α-Glucosidase and Protein Tyrosine Phosphatase 1B Inhibitory Activity of Plastoquinones from Marine Brown Alga Sargassum serratifolium

 ,
, 
Abstract
1. Introduction
2. Results
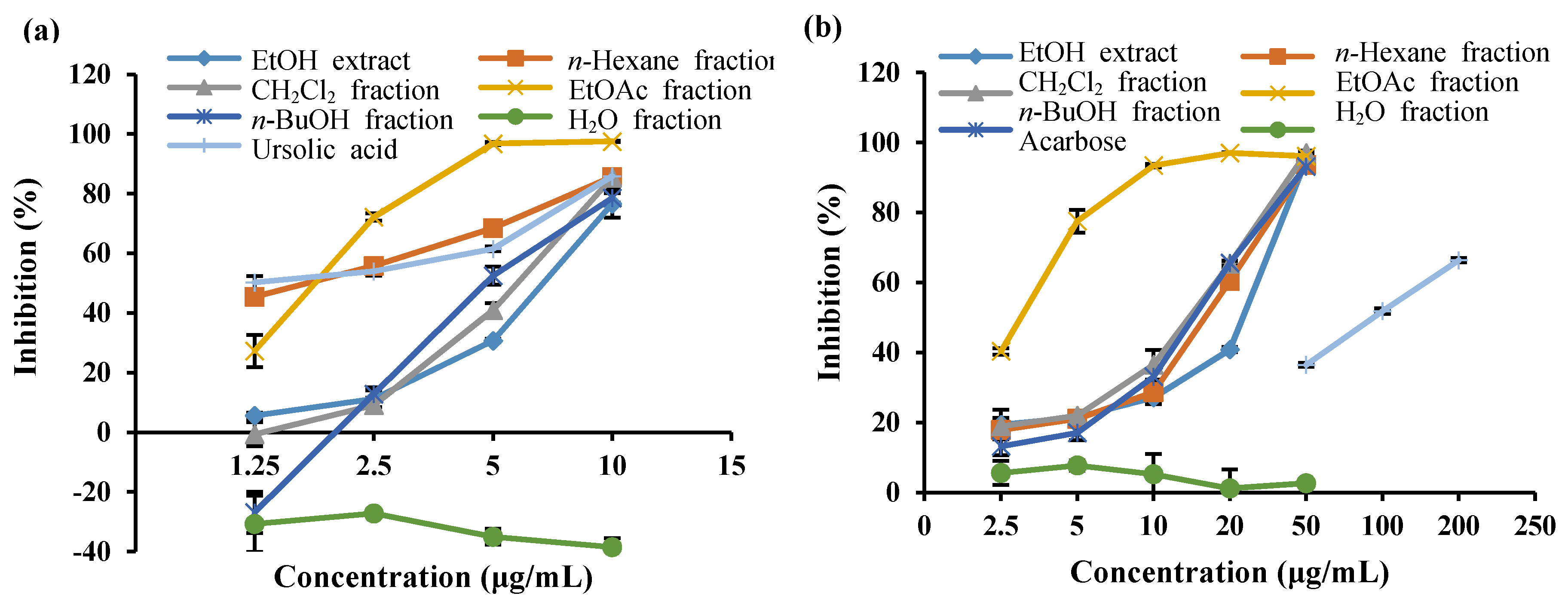
2.1. PTP1B and α-Glucosidase Inhibitory Activity of the EtOH Extract and the Solvent-Soluble Fractions of S. serratifolium
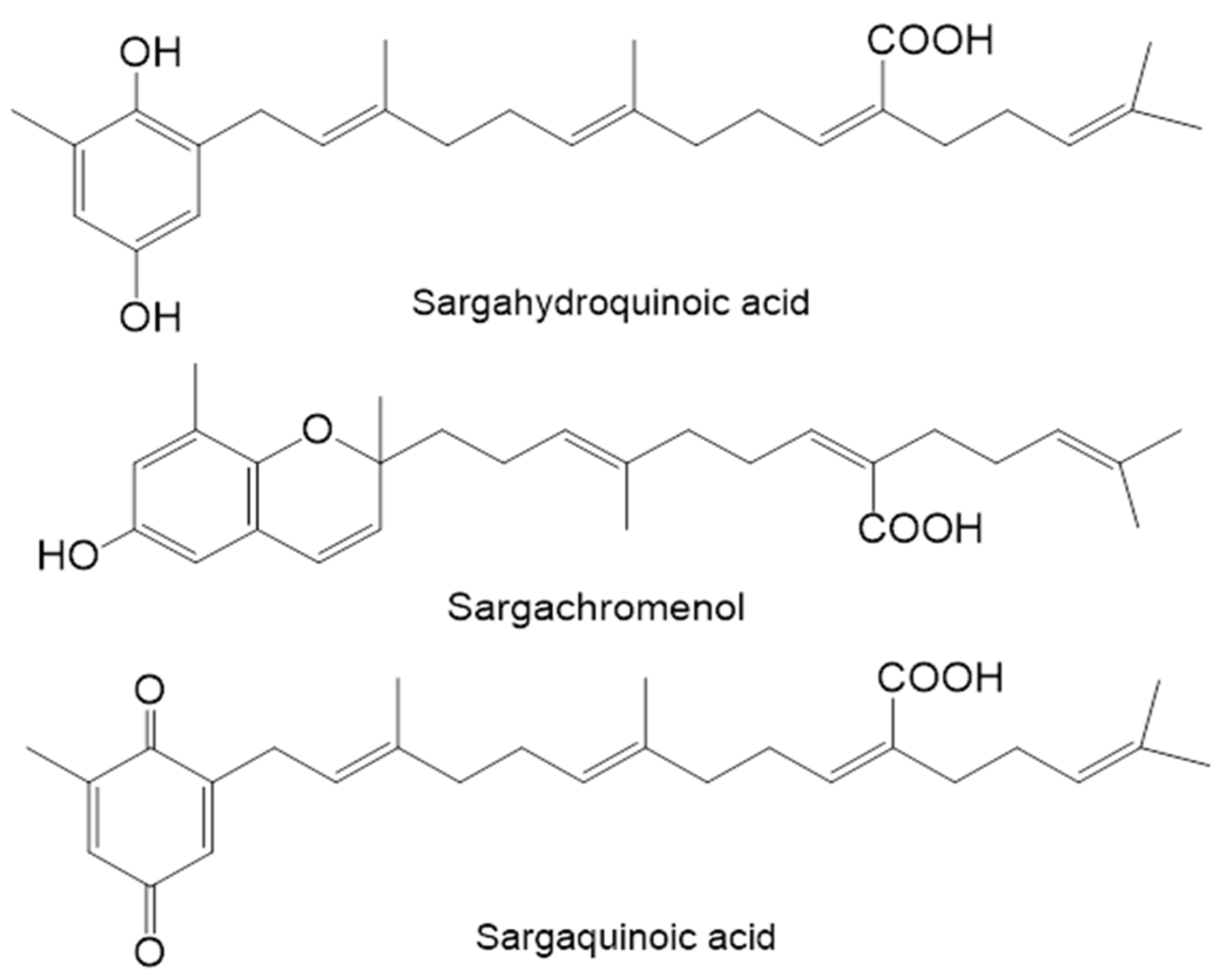
2.2. Inhibitory Activity of Plastoquinones from S. serratifolium against PTP1B and α-Glucosidase
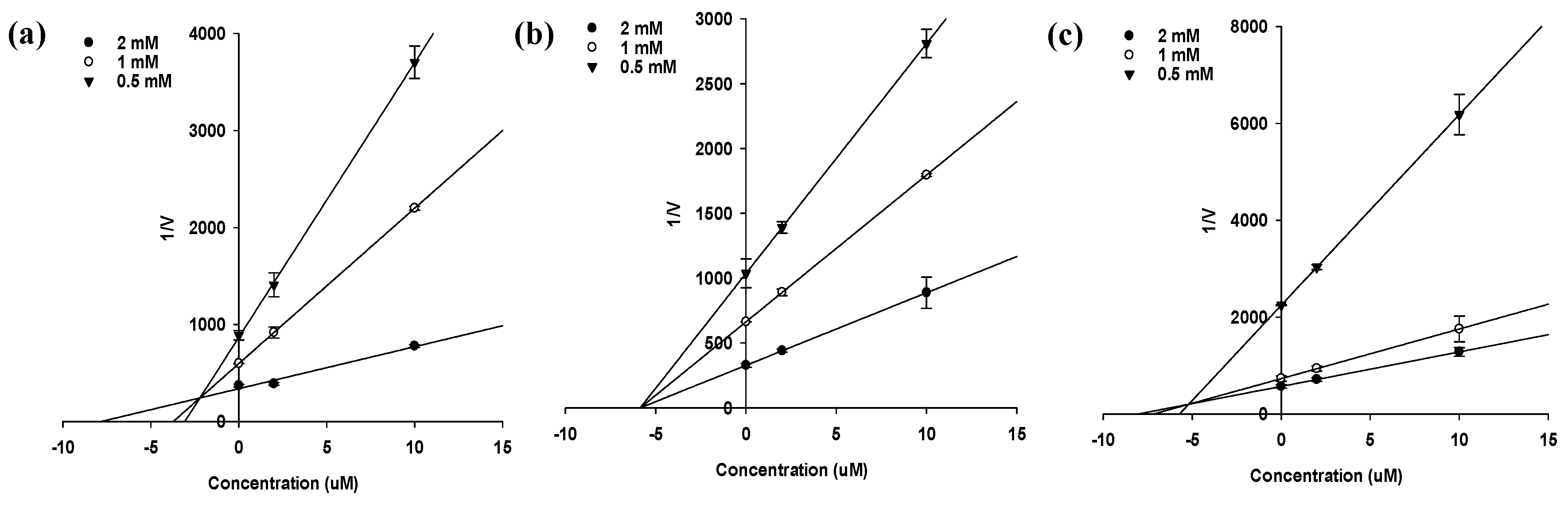
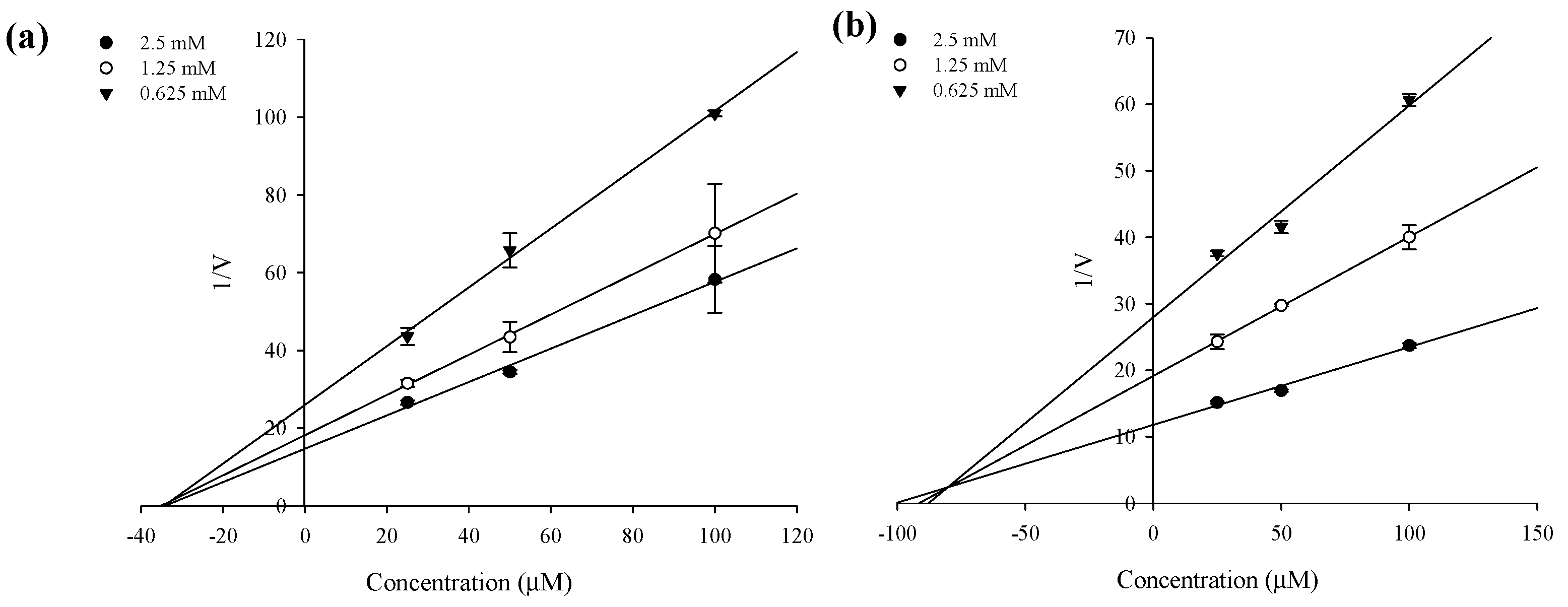
2.3. Enzyme Kinetics of PTP1B and α-Glucosidase Inhibition
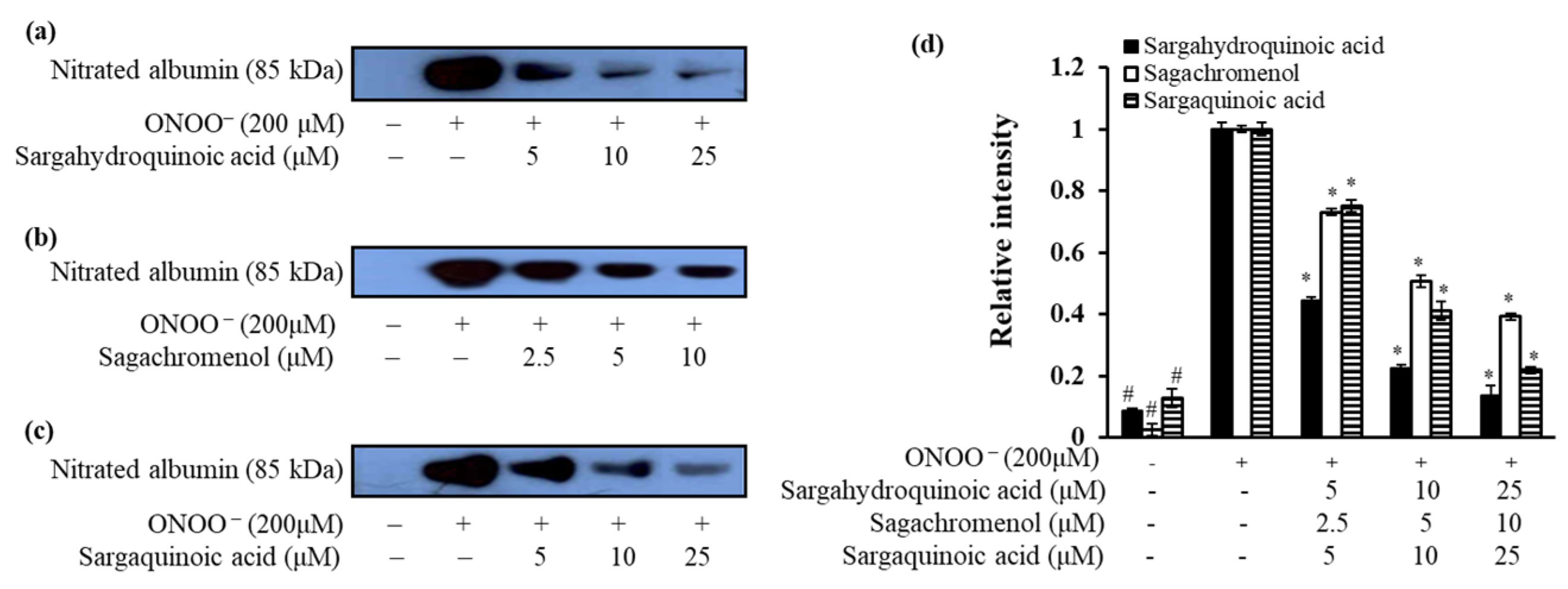
2.4. Inhibitory Effect of Plastoquinones on ONOO–-Mediated Albumin Nitration
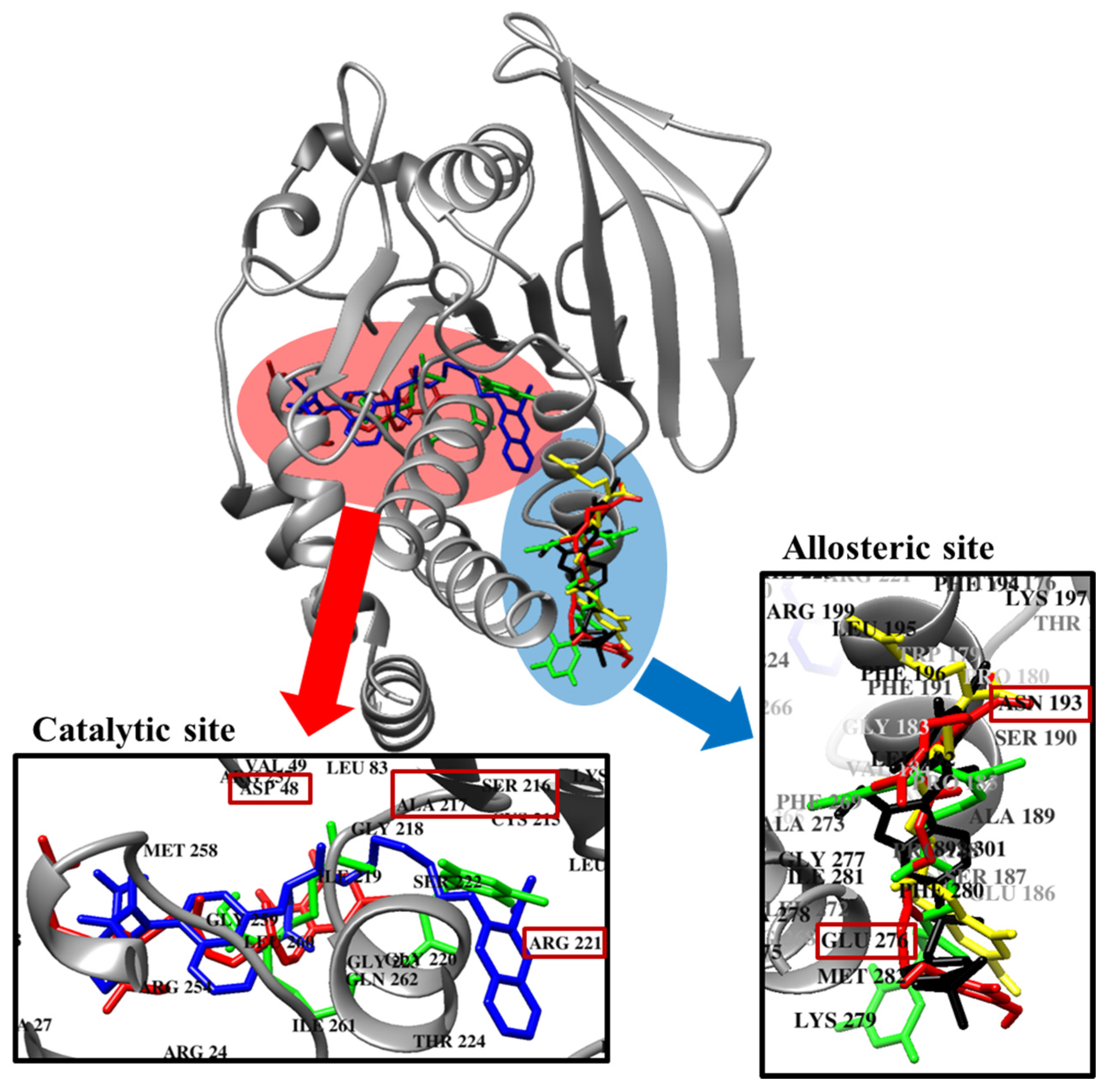
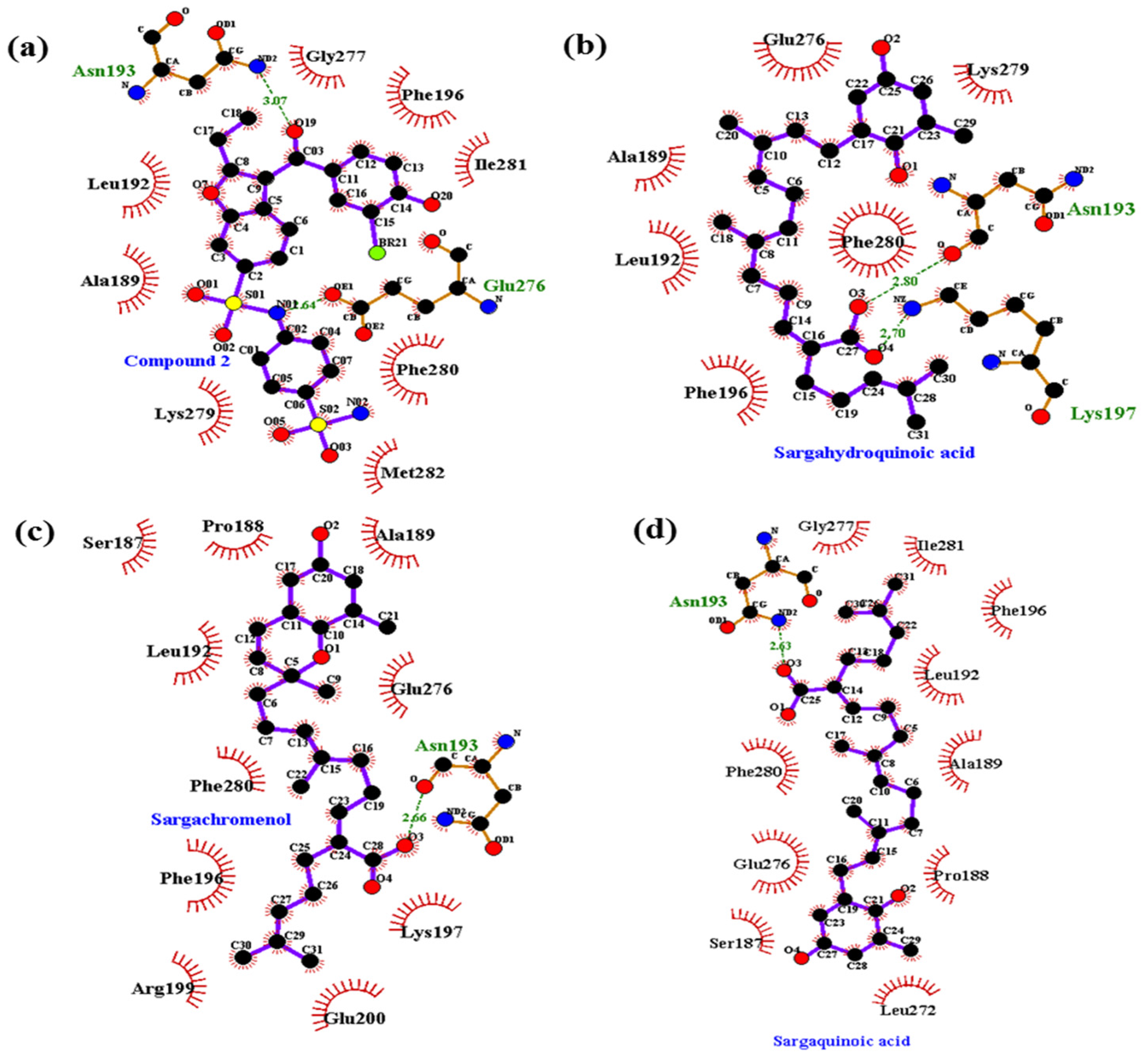
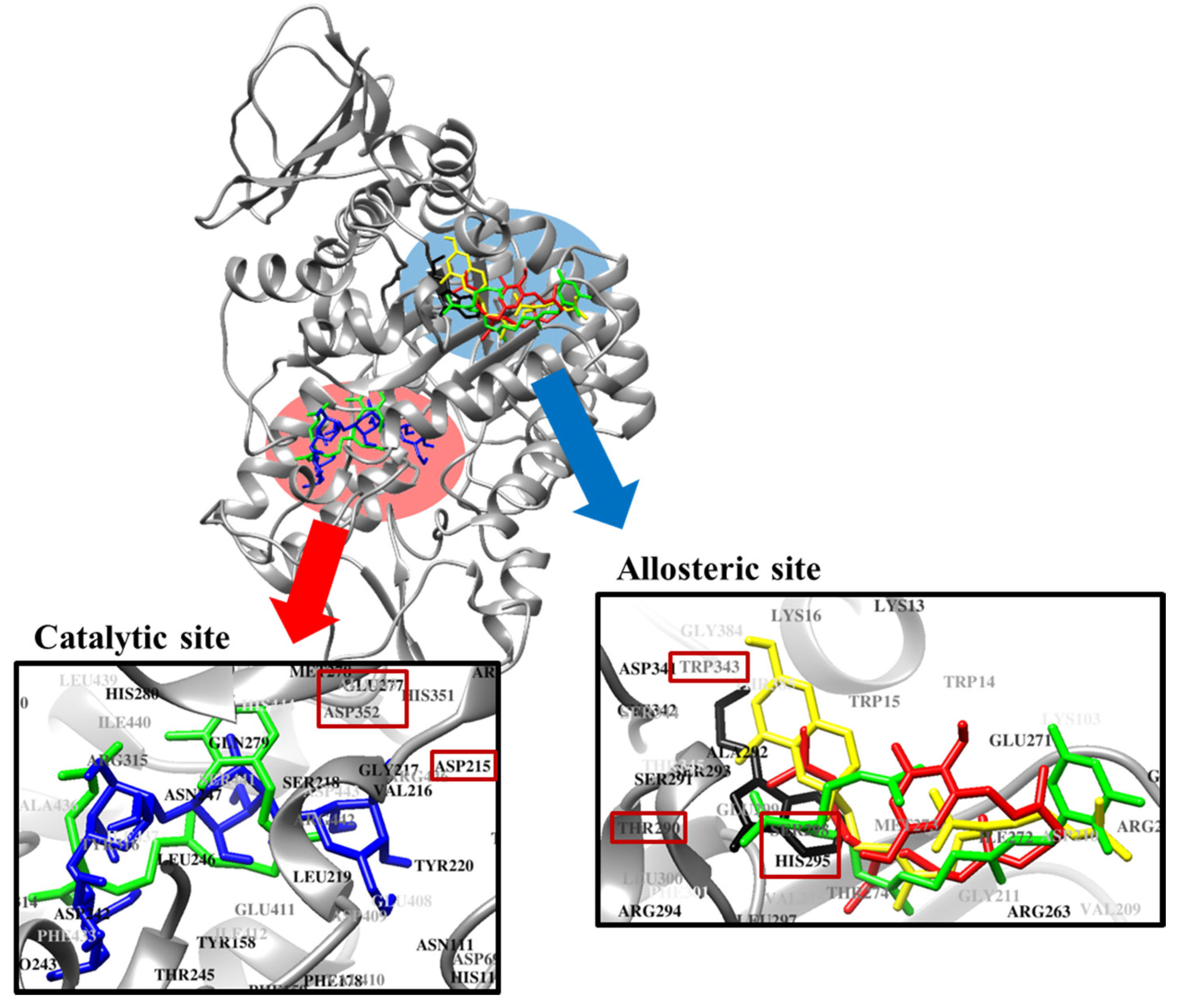
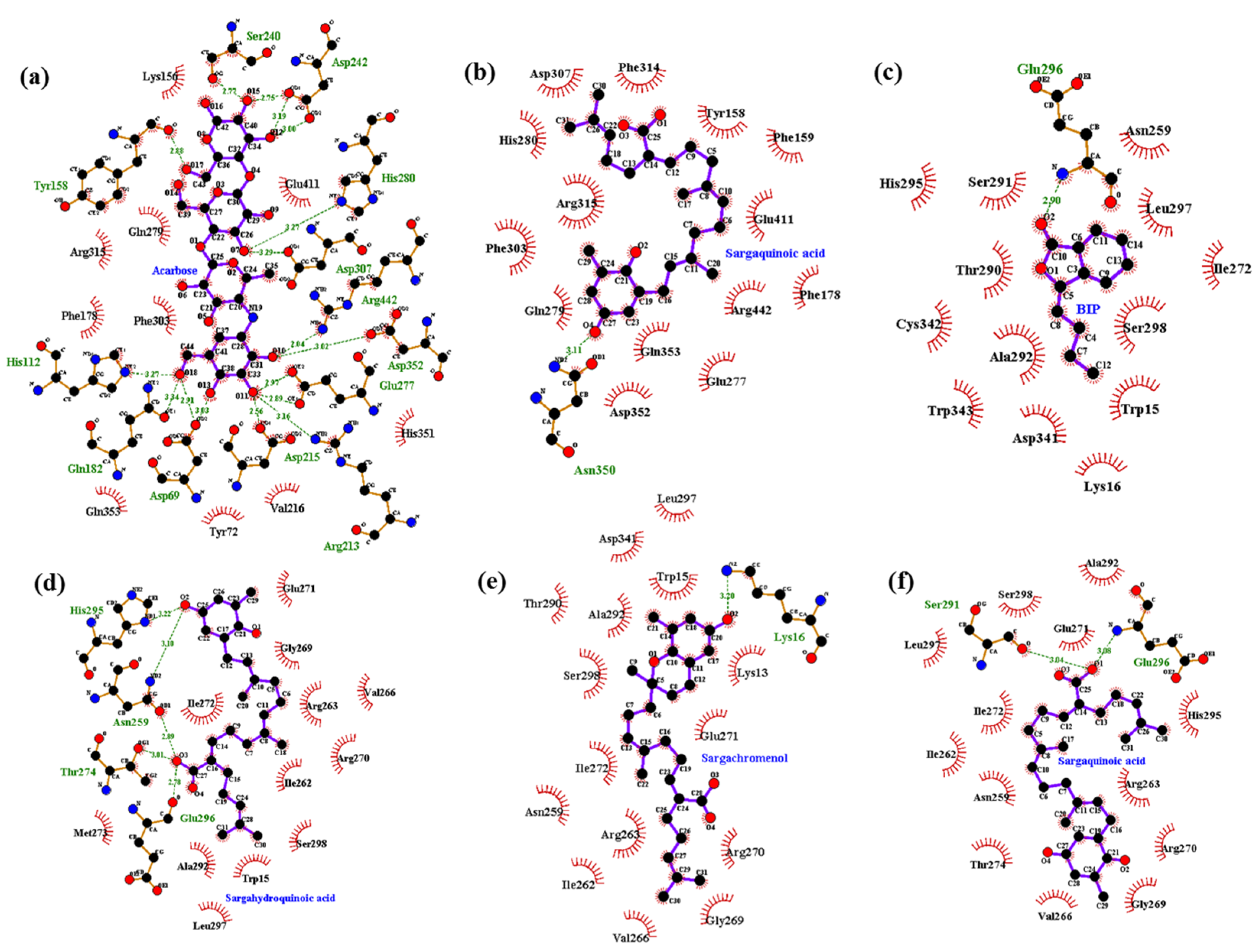
2.5. Molecular Docking Simulation of PTP1B Inhibition
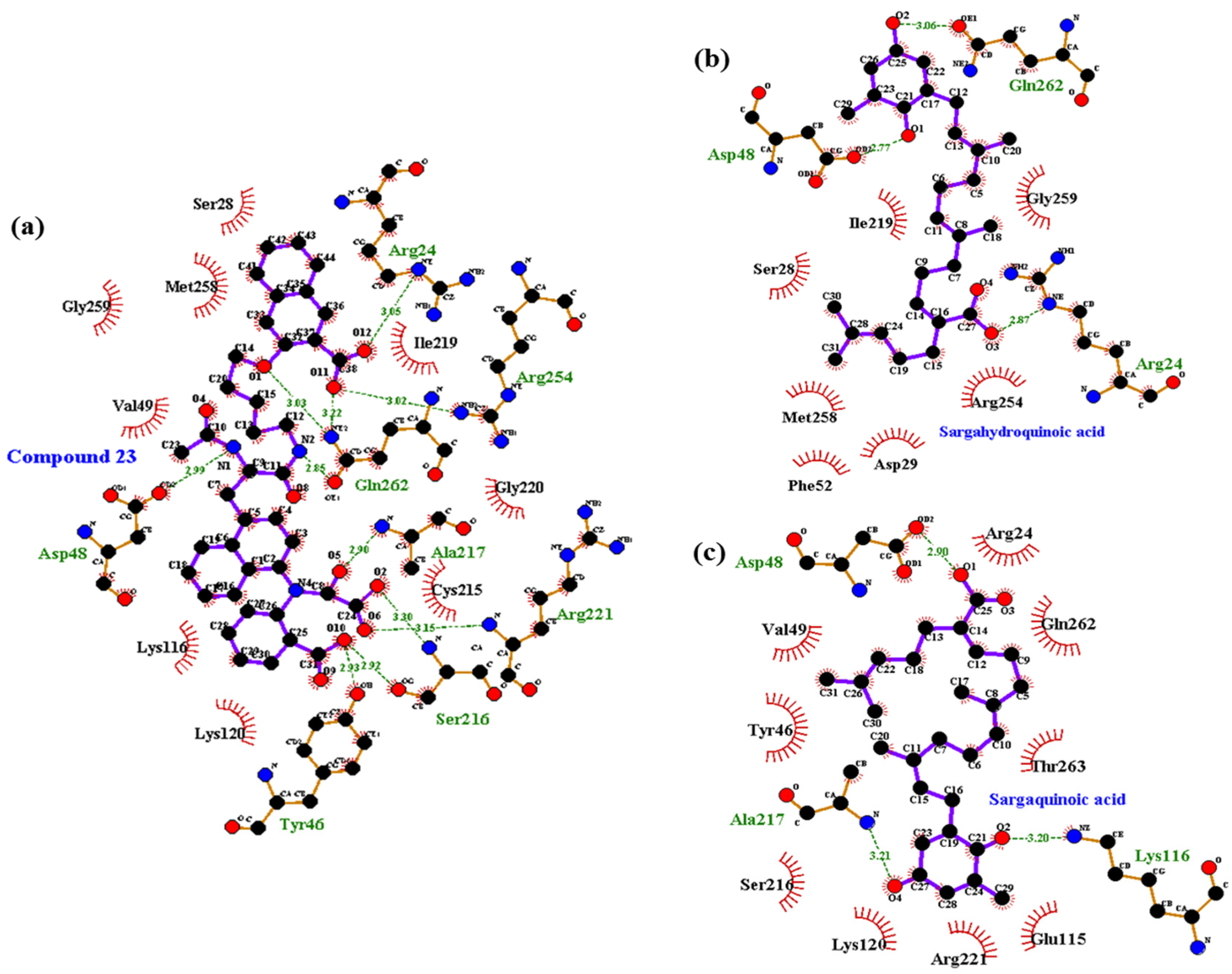
2.6. Molecular Docking Simulation of α-Glucosidase Inhibition
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Chemicals and Reagents
4.3. Extraction, Fractionation, and Isolation
4.4. PTP1B Inhibitory Assay
4.5. α-Glucosidase Inhibitory Assay
4.6. Inhibition of ONOO–-Mediated Albumin Nitration
4.7. Kinetic Study of Plastoquinones against PTP1B and α-Glucosidase
4.8. Molecular Docking Simulation of PTP1B and α-Glucosidase Inhibition
4.9. Statistics
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ramirez-Espinosa, J.J.; Rios, M.Y.; Lopez-Martinez, S.; Lopez-Vallejo, F.; Medina-Franco, J.L.; Paoli, P.; Camici, G.; Navarrete-Vazquez, G.; Ortiz-Andrade, R.; Estrada-Soto, S. Antidiabetic activity of some pentacyclic acid triterpenoids, role of PTP1B: In vitro, in silico, and in vivo approaches. Eur. J. Med. Chem. 2011, 46, 2243–2251. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, H.P.; Arai, R.J.; Travassos, L.R. Protein tyrosine phosphorylation and protein tyrosine nitration in redox signaling. Antioxid. Redox Signal. 2008, 10, 843–889. [Google Scholar] [CrossRef] [PubMed]
- Norrisa, K.; Norrisa, F.; Konod, D.H.; Vestergaardc, H.; Pedersenc, O.; Theofilopoulosd, A.N.; Mollerb, N.P.H. Expression of protein-tyrosine phosphatases in the major insulin target tissues. FEBS Lett. 1997, 415, 243–248. [Google Scholar] [CrossRef]
- Johnson, T.O.; Ermolieff, J.; Jirousek, M.R. Protein tyrosine phosphatase 1B inhibitors for diabetes. Nat. Rev. Drug Discov. 2002, 1, 696–709. [Google Scholar] [CrossRef] [PubMed]
- Seely, B.L.; Staubs, P.A.; Reichart, D.R.; Berhanu, P.; Milarski, K.L.; Saltiel, A.R.; Kusari, J.; Olefsky, J.M. Protein tyrosine phosphatase 1B interacts with the activated insulin receptor. Diabetes 1996, 45, 1379–1385. [Google Scholar] [CrossRef] [PubMed]
