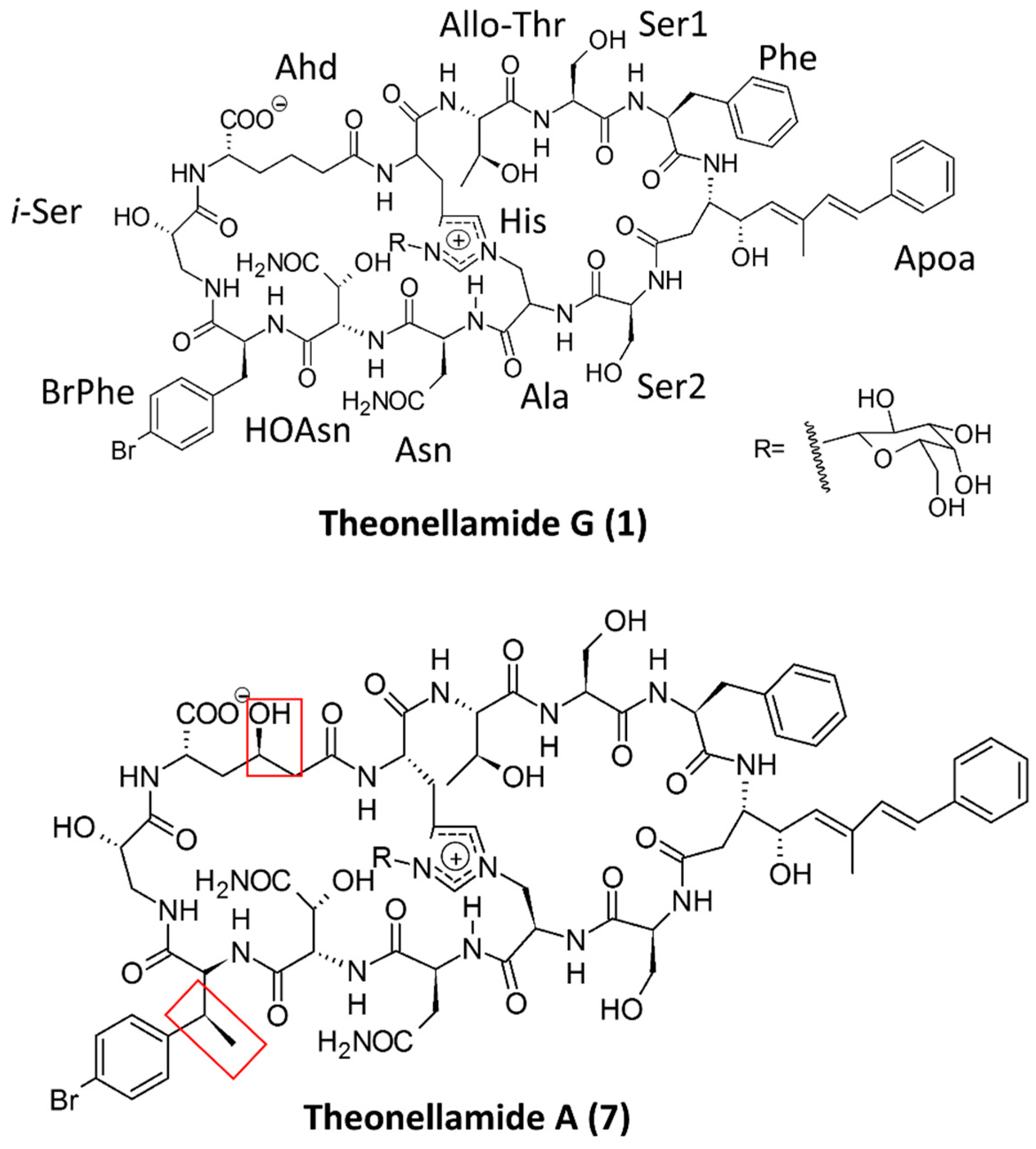
- Youssef, D.T.A.; Shaala, L.A.; Mohamed, G.A.; Badr, J.M.; Bamanie, F.H.; Ibrahim, S.R.M. Theonellamide G, a Potent Antifungal and Cytotoxic Bicyclic Glycopeptide from the Red Sea Marine Sponge Theonella swinhoei. Mar. Drugs 2014, 12, 1911–1923. [Google Scholar] [CrossRef] [PubMed]
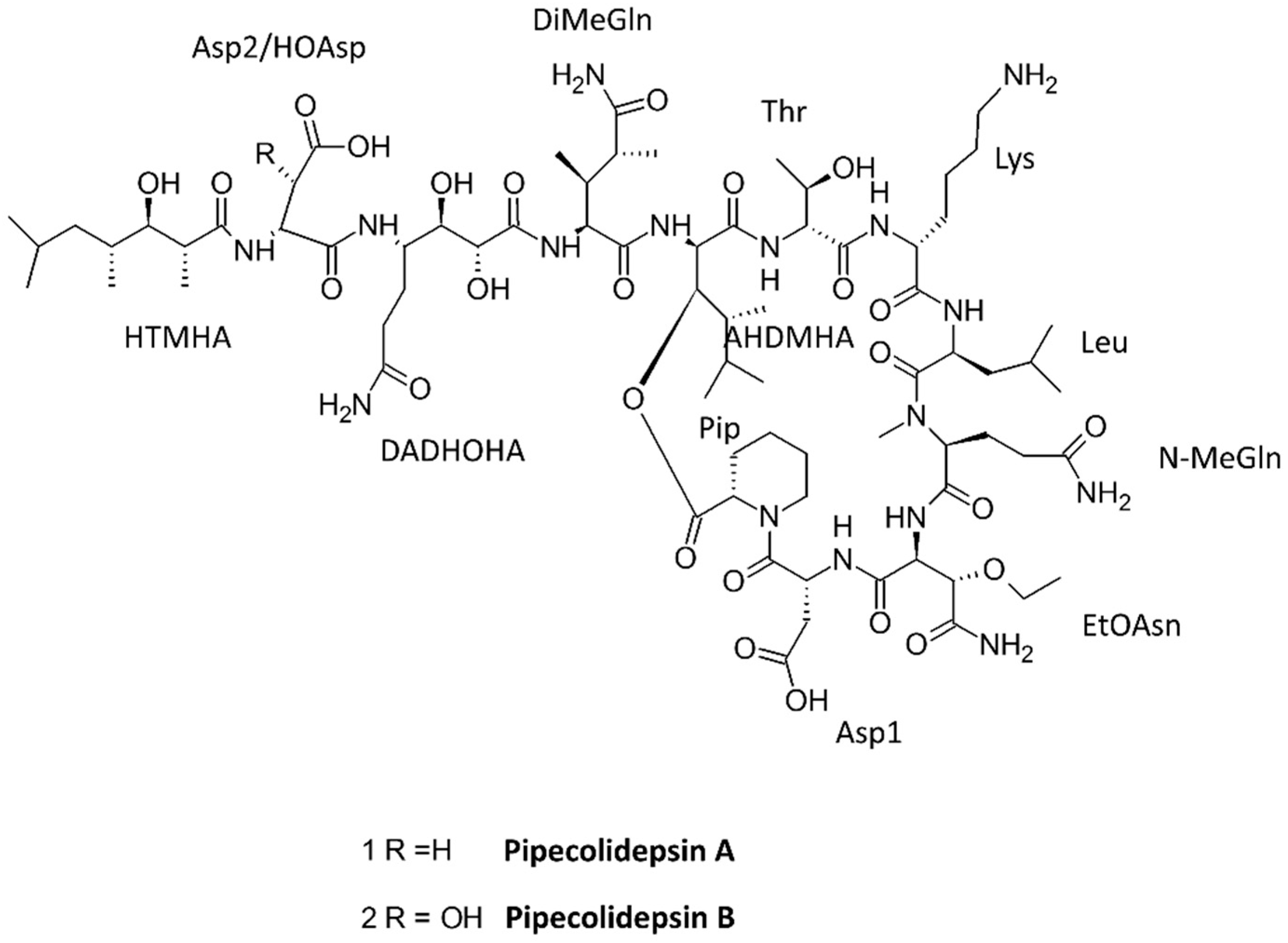
- Coello, L.; Reyes, F.; Martin, M.J.; Cuevas, C.; Fernandez, R. Isolation and structures of pipecolidepsins A and B, cytotoxic cyclic depsipeptides from the Madagascan sponge Homophymia lamellosa. J. Nat. Prod. 2014, 77, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Espiritu, R.A.; Cornelio, K.; Kinoshita, M.; Matsumori, N.; Murata, M.; Nishimura, S.; Kakeya, H.; Yoshida, M.; Matsunaga, S. Marine sponge cyclic peptide theonellamide A disrupts lipid bilayer integrity without forming distinct membrane pores. Biochim. Biophys. Acta 2016, 1858, 1373–1379. [Google Scholar] [CrossRef] [PubMed]
- Youssef, D.T.; Shaala, L.A.; Mohamed, G.A.; Badr, J.M.; Bamanie, F.H.; Ibrahim, S.R. Theonellamide G, a potent antifungal and cytotoxic bicyclic glycopeptide from the Red Sea marine sponge Theonella swinhoei. Mar. Drugs 2014, 12, 1911–1923. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.D.; Pham, N.B.; Fechner, G.A.; Hooper, J.N.; Quinn, R.J. Potent cytotoxic peptides from the Australian marine sponge Pipestela candelabra. Mar. Drugs 2014, 12, 3399–3415. [Google Scholar] [CrossRef] [PubMed]
- Carstens, B.B.; Rosengren, K.J.; Gunasekera, S.; Schempp, S.; Bohlin, L.; Dahlstrom, M.; Clark, R.J.; Goransson, U. Isolation, Characterization, and Synthesis of the Barrettides: Disulfide-Containing Peptides from the Marine Sponge Geodia barretti. J. Nat. Prod. 2015, 78, 1886–1893. [Google Scholar] [CrossRef] [PubMed]
- Plaza, A.; Bifulco, G.; Masullo, M.; Lloyd, J.R.; Keffer, J.L.; Colin, P.L.; Hooper, J.N.A.; Bell, L.J.; Bewley, C.A. Mutremdamide A and Koshikamides C–H, Peptide Inhibitors of HIV-1 Entry from Different Theonella Species. J. Org. Chem. 2010, 75, 4344–4355. [Google Scholar] [CrossRef] [PubMed]
- Lazcano-Perez, F.; Castro, H.; Arenas, I.; Garcia, D.E.; Gonzalez-Munoz, R.; Arreguin-Espinosa, R. Activity of Palythoa caribaeorum Venom on Voltage-Gated Ion Channels in Mammalian Superior Cervical Ganglion Neurons. Toxins (Basel) 2016, 8, 135. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Sun, L.; Zhang, Y.; Liu, G. Antihypertensive effect of long-term oral administration of jellyfish (Rhopilema esculentum) collagen peptides on renovascular hypertension. Mar. Drugs 2012, 10, 417–426. [Google Scholar] [CrossRef] [PubMed]
- So, P.B.; Rubio, P.; Lirio, S.; Macabeo, A.P.; Huang, H.Y.; Corpuz, M.J.; Villaflores, O.B. In vitro angiotensin I converting enzyme inhibition by a peptide isolated from Chiropsalmus quadrigatus Haeckel (box jellyfish) venom hydrolysate. Toxicon 2016, 119, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Gur Barzilai, M.; Kahn, R.; Regev, N.; Gordon, D.; Moran, Y.; Gurevitz, M. The specificity of Av3 sea anemone toxin for arthropods is determined at linker DI/SS2-S6 in the pore module of target sodium channels. Biochem. J. 2014, 463, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Shilabin, A.G.; Hamann, M.T. In vitro and in vivo evaluation of select kahalalide F analogs with antitumor and antifungal activities. Bioorg. Med. Chem. 2011, 19, 6628–6632. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Abarrategui, C.; Alba, A.; Silva, O.N.; Reyes-Acosta, O.; Vasconcelos, I.M.; Oliveira, J.T.; Migliolo, L.; Costa, M.P.; Costa, C.R.; Silva, M.R.; et al. Functional characterization of a synthetic hydrophilic antifungal peptide derived from the marine snail Cenchritis muricatus. Biochimie 2012, 94, 968–974. [Google Scholar] [CrossRef] [PubMed]
- López-Abarrategui, C.; McBeth, C.; Mandal, S.M.; Sun, Z.J.; Heffron, G.; Alba-Menéndez, A.; Migliolo, L.; Reyes-Acosta, O.; García-Villarino, M.; Nolasco, D.O.; et al. Cm-p5: An antifungal hydrophilic peptide derived from the coastal mollusk Cenchritis muricatus (Gastropoda: Littorinidae). FASEB J. 2015, 29, 3315–3325. [Google Scholar] [CrossRef] [PubMed]
- Franklin, J.B.; Rajesh, R.P. A sleep-inducing peptide from the venom of the Indian cone snail Conus araneosus. Toxicon 2015, 103, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.K.; Zhang, L.; Smith, M.D.; Walewska, A.; Vellore, N.A.; Baron, R.; McIntosh, J.M.; White, H.S.; Olivera, B.M.; Bulaj, G. A marine analgesic peptide, Contulakin-G, and neurotensin are distinct agonists for neurotensin receptors: Uncovering structural determinants of desensitization properties. Front. Pharmacol. 2015, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.-C.; Kim, J.-I.; Park, S.-J.; Jung, W.-K.; Jeon, Y.-J. Antihypertensive peptide purified from Styela clava flesh tissue stimulates glucose uptake through AMP-activated protein kinase (AMPK) activation in skeletal muscle cells. Eur. Food Res. Technol. 2016, 242, 163–170. [Google Scholar] [CrossRef]
