
Antimicrobials from Cnidarians. A New Perspective for Anti-Infective Therapy?


Abstract
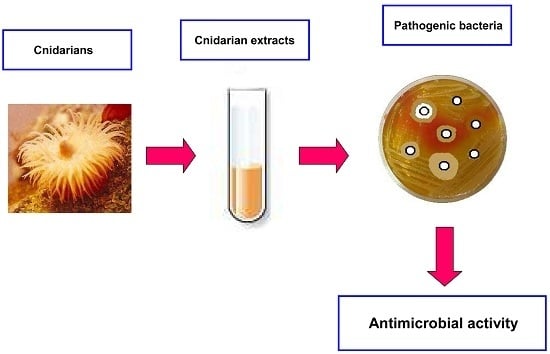
:
1. The Discovery of Antibiotics and the Antibiotic Era
2. Antibiotic/Antimicrobial Resistance
3. The Role of Natural Compounds in Drug Discovery
4. Cnidarians as a New Option for the Development of Drugs?
5. Antimicrobials from Cnidaria
5.1. Freshwater Cnidaria
Antimicrobials from Hydrozoa
5.2. Marine Cnidaria
5.2.1. Antimicrobials from Anthozoa
5.2.2. Antimicrobials from Scyphozoa and Cubozoa
5.3. Antiviral Activity of Cnidarian Extracts
5.4. Antiparasitic Activity of Cnidarian Extracts
6. Conclusions
Conflicts of Interest
References
- Aminov, R.I. A brief history of the antibiotic era: Lessons learned and challenges for the future. Front. Microbiol. 2010, 1, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bassett, E.J.; Keith, M.S.; Armelagos, G.J.; Martin, D.L.; Villanueva, A.R. Tetracycline-labeled human bone from ancient Sudanese Nubia (A.D. 350). Science 1980, 209, 1532–1534. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.L.; Dinardo, A.; Hochberg, J.; Armelagos, G.J. Brief communication: Mass spectroscopic characterization of tetracycline in the skeletal remains of an ancient population from Sudanese Nubia 350–550 CE. Am. J. Phys. Anthropol. 2010, 143, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.; Molto, E.; Anderson, C. Fluorochrome labelling in roman period skeletons from Dakhleh oasis, Egypt. Am. J. Phys. Anthropol. 1989, 80, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Armelagos, G.J. Disease in ancient Nubia. Science 1969, 163, 225–258. [Google Scholar] [CrossRef]
- Sköld, O. Antibiotics and Antibiotic Resistance; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011; pp. 1–207. [Google Scholar]
- Tiberio, V. Sugli estratti di alcune muffe. Ann. Ig. Sper. 1895, 1, 91–103. [Google Scholar]
- Schwartz, M. Historical streptococci. In Streptococci and the Host; Horaud, T., Bouvet, A., Leclercq, R., de Montclos, H., Sicard, M., Eds.; Plenum Press: New York, NY, USA, 1997; Chapter 1; pp. 1–2. [Google Scholar]
- Fleming, A. On the antibacterial action of cultures of a Penicillium, with special reference to their use in the isolation of B. influenzae. Br. J. Exp. Pathol. 1929, 10, 226–236. [Google Scholar] [CrossRef]
- Schatz, A.; Bugle, E.; Waksman, S.A. Streptomycin, a substance exhibiting antibiotic activity against gram-positive and gram-negative bacteria. Proc. Soc. Exp. Biol. Med. 1944, 55, 66–69. [Google Scholar] [CrossRef]
- Brotzu, G. Ricerche su di un Nuovo Antibiotico. Available online: http://medicina.unica.it/pacs/brotzu.pdf (accessed on 7 March 2016).
- Bo, G. Giuseppe Brotzu and the discovery of cephalosporins. Clin. Microbiol. Infect. 2000, 6 (Suppl. S3), 6–8. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, S. The origin of cephalosporins. Yakushigaku Zasshi 2003, 37, 119–127. [Google Scholar] [PubMed]
- Abraham, E.P.; Newton, G.G.F.; Crawford, K.; Burton, H.S.; Hale, C.W. Cephalosporin N: A new type of penicillin. Nature 1953, 171, 343. [Google Scholar] [CrossRef] [PubMed]
- Abraham, E.P.; Newton, G.G.F.; Hale, C.W. Purification and some properties of cephalosporin N, a new penicillin. Biochem. J. 1954, 58, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Abraham, E.P.; Newton, G.G.F. Experiments on the degradation of cephalosporin C. Biochem. J. 1956, 62, 658–665. [Google Scholar] [CrossRef] [PubMed]
- Crawfork, K.; Abraham, E.P. The synergistic action of Cephalosporin C and Benzylpenicillin against a penicillinase-producing strain of Staphylococcus aureus. J. Gen. Microbiol. 1957, 16, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Ganapathi, K. Twenty years of antibiotics—A review. Indian J. Physiol. Pharmacol. 1961, 7, 91–112. [Google Scholar]
- Bennett, J.W. What is an Antibiotic? In Antibiotics: Current Innovations and Future Trends; Sánchez, S., Demain, A.L., Eds.; Caister Academic Press: Poole, UK, 2015; Chapter 1; pp. 1–18. [Google Scholar]
- Michigan State University. Antimicrobials: An introduction. Antimicrobial Resistance Learning Site. Pharmacology. 2011. Available online: http://amrls.cvm.msu.edu/pharmacology/antimicrobials/antimicrobials-an-introduction (accessed on 27 November 2015).
