
Porifera Lectins: Diversity, Physiological Roles and Biotechnological Potential



 ,
, 
Abstract
:1. Introduction
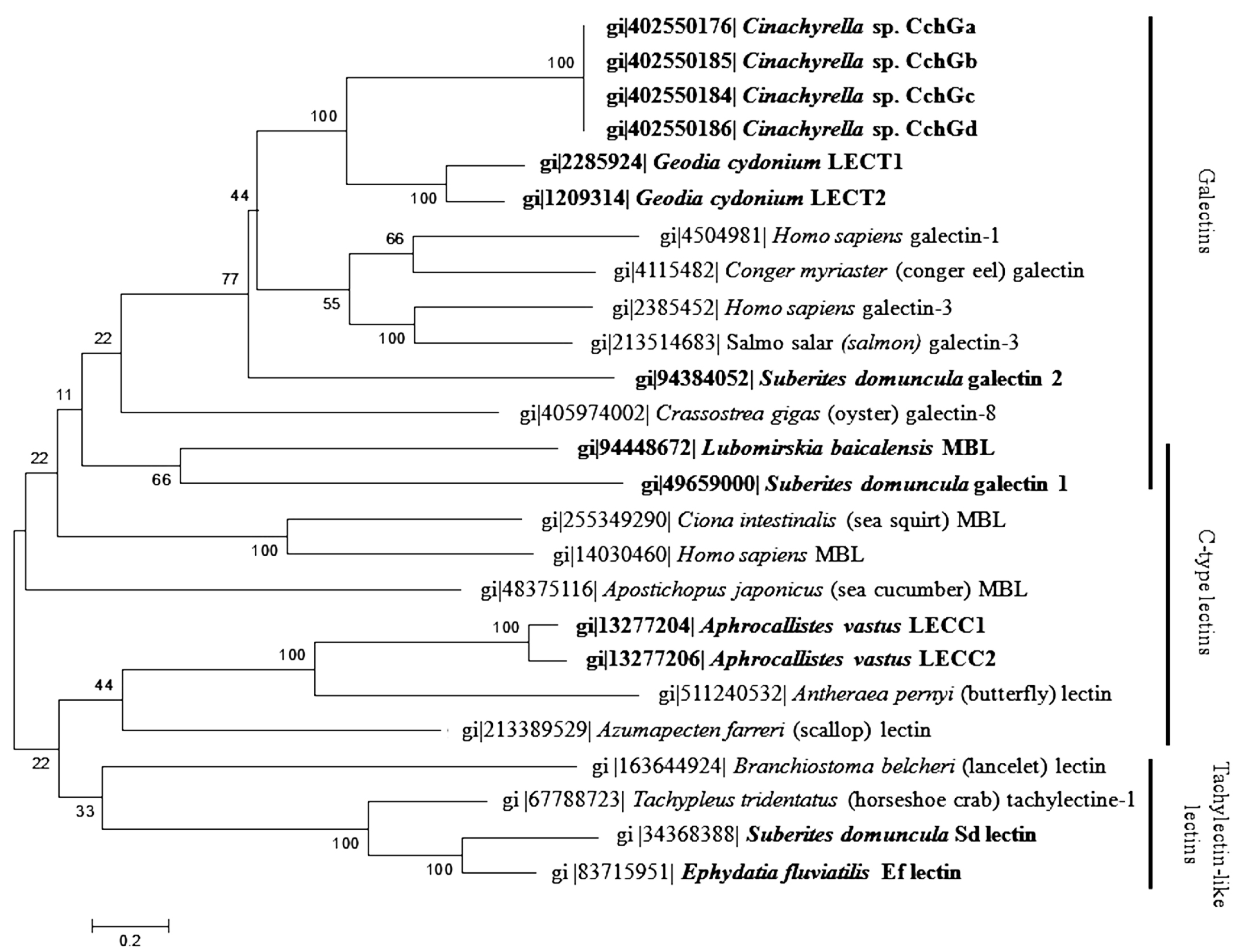
2. Classification

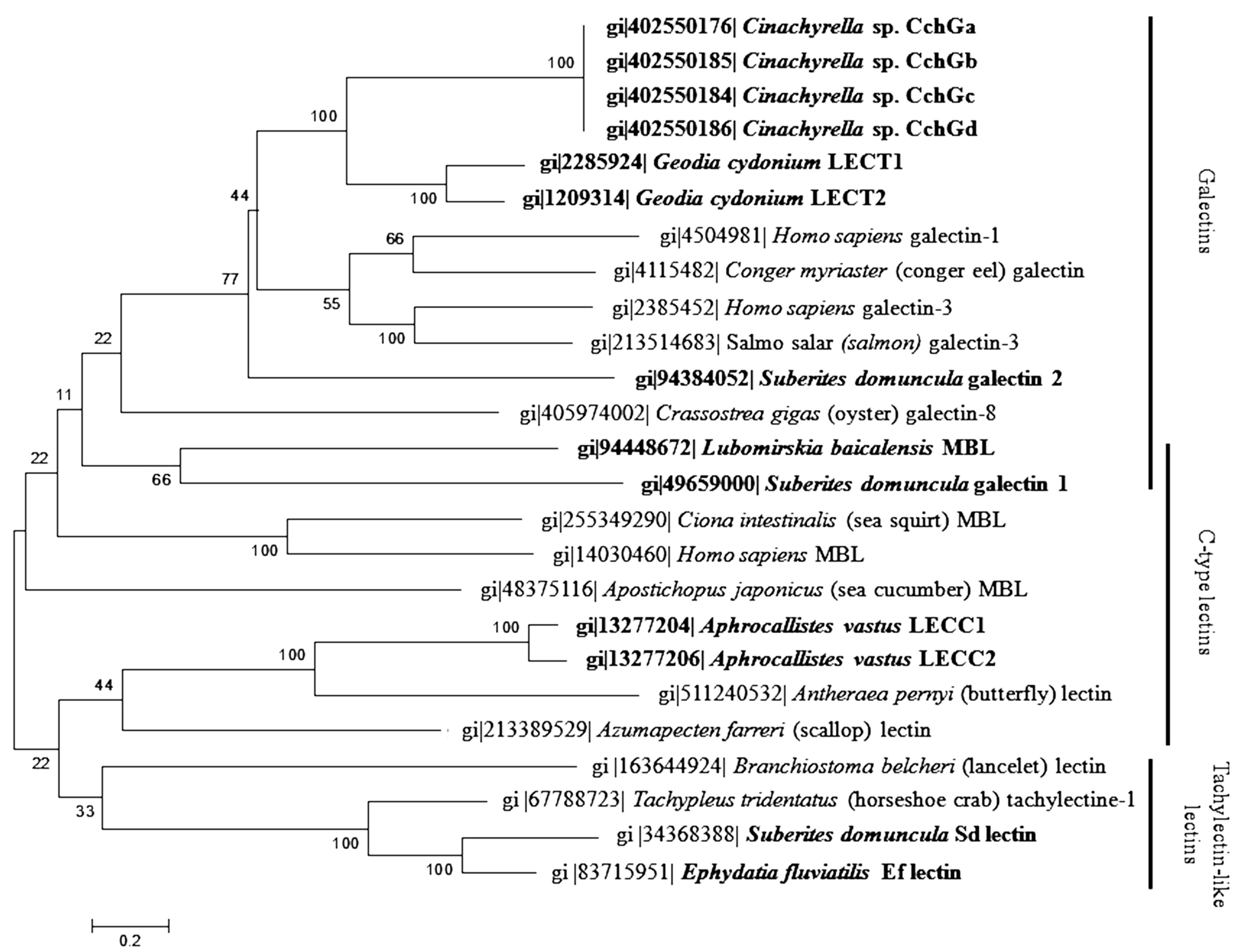{kind=link}
{kind=link}
{kind=link}
| Name | Species | Order (Class) | Size in kDa (Subunits) | Carbohydrates | pI | Cations | Disulfide Bridges | pH/T °C Activity |
|---|---|---|---|---|---|---|---|---|
| Sponge galectins | ||||||||
| CchG 1 | Cinachyrella sp. | Spirophorida(D) | 50.0 (4) | galactoside residues | nd | no | no | nd/<100 °C |
| CchG 2 | Cinachyrella sp. | Spirophorida (D) | 50.0 (4) | galactoside residues | nd | no | no | nd/<100 °C |
| GCG | Geodia cydonium | Astrophorida (D) | 60.0 (4) | galactoside residues | 4.4 | Ca2+ | no | nd/nd |
| HoL-30 | Halichondria okadai | Halichondrida (D) | 60.0 (2) | galactoside residues | 6.7 | no | no | nd/nd |
| Sd galectin 1 | Suberites domuncula | Hadromerida (D) | 22.1 (nd) | galactoside residues | nd | no | nd | nd/nd |
| Sd galectin 2 | Suberites domuncula | Hadromerida (D) | 35.0 (nd) | galactose | nd | Ca2+ | no | nd/nd |
| Sponge C-type lectins | ||||||||
| AaL | Aplysina archeri | Verongida (D) | 63.0 (4) | non reducing galactoside residues | nd | Ca2+/Mg2+ | no | nd/nd |
| AlL | Aplysina lacunosa | Verongida (D) | 63.0 (4) | non reducing galactoside residues | nd | Ca2+/Mg2+ | no | nd/nd |
| AvL | Aphrocallistes vastus | Hexactinosida (H) | 34.0 (1) | galactoside residues | nd | Ca2+ | no | nd/nd |