- Chi, C.-F.; Hu, F.-Y.; Wang, B.; Ren, X.-J.; Deng, S.-G.; Wu, C.-W. Purification and characterization of three antioxidant peptides from protein hydrolyzate of croceine croaker (Pseudosciaena crocea) muscle. Food Chem. 2015, 168, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Pangestuti, R.; Ryu, B.; Himaya, S.; Kim, S.-K. Optimization of hydrolysis conditions, isolation, and identification of neuroprotective peptides derived from seahorse Hippocampus trimaculatus. Amino Acids 2013, 45, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Wei, J.; Huang, X.; Huang, Y.; Qin, Q. Antiviral effects of beta-defensin derived from orange-spotted grouper (Epinephelus coioides). Fish Shellfish Immunol. 2012, 32, 828–838. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Jimenez, G.M.; Burgos-Hernandez, A.; Ezquerra-Brauer, J.M. Bioactive peptides and depsipeptides with anticancer potential: Sources from marine animals. Mar. Drugs 2012, 10, 963–986. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.Y.; Chi, C.F.; Wang, B.; Deng, S.G. Two novel antioxidant nonapeptides from protein hydrolysate of skate (Raja porosa) muscle. Mar. Drugs 2015, 13, 1993–2009. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.F.; Hu, F.Y.; Wang, B.; Li, Z.R.; Luo, H.Y. Influence of Amino Acid Compositions and Peptide Profiles on Antioxidant Capacities of Two Protein Hydrolysates from Skipjack Tuna (Katsuwonus pelamis) Dark Muscle. Mar. Drugs 2015, 13, 2580–2601. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.C.; Hsieh, H.J.; Hwang, D.F. Cytotoxic and apoptotic activities of the plancitoxin I from the venom of crown-of-thorns starfish (Acanthaster planci) on A375.S2 cells. J. Appl. Toxicol. 2015, 35, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Haug, T.; Moe, M.K.; Styrvold, O.B.; Stensvag, K. Centrocins: Isolation and characterization of novel dimeric antimicrobial peptides from the green sea urchin, Strongylocentrotus droebachiensis. Dev. Comp. Immunol. 2010, 34, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Shan, Z.; Zhu, K.; Peng, H.; Chen, B.; Liu, J.; Chen, F.; Ma, X.; Wang, S.; Qiao, K.; Wang, K. The New Antimicrobial Peptide SpHyastatin from the Mud Crab Scylla paramamosain with Multiple Antimicrobial Mechanisms and High Effect on Bacterial Infection. Front. Microbiol. 2016, 7, 1140. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Fan, D.Q.; Zhu, K.X.; Shan, Z.G.; Chen, F.Y.; Hou, L.; Cai, L.; Wang, K.J. Mechanism study on a new antimicrobial peptide Sphistin derived from the N-terminus of crab histone H2A identified in haemolymphs of Scylla paramamosain. Fish Shellfish Immunol. 2015, 47, 833–846. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.N.; Rajanbabu, V.; Pan, C.Y.; Chan, Y.L.; Chen, J.Y.; Wu, C.J. Enhanced Control of Bladder-Associated Tumors Using Shrimp Anti-Lipopolysaccharide Factor (SALF) Antimicrobial Peptide as a Cancer Vaccine Adjuvant in Mice. Mar. Drugs 2015, 13, 3241–3258. [Google Scholar] [CrossRef] [PubMed]
- Rolland, J.L.; Abdelouahab, M.; Dupont, J.; Lefevre, F.; Bachere, E.; Romestand, B. Stylicins, a new family of antimicrobial peptides from the Pacific blue shrimp Litopenaeus stylirostris. Mol. Immunol. 2010, 47, 1269–1277. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.J.; Liu, R.F.; Li, Y.G.; Tao, L.M.; Tian, L. Two new antifungal cyclic lipopeptides from Bacillus marinus B-9987. Chem. Pharm. Bull. (Tokyo) 2010, 58, 1630–1634. [Google Scholar] [CrossRef] [PubMed]
- Motohashi, K.; Takagi, M.; Shin-ya, K. Tetrapeptides Possessing a Unique Skeleton, JBIR-34 and JBIR-35, Isolated from a Sponge-Derived Actinomycete, Streptomyces sp. Sp080513GE-23. J. Nat. Prod. 2010, 73, 226–228. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Wang, N.; Hu, J.; Wang, S. Isolation and characterization of a new iturinic lipopeptide, mojavensin A produced by a marine-derived bacterium Bacillus mojavensis B0621A. J. Antibiot. (Tokyo) 2012, 65, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Um, S.; Pyee, Y.; Kim, E.H.; Lee, S.K.; Shin, J.; Oh, D.C. Thalassospiramide G, a new gamma-amino-acid-bearing peptide from the marine bacterium Thalassospira sp. Mar. Drugs 2013, 11, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.; Chung, B.; Oh, K.B.; Shin, J.; Oh, D.C. Hormaomycins B and C: New Antibiotic Cyclic Depsipeptides from a Marine Mudflat-Derived Streptomyces sp. Mar. Drugs 2015, 13, 5187–5200. [Google Scholar] [CrossRef] [PubMed]
- Pruksakorn, P.; Arai, M.; Kotoku, N.; Vilcheze, C.; Baughn, A.D.; Moodley, P.; Jacobs, W.R., Jr.; Kobayashi, M. Trichoderins, novel aminolipopeptides from a marine sponge-derived Trichoderma sp., are active against dormant mycobacteria. Bioorg. Med. Chem. Lett. 2010, 20, 3658–3663. [Google Scholar] [CrossRef] [PubMed]
- You, M.; Liao, L.; Hong, S.H.; Park, W.; Kwon, D.I.; Lee, J.; Noh, M.; Oh, D.-C.; Oh, K.-B.; Shin, J. Lumazine Peptides from the Marine-Derived Fungus Aspergillus terreus. Mar. Drugs 2015, 13, 1290–1303. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.A.; Kim, I.H.; Nam, T.J. Bioactive peptide from Pyropia yezoensis and its anti-inflammatory activities. Int. J. Mol. Med. 2015, 36, 1701–1706. [Google Scholar] [CrossRef] [PubMed]
- Falkenberg, S.S. Discovery and Characterization of Novel Bioacive Peptides from Marine Secondary Products. Ph.D. Thesis, Electronic, Technical University of Denmark, Kongens Lyngby, Denmark, 2014. [Google Scholar]
- Lemes, A.C.; Sala, L.; Ores Jda, C.; Braga, A.R.; Egea, M.B.; Fernandes, K.F. A Review of the Latest Advances in Encrypted Bioactive Peptides from Protein-Rich Waste. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Cole, A.M.; Weis, P.; Diamond, G. Isolation and characterization of pleurocidin, an antimicrobial peptide in the skin secretions of winter flounder. J. Biol. Chem. 1997, 272, 12008–12013. [Google Scholar] [CrossRef] [PubMed]