- WHO. Antimicrobial Resistance. Fact Sheet N°194. Updated April 2015. Available online: http://www.who.int/mediacentre/factsheets/fs194/en/ (accessed on 27 November 2015).
- Alekshun, M.N.; Levy, S.B. Molecular mechanisms of antibacterial multidrug resistance. Cell 2007, 128, 1037–1050. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [PubMed]
- Donné, J.; Dewilde, S. The challenging world of biofilm physiology. Adv. Microb. Physiol. 2015, 67, 235–292. [Google Scholar] [PubMed]
- Pane, G.; Cacciola, G.; Giacco, E.; Mariottini, G.L.; Coppo, E. Assessment of the antimicrobial activity of algae extracts on bacteria responsible of external otitis. Mar. Drugs 2015, 13, 6440–6452. [Google Scholar] [CrossRef] [PubMed]
- Cash, P. Proteomics in medical microbiology. Electrophoresis 2000, 21, 1187–1201. [Google Scholar] [CrossRef]
- Dos Santos, K.V.; Galuppo Diniz, C.; de Castro Veloso, L.; Monteiro de Andrade, H.; da Silva Giusta, M.; da Fonseca Pires, S.; Valadares Santos, A.; Morais Apolônio, A.C.; Roque de Carvalho, M.A.; de Macêdo Farias, L. Proteomic analysis of Escherichia coli with experimentally induced resistance topiperacillin/tazobactam. Res. Microbiol. 2010, 161, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Groisman, E.A.; Ochman, H. Pathogenicity islands: Bacterial evolution in quantum leaps. Cell 1996, 87, 791–794. [Google Scholar] [CrossRef]
- Ochman, H.; Lawrence, J.G.; Groisman, E.A. Lateral gene transfer and the nature of bacterial innovation. Nature 2000, 405, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Tomazella, G.G.; Risberg, K.; Mylvaganam, H.; Lindemann, P.C.; Thiede, B.; de Souza, G.A.; Wiker, H.G. Proteomic analysis of a multi-resistant clinical Escherichia coli isolate of unknown genomic background. J. Proteom. 2012, 75, 1830–1837. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.; Bont, L.; Engelhard, D.; Moore, E.; Fernández, D.; Kreisberg-Greenblatt, R.; Oved, K.; Eden, E.; Hays, J.P. A multifaceted “omics” approach for addressing the challenge of antimicrobial resistance. Future Microbiol. 2015, 10, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, R.; Correia, S.; Poeta, P.; Pinto, L.; Igrejas, G. The role of proteomics in elucidating multiple antibiotic resistance in Salmonella and in novel antibacterial discovery. In Salmonella—Distribution, Adaptation, Control Measures and Molecular Technologies; Annous, B.A., Gurtler, J.B., Eds.; InTech: Rijeka, Croatia, 2012; Chapter 10; pp. 187–220. [Google Scholar]
- Newman, D.J.; Cragg, G.M.; Snader, K.M. Natural products as sources of new drugs over the period 1981–2002. J. Nat. Prod. 2003, 66, 1022–1037. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M.; Snader, K.M. The influence of natural products upon drug discovery. Nat. Prod. Rep. 2000, 17, 215–234. [Google Scholar] [CrossRef] [PubMed]
- Proksch, P.; Edrada-Ebel, R.A.; Ebel, R. Drugs from the sea—Opportunities and obstacles. Mar. Drugs 2003, 1, 5–17. [Google Scholar] [CrossRef]
- Cragg, G.M.; Newman, D.J. Natural products: A continuing source of novel drug leads. Biochim. Biophys. Acta 2013, 1830, 3670–3695. [Google Scholar] [CrossRef] [PubMed]
- Leone, A.; Lecci, R.M.; Durante, M.; Piraino, S. Extract from the zooxanthellate jellyfish Cotylorhiza tuberculata modulates gap junction intercellular communication in human cell cultures. Mar. Drugs 2013, 11, 1728–1762. [Google Scholar] [CrossRef] [PubMed]
- Jones, D. Structure and experimental uses of arthropod venom proteins. Adv. Exp. Med. Biol. 1996, 391, 379–386. [Google Scholar] [PubMed]
- Skarnes, R.C. L-Amino acid oxidase, a bactericidal system. Nature 1970, 225, 1072–1073. [Google Scholar] [CrossRef] [PubMed]
- Tan, N.H.; Fung, S.Y. Snake venom l-amino acid oxidase. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S., Ed.; CRC Press: New York, NY, USA, 2009; pp. 219–232. [Google Scholar]