| CvL | Cliona varians | Hadromerida (D) | 114.0 (4) | galactose/sucrose | nd | Ca2+ | yes | 6.0–8.0/<60 °C |
| Lb MBL | Lubomirskia baicalensis | Haplosclerida (D) | 13 | mannose | nd | nd | nd | nd/nd |
| PsL | Pellina semitubulosa | Halichondrida (D) | 200.0 (6) | galactose/arabinose | 6.1 | Ca2+ | no | nd/nd |
| Sponge tachylectine-like lectins | ||||||||
| Ef lectin | Ephydatia fluviatilis | Haplosclerida (D) | 24.0 (1) | nd | nd | no | no | nd/nd |
| Sd lectin | Suberites domuncula | Hadromerida (D) | 27.0 (1) | lipopolysaccharides | nd | no | no | nd/nd |
| Sponge F-type lectin | ||||||||
| CcL | Crambe crambe | Poecilosclerida (D) | 14.0 (1) | fucose | nd | no | no | nd/nd |
| Unclassified sponge lectins | ||||||||
| AcL I | Axinella corrugata | Halichondrida (D) | 78.5 (6) | N-acetyl-derived residues | 6.3 | no | yes | 6.5–8.5/<65 °C |
| AcL II | Axinella corrugata | Halichondrida (D) | 80 | galatose/chitin/fetuin/N-acetyl-derived residues | nd | nd | nd | 2.0–6.0/<65 °C |
| ApaL I | Aaptos papillata | Hadromerida (D) | 21.0 (2) | N-acetyl-d-glucosamine | 42,131 | nd | no | nd/nd |
| ApaL II | Aaptos papillata | Hadromerida (D) | 16.0 (1) | N-acetyl-d-glucosamine/N-acetyl-d-galactosamine/sialic acid residues | 3.4/5 | nd | no | nd/nd |
| ApaL III | Aaptos papillata | Hadromerida (D) | 16.0 (1) | N-acetyl-d-glucosamine/N-acetyl-d-galactosamine/sialic acid residues | 3.4/5 | nd | no | nd/nd |
| ApL I | Axinella polypoides | Halichondrida (D) | nd (2) | galactoside residues | 3.9 | no | yes1 | nd/nd |