- Song, R.; Wei, R.B.; Luo, H.Y.; Wang, D.F. Isolation and characterization of an antibacterial peptide fraction from the pepsin hydrolysate of half-fin anchovy (Setipinna taty). Molecules 2012, 17, 2980–2991. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sadiq, F.A.; Fu, L.; Zhu, H.; Zhong, M.; Sohail, M. Identification of Angiotensin I-Converting Enzyme Inhibitory Peptides Derived from Enzymatic Hydrolysates of Razor Clam Sinonovacula constricta. Mar. Drugs 2016, 14, 110. [Google Scholar] [CrossRef] [PubMed]
- Kiran N, S.G.; Khan, A.N.; Ibrar, K.; Tushar, P. Extraction and Screening of Bioactive Compounds with Antimicrobial Properties from Selected Species of Mollusk and Crustacean. J. Clin. Cell Immunol. 2014, 5, 1–5. [Google Scholar]
- Riguera, R. Isolating bioactive compounds from marine organisms. J. Mar. Biotechnol. 1997, 5, 187–193. [Google Scholar]
- Sruthy, K.S.; Chaithanya, E.R.; Sathyan, N.; Anilkumar, P.R.; Singh, I.S.B.; Philip, R. Isolation, Purification and Characterization of Antimicrobial Peptides from Indian Ruff, Psenopsis cyanea (Alcock, 1890). Int. J. Res. Biosci. 2015, 5, 1–6. [Google Scholar]
- Martínez-Maqueda, D.; Hernández-Ledesma, B.; Amigo, L.; Miralles, B.; Gómez-Ruiz, J.Á. Extraction/fractionation techniques for proteins and peptides and protein digestion. In Proteomics in Foods: Principles and Applications; Toldrá, F., Nollet, L.M.L., Eds.; Springer: Boston, MA, USA, 2013; pp. 21–50. [Google Scholar]
- Rajalingam, D.; Loftis, C.; Xu, J.J.; Kumar, T.K. Trichloroacetic acid-induced protein precipitation involves the reversible association of a stable partially structured intermediate. Protein Sci. 2009, 18, 980–993. [Google Scholar] [CrossRef] [PubMed]
- Se-Kwon Kim, I.W. Development and biological activities of marine-derived bioactive peptides: A review. J. Funct. Foods 2010, 2, 1–9. [Google Scholar]
- Dai-Hung Ngo, T.-S.V.; Kim, S.-K. Marine Proteins and Peptides: Biological Activities and Applications, 1st ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2013; p. 816. [Google Scholar]
- Sampath Kumar, N.S.; Nazeer, R.A.; Jaiganesh, R. Purification and identification of antioxidant peptides from the skin protein hydrolysate of two marine fishes, horse mackerel (Magalaspis cordyla) and croaker (Otolithes ruber). Amino Acids 2012, 42, 1641–1649. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Bai, L.; Zhu, L.; Yang, L.; Zhang, X. Marine algae-derived bioactive peptides for human nutrition and health. J. Agric. Food Chem. 2014, 62, 9211–9222. [Google Scholar] [CrossRef] [PubMed]
- Gobbetti, M.; Stepaniak, L.; De Angelis, M.; Corsetti, A.; Di Cagno, R. Latent bioactive peptides in milk proteins: Proteolytic activation and significance in dairy processing. Crit. Rev. Food Sci. Nutr. 2002, 42, 223–239. [Google Scholar] [CrossRef] [PubMed]
- Sipola, M.; Finckenberg, P.; Korpela, R.; Vapaatalo, H.; Nurminen, M.L. Effect of long-term intake of milk products on blood pressure in hypertensive rats. J. Dairy Res. 2002, 69, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Ryan, J.T.; Ross, R.P.; Bolton, D.; Fitzgerald, G.F.; Stanton, C. Bioactive peptides from muscle sources: Meat and fish. Nutrients 2011, 3, 765–791. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.K.; He, H.L.; Chen, X.L.; Sun, C.Y.; Zhang, Y.Z.; Zhou, B.C. Production of novel angiotensin I-converting enzyme inhibitory peptides by fermentation of marine shrimp Acetes chinensis with Lactobacillus fermentum SM 605. Appl. Microbiol. Biotechnol. 2008, 79, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, J.; Hu, G.; Yu, J.; Zhu, X.; Lin, Y.; Chen, S.; Yuan, J. Statistical research on the bioactivity of new marine natural products discovered during the 28 years from 1985 to 2012. Mar. Drugs 2015, 13, 202–221. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.P.; Yuan, J.; Sun, L.; She, Z.G.; Wu, J.H.; Lan, X.J.; Zhu, X.; Lin, Y.C.; Chen, S.P. Statistical research on marine natural products based on data obtained between 1985 and 2008. Mar. Drugs 2011, 9, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Zhao, Q.Y.; Xue, S.; Zhang, W. Bioactive compounds from marine sponges and cell culture of marine sponges. Sheng Wu Gong Cheng Xue Bao 2002, 18, 10–15. [Google Scholar] [PubMed]
- Ibrahim, S.R.; Min, C.C.; Teuscher, F.; Ebel, R.; Kakoschke, C.; Lin, W.; Wray, V.; Edrada-Ebel, R.; Proksch, P. Callyaerins A–F and H, new cytotoxic cyclic peptides from the Indonesian marine sponge Callyspongia aerizusa. Bioorg. Med. Chem. 2010, 18, 4947–4956. [Google Scholar] [CrossRef] [PubMed]
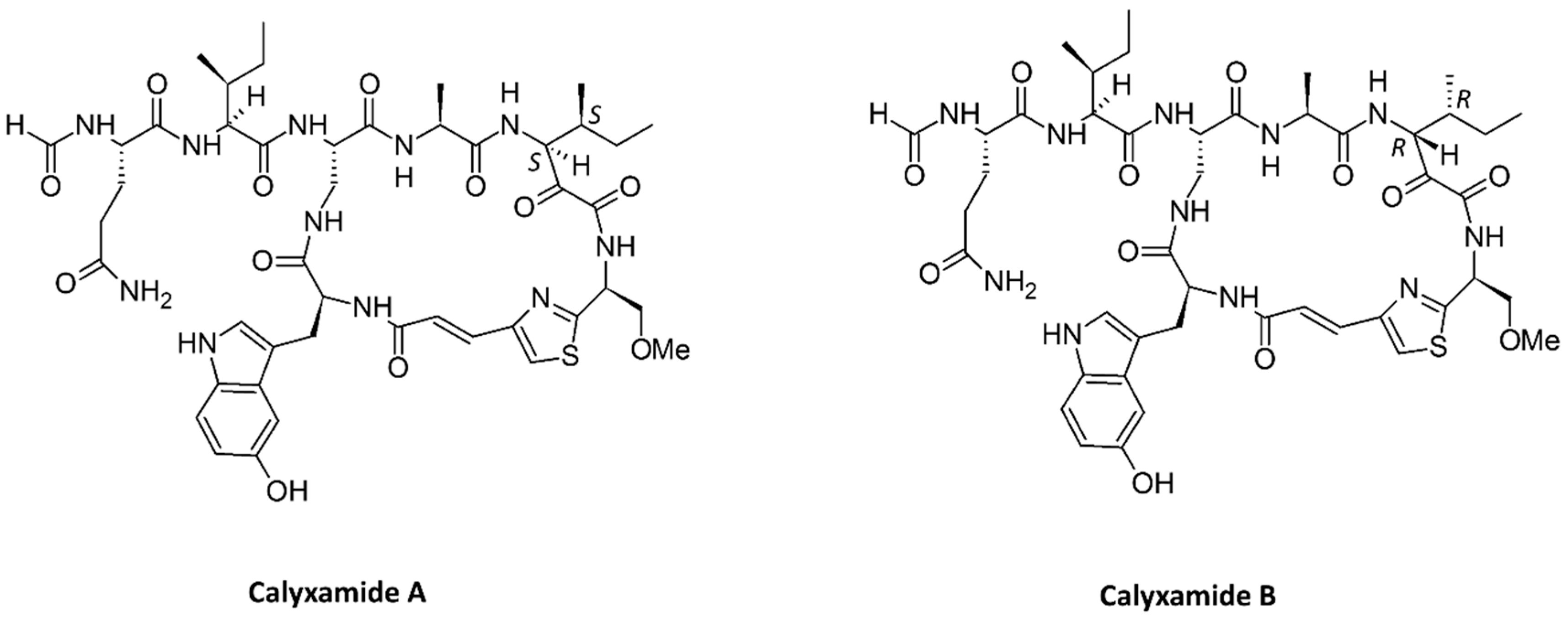
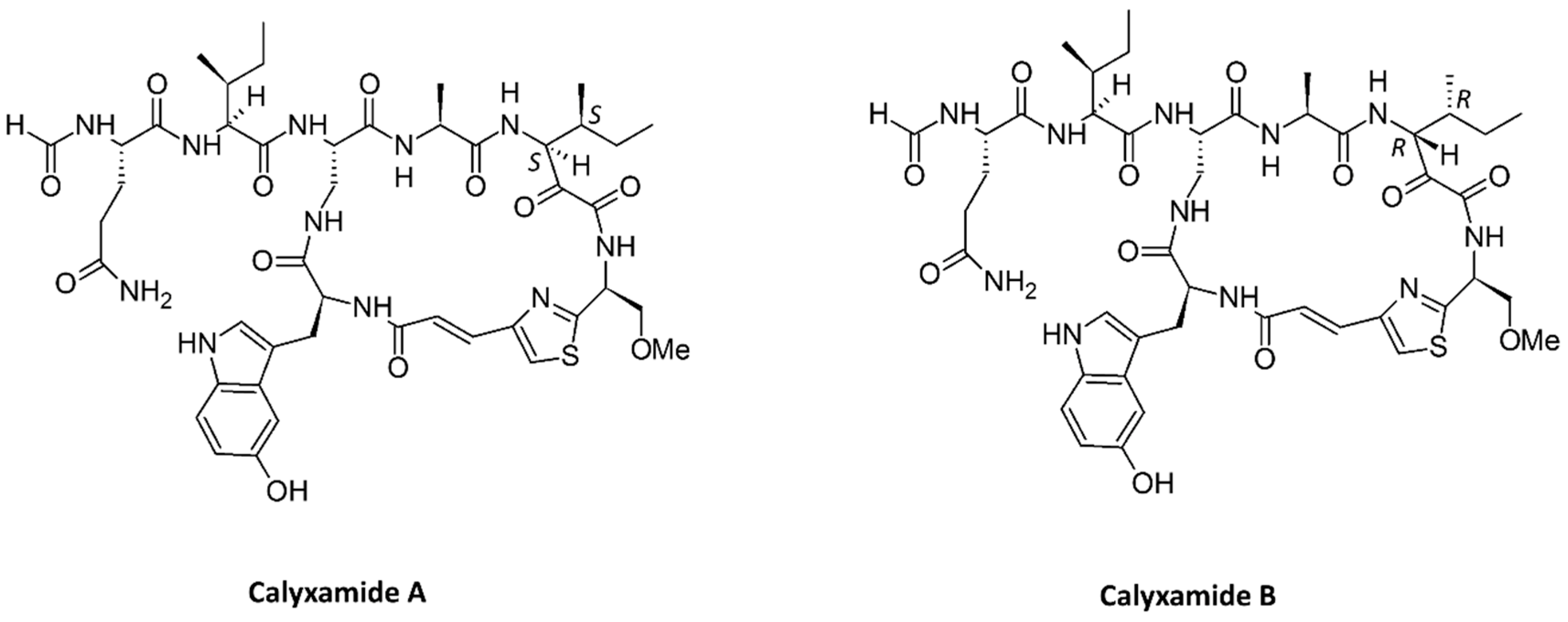
- Kimura, M.; Wakimoto, T.; Egami, Y.; Tan, K.C.; Ise, Y.; Abe, I. Calyxamides A and B, cytotoxic cyclic peptides from the marine sponge Discodermia calyx. J. Nat. Prod. 2012, 75, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Plaza, A.; Bifulco, G.; Masullo, M.; Lloyd, J.R.; Keffer, J.L.; Colin, P.L.; Hooper, J.N.; Bell, L.J.; Bewley, C.A. Mutremdamide A and koshikamides C-H, peptide inhibitors of HIV-1 entry from different Theonella species. J. Org. Chem. 2010, 75, 4344–4355. [Google Scholar] [CrossRef] [PubMed]
- da Mata, E.C.; Mourao, C.B.; Rangel, M.; Schwartz, E.F. Antiviral activity of animal venom peptides and related compounds. J. Venom Anim. Toxins Incl. Trop. Dis. 2017, 23, 3. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Ma, D. Total synthesis of salinamide A: A potent anti-inflammatory bicyclic depsipeptide. Angew. Chem. Int. Ed. Engl. 2008, 47, 3614–3617. [Google Scholar] [CrossRef] [PubMed]