- Yang, H.; Johnson, P.M.; Ko, K.C.; Kamio, M.; Germann, M.W.; Derby, C.D.; Tai, P.C. Cloning, characterization and expression of escapin, a broadly antimicrobial FAD-containing l-amino acid oxidase from ink of the sea hare Aplysia californica. J. Exp. Biol. 2005, 208, 3609–3622. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, Y.; Tsukamoto, C.; Kitani, Y.; Ishizaki, S.; Nagai, H.; Yanagimoto, T. Isolation and cDNA cloning of an antibacterial l-amino acid oxidase from the skin mucus of the great sculpin Myoxocephalus polyacanthocephalus. Comp. Biochem. Physiol. B 2009, 154, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Abdelkafi-Koubaa, Z.; Aissa, I.; Morjen, M.; Kharrat, N.; El Ayeb, M.; Gargouri, Y.; Srairi-Abid, N.; Marrakchi, N. Interaction of a snake venom l-amino acid oxidase with different cell types membrane. Int. J. Biol. Macromol. 2015, in press. Available online: http://dx.doi.org/10.1016/j.ijbiomac.2015.09.065 (accessed on 9 October 2015). [Google Scholar]
- Mariscal, R.N. Nematocysts. In Coelenterate Biology; Muscatine, L., Lenhoff, H.M., Eds.; Academic Press: New York, NY, USA, 1974; pp. 129–178. [Google Scholar]
- Rocha, J.; Peixe, L.; Gomes, N.C.M.; Calado, R. Cnidarians as a source of new marine bioactive compounds—An overview of the last decade and future steps for bioprospecting. Mar. Drugs 2011, 9, 1860–1886. [Google Scholar] [CrossRef] [PubMed]
- Purcell, J.E. Predation on zooplankton by large jellyfish, Aurelia labiata, Cyanea capillata and Aequorea aequorea, in Prince William Sound, Alaska. Mar. Ecol. Prog. Ser. 2003, 246, 137–152. [Google Scholar] [CrossRef]
- Lynam, C.P.; Gibbons, M.J.; Axelsen, B.E.; Sparks, C.A.; Coetzee, J.; Heywood, B.G.; Brierley, A.S. Jellyfish overtake fish in a heavily fished ecosystem. Curr. Biol. 2006, 16, 492–493. [Google Scholar] [CrossRef] [PubMed]
- Riisgård, H.U.; Madsen, C.V.; Barth-Jensen, C.; Purcell, J.E. Population dynamics and zooplankton-predation impact of the indigenous scyphozoan Aurelia aurita and the invasive ctenophore Mnemiopsis leidyi in Limfjorden (Denmark). Aquat. Invasion 2012, 7, 147–162. [Google Scholar] [CrossRef]
- Boero, F. Review of Jellyfish Blooms in the Mediterranean and Black Sea; No. 92; Food and Agriculture Organisation: Rome, Italy, 2013. [Google Scholar]
- Allavena, A.; Mariottini, G.L.; Carli, A.M.; Contini, S.; Martelli, A. In vitro evaluation of the cytotoxic, hemolytic and clastogenic activities of Rhizostoma pulmo toxin(s). Toxicon 1998, 36, 933–936. [Google Scholar] [CrossRef]
- Marino, A.; Morabito, R.; La Spada, G. Physiology of Nematocytes. In Proceedings of the 83th National Congress of the SIBS, Palermo, Italy, 24–25 October 2013. Abstract No. 36.
- Duarte, C.M.; Pitt, K.A.; Lucas, C.H.; Purcell, J.E.; Uye, S.-I.; Robinson, K.; Brotz, L.; Decker, M.B.; Sutherland, K.R.; Malej, A.; et al. Is global ocean sprawl a cause of jellyfish blooms? Front. Ecol. Environ. 2013, 11, 91–97. [Google Scholar] [CrossRef]
- De Donno, A.; Idolo, A.; Bagordo, F.; Grassi, T.; Leomanni, A.; Serio, F.; Guido, M.; Canitano, M.; Zampardi, S.; Boero, F.; et al. Impact of stinging jellyfish proliferations along South Italian coasts: Human health hazards, treatment and social costs. Int. J. Environ. Res. Public Health 2014, 11, 2488–2503. [Google Scholar] [CrossRef] [PubMed]
- Benedetti-Cecchi, L.; Canepa, A.M.; Fuentes, V.; Tamburello, L.; Purcell, J.E.; Piraino, S.; Roberts, J.; Boero, F.; Halpin, P. Deterministic factors overwhelm stochastic environmental fluctuations as drivers of jellyfish outbreaks. PLoS ONE 2015, 10, e0141060. [Google Scholar]
- Lassen, S.; Helmholz, H.; Ruhnau, C.; Prange, A. A novel proteinaceous cytotoxin from the northern Scyphozoa Cyanea capillata (L.) with structural homology to cubozoan haemolysins. Toxicon 2011, 57, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Morabito, R.; Condello, S.; Currò, M.; Marino, A.; Ientile, R.; La Spada, G. Oxidative stress induced by crude venom from the jellyfish Pelagia noctiluca in neuronal-like differentiated SH-SY5Y cells. Toxicol. Vitro 2012, 26, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Mariottini, G.L. Hemolytic venoms from marine cnidarian jellyfish—An overview. J. Venom Res. 2014, 5, 22–32. [Google Scholar] [PubMed]