- Agouni, A.; Mody, N.; Owen, C.; Czopek, A.; Zimmer, D.; Bentires-Alj, M.; Bence, K.K.; Delibegovic, M. Liver-specific deletion of protein tyrosine phosphatase (PTP) 1B improves obesity- and pharmacologically induced endoplasmic reticulum stress. Biochem. J. 2011, 438, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Wiesmann, C.; Barr, K.J.; Kung, J.; Zhu, J.; Erlanson, D.A.; Shen, W.; Fahr, B.J.; Zhong, M.; Taylor, L.; Randal, M.; et al. Allosteric inhibition of protein tyrosine phosphatase 1B. Nat. Struct. Mol. Biol. 2004, 11, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Lebovitz, H.E. A new oral therapy for diabetes management: Alpha-glucosidase inhibition with acarbose. Clin. Diabetes 1995, 13, 99–103. [Google Scholar]
- van de Laar, F.A.; Lucassen, P.L.; Akkermans, R.P.; von de Lisdonk, E.H.; Rutten, G.E.; van Weel, C. α-Glucosidase inhibitors for patients with type 2 diabetes: Results from a Cochrane systematic review and metaanalysis. Diabetes Care 2005, 28, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Derosa, G.; Maffioli, P. α-Glucosidase inhibitors and their use in clinical practice. Arch. Med. Sci. 2012, 8, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Ischiropoulos, H. Biological tyrosine nitration: A pathophysiological function of nitric oxide and reactive oxygen species. Arch. Biochem. Biophys. 1998, 356, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ceriello, A.; Mercuri, F.; Quagliaro, L.; Assaloni, R.; Motz, E.; Tonutti, L.; Tabaga, C. Detection of nitrotyrosine in the diabetic plasma: Evidence of oxidative stress. Diabetologia 2001, 44, 834–838. [Google Scholar] [PubMed]
- Ceriello, A.; Quagliaro, L.; Catone, B.; Pascon, R.; Piazzola, M.; Bais, B.; Marra, G.; Tonutti, L.; Taboga, C.; Motz, E. Role of hyperglycemia in nitrotyrosine postprandial generation. Diabetes Care 2002, 25, 1439–1443. [Google Scholar] [CrossRef] [PubMed]
- Vadala, M.; Palmieri, B. From algae to “functional foods”. Clin. Ther. 2015, 166, 281–300. [Google Scholar]
- Kim, Y.H.; Kim, J.H.; Kim, D.H.; Kim, S.H.; Kim, H.R.; Kim, Y.M. Synergistic antimicrobial effect of Sargassum serratifolium (C. Agardh) C. Agardh extract against human skin pathogens. Korean J. Food Sci. Technol. 2016, 48, 241–246. [Google Scholar] [CrossRef]
- Kang, C.W.; Park, M.S.; Kim, N.H.; Lee, J.H.; Oh, C.W.; Kim, H.R.; Kim, G.D. Hexane extract from Sargassum serratifolium inhibits the cell proliferation and metastatic ability of human glioblastoma U87MG cells. Oncol. Rep. 2015, 34, 2602–2608. [Google Scholar] [CrossRef] [PubMed]
- Joung, E.J.; Gwon, W.G.; Shin, T.; Jung, B.M.; Choi, J.S.; Kim, H.R. Anti-inflammatory action of the ethanolic extract from Sargassum serratifolium on lipopolysaccharide-stimulated mouse peritoneal macrophages and identification of active components. J. Appl. Phycol. 2016, 29, 563–573. [Google Scholar] [CrossRef]
- Gwon, W.G.; Lee, B.; Joung, E.J.; Choi, M.W.; Yoon, N.; Shin, T.; Oh, C.W.; Kim, H.R. Sargaquinoic acid inhibits TNF-α-induced NF-κB signaling, thereby contributing to decreased monocyte adhesion to human umbilical vein endothelial cells (HUVECs). J. Agric. Food Chem. 2015, 63, 9053–9061. [Google Scholar] [CrossRef] [PubMed]