- Degen, D.; Feng, Y.; Zhang, Y.; Ebright, K.Y.; Ebright, Y.W.; Gigliotti, M.; Vahedian-Movahed, H.; Mandal, S.; Talaue, M.; Connell, N.; et al. Transcription inhibition by the depsipeptide antibiotic salinamide A. Elife 2014, 3, e02451. [Google Scholar] [CrossRef] [PubMed]
- Hassan, H.M.; Degen, D.; Jang, K.H.; Ebright, R.H.; Fenical, W. Salinamide F, new depsipeptide antibiotic and inhibitor of bacterial RNA polymerase from a marine-derived Streptomyces sp. J. Antibiot. (Tokyo) 2015, 68, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Rawat, D.S.; Joshi, M.C.; Joshi, P.; Atheaya, H. Marine peptides and related compounds in clinical trial. Anticancer Agents Med. Chem. 2006, 6, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Yao, G.; Gao, B.M.; Fan, C.X.; Bian, C.; Wang, J.; Cao, Y.; Wen, B.; Zhu, Y.; Ruan, Z.; et al. High-throughput identification of novel conotoxins from the Chinese tubular cone snail (Conus betulinus) by multi-transcriptome sequencing. Gigascience 2016, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.; Jones, A.; Lewis, R.J. Remarkable inter- and intra-species complexity of conotoxins revealed by LC/MS. Peptides 2009, 30, 1222–1227. [Google Scholar] [CrossRef] [PubMed]
- McGivern, J.G. Ziconotide: A review of its pharmacology and use in the treatment of pain. Neuropsychiatr. Dis. Treat. 2007, 3, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, C.I.; Swedberg, J.E.; Craik, D.J. Recent progress towards pharmaceutical applications of disulfide-rich cyclic peptides. Curr. Protein Pept. Sci. 2013, 14, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.J.; Jensen, J.; Nevin, S.T.; Callaghan, B.P.; Adams, D.J.; Craik, D.J. The engineering of an orally active conotoxin for the treatment of neuropathic pain. Angew. Chem. Int. Ed. Engl. 2010, 49, 6545–6548. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J. Conotoxins as selective inhibitors of neuronal ion channels, receptors and transporters. IUBMB Life 2004, 56, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Gowd, K.H.; Twede, V.; Watkins, M.; Krishnan, K.S.; Teichert, R.W.; Bulaj, G.; Olivera, B.M. Conantokin-P, an unusual conantokin with a long disulfide loop. Toxicon 2008, 52, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Mena, E.E.; Gullak, M.F.; Pagnozzi, M.J.; Richter, K.E.; Rivier, J.; Cruz, L.J.; Olivera, B.M. Conantokin-G: A novel peptide antagonist to the N-methyl-d-aspartic acid (NMDA) receptor. Neurosci. Lett. 1990, 118, 241–244. [Google Scholar] [CrossRef]
- Christensen, S.B.; Bandyopadhyay, P.K.; Olivera, B.M.; McIntosh, J.M. Alpha-S-conotoxin GVIIIB potently and selectively blocks alpha9alpha10 nicotinic acetylcholine receptors. Biochem. Pharmacol. 2015, 96, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Menting, J.G.; Gajewiak, J.; MacRaild, C.A.; Chou, D.H.; Disotuar, M.M.; Smith, N.A.; Miller, C.; Erchegyi, J.; Rivier, J.E.; Olivera, B.M.; et al. A minimized human insulin-receptor-binding motif revealed in a Conus geographus venom insulin. Nat. Struct. Mol. Biol. 2016, 23, 916–920. [Google Scholar] [CrossRef] [PubMed]
- Mir, R.; Karim, S.; Kamal, M.A.; Wilson, C.M.; Mirza, Z. Conotoxins: Structure, Therapeutic Potential and Pharmacological Applications. Curr. Pharm. Des. 2016, 22, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Durek, T.; Craik, D.J. Therapeutic conotoxins: A US patent literature survey. Expert Opin. Ther. Pat. 2015, 25, 1159–1173. [Google Scholar] [CrossRef] [PubMed]
- What Is Aplidin? Available online: https://www.themmrf.org/multiple-myeloma-knowledge-center/experimental-treatments/aplidin/ (accessed on 20 April 2017).
- Broggini, M.; Marchini, S.V.; Galliera, E.; Borsotti, P.; Taraboletti, G.; Erba, E.; Sironi, M.; Jimeno, J.; Faircloth, G.T.; Giavazzi, R.; et al. Aplidine, a new anticancer agent of marine origin, inhibits vascular endothelial growth factor (VEGF) secretion and blocks VEGF-VEGFR-1 (flt-1) autocrine loop in human leukemia cells MOLT-4. Leukemia 2003, 17, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Kersten, R.D.; Nam, S.J.; Lu, L.; Al-Suwailem, A.M.; Zheng, H.; Fenical, W.; Dorrestein, P.C.; Moore, B.S.; Qian, P.Y. Bacterial biosynthesis and maturation of the didemnin anti-cancer agents. J. Am. Chem. Soc. 2012, 134, 8625–8632. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.-C.; Kim, D.G.; Han, C.-H.; Lee, Y.J.; Lee, J.-K.; Byun, H.-G.; Lee, S.-C.; Park, S.-J.; Lee, D.-H.; Jeon, Y.-J. Nitric oxide-mediated vasorelaxation effects of anti-angiotensin I-converting enzyme (ACE) peptide from Styela clava flesh tissue and its anti-hypertensive effect in spontaneously hypertensive rats. Food Chem. 2012, 134, 1141–1145. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.C.; Jeon, Y.J. Anti-inflammatory effect of enzymatic hydrolysates from Styela clava flesh tissue in lipopolysaccharide-stimulated RAW 264.7 macrophages and in vivo zebrafish model. Nutr. Res. Pract. 2015, 9, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Humbert, X.; Alexandre, J.; Sassier, M.; Default, A.; Gouraud, A.; Yelehe-Okouma, M.; Puddu, P.E.; Fedrizzi, S. Long delay to onset of ACE inhibitors-induced cough: Reason of difficult diagnosis in primary care? Eur. J. Intern. Med. 2017, 37, e50–e51. [Google Scholar] [CrossRef] [PubMed]
- Vena, G.A.; Cassano, N.; Coco, V.; De Simone, C. Eczematous reactions due to angiotensin-converting enzyme inhibitors or angiotensin II receptor blockers. Immunopharmacol. Immunotoxicol. 2013, 35, 447–450. [Google Scholar] [CrossRef] [PubMed]