- Mariottini, G.L.; Pane, L. Cytotoxic and cytolytic cnidarian venoms. A review on health implications and possible therapeutic applications. Toxins 2014, 6, 108–151. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.-H.P.; Rudloe, J. Potential of utilizing jellyfish as food in Western countries. Trends Food Sci. Technol. 1994, 5, 225–229. [Google Scholar] [CrossRef]
- Hsieh, Y.-H.P.; Leong, F.-M.; Rudloe, J. Jellyfish as food. Hydrobiologia 2001, 451, 11–17. [Google Scholar] [CrossRef]
- Gopal, R.; Vijayakumaran, M.; Venkatesan, R.; Kathiroli, S. Marine organisms in Indian medicine and their future prospects. Nat. Prod. Rad. 2008, 7, 139–145. [Google Scholar]
- De Zoysa, M. Medicinal benefits of marine invertebrates: Sources for discovering natural drug candidates. Adv. Food Nutr. Res. 2012, 65, 153–169. [Google Scholar] [PubMed]
- Kohl, A.C.; Kerr, R.G. Pseudopterosin biosynthesis: Aromatization of the diterpene cyclase product, Elisabethatriene. Mar. Drugs 2003, 1, 54–65. [Google Scholar] [CrossRef]
- Mariottini, G.L.; Pane, L. The role of Cnidaria in drug discovery. A review on CNS implications and new perspectives. Recent Pat. CNS Drug Discov. 2013, 8, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Bosch, T.C.G. The path less explored: Innate immune reactions in cnidarians. In Innate Immunity of Plants, Animals, and Humans. Nucleic Acids and Molecular Biology; Heine, H., Ed.; Springer-Verlag: Berlin, Germany; Heidelberg, Germany, 2008; pp. 27–42. [Google Scholar]
- Bosch, T.C.G.; Augustin, R.; Anton-Erxleben, F.; Fraune, S.; Hemmrich, G.; Zill, H.; Rosenstiel, P.; Jacobs, G.; Schreiber, S.; Leippe, M.; et al. Uncovering the evolutionary history of innate immunity: The simple metazoan Hydra uses epithelial cells for host defence. Dev. Comp. Immunol. 2009, 33, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Dingley, A.J.; Augustin, R.; Anton-Erxleben, F.; Stanisak, M.; Gelhaus, C.; Gutsmann, T.; Hammer, M.U.; Podschun, R.; Bonvin, A.M.J.J.; et al. Hydramacin-1, structure and antibacterial activity of a protein from the basal metazoan Hydra. J. Biol. Chem. 2009, 284, 1896–1905. [Google Scholar] [CrossRef] [PubMed]
- Augustin, R.; Anton-Erxleben, F.; Jungnickel, S.; Hemmrich, G.; Spudy, B.; Podschun, R.; Bosch, T.C.G. Activity of the novel peptide arminin against multiresistant human pathogens shows the considerable potential of phylogenetically ancient organisms as drug sources. Antimicrob. Agents Chemother. 2009, 53, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Augustin, R.; Siebert, S.; Bosch, T.C.G. Identification of a kazal-type serine protease inhibitor with potent anti-staphylococcal activity as part of Hydra’s innate immune system. Dev. Comp. Immunol. 2009, 33, 830–837. [Google Scholar] [CrossRef] [PubMed]
- Franzenburg, S.; Walter, J.; Künzel, S.; Wang, J.; Baines, J.F.; Bosch, T.C.G.; Fraune, S. Distinct antimicrobial peptide expression determines host species-specific bacterial associations. Proc. Natl. Acad. Sci. USA 2013, 110, E3730–E3738. [Google Scholar] [CrossRef] [PubMed]
- Kasahara, S.; Bosch, T.C.G. Enhanced antibacterial activity in Hydra polyps lacking nerve cells. Dev. Comp. Immunol. 2003, 27, 79–85. [Google Scholar] [CrossRef]
- Kim, K. Antimicrobial activity in gorgonian corals (Coelenterata: Octocorallia). Coral Reefs 1994, 13, 75–80. [Google Scholar] [CrossRef]
- Rodríguez, A.D.; Ramirez, C.; Rodríguez, I.I.; González, E. Novel antimycobacterial benzoxazole alkaloids, from the West Indian sea whip Pseudopterogorgia elisabethae. Org. Lett. 1999, 1, 527–530. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, I.I.; Rodríguez, A.D. Homopseudopteroxazole, a new antimycobacterial diterpene alkaloid from Pseudopterogorgia elisabethae. J. Nat. Prod. 2003, 66, 855–857. [Google Scholar] [CrossRef] [PubMed]