- Seong, S.H.; Ali, M.Y.; Kim, H.R.; Jung, H.A.; Choi, J.S. BACE1 inhibitory activity and molecular docking analysis of meroterpenoids from Sargassum serratifolium. Bioorg. Med. Chem. 2017, 25, 3964–3970. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Heinrich, M.; Myers, S.; Dworjanyn, S.A. Towards a better understanding of medicinal uses of the brown seaweed Sargassum in traditional Chinese medicine: A phytochemical and pharmacological review. J. Ethnopharmacol. 2012, 142, 591–619. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; Ianora, A. Marine organisms with anti-diabetes properties. Mar. Drugs 2016, 14, 220. [Google Scholar] [CrossRef] [PubMed]
- Pietra, F. Secondary metabolites from marine microorganisms; bacteria, protozoa, algae and fungi: Achievements and perspective. Nat. Prod. Rep. 1997, 14, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Mehdinezhad, N.; Ghannadi, A.; Yegdaneh, A. Phytochemical and biological evaluation of some Sargassum species from Persian Gulf. Res. Pharm. Sci. 2016, 11, 243–249. [Google Scholar] [PubMed]
- Lee, Y.P.; Kang, S.Y. A Catalogue of the Seaweeds in Korea; Jeju National University Press: Jeju, Korea, 2001. [Google Scholar]
- Itoh, H.; Noda, H.; Amano, H.; Zhuaug, C.; Mizuno, T.; Ito, H. Antitumor activity and immunological properties of marine algal polysaccharides, especially fucoidan, prepared from Sargassum thunbergii of Phaeophyceae. Anticancer Res. 1993, 13, 2045–2052. [Google Scholar] [PubMed]
- Choi, B.W.; Ryu, G.; Park, S.H.; Kim, E.S.; Shin, J.; Roh, S.S.; Shin, H.C.; Lee, B.H. Anticholinesterase activity of plastoquinones from Sargassum sagamianum: Lead compounds for Alzheimer’s disease therapy. Phytother. Res. 2007, 21, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.H.; Kang, S.E.; Cho, J.Y.; Kim, A.R.; Park, S.M.; Hong, Y.K.; Ahn, D.H. The antioxidant properties of brown seaweed (Sargassum siliquastrum) extracts. J. Med. Food. 2010, 10, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Ghannadi, A.; Plubrukarn, A.; Zandi, K.; Sartavi, K.; Yegdaneh, A. Screening for antimalarial and acetylcholinesterase inhibitory activities of some Iranian seaweeds. Res. Pharm. Sci. 2013, 8, 113–118. [Google Scholar] [PubMed]
- Vinoth Kumar, T.; Lakshmanasenthil, S.; Geetharamani, D.; Marudhupandi, T.; Suja, G.; Suganya, P. Fucoidan—A α-d-glucosidase inhibitor from Sargassum wightii with relevance to type 2 diabetes mellitus therapy. Int. J. Biol. Macromol. 2015, 72, 1044–1047. [Google Scholar] [CrossRef] [PubMed]
- He, W.F.; Yao, L.G.; Liu, H.L.; Guo, Y.W. Thunberol, a new sterol from the Chinese brown alga Sargassum thunbergii. Asian Nat. Prod. Res. 2014, 16, 685–690. [Google Scholar] [CrossRef] [PubMed]
- D’Orazio, N.; Gammone, M.A.; Gemello, E.; De Girolamo, M.; Cusenza, S.; Riccioni, G. Marine bioactives: Pharmacological properties and potential applications against inflammatory diseases. Mar. Drugs 2012, 10, 812–833. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.A.; Yoon, N.Y.; Woo, M.H.; Choi, J.S. Inhibitory activities of extracts from several kinds of seaweeds and phlorotannins from the brown alga Ecklonia stolonifera on glucose-mediated protein damage and rat lens aldose reductase. Fish. Sci. 2008, 74, 1363–1365. [Google Scholar] [CrossRef]