- Wijesekara, I.; Qian, Z.-J.; Ryu, B.; Ngo, D.-H.; Kim, S.-K. Purification and identification of antihypertensive peptides from seaweed pipefish (Syngnathus schlegeli) muscle protein hydrolysate. Food Res. Int. 2011, 44, 703–707. [Google Scholar] [CrossRef]
- Pangestuti, R.; Kim, S.-K. Peptide-derived from Seahorse Exerts a Protective Effect against Cholinergic Neuronal Death in in vitro Model of Alzheimer’s Disease. Procedia Chem. 2015, 14, 343–352. [Google Scholar] [CrossRef]
- Tamamura, H.; Xu, Y.; Hattori, T.; Zhang, X.; Arakaki, R.; Kanbara, K.; Omagari, A.; Otaka, A.; Ibuka, T.; Yamamoto, N.; et al. A low-molecular-weight inhibitor against the chemokine receptor CXCR4: A strong anti-HIV peptide T140. Biochem. Biophys. Res. Commun. 1998, 253, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Tziveleka, L.A.; Vagias, C.; Roussis, V. Natural products with anti-HIV activity from marine organisms. Curr. Top. Med. Chem. 2003, 3, 1512–1535. [Google Scholar] [CrossRef] [PubMed]
- An, M.Y.; Gao, J.; Zhao, X.F.; Wang, J.X. A new subfamily of penaeidin with an additional serine-rich region from kuruma shrimp (Marsupenaeus japonicus) contributes to antimicrobial and phagocytic activities. Dev. Comp. Immunol. 2016, 59, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Afsal, V.V.; Antony, S.P.; Philip, R.; Bright Singh, I.S. Molecular Characterization of a Newly Identified Subfamily Member of Penaeidin from two Penaeid Shrimps, Fenneropenaeus indicus and Metapenaeus monoceros. Probiotics Antimicrob. Proteins 2016, 8, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Destoumieux, D.; Bulet, P.; Loew, D.; Van Dorsselaer, A.; Rodriguez, J.; Bachere, E. Penaeidins, a new family of antimicrobial peptides isolated from the shrimp Penaeus vannamei (Decapoda). J. Biol. Chem. 1997, 272, 28398–28406. [Google Scholar] [CrossRef] [PubMed]
- Destoumieux, D.; Munoz, M.; Cosseau, C.; Rodriguez, J.; Bulet, P.; Comps, M.; Bachere, E. Penaeidins, antimicrobial peptides with chitin-binding activity, are produced and stored in shrimp granulocytes and released after microbial challenge. J. Cell Sci. 2000, 113, 461–469. [Google Scholar] [PubMed]
- Petit, V.W.; Rolland, J.L.; Blond, A.; Cazevieille, C.; Djediat, C.; Peduzzi, J.; Goulard, C.; Bachere, E.; Dupont, J.; Destoumieux-Garzon, D.; Rebuffat, S. A hemocyanin-derived antimicrobial peptide from the penaeid shrimp adopts an alpha-helical structure that specifically permeabilizes fungal membranes. Biochim. Biophys. Acta 2016, 1860, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Poirier, L.; Quiniou, F.; Ruiz, N.; Montagu, M.; Amiard, J.C.; Pouchus, Y.F. Toxicity assessment of peptaibols and contaminated sediments on Crassostrea gigas embryos. Aquat. Toxicol. 2007, 83, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, N.; Wielgosz-Collin, G.; Poirier, L.; Grovel, O.; Petit, K.E.; Mohamed-Benkada, M.; du Pont, T.R.; Bissett, J.; Verite, P.; Barnathan, G.; et al. New Trichobrachins, 11-residue peptaibols from a marine strain of Trichoderma longibrachiatum. Peptides 2007, 28, 1351–1358. [Google Scholar] [CrossRef] [PubMed]
- Su, H.-N.; Chen, Z.-H.; Song, X.-Y.; Chen, X.-L.; Shi, M.; Zhou, B.-C.; Zhao, X.; Zhang, Y.-Z. Antimicrobial Peptide Trichokonin VI-Induced Alterations in the Morphological and Nanomechanical Properties of Bacillus subtilis. PLoS ONE 2012, 7, e45818. [Google Scholar] [CrossRef] [PubMed]
- Du, F.Y.; Zhang, P.; Li, X.M.; Li, C.S.; Cui, C.M.; Wang, B.G. Cyclohexadepsipeptides of the isaridin class from the marine-derived fungus Beauveria felina EN-135. J. Nat. Prod. 2014, 77, 1164–1169. [Google Scholar] [CrossRef] [PubMed]
- Raju, R.; Khalil, Z.G.; Piggott, A.M.; Blumenthal, A.; Gardiner, D.L.; Skinner-Adams, T.S.; Capon, R.J. Mollemycin A: An antimalarial and antibacterial glyco-hexadepsipeptide-polyketide from an Australian marine-derived Streptomyces sp. (CMB-M0244). Org. Lett. 2014, 16, 1716–1719. [Google Scholar] [CrossRef] [PubMed]
- Wyche, T.P.; Hou, Y.; Vazquez-Rivera, E.; Braun, D.; Bugni, T.S. Peptidolipins B-F, antibacterial lipopeptides from an ascidian-derived Nocardia sp. J. Nat. Prod. 2012, 75, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Marine natural products and related compounds in clinical and advanced preclinical trials. J. Nat. Prod. 2004, 67, 1216–1238. [Google Scholar] [CrossRef] [PubMed]
- Tu, A.T. Sea snake venoms and neurotoxins. J. Agric. Food Chem. 1974, 22, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Marr, A.K.; Gooderham, W.J.; Hancock, R.E. Antibacterial peptides for therapeutic use: Obstacles and realistic outlook. Curr. Opin. Pharmacol. 2006, 6, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, A.; Pirri, G.; Nicoletto, S.F. Antimicrobial peptides: An overview of a promising class of therapeutics. Cent. Eur. J. Biol. 2007, 2, 1–33. [Google Scholar] [CrossRef]
- Mygind, P.H.; Fischer, R.L.; Schnorr, K.M.; Hansen, M.T.; Sonksen, C.P.; Ludvigsen, S.; Raventos, D.; Buskov, S.; Christensen, B.; De Maria, L.; et al. Plectasin is a peptide antibiotic with therapeutic potential from a saprophytic fungus. Nature 2005, 437, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Opar, A. ‘Pharmers’ hope for first plant drug harvest. Nat. Rev. Drug Discov. 2011, 10, 81–82. [Google Scholar] [CrossRef] [PubMed]
- Dornenburg, H. Progress in kalata peptide production via plant cell bioprocessing. Biotechnol. J. 2009, 4, 632–645. [Google Scholar] [CrossRef] [PubMed]