- McCulloch, M.W.B.; Haltli, B.; Marchbank, D.H.; Kerr, R.G. Evaluation of pseudopteroxazole and pseudopterosin derivatives against Mycobacterium tuberculosis and other pathogens. Mar. Drugs 2012, 10, 1711–1728. [Google Scholar] [CrossRef] [PubMed]
- Correa, H.; Aristizabal, F.; Duque, C.; Kerr, R. Cytotoxic and antimicrobial activity of pseudopterosins and secopseudopterosins isolated from the octocoral Pseudopterogorgia elisabethae of San Andrés and Providencia Islands (Southwest Caribbean Sea). Mar. Drugs 2011, 9, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Ata, A.; Win, H.Y.; Holt, D.; Holloway, P.; Segstro, E.P.; Jayatilake, G.S. New antibacterial diterpenes from Pseudopterogorgia elisabethae. Helv. Chim. Acta 2004, 87, 1090–1098. [Google Scholar] [CrossRef]
- Shapo, J.L.; Moeller, P.D.; Galloway, S.B. Antimicrobial activity in the common seawhip, Leptogorgia virgulata (Cnidaria: Gorgonaceae). Comp. Biochem. Physiol. B 2007, 148, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Tadesse, M.; Gulliksen, B.; Strøm, M.B.; Styrvold, O.B.; Haug, T. Screening for antibacterial and antifungal activities in marine benthic invertebrates from northern Norway. J. Invertebr. Pathol. 2008, 99, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Yu, S.; van Ofwegen, L.; Proksch, P.; Lin, W. Anthogorgienes A−O, new guaiazulene-derived terpenoids from a Chinese gorgonian Anthogorgia species, and their antifouling and antibiotic activities. J. Agric. Food Chem. 2012, 60, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; La, M.-P.; Tang, H.; Pan, W.-H.; Sun, P.; Krohn, K.; Yi, Y.-H.; Li, L.; Zhang, W. Bioactive briarane diterpenoids from the South China Sea gorgonian Dichotella gemmacea. Bioorg. Med. Chem. Lett. 2012, 22, 4368–4372. [Google Scholar] [CrossRef] [PubMed]
- Encarnación, D.R.; Franzblau, S.G.; Tapia, C.A.; Cedillo-Rivera, R. Screening of marine organisms for antimicrobial and antiprotozoal activity. Pharm. Biol. 2000, 38, 379–384. [Google Scholar] [CrossRef]
- El Sayed, K.A.; Bartyzel, P.; Shen, X.; Perry, T.L.; Zjawiony, J.K.; Hamann, M.T. Marine natural products as antituberculosis agents. Tetrahedron 2000, 56, 949–953. [Google Scholar] [CrossRef]
- Wei, X.; Nieves, K.; Rodríguez, A.D. Bioactive cubitane diterpenoids from a Colombian gorgonian species of the genus Eunicea. Pure Appl. Chem. 2012, 84, 1847–1855. [Google Scholar] [CrossRef]
- Cheng, S.-Y.; Chuang, C.-T.; Wang, S.-K.; Wen, Z.-H.; Chiou, S.-F.; Hsu, C.-H.; Dai, C.-F.; Duh, C.-Y. Antiviral and antiinflammatory diterpenoids from the soft coral Sinularia gyrosa. J. Nat. Prod. 2010, 73, 1184–1187. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Meng, L.-Y.; Tang, H.; Liu, B.-S.; Li, L.; Yi, Y.; Zhang, W. Sinularosides A and B, bioactive 9,11-secosteroidal glycosides from the South China Sea soft coral Sinularia humilis Ofwegen. J. Nat. Prod. 2012, 75, 1656–1659. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.-Y.; Huang, Y.-C.; Wen, Z.-H.; Chiou, S.-F.; Wang, S.-K.; Hsu, C.-H.; Dai, C.-F.; Duh, C.-Y. Novel sesquiterpenes and norergosterol from the soft corals Nephthea erecta and Nephthea chabroli. Tetrahedron Lett. 2009, 50, 802–806. [Google Scholar] [CrossRef]
- Badria, F.A.; Guirguis, A.N.; Perovic, S.; Steffen, R.; Müller, W.E.G.; Schröder, H.C. Sarcophytolide: A new neuroprotective compound from the soft coral Sarcophyton glaucum. Toxicology 1998, 131, 133–143. [Google Scholar] [CrossRef]
- Liang, L.-F.; Lan, L.-F.; Taglialatela-Scafati, O.; Guo, Y.-W. Sartrolides A-G and bissartrolide, new cembranolides from the South China Sea soft coral Sarcophyton trocheliophorum Marenzeller. Tetrahedron 2013, 69, 7381–7386. [Google Scholar] [CrossRef]
- Kelman, D.; Kashman, Y.; Rosenberg, E.; Kushmaro, A.; Loya, Y. Antimicrobial activity of Red Sea corals. Mar. Biol. 2006, 149, 357–363. [Google Scholar] [CrossRef]