- Thomas, N.V.; Kim, S.K. Potential pharmacological applications of polyphenolic derivatives from marine brown algae. Environ. Toxicol. Pharmacol. 2011, 32, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Ishimaru, A.; Suzuki, R.; Okuyama, T. A new phloroglucinol derivative from the brown alga Eisenia bicyclis: Potential for the effective treatment of diabetic complications. J. Nat. Prod. 2004, 67, 103–105. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.J.; Hwang, J.Y.; Choi, J.I.; Han, J.S.; Kim, H.J.; Jeon, Y.J. Diphlorethohydroxycarmalol isolated from Ishige okamurae, a brown algae, a potent alpha-glucosidase and alpha-amylase inhibitor, alleviates postprandial hyperglycemia in diabetic mice. Eur. J. Pharmacol. 2009, 615, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Ohigashi, H.; Sakai, Y.; Yamaguchi, K.; Umezaki, I.; Koshimizu, K. Possible anti-tumor promoting properties of marine algae and in vivo activity of Wakame seaweed extract. Biosci. Biotechnol. Biochem. 1992, 56, 994–995. [Google Scholar] [CrossRef] [PubMed]
- Kusumi, T.; Shibata, Y.; Ishitsuka, M.; Kinoshita, T.; Kakisawa, H. Structures of new plastoquinones from the brown alga Sargassum serratifolium. Chem. Lett. 1979, 8, 277–278. [Google Scholar] [CrossRef]
- Hur, S.; Lee, H.; Kim, Y.; Lee, B.H.; Shin, J.; Kim, T.Y. Sargaquinoic acid and sargachromenol, extracts of Sargassum sagamianum, induce apoptosis in HaCaT cells and mice skin: Its potentiation of UVB-induced apoptosis. Eur. J. Pharmacol. 2008, 17, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kamei, Y.; Tsang, C.K. Sargaquinoic acid promotes neurite outgrowth via protein kinase A and MAP kinases-mediated signaling pathways in PC12D cells. Int. J. Dev. Neurosci. 2003, 21, 255–262. [Google Scholar] [CrossRef]
- Tsang, C.K.; Kamei, Y. Sargaquinoic acid supports the survival of neuronal PC12D cells in a nerve growth factor-independent manner. Eur. J. Pharmacol. 2004, 488, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Fernando, P.M.; Piao, M.J.; Hewage, S.R.; Kang, H.K.; Yoo, E.S.; Koh, Y.S.; Ko, M.H.; Ko, C.S.; Byeon, S.H.; Mun, S.R.; et al. Photo-protective effect of sargachromenol against UVB radiation-induced damage through modulating cellular antioxidant systems and apoptosis in human keratinocytes. Environ. Toxicol. Pharmacol. 2016, 23, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Kang, G.J.; Han, S.C.; Yoon, W.J.; Koh, Y.S.; Hyun, J.W.; Kang, H.K.; Cho, J.Y.; Yoo, E.S. Sargaquinoic acid isolated from Sargassum siliquastrum inhibits lipopolysaccharide-induced nitric oxide production in macrophages via modulation of nuclear factor-κ B and c-Jun N-terminal kinase pathways. Immunopharmacol. Immunotoxicol. 