- Company, N.; Nadal, A.; Ruiz, C.; Pla, M. Production of phytotoxic cationic alpha-helical antimicrobial peptides in plant cells using inducible promoters. PLoS ONE 2014, 9, e109990. [Google Scholar] [CrossRef] [PubMed]
- Santos, G.B.; Ganesan, A.; Emery, F.S. Oral Administration of Peptide-Based Drugs: Beyond Lipinski’s Rule. ChemMedChem 2016, 11, 2245–2251. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.F.; Peng, H.B.; Liu, G.Q.; Zhang, F.; Li, Y. Beneficial effects of oligopeptides from marine salmon skin in a rat model of type 2 diabetes. Nutrition 2010, 26, 1014–1020. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.F.; Li, G.Z.; Peng, H.B.; Zhang, F.; Chen, Y.; Li, Y. Treatment with marine collagen peptides modulates glucose and lipid metabolism in Chinese patients with type 2 diabetes mellitus. Appl. Physiol. Nutr. Metab. 2010, 35, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Wyche, T.P.; Dammalapati, A.; Cho, H.; Harrison, A.D.; Kwon, G.S.; Chen, H.; Bugni, T.S.; Jaskula-Sztul, R. Thiocoraline activates the Notch pathway in carcinoids and reduces tumor progression in vivo. Cancer Gene. Ther. 2014, 21, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Tulla-Puche, J.; Auriemma, S.; Falciani, C.; Albericio, F. Orthogonal chemistry for the synthesis of thiocoraline-triostin hybrids. Exploring their structure-activity relationship. J. Med. Chem. 2013, 56, 5587–5600. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.Y.; Liu, Y.; Luesch, H. Systematic Chemical Mutagenesis Identifies a Potent Novel Apratoxin A/E Hybrid with Improved in Vivo Antitumor Activity. ACS Med. Chem. Lett. 2011, 2, 861–865. [Google Scholar] [CrossRef] [PubMed]
- Salvador, L.A.; Park, H.; Al-Awadhi, F.H.; Liu, Y.; Kim, B.; Zeller, S.L.; Chen, Q.Y.; Hong, J.; Luesch, H. Modulation of Activity Profiles for Largazole-Based HDAC Inhibitors through Alteration of Prodrug Properties. ACS Med. Chem. Lett. 2014, 5, 905–910. [Google Scholar] [CrossRef] [PubMed]
- Schweikart, K.; Guo, L.; Shuler, Z.; Abrams, R.; Chiao, E.T.; Kolaja, K.L.; Davis, M. The effects of jaspamide on human cardiomyocyte function and cardiac ion channel activity. Toxicol. In Vitro 2013, 27, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Randazzo, A.; Bifulco, G.; Giannini, C.; Bucci, M.; Debitus, C.; Cirino, G.; Gomez-Paloma, L. Halipeptins A and B: Two novel potent anti-inflammatory cyclic depsipeptides from the Vanuatu marine sponge Haliclona species. J. Am. Chem. Soc. 2001, 123, 10870–10876. [Google Scholar] [CrossRef] [PubMed]
- Pope, J.E.; Deer, T.R. Ziconotide: A clinical update and pharmacologic review. Expert Opin Pharmacother. 2013, 14, 957–966. [Google Scholar] [CrossRef] [PubMed]
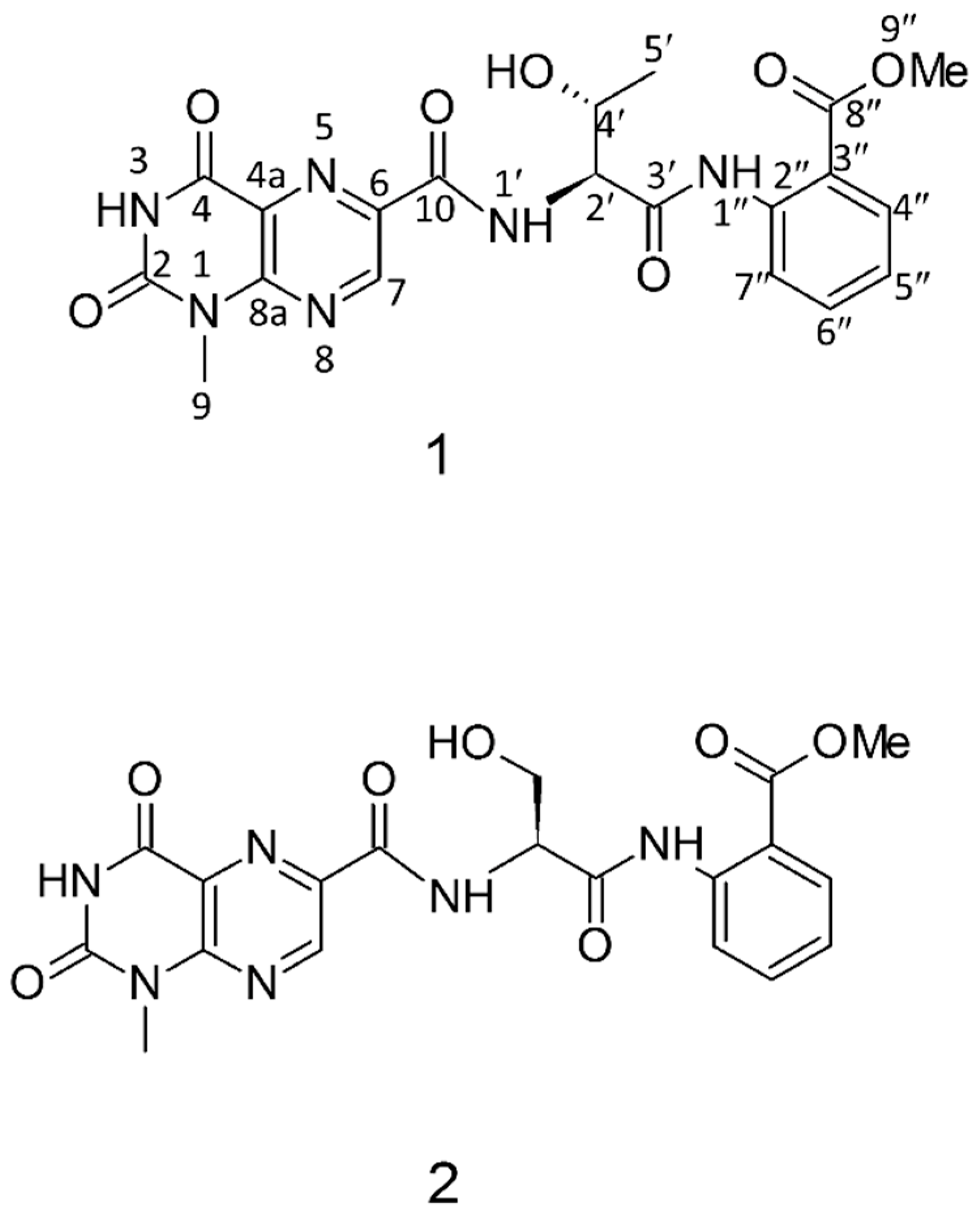
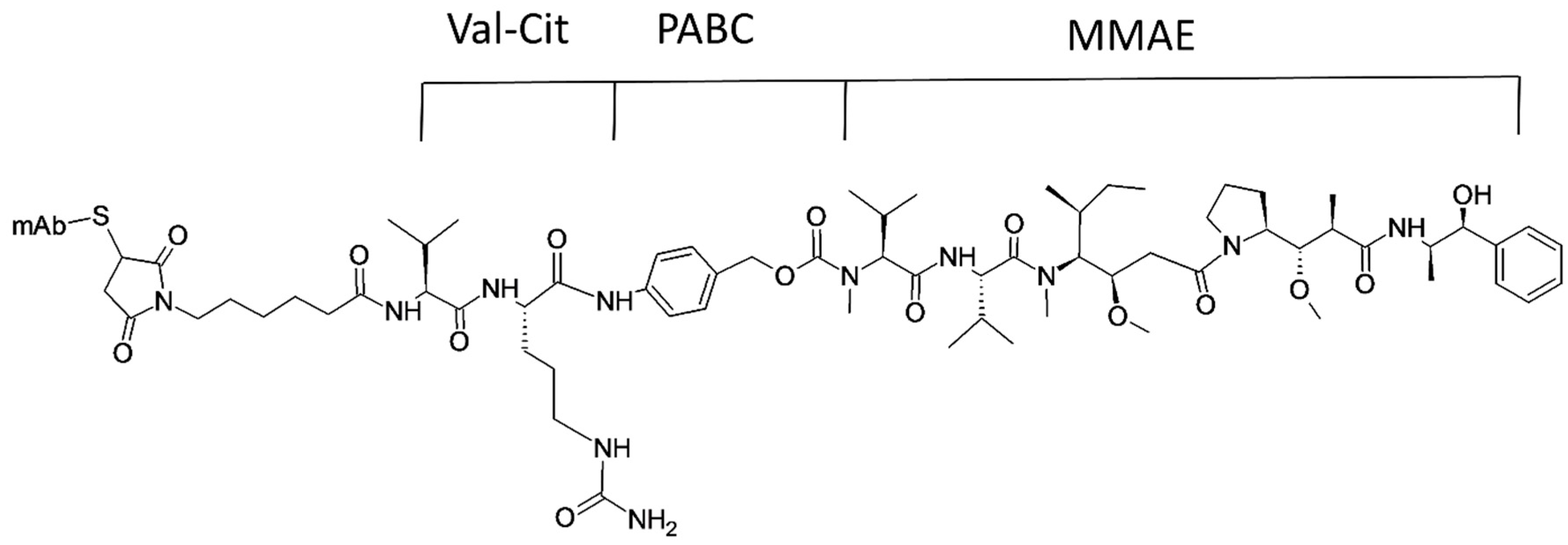
- Katz, J.; Janik, J.E.; Younes, A. Brentuximab Vedotin (SGN-35). Clin. Cancer Res. 2011, 17, 6428. [Google Scholar] [CrossRef] [PubMed]
- Martin-Algarra, S.; Espinosa, E.; Rubio, J.; Lopez, J.J.; Manzano, J.L.; Carrion, L.A.; Plazaola, A.; Tanovic, A.; Paz-Ares, L. Phase II study of weekly Kahalalide F in patients with advanced malignant melanoma. Eur. J. Cancer 2009, 45, 732–735. [Google Scholar] [CrossRef] [PubMed]
- Ratain, M.J.; Geary, D.; Undevia, S.D.; Coronado, C.; Alfaro, V.; Iglesias, J.L.; Schilsky, R.L.; Miguel-Lillo, B. First-in-human, phase I study of elisidepsin (PM02734) administered as a 30-min or as a 3-hour intravenous infusion every three weeks in patients with advanced solid tumors. Investig. New Drugs 2015, 33, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.; Glaser, K.B.; Cuevas, C.; Jacobs, R.S.; Kem, W.; Little, R.D.; McIntosh, J.M.; Newman, D.J.; Potts, B.C.; Shuster, D.E. The odyssey of marine pharmaceuticals: A current pipeline perspective. Trends Pharmacol. Sci. 2010, 31, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Vaklavas, C.; Forero, A. Management of metastatic breast cancer with second-generation antibody-drug conjugates: Focus on glembatumumab vedotin (CDX-011, CR011-vcMMAE). BioDrugs 2014, 28, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Keohan, M.L.; Saif, M.W.; Rushing, D.; Baez, L.; Feit, K.; DeJager, R.; Anderson, S. Phase II study of intravenous TZT-1027 in patients with advanced or metastatic soft-tissue sarcomas with prior exposure to anthracycline-based chemotherapy. Cancer 2006, 107, 2881–2887. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Lima, C.M.; Bayraktar, S.; Macintyre, J.; Raez, L.; Flores, A.M.; Ferrell, A.; Rubin, E.H.; Poplin, E.A.; Tan, A.R.; Lucarelli, A.; et al. A phase 1 trial of E7974 administered on day 1 of a 21-day cycle in patients with advanced solid tumors. Cancer 2012, 118, 4262–4270. [Google Scholar] [CrossRef] [PubMed]
- Hadaschik, B.A.; Ettinger, S.; Sowery, R.D.; Zoubeidi, A.; Andersen, R.J.; Roberge, M.; Gleave, M.E. Targeting prostate cancer with HTI-286, a synthetic analog of the marine sponge product hemiasterlin. Int. J. Cancer 2008, 122, 2368–2376. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Marine-sourced anti-cancer and cancer pain control agents in clinical and late preclinical development. Mar. Drugs 2014, 12, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Okkerse, P.; Hay, J.L.; Sitsen, E.; Dahan, A.; Klaassen, E.; Houghton, W.; Groeneveld, G.J. Pharmacokinetics and pharmacodynamics of intrathecally administered Xen2174, a synthetic conopeptide with norepinephrine reuptake inhibitor and analgesic properties. Br. J. Clin. Pharmacol. 2017, 83, 751–763. [Google Scholar] [CrossRef] [PubMed]