- Koh, E.G.L. Do scleractinian corals engage in chemical warfare against microbes? J. Chem. Ecol. 1997, 23, 379–398. [Google Scholar] [CrossRef]
- Marquis, C.P.; Baird, A.H.; de Nys, R.; Holmström, C.; Koziumi, N. An evaluation of the antimicrobial properties of the eggs of 11 species of scleractinian corals. Coral Reefs 2005, 24, 248–253. [Google Scholar] [CrossRef]
- Barresi, G.; di Carlo, E.; Trapani, M.R.; Parisi, M.G.; Chille, C.; Mule, M.F.; Cammarata, M.; Palla, F. Marine organisms as source of bioactive molecules applied in restoration projects. Herit. Sci. 2015, 3. [Google Scholar] [CrossRef]
- Stabili, L.; Schirosi, R.; Parisi, M.G.; Piraino, S.; Cammarata, M. The mucus of Actinia equina (Anthozoa, Cnidaria): An unexplored resource for potential applicative purposes. Mar. Drugs 2015, 13, 5276–5296. [Google Scholar] [CrossRef] [PubMed]
- Fredrick, W.S.; Ravichandran, S. Anti microbial activity of the cnidarian blue button Porpita porpita (Linnaeus, 1758). Middle-East J. Sci. Res. 2010, 5, 355–358. [Google Scholar]
- ElAhwany, A.M.D.; Ghozlan, H.A.; ElSharif, H.A.; Sabry, S.A. Phylogenetic diversity and antimicrobial activity of marine bacteria associated with the soft coral Sarcophyton glaucum. J. Basic Microbiol. 2015, 55, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.-Y.; Yang, K.-L.; Li, J.; Wang, C.-Y.; Shao, C.-L. Phylogenetic diversity and antibacterial activity of culturable Fungi derived from the Zoanthid Palythoa haddoni in the South China Sea. Mar. Biotechnol. 2015, 17, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikova, T.V.; Balandin, S.V.; Aleshina, G.M.; Tagaev, A.A.; Leonova, Y.F.; Krasnodembsky, E.D.; Men’shenin, A.V.; Kokryakov, V.N. Aurelin, a novel antimicrobial peptide from jellyfish Aurelia aurita with structural features of defensins and channel-blocking toxins. Biochem. Biophys. Res. Commun. 2006, 348, 514–523. [Google Scholar] [CrossRef] [PubMed]
- Shenkarev, Z.O.; Panteleev, P.V.; Balandin, S.V.; Gizatullina, A.K.; Altukhov, D.A.; Finkina, E.I.; Kokryakov, V.N.; Arseniev, A.S.; Ovchinnikova, T.V. Recombinant expression and solution structure of antimicrobial peptide aurelin from jellyfish Aurelia aurita. Biochem. Biophys. Res. Commun. 2012, 429, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Grant, S.; Gisondi, A.; Hortano, W.; Defilippo, J.; Beck, G. Isolation and preliminary characterization of antimicrobial proteins and peptides from Ctenophores and Cnidaria. In Comprehensive Bioactive Natural Products, Vol 6, Extraction, Isolation & Characterization; Gupta, V.K., Taneja, S.C., Gupta, B.D., Eds.; Studium Press LLC: New Delhi, India, 2010; pp. 258–279. [Google Scholar]
- Suganthi, K.; Bragadeeswaran, S. Antimicrobial and immunomodulatory activities of jellyfish (Chrysaora quinquecirrha) venom. In Prospects in Bioscience: Addressing the Issues; Sabu, A., Augustine, A., Eds.; Springer: New Delhi, India, 2013; pp. 283–292. [Google Scholar]
- Morales-Landa, J.L.; Zapata-Pérez, O.; Cedillo-Rivera, R.; Segura-Puertas, L.; Simá-Alvarez, R.; Sánchez-Rodríguez, J. Antimicrobial, antiprotozoal, and toxic activities of cnidarian extracts from the Mexican Caribbean Sea. Pharm. Biol. 2007, 45, 37–43. [Google Scholar] [CrossRef]
- Yeh, T.-T.; Wang, S.-K.; Dai, C.-F.; Duh, C.-Y. Briacavatolides A–C, new briaranes from the Taiwanese octocoral Briareum excavatum. Mar. Drugs 2012, 10, 1019–1026. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-K.; Yeh, T.-T.; Duh, C.-Y. Briacavatolides D–F, new briaranes from the Taiwanese octocoral Briareum excavatum. Mar. Drugs 2012, 10, 2103–2110. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.-J.; Lee, Y.-S.; Wang, S.-K.; Sheu, J.-H.; Duh, C.-Y. Parathyrsoidins A–D, four new sesquiterpenoids from the Soft Coral Paralemnalia thyrsoides. Mar. Drugs 2013, 11, 2501–2509. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.K.; Hsieh, M.K.; Duh, C.Y. Three new cembranoids from the Taiwanese soft coral Sarcophyton ehrenbergi. Mar. Drugs 2012, 10, 1433–1444. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-K.; Hsieh, M.-K.; Duh, C.-Y. New diterpenoids from soft coral Sarcophyton ehrenbergi. Mar. Drugs 2013, 11, 4318–4327. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.; Shao, C.-L.; Chen, M.; Zhang, M.-Q.; Xu, K.-X.; Meng, H.; Wang, C.-Y. Antiviral C-25 epimers of 26-acetoxy steroids from the South China Sea gorgonian Echinogorgia rebekka. J. Nat. Prod. 2014, 77, 1488–1493. [Google Scholar] [CrossRef] [PubMed]
- Lillsunde, K.-E.; Festa, C.; Adel, H.; De Marino, S.; Lombardi, V.; Tilvi, S.; Nawrot, D.A.; Zampella, A.; D’Souza, L.; D’Auria, M.V.; et al. Bioactive cembrane derivatives from the Indian Ocean soft coral, Sinularia kavarattiensis. Mar. Drugs 2014, 12, 4045–4068. [Google Scholar] [CrossRef] [PubMed]
- Tejuca, M.; Anderluh, G.; Maček, P.; Marcet, R.; Torres, D.; Sarracent, J.; Alvarez, C.; Lanio, M.E.; Dalla Serra, M.; Menestrina, G. Antiparasite activity of sea anemone cytolysins on Giardia duodenalis and specific targeting with anti-Giardia antibodies. Int. J. Parasitol. 1999, 29, 489–498. [Google Scholar] [CrossRef]
- Reimão, J.Q.; Migotto, A.E.; Kossuga, M.H.; Berlinck, R.G.S.; Tempo, A.G. Antiprotozoan activity of Brazilian marine cnidarian extracts and of a modified steroid from the octocoral Carijoa riisei. Parasitol. Res. 2008, 103, 1445–1450. [Google Scholar] [CrossRef] [PubMed]
- Ishigami, S.-T.; Goto, Y.; Inoue, N.; Kawazu, S.-I.; Matsumoto, Y.; Imahara, Y.; Tarumi, M.; Nakai, H.; Fusetani, N.; Nakao, Y. Cristaxenicin A, an antiprotozoal xenicane diterpenoid from the deep sea gorgonian Acanthoprimnoa cristata. J. Org. Chem. 2012, 77, 10962–10966. [Google Scholar] [CrossRef] [PubMed]
- Tenover, F.C. Mechanisms of antimicrobial resistance in bacteria. AJIC 2006, 34 (Suppl. S1), S3–S10. [Google Scholar]
- Walsh, T.R. Combinatorial genetic evolution of multiresistance. Curr. Opin. Microbiol. 2006, 9, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. Resisting resistance: New chemical strategies for battling superbugs. Chem. Biol. 2000, 7, R127–R132. [Google Scholar] [CrossRef]
- Daly, N.L.; Seymour, J.; Wilson, D. Exploring the therapeutic potential of jellyfish venom. Future Med. Chem. 2014, 6, 1715–1724. [Google Scholar] [CrossRef] [PubMed]
- Yanagihara, A.A.; Shohet, R.V. Cubozoan venom-induced cardiovascular collapse is caused by hyperkalemia and prevented by zinc gluconate in mice. PLoS ONE 2012, 7, e51368. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, D.L.; Burnell, J.N. Biochemical and molecular characterisation of cubozoan protein toxins. Toxicon 2009, 54, 1162–1173. [Google Scholar] [CrossRef] [PubMed]
- Mariottini, G.L.; Bonello, G.; Giacco, E.; Pane, L. Neurotoxic and neuroactive compounds from Cnidaria: Five decades of research and more. CNS Agents Med. Chem. 2015, 15, 74–80. [Google Scholar]
- Ganz, T. Defensins: Antimicrobial peptides of innate immunity. Nat. Rev. Immunol. 2003, 3, 710–720. [Google Scholar] [CrossRef] [PubMed]

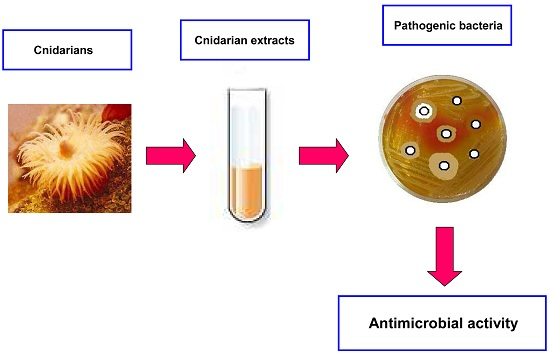{kind=link}
| Species | Compound | Sensitive Microbes | Activity | Reference | |
|---|---|---|---|---|---|
| MBC (μM) | LD90 (μM) | ||||
| Hydra magnipapillata | Hydramacin-1 (peptide) | Citrobacter freundii | 0.9 | 0.2 | [66] |
| Citrobacter freundii | 4.0 ± 4.4 | 0.7 ± 0.3 | [67] | ||
| Enterobacter cloacae | 1.8 | 0.4 | [66] | ||
| Enterobacter cloacae (1) | 7.6 ± 9.5 | 0.7 ± 0.3 | [67] | ||
| Escherichia coli (10) | 1.5 ± 2.3 | 0.3 ± 0.1 | [66] | ||
| Escherichia coli (2) | 2.25 ± 1.9 | 0.55 ± 0.5 | [67] | ||
| Salmonella typhimurium | 0.9 | 0.2 | [66] | ||
| Salmonella typhimurium (1) | 0.9 ± 0 | 0.5 ± 0 | [67] | ||
| Yersinia enterocolitica | 0.4 | 0.2 | [66] | ||
| Yersinia enterocolitica | 0.9 | 0.2 | [67] | ||
| Klebsiella oxytoca (7) | 0.9 ± 0 | 0.35 ± 0.1 | [66] | ||