2013, 35, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.J.; Ham, Y.M.; Yang, K.W.; Lee, N.H.; Hyun, C.G. Sargachromenol from Sargassum micracanthum inhibits the lipopolysaccharide-induced production of inflammatory mediators in RAW 264.7 macrophages. Sci. World J. 2013. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Lee, M.S.; Lee, B.G.; Gwon, W.G.; Joung, E.J.; Yoon, N.Y.; Kim, H.R. Anti-inflammatory effects of sargachromenol-rich ethanolic extract of Myagropsis myagroides on lipopolysaccharide-stimulated BV-2 cells. BMC Compliment. Altern. Med. 2014, 9, 231–242. [Google Scholar]
- Jeon, Y.; Jung, Y.; Kim, M.C.; Kwon, H.C.; Kang, K.S.; Kim, Y.K.; Kim, S.N. Sargahydroquinoic acid inhibits TNFα-induced AP-1 and NF-κB signaling in HaCaT cells through PPARα activation. Biochem. Biophys. Res. Commun. 2014, 450, 1553–1559. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.N.; Lee, W.; Bae, G.U.; Kim, Y.K. Anti-diabetic and hypolipidemic effects of Sargassum yezoense in db/db mice. Biochem. Biophys. Res. Commun. 2012, 424, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.N.; Choi, H.Y.; Lee, W.; Park, G.M.; Shin, W.S.; Kim, Y.K. Sargaquinoic acid and sargahydroquinoic acid from Sargassum yezoense stimulate adipocyte differentiation through PPAR α/γ activation in 3T3-L1 cells. FEBS Lett. 2008, 582, 3465–3472. [Google Scholar] [CrossRef] [PubMed]
- de la Mare, J.A.; Lawson, J.C.; Chiwakata, M.T.; Beukes, D.R.; Edkins, A.L.; Blatch, G.L. Quinones and halogenated monoterpenes of algal origin show anti-proliferative effects against breast cancer cells in vitro. Investig. New Drugs 2012, 30, 2187–2200. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Lee, S.Y. PTP1B inhibitors as potential therapeutics in the treatment of type 2 diabetes and obesity. Expert Opin. Investig. Drugs 2003, 12, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Tiganis, T.; Bennett, A.M. Protein tyrosine phosphatase function: the substrate perspective. Biochem. J. 2007, 402, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bakke, J.; Haj, F.G. Protein tyrosine phosphatase 1B substrates and metabolic regulation. Semin. Cell Dev. Biol. 2015, 37, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, D.; Kusari, A.; Kenner, K.A.; Liu, F.; Chernoff, J.; Gustafson, T.A.; Kusari, J. Protein-tyrosine phosphatase 1B complexes with the insulin receptor in vivo and is tyrosine phosphorylated in the presence of insulin. J. Biol. Chem. 1997, 272, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- Salmeen, A.; Andersen, J.N.; Myers, M.P.; Tonks, N.K.; Barford, D. Molecular basis for the dephosphorylation of the activation segment of the insulin receptor by protein tyrosine phosphatase 1B. Mol. Cell 2000, 6, 1401–1412. [Google Scholar] [CrossRef]
- Zabolotny, J.M.; Bence-Hanulec, K.K.; Stricker-Krongrad, A.; Haj, F.; Wang, Y.; Minokoshi, Y.; Kim, Y.B.; Elmquist, J.K.; Tartaglia, L.A.; Kahn, B.B.; et al. PTP1B regulates leptin signal transduction in vivo. Dev. Cell 2002, 2, 489–495. [Google Scholar] [CrossRef]