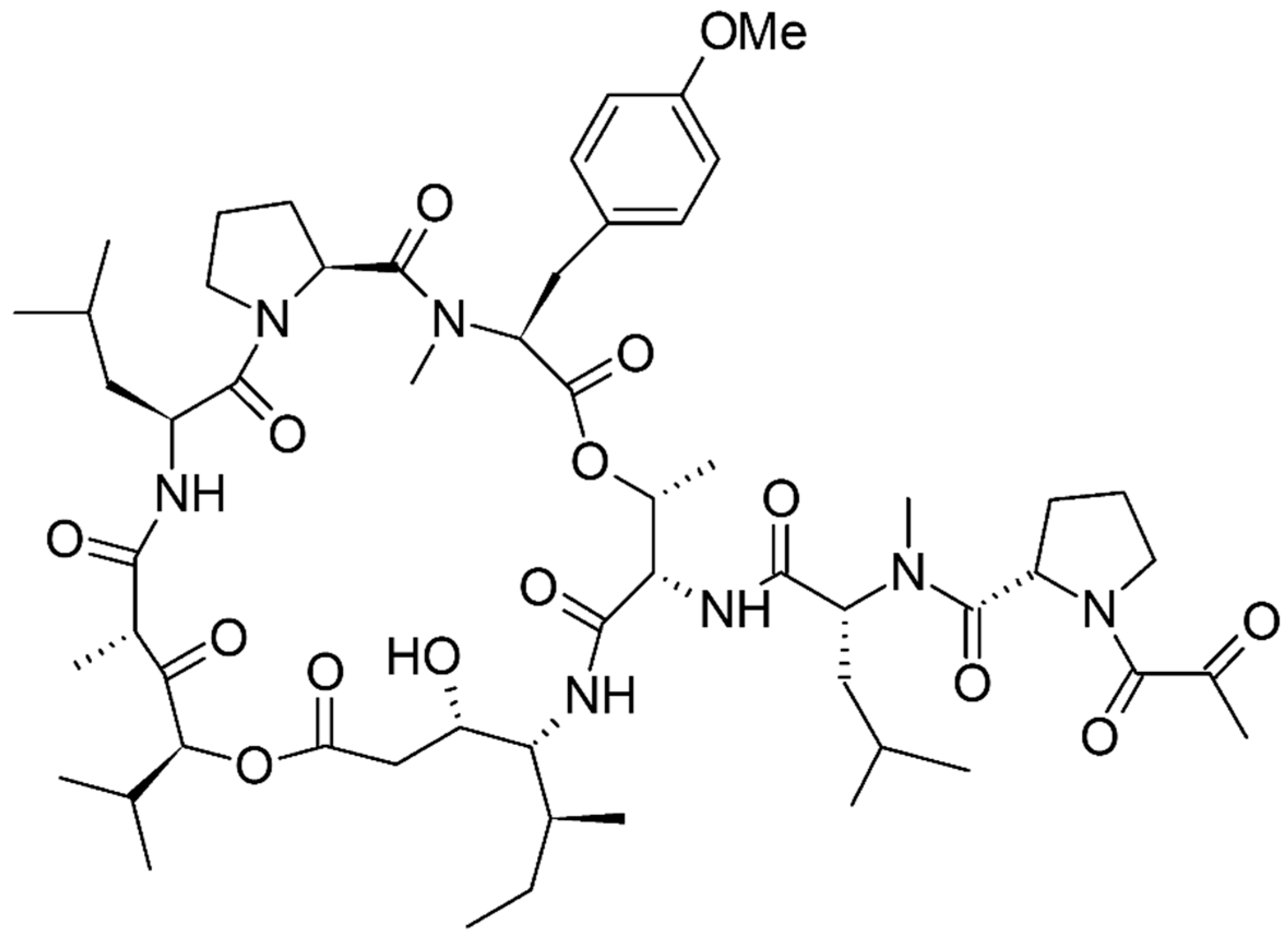
- Ribrag, V.; Caballero, D.; Ferme, C.; Zucca, E.; Arranz, R.; Briones, J.; Gisselbrecht, C.; Salles, G.; Gianni, A.M.; Gomez, H.; et al. Multicenter phase II study of plitidepsin in patients with relapsed/refractory non-Hodgkin’s lymphoma. Haematologica 2013, 98, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Van Andel, L.; Fudio, S.; Rosing, H.; Munt, S.; Miguel-Lillo, B.; González, I.; Tibben, M.; de Vries, N.; de Vries Schultink, A.; Schellens, J. Pharmacokinetics and excretion of 14C–Plitidepsin in patients with advanced cancer. Investig. New Drugs 2017, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Perez, E.A.; Hillman, D.W.; Fishkin, P.A.; Krook, J.E.; Tan, W.W.; Kuriakose, P.A.; Alberts, S.R.; Dakhil, S.R. Phase II trial of dolastatin-10 in patients with advanced breast cancer. Investig. New Drugs 2005, 23, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Supko, J.G.; Lynch, T.J.; Clark, J.W.; Fram, R.; Allen, L.F.; Velagapudi, R.; Kufe, D.W.; Eder, J.P., Jr. A phase I clinical and pharmacokinetic study of the dolastatin analogue cemadotin administered as a 5-day continuous intravenous infusion. Cancer Chemother. Pharmacol. 2000, 46, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Lago, J.; Rodriguez, L.P.; Blanco, L.; Vieites, J.M.; Cabado, A.G. Tetrodotoxin, an Extremely Potent Marine Neurotoxin: Distribution, Toxicity, Origin and Therapeutical Uses. Mar. Drugs 2015, 13, 6384–6406. [Google Scholar] [CrossRef] [PubMed]
- Molina-Guijarro, J.M.; Macias, A.; Garcia, C.; Munoz, E.; Garcia-Fernandez, L.F.; David, M.; Nunez, L.; Martinez-Leal, J.F.; Moneo, V.; Cuevas, C.; et al. Irvalec inserts into the plasma membrane causing rapid loss of integrity and necrotic cell death in tumor cells. PLoS ONE 2011, 6, e19042. [Google Scholar] [CrossRef] [PubMed]
- Molina-Guijarro, J.M.; Garcia, C.; Macias, A.; Garcia-Fernandez, L.F.; Moreno, C.; Reyes, F.; Martinez-Leal, J.F.; Fernandez, R.; Martinez, V.; Valenzuela, C.; et al. Elisidepsin Interacts Directly with Glycosylceramides in the Plasma Membrane of Tumor Cells to Induce Necrotic Cell Death. PLoS ONE 2015, 10, e0140782. [Google Scholar]
- Cruz, L.J.; Luque-Ortega, J.R.; Rivas, L.; Albericio, F. Kahalalide F, an antitumor depsipeptide in clinical trials, and its analogues as effective antileishmanial agents. Mol. Pharm. 2009, 6, 813–824. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Number of Amino Acids/Unnatural Amino Acids/Molecular Weight | Marine Source | Possible Application | Reference |
|---|---|---|---|---|
| Porifera | ||||
| Cyclic depsipeptides Mirabamides | 3 N, 8 UNA | Sponge Stelletta clavosa | Anti-HIV and antibacterial | [63,64] |
| Papuamides A–F | 11 UNA | Sponges Theonella mirabilis and Theonella swinhoei | Anti-HIV and cytotoxic | [65,66] |
| Celebesides A–C and | 5 UNA | Sponge Siliquariaspongia mirabilis | Anti-HIV, cytotoxic, and antifungal | [67] |
| Theopapuamides B–D | 2 N, 9 UNA | Sponge Siliquariaspongia mirabilis | Anti-HIV, cytotoxic, and antifungal | [67] |
| Callipeltins | 5 N, 5 UNA | Sponge Callipelta sp. | Anti-HIV | [68,69] |
| Callyaerins A–G, I–M | 9-12 N | Sponge Callyspongia aerizusa | Antimicrobial, antitubercular and antiviral | [70] |
| Discodermin A | 11 N, 3UNA | Sponge Discodermia kiiensis, | Broad spectrum antifungal and antibacterial, inhibited starfish embryo development | [71] |
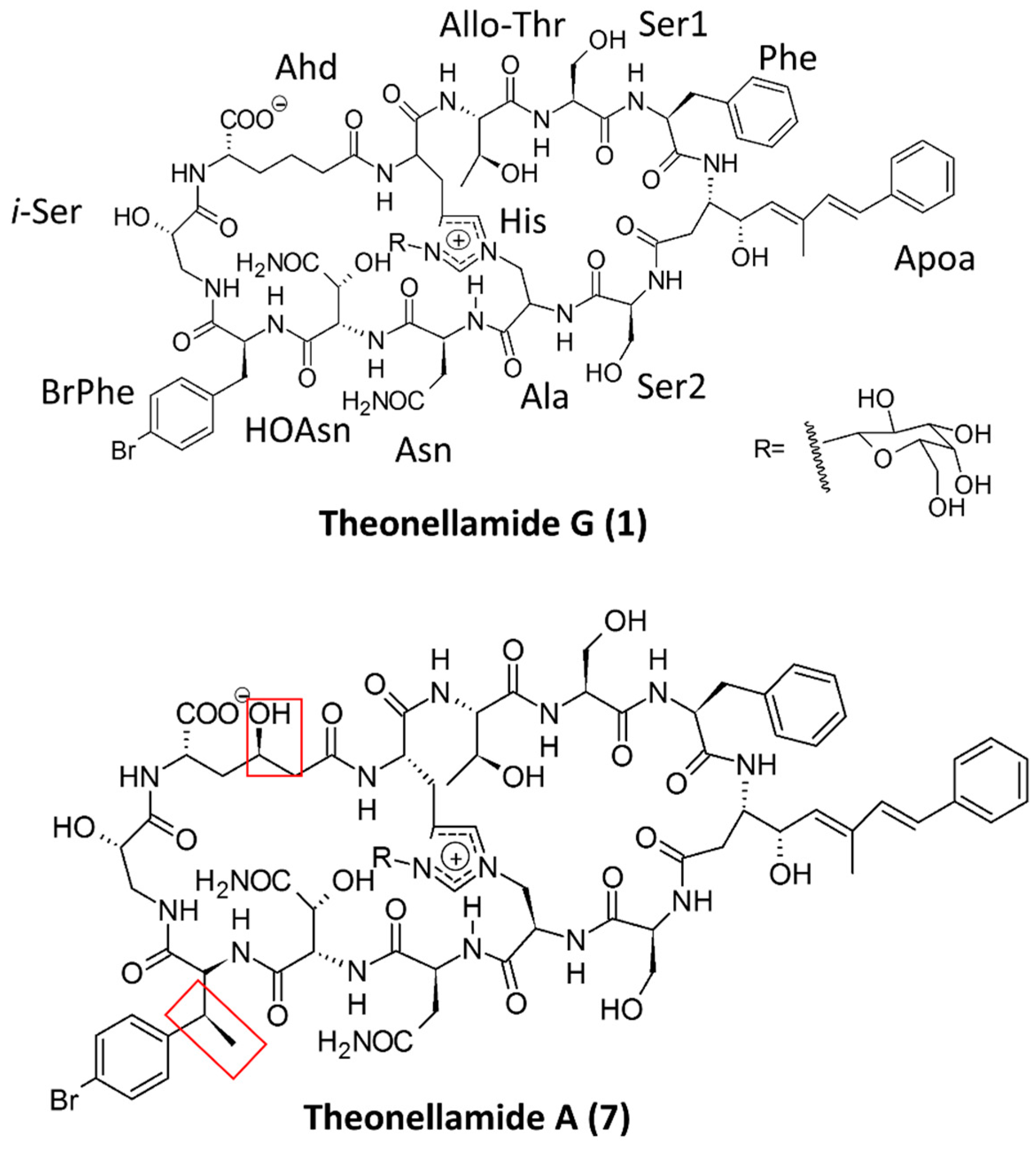
| Theonellamide G | 7 N, 5 UNA | Red Sea sponge Theonella swinhoei | Antifungal and cytotoxic | [72] |
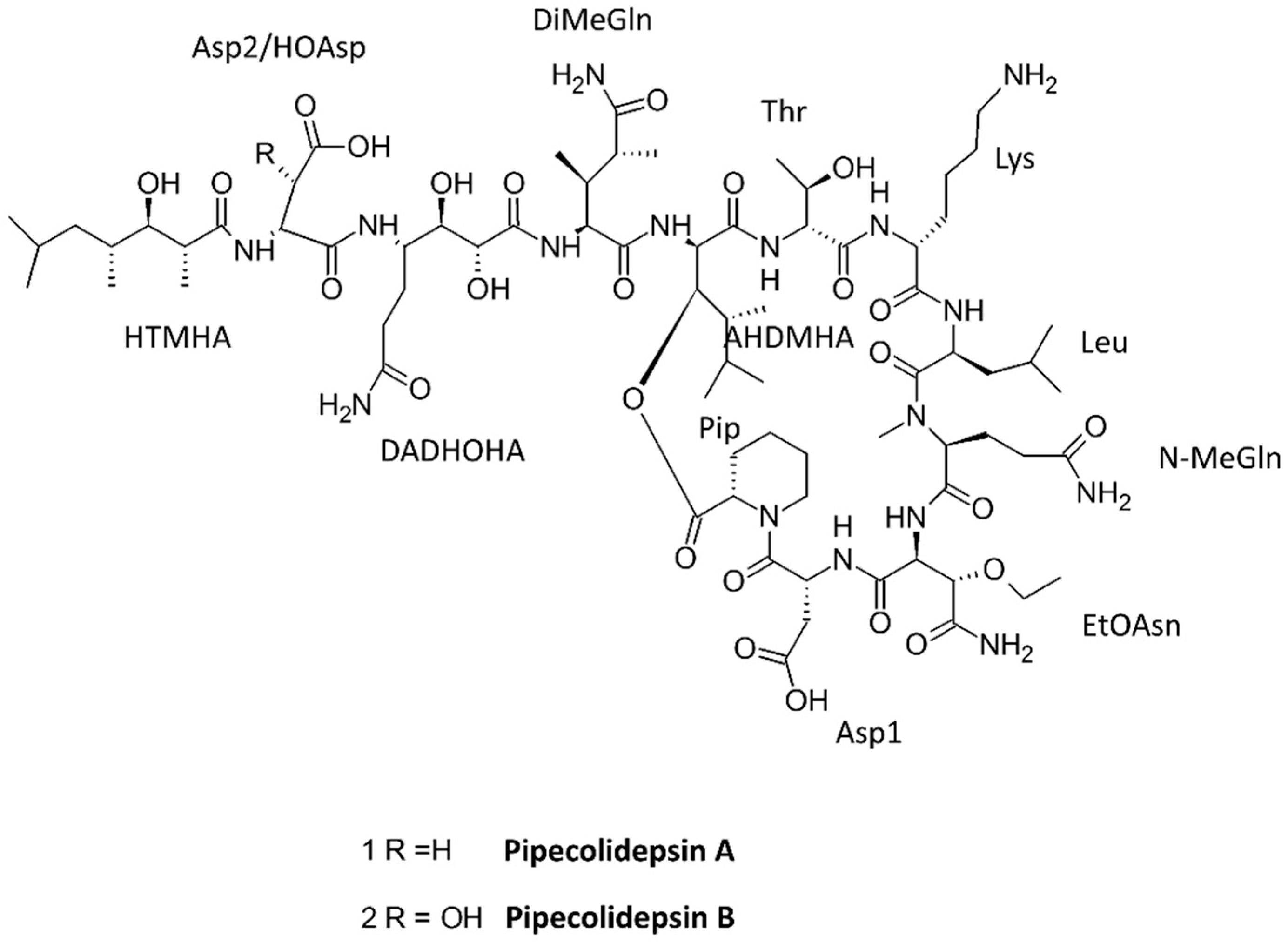
| Pipecolidepsins A and B | 3 N, 8 UNA | Sponge Homophymia lamellose | Cytotoxic | [73] |
| Theonellamide A–E, G | 7 N, 5 UNA | Sponge Theonella sp. | Cytotoxic and antifungal | [74,75] |
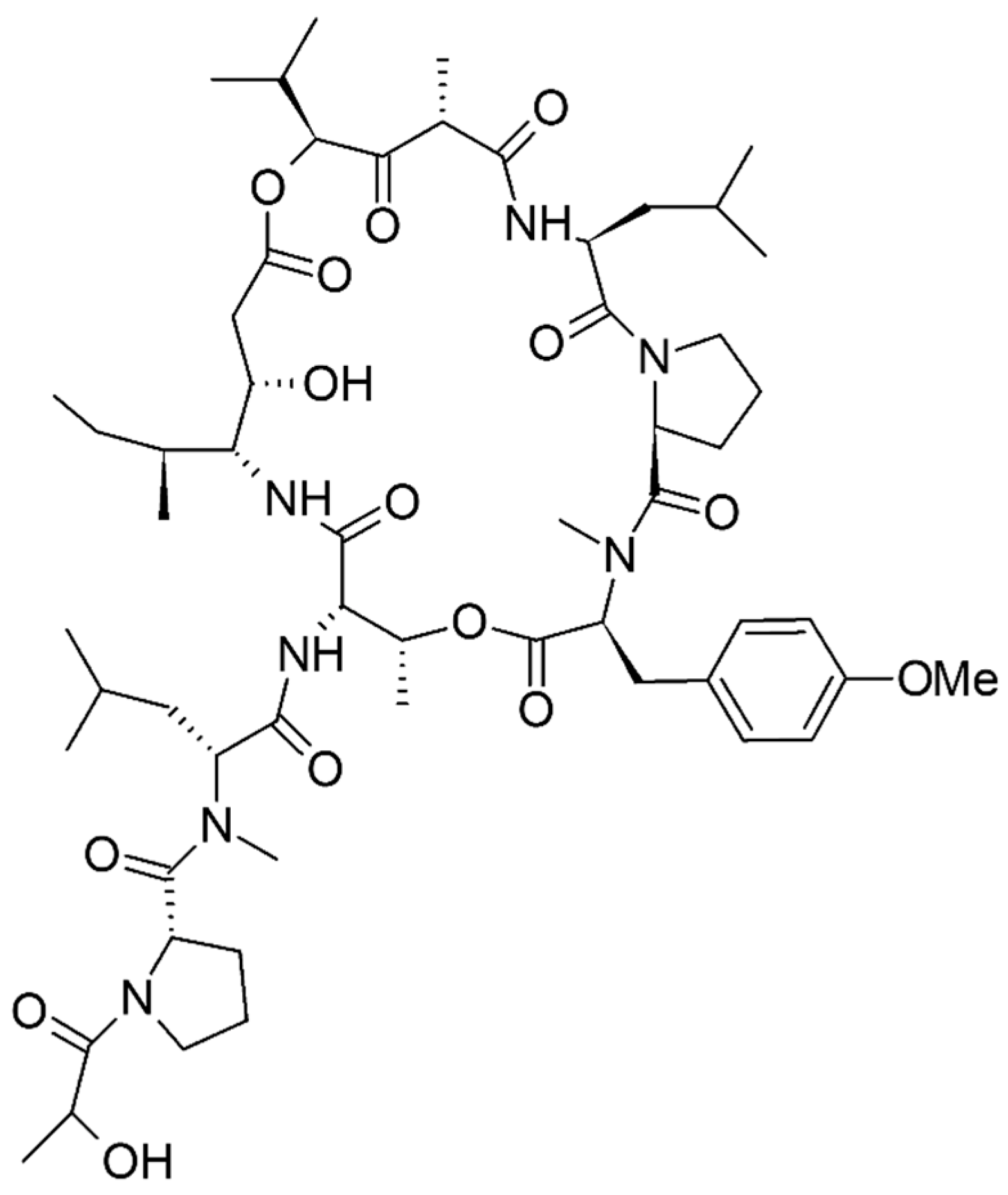
| Milnamide A | 3 UNA | Sponge Pipestela candelabra | Antiproliferative and antitumor | [76] |
| Barrettides A and B | 31 N | Sponge Geodia barrette | Antifouling effect with barnacle larvae | [77] |
| Koshikamides F–H | 4N, 13 UNA | Sponges Theonella swinhoei and Theonella cupola | Anti-HIV | [78] |
| Cnidaria | ||||
| Toxin of Palythoa caribaeorum | 1800–9000 Da peptides | Zoanthid Palythoa caribaeorum | Neuronal function modulation | [79] |
| Jellyfish collagen peptides (JCP) | 200–600 Da | Jellyfish Rhopilema esculentum | Antihypertensive | [80] |
| Peptide Ala-Cys-Pro-Gly-Pro-Asn-Pro-Gly-Arg-Pro | 10 N | Box jellyfish Chiropsalmus quadrigatus | Antihypertensive | [81] |
| Neurotoxin AV3 | 27 N | Sea anemone Anemonia viridis | Modulation of voltage gated sodium channel | [82] |
| Molluska | ||||
| Kahalalide F Analogs | 8 N, 5 UNA | Mollusks Sacoglossan sp. | Antitumor and antifungal | [83] |
| Cm-p1 and Cm-p5 | 10 N | Marine snail Cenchritis muricatus | Antifungal | [84,85] |
| Crude peptide extract | Cone snail Conus araneosus | Sleep inductive | [86] | |
| Hemocyanin 1 and 2 | 38 and 24 N | Sea snail Rapana thomasiana | Antiviral against herpes simplex virus (HSV) | [59] |
| Neurotensin (NT) and Contulakin-G | 13 N in NT, and 15 N and 1 UNA in contulakin-G | Cone snail Conus geographus | Analgesic | [87] |
| Chordata | ||||
| Peptide (Ala-His-Ile-Ile-Ile, MW: 565.3 Da) | 5 N | Tunicate Styela clava | Antihypertensive, antioxidant, cytotoxic, hepatoprotective | [88] |
| PC-1, PC-2 and PC-3 | 5 N | Large yellow croaker Pseudosciaena crocea | Antioxidant | [89] |
| HTP-1 | 8 N | Seahorse Hippocampus trimaculatus | Neuroprotective | [90] |