| ApL II | Axinella polypoides | Halichondrida (D) | nd | galactoside residues | nd | nd | nd | nd/nd |
| ApL III | Axinella polypoides | Halichondrida (D) | nd | galactoside residues | nd | nd | nd | nd/nd |
| ApL IV | Axinella polypoides | Halichondrida (D) | nd | hexuronic acids | nd | nd | nd | nd/nd |
| ApL V | Axinella polypoides | Halichondrida (D) | nd | galactoside residues | nd | nd | nd | nd/nd |
| CaL | Cinachyrella apion | Spirophorida (D) | 124.0 (8) | lactose | nd | no | no | nd/nd |
| CalL | Cinachyrella alloclada | Spirophorida (D) | nd | galactoside residues | nd | nd | yes | nd/nd |
| CauL | Craniella australiensis | Spirophorida (D) | 54.0 (3) | porcine stomach mucin/asialo-porcine stomach mucin | nd | no | yes | 5.0–8.0/20–70 °C |
| CnL | Chondrilla nucula | Chondrosida (D) | 70.0 (4) | galactose | nd | no | no | 4.5–8.5/<60 °C |
| CtL | Cinachyrella tenuiviolacea | Spiroporida (D) | 22 | lactose | nd | nd | nd | nd/nd |
| DaL | Desmapsamma anchorata | nd | nd (2) | galactose | nd | nd | nd | nd/nd |
| Halilectin 1 (H-1) | Haliclona caerulea | Haplosclerida (D) | 15.0 (1) | nd | nd | no | no | 9/<70 °C |
| Halilectin 2 (H-2) | Haliclona caerulea | Haplosclerida (D) | 27.0 (2) | porcine stomach mucin | nd | no | no | 4.0–5.0/<80 °C |
| Halilectin 3 (H-3) | Haliclona caerulea | Haplosclerida (D) | nd (4) | porcine stomach mucin/N-acetyl-galactosamine | nd | no | no | 4.0–7.0/<70 °C |
| HcL | Haliclona cratera | Haplosclerida (D) | nd | orosomucoid | 8.6 | no | no | 4.6–10.2/56 °C |
| HL | Haliclona sp