- Cheng, A.; Uetani, N.; Simoncic, P.D.; Chaubey, V.P.; Lee-Loy, A.; McGlade, C.J.; Kennedy, B.P.; Tremblay, M.L. Attenuation of leptin action and regulation of obesity by protein tyrosine phosphatase 1B. Dev. Cell 2002, 2, 497–503. [Google Scholar] [CrossRef]
- Liu, Z.; chai, Q.; Li, Y.Y.; Shen, Q.; Ma, L.P.; Zhang, L.N.; Wang, X.; Sheng, L.; Li, J.Y.; Li, J.; et al. Discovery of novel PTP1B inhibitors with antihyperglycemic activity. Acta Pharmacol. Sin. 2010, 31, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.J.; Jiang, B.; Wu, N.; Wang, S.Y.; Shi, D.Y. Small molecules as potent protein tyrosine phosphatase 1B (PTP1B) inhibitors documented in patents from 2009 to 2013. Mini-Rev. Med. Chem. 2015, 15, 104–122. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, N.; Koveal, D.; Miller, D.H.; Xue, B.; Akshinthala, S.D.; Kragelj, J.; Jensen, M.R.; Gauss, C.M.; Page, R.; Blackledge, M.; et al. Targeting the disordered C terminus of PTP1B with an allosteric inhibitor. Nat. Chem. Biol. 2014, 10, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Szczepankiewicz, B.G.; Liu, G.; Hajduk, P.J.; Abad-Zapatero, C.; Pei, Z.; Xin, Z.; Lubben, T.H.; Trevillyan, J.M.; Stashko, M.A.; Ballaron, S.J.; et al. Discovery of a potent, selective protein tyrosine phosphatase 1B inhibitor using a linked-fragment strategy. J. Am. Chem. Soc. 2003, 125, 4087–4096. [Google Scholar] [CrossRef] [PubMed]
- Braun, C.; Brayer, G.D.; Withers, S.G.J. Mechanism-based inhibition of yeast alpha-glucosidase and human pancreatic alpha-amylase by a new class of inhibitors. 2-Deoxy-2,2-difluoro-alpha-glycosides. Biol. Chem. 1995, 270, 26778–26781. [Google Scholar] [CrossRef]
- Robinson, K.M.; Begovic, M.E.; Rhinehart, B.L.; Heineke, E.W.; Ducep, J.B.; Kastner, P.R.; Marshall, F.N.; Danzin, C. New potent α-glucohydrolase inhibitor MDL 73945 with long duration of action in rats. Diabetes 1991, 40, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Oki, T.; Matsui, T.; Osajima, Y. Inhibitory effect of alpha-glucosidase inhibitors varies according to its origin. J. Agric. Food Chem. 1999, 47, 550–553. [Google Scholar] [CrossRef] [PubMed]
- Oku, T.; Tanabe, K.; Ogawa, S.; Sadamori, N.; Nakamura, S. Similarity of hydrolyzing activity of human and rat small intestinal disaccharidases. Clin. Exp. Gastroenterol. 2011, 4, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Takakura, K.; Beckman, J.S.; MacMillan-Crow, L.A.; Crow, J.P. Rapid and irreversible inactivation of protein tyrosine phosphatases PTP1B, CD45, and LAR by peroxynitrite. Arch. Biochem. Biophys. 1999, 369, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Nowicka, B.; Kruk, J. Occurrence, biosynthesis and function of isoprenoid quinones. Biochim. Biophys. Acta 2010, 1797, 1587–1605. [Google Scholar] [CrossRef] [PubMed]
- Mori, J.; Hayashi, T.; Iwashima, M.; Matsunaga, T.; Saito, H. Effects of plastoquinones from the brown alga Sargassum micracanthum and a new chromene derivative converted from the plastoquinones on acute gastric lesions in rats. Biol. Pharm. Bull. 2006, 29, 1197–1201. [Google Scholar] [CrossRef] [PubMed]