| EcDefensin | 63 N | Estuary cod Epinephelus coioides | Antiviral against Singapore grouper iridovirus and viral nervous necrosis virus | [91] |
| Trunkamide A | 3 N and 4 UNA | Ascidian Lissoclinum patella | Antitumor | [92] |
| SP-A and SP-B | 9 N | Skate Raja porosa | Antioxidant | [93] |
| Hydrolysate fractions FrA3 and FrB2 | 585.91 and 528.78 Da peptides | Skipjack tuna Katsuwonus pelamis | Antioxidant | [94] |
| Echinodermata | ||||
| Plancitoxin I | 358 N | Spine of crown of thorns starfish, Acanthaster plansi | Cytotoxic and apoptotic | [95] |
| Centrocins 1 and 2 | 119 and 118 N | Green sea urchin Strongylocentrotus droebachiensis | Broad spectrum antibacterial | [96] |
| Arthopoda | ||||
| SpHyastatin | 135 N | Mud crab Scylla paramamosain | Antibacterial | [97] |
| Scygonadin | 102 N | Mud crab Scylla paramamosain | Antibacterial and immune booster, antiviral against white spot syndrome virus | [59] |
| Histone H2A derivative sphistin | 38 N | Mud crab Scylla paramamosain | Antimicrobial | [98] |
| Shrimp anti-lipopolysaccharide factor (SALF) | 24 N | Shrimp Penaeus monodon | Increased expression of immune cells in mice, can be used as adjuvant in cancer vaccine, antibacterial | [99] |
| Ls-Stylicin1 | 82 N | Shrimp Litopenaeus stylirostris | Antibacterial, antifungal | [100] |
| Algae, fungi and bacteria | ||||
| Cyclic lipopeptides, Maribasins A and B | 7 N and 1 UNA | Fermentation broth of the marine microorganisms Bacillus marinus B-9987 | Broad spectrum antifungal | [101] |
| JBIR-34 and JBIR-35 | 4 UNA | Sponge derived actinomycete Streptomyces sp. | Antibacterial, cytotoxic | [102] |
| Mojavensin A, iso-C16 fengycin B, and anteiso-C17 fengycin B | 7 N and 1 UNA | Bacterium Bacillus mojavensis B0621A | Cytotoxic, antifungal | [103] |
| Thalassospiramides A, D and G | (1, 4 and 2 NA) and (6, 3 and 5 UNA) | Bacteria Thalassospira strain CNJ328 | Anti-inflammatory | [104] |
| Hormaomycins B and C | 2 N and 6 UNA | Mudflat-derived Streptomyces sp. | Antibacterial | [105] |
| Aminolipopeptides Trichoderins A, A1 and B | 2 N and 6 UNA | Marine sponge-derived fungus of Trichoderma sp. | Antimycobacterial activity against Mycobacterium smegmatis, Mycobacterium bovis, and Mycobacterium tuberculosis | [106] |
| Terrelumamides A and B | 1 N | Marine fungi Aspergillus terreus | Antidiabetic (increased insulin sensitivity) | [107] |
| PPY1 | 5 N | Algae Pyropia yezoensis | Anti-inflammatory | [108] |
| Major Source | Proteolytic Enzyme | pH | Temperature (°C) |
|---|---|---|---|
| Animals | Trypsin | 8.0 | 37 |
| Pepsin | 2.0 | 37 | |
| α-Chymotrypsin | 8.0 | 37 | |
| Plants | Papain | 6.5 | 50 |
| Microorganisms | Alcalase | 9.5 | 50 |
| Neutrase | 7.0 | 60 | |
| Flavourzyme | 7.0 | 55 | |
| Protamex | 6.0 | 40 | |
| Kojizyme | 6.0 | 40 |
| Marine Peptides in Clinical Trial and on the Market | Marine Source | Therapeutic Application | Reference |
|---|---|---|---|
| Ziconotide | Marine cone snail Conus magus | Analgesic drug (FDA-approved) | [189] |
| Brentuximab Vedotin | Marine mollusk Dolabella auricularia Cyanobacteria Symploca sp. (Antibody-peptide conjugate) | Anticancer (FDA-approved) | [190] |
| Kahalalide F | Mollusk Elysia rufescens, and its diet green algae Bryopsis sp. | Anticancer (phase I clinical study) | [191] |
| Elisidepsin (PM02734) | Synthetic analogue of kahalalide F | Malignant solid tumors (phase I clinical study) | [192] |
| Tasidotin (ILX-651) | Dolastatin 15 synthetic analogues, marine mollusk Dolabella auricularia | Solid tumors, microtubule assembly, lung cancer; phase III clinical study, trials under review | [193] |
| Glembatumumab Vedotin | Derivative of dolastatin 10, marine mollusk Dolabella auricularia (Antibody-peptide conjugate) | Phase I/II for treatment of breast cancer | [194] |
| Soblidotin (TZT-1027) | Derivative of dolastatin 10, marine mollusk Dolabella auricularia | Phase II clinical trial for soft tissue sarcoma | [195] |
| E7974 | Derivative of hemiasterlin, sponge Hemiasterella minor | Phase I study of colorectal, prostate, and larynx carcinomas; recommended for phase II | [196] |
| HTI-286 | Derivative of hemiasterlin, sponge Hemiasterella minor | Metastatic prostate cancer (preclinical study) | [137,197] |
| XEN-2174 | Derivative of ziconotide, marine cone snail Conus magus | Analgesic (phase I-II open label study) | [198,199] |
| Plitidepsin | Ascidian Aplidium albicans | Anticancer (phase I/II clinical study) | [200,201] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sable, R.; Parajuli, P.; Jois, S. Peptides, Peptidomimetics, and Polypeptides from Marine Sources: A Wealth of Natural Sources for Pharmaceutical Applications. Mar. Drugs 2017, 15, 124. https://doi.org/10.3390/md15040124
Sable R, Parajuli P, Jois S. Peptides, Peptidomimetics, and Polypeptides from Marine Sources: A Wealth of Natural Sources for Pharmaceutical Applications. Marine Drugs. 2017; 15(4):124. https://doi.org/10.3390/md15040124
Chicago/Turabian StyleSable, Rushikesh, Pravin Parajuli, and Seetharama Jois. 2017. "Peptides, Peptidomimetics, and Polypeptides from Marine Sources: A Wealth of Natural Sources for Pharmaceutical Applications" Marine Drugs 15, no. 4: 124. https://doi.org/10.3390/md15040124
APA StyleSable, R., Parajuli, P., & Jois, S. (2017). Peptides, Peptidomimetics, and Polypeptides from Marine Sources: A Wealth of Natural Sources for Pharmaceutical Applications. Marine Drugs, 15(4), 124. https://doi.org/10.3390/md15040124