| Klebsiella oxytoca (4) | 3.9 ± 3.1 | 0.8 ± 0.2 | [67] | ||
| Klebsiella pneumoniae (8) | 1.8 ± 1.3 | 0.5 ± 0.25 | [66] | ||
| Klebsiella pneumoniae (2) | 2.25 ± 1.9 | 0.7 ± 0.3 | [67] | ||
| Acinetobacter baumannii | 7.1 | 1.8 | [67] | ||
| Proteus mirabilis | 14.3 | 0.9 | [67] | ||
| Proteus vulgaris | >14.3 | 3.6 | [67] | ||
| Pseudomonas aeruginosa | >14.3 | >14.3 | [66] | ||
| Serratia marcescens (3) | >14.3 | 6.6 ± 6.7 | [67] | ||
| Bacillus megaterium | 0.2 | 0.1 | [66] | ||
| Enterococcus faecalis | 14.3 | 0.9 | [66] | ||
| Enterococcus faecalis (10) | >14.3 | 7.1 | [67] | ||
| Staphylococcus aureus | >14.3 | 14.3 | [66] | ||
| Staphylococcus hominis | >14.3 | 3.6 | [66] | ||
| Staphylococcus hemolyticus | 1.8 | 0.9 | [67] | ||
| Streptococcus pyogenes | >14.3 | 7.1 | [67] | ||
| Periculin-1 (recombinant peptide) | Bacillus megaterium | Not given | 0.2–0.4 | [66] | |
| c-Arminin 1a (recombinant peptide) | ESBL-producing K. pneumoniae (3) | 0.5 ± 0.2 | 0.2 ± 0 | [68] | |
| Escherichia coli (5) | 0.3 ± 0.1 | 0.08 ± 0.03 | [68] | ||
| Bacillus megaterium | 0.1 | 0.01 | [68] | ||
| Staphylococcus aureus (9) | 0.5 ± 0.2 | 0.17 ± 0.06 | [68] | ||
| Enterococcus faecalis (VRE) | 1.6 | 0.8 | [68] | ||
| Enterococcus faecium (VRE) (6) | 0.5 ± 0.2 | 0.2 ± 0 | [68] | ||
| Kazal-2 (native protein) | Staphylococcus aureus | 0.7–0.8 | - | [69] | |
| Kazal-2 (recombinant protein) (*) | Staphylococcus aureus | 35.7 ± 2.5 | - | [69] | |
| Hydra spp. | Arminin (peptide) | Escherichia coli | 1.823 ± 1.19 (^) | [70] | |
| Species | Compound | Sensitive Microbes | MIC (μg/mL) | Reference |
|---|---|---|---|---|
| Pseudopterogorgia elisabethae | Pseudopteroxazole (diterpenoid alkaloid) | Mycobacterium tuberculosis H37Rv | 15 | [73,75] |
| Pseudopteroxazole | Mycobacterium tuberculosis H37Rv | 15 (*) | [75] | |
| (Ptx-CH2-(1H-imidazol-5-yl)) | Mycobacterium tuberculosis H37Rv | 13 (*) | [75] | |
| Mycobacterium smegmatis | 4 | |||
| Mycobacterium diernhoferi | 2 | |||
| iso-Ptx-H | Mycobacterium tuberculosis H37Rv | 14 (*) | [75] | |
| Ptx-CH3 | Mycobacterium tuberculosis H37Rv | 15 (*) | [75] | |
| Pseudopterosin P | Streptococcus pyogenes | 0.8 | [77] | |
| Staphylococcus aureus | 2.0 | |||
| Enterococcus faecalis | 3.5 | |||
| Pseudopterosin Q | Streptococcus pyogenes | 1.0 | [77] | |
| Staphylococcus aureus | 2.3 | |||
| Enterococcus faecalis | 3.6 | |||
| Homopseudopteroxazole (diterpene alkaloid) | Mycobacterium tuberculosis H37Rv | 12.5 | [74] | |
| Nephthea sp. | Litosterol | Mycobacterium tuberculosis | 3.13 | [83] |
| Nephalsterol C | Mycobacterium tuberculosis | 12.5 | [83] | |
| Sarcophyton glaucum | Sarcophytolide (lactone cembrane diterpene) | Staphylococcus aureus | 0.19 | [88] |
| Pseudomonas aeruginosa | 0.22 | |||
| Saccharomyces cerevisiae | 0.13 | |||
| Sarcophyton trocheliophorum | Cembranolide | Staphylococcus aureus | 125 | [89] |
| Xenia macrospiculata | Desoxyhavannahine | Marine bacteria | 48 | [90] |
| Aurelia aurita (^) | Aurelin (peptide) | Listeria monocytogenes | 5.27 | [98,99] |
| Escherichia coli | 1.78 | |||
| Bacillus megaterium | 10 | |||
| Micrococcus luteus | 40 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mariottini, G.L.; Grice, I.D. Antimicrobials from Cnidarians. A New Perspective for Anti-Infective Therapy? Mar. Drugs 2016, 14, 48. https://doi.org/10.3390/md14030048
Mariottini GL, Grice ID. Antimicrobials from Cnidarians. A New Perspective for Anti-Infective Therapy? Marine Drugs. 2016; 14(3):48. https://doi.org/10.3390/md14030048
Chicago/Turabian StyleMariottini, Gian Luigi, and Irwin Darren Grice. 2016. "Antimicrobials from Cnidarians. A New Perspective for Anti-Infective Therapy?" Marine Drugs 14, no. 3: 48. https://doi.org/10.3390/md14030048
APA StyleMariottini, G. L., & Grice, I. D. (2016). Antimicrobials from Cnidarians. A New Perspective for Anti-Infective Therapy? Marine Drugs, 14(3), 48. https://doi.org/10.3390/md14030048