- Park, B.G.; Shin, W.S.; Um, Y.; Cho, S.; Park, G.M.; Yeon, D.S.; Kwon, S.C.; Ham, J.; Choi, B.W.; Lee, S. Selective vasodilatation effect of sargahydroquinoic acid, an active constituent of Sargassum micracanthum, on the basilar arteries of rabbits. Bioorg. Med. Chem. Lett. 2008, 18, 2624–2627. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Na, M.; Oh, H.; Bae, E.Y.; Jeong, D.G.; Ryu, S.E.; Kim, S.; Kim, B.Y.; Oh, W.K.; Ahn, J.S. Protein tyrosine phosphatase 1B inhibitors from Morus root bark. Bioorg. Med. Chem. Lett. 2006, 16, 1426–1429. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhang, X.D.; Song, Y.W.; Liu, J.W. A microplate-based screening method for α-glucosidase inhibitors. Chin. J. Clin. Pharmacol. Ther. 2005, 10, 1128–1134. [Google Scholar]
- Aulak, K.S.; Miyagi, M.; Yan, L.; West, K.A.; Massillon, D.; Crabb, J.W.; Stuehr, D.J. Proteomic method identifies proteins nitrated in vivo during inflammatory challenge. Proc. Natl. Acad. Sci. USA 2001, 98, 12056–12061. [Google Scholar] [CrossRef] [PubMed]
- Dixon, M. The determination of enzyme inhibitor constant. Biochem. J. 1953, 55, 170–171. [Google Scholar] [CrossRef] [PubMed]
- Cornish-Bowden, A. A simple graphical method for determining the inhibition constants of mixed, uncompetitive and non-competitive inhibitors. Biochem. J. 1974, 137, 143–144. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zhang, G.; Pan, J.; Wang, Y. α-Glucosidase inhibition by luteolin: Kinetics, interaction and molecular docking. Int. J. Biol. Macromol. 2014, 64, 213–223. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | IC50 Values (Mean ± SEM) a | |
|---|---|---|
| PTP1B | α-Glucosidase | |
| EtOH extract | 7.04 ± 0.26 d | 24.16 ± 0.31 e |
| n-Hexane fraction | 1.83 ± 0.06 g | 16.73 ± 0.14 f |
| CH2Cl2 fraction | 6.32 ± 0.04 e | 14.61 ± 0.99 g |
| EtOAc fraction | 1.88 ± 0.09 g | 3.16 ± 0.10 h |
| n-BuOH fraction | 4.87 ± 0.24 f | 15.22 ± 0.25 g |
| H2O fraction | >100 | >100 |
| Ursolic acid b | 1.12 ± 0.19 h | |
| Acarbose c | 108.74 ± 2.96 d | |
| Compound | PTP1B | α-Glucosidase | ||||
|---|---|---|---|---|---|---|
| IC50 (μM) a | Inhibition Type b | Ki (μM) c | IC50 (μM) a | Inhibition Type b | Ki (μM) c | |
| Sargahydroquinoic acid | 5.14 ± 0.07 h | Mixed | 2.21 | >100 | - | - |
| Sargachromenol | 11.80 ± 3.35 f | Non-competitive | 5.85 | 42.41 ± 3.09 h | Non-competitive | 33.95 |
| Sargaquinoic acid | 14.15 ± 0.02 f | Mixed | 5.20 | 96.17 ± 3.48 g | Mixed | 79.68 |
| Ursolic acid d | 6.09 ± 0.02 g | |||||
| Acarbose e | 210.76 ± 4.52 f | |||||
| Compound | Binding Energy a (kcal/mol) | No. of H-Bond b | H-Bond Interacting Residues c | van der Waals Bond Interacting Residues c |
|---|---|---|---|---|
| Compound 23 d (catalytic inhibitor) | −11.23 | 11 | Arg24, Tyr46, Asp48, Ser216, Ala217, Arg221, Arg254, Gln262 | Ser28, Val49, Lys116, Lys120, Cys215, Ile219, Gly220, Met258, Gly259 |
| Compound 2 d (allosteric inhibitor) | −10.98 | 2 | Asn193, Glu276 | Ala189, Leu192, Phe196, Gly277, Lys279, Phe280, Ile281, Met282 |
| Sargahydroquinoic acid e | −5.09 | 3 | Arg24, Asp48, Gln262 | Ile219, Ser28, Met258, Phe52, Asp29, Arg254, Gly259 |
| −5.95 | 2 | Asn193, Lys197 | Ala189, Leu192, Phe196, Glu276, Lys279, Phe280 | |
| Sargachromenol | −8.84 | 1 | Asn193 | Ser187, Pro188, Ala189, Leu192, Phe196, Lys197, Arg199, Glu200, Glu276, Phe280 |
| Sargaquinoic acid e | −3.13 | 3 | Asp48, Lys116, Ala217 | Arg24, Gln262, Thr263, Glu115, Arg221, Lys120, Ser216, Tyr46, Val49 |
| −6.83 | 1 | Asn193 | Ser187, Pro188, Ala189, Leu192, Phe196, Leu272, Glu276, Gly277, Phe280, Ile281 |
| Compound | Binding Energy a (kcal/mol) | No. of H-Bond b | H-Bond Interacting Residues c | van der Waals Bond Interacting Residues c |
|---|---|---|---|---|
| Acarbose d (catalytic inhibitor) | −8.6 | 17 | Asp69, Gln82, His112, Tyr158, Arg213, Asp215, Ser240, Asp242, Glu277, His280, Asp307, Asp352, Arg442 | Tyr72, Lys156, Phe178, Val216, Gln279, Phe303, Arg315, His351, Gln353, Glu411 |
| BIP d (allosteric inhibitor) | −6.75 | 1 | Glu296 | Trp15, Lys16, Asn259, Arg263, Val266, Gly269, Glu271, Ile272, Thr290, Ser291, Ala292,His295, Leu297, Ser298, Asp341, Cys342, Trp343 |
| Sargahydroquinoic acid | −8.0 | 5 | Glu296, Asn259, Thr274, His295 | Trp15, Ile262, Arg270, Ile272, Val266, Ala292, Met273, Leu297, Ser298, Gly269, Glu271,Arg263 |
| Sargachromenol | −7.3 | 1 | Lys16 | Lys13, Trp15, Asn259, Ile262, Arg263, Val266, Gly269, Arg270, Glu271, Ile272,Thr290, Ala292, Leu297, Ser298,Asp341 |
| Sargaquinoic acid e | −5.38 | 1 | Asn350 | Tyr158, Phe159, Phe178, Glu277, Gln279, His280, Phe303, Asp307, Phe314, Arg315, Asp352, Gln353, Glu411, Arg442 |
| −7.1 | 2 | Ser291, Glu296 | Asn259, Ile262,Arg263, Val266, Gly269, Arg270, Glu271,Ile272, Thr274, Leu297, Ala292, His295, Ser298 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, M.Y.; Kim, D.H.; Seong, S.H.; Kim, H.-R.; Jung, H.A.; Choi, J.S. α-Glucosidase and Protein Tyrosine Phosphatase 1B Inhibitory Activity of Plastoquinones from Marine Brown Alga Sargassum serratifolium. Mar. Drugs 2017, 15, 368. https://doi.org/10.3390/md15120368
Ali MY, Kim DH, Seong SH, Kim H-R, Jung HA, Choi JS. α-Glucosidase and Protein Tyrosine Phosphatase 1B Inhibitory Activity of Plastoquinones from Marine Brown Alga Sargassum serratifolium. Marine Drugs. 2017; 15(12):368. https://doi.org/10.3390/md15120368
Chicago/Turabian StyleAli, Md. Yousof, Da Hye Kim, Su Hui Seong, Hyeung-Rak Kim, Hyun Ah Jung, and Jae Sue Choi. 2017. "α-Glucosidase and Protein Tyrosine Phosphatase 1B Inhibitory Activity of Plastoquinones from Marine Brown Alga Sargassum serratifolium" Marine Drugs 15, no. 12: 368. https://doi.org/10.3390/md15120368
APA StyleAli, M. Y., Kim, D. H., Seong, S. H., Kim, H.-R., Jung, H. A., & Choi, J. S. (2017). α-Glucosidase and Protein Tyrosine Phosphatase 1B Inhibitory Activity of Plastoquinones from Marine Brown Alga Sargassum serratifolium. Marine Drugs, 15(12), 368. https://doi.org/10.3390/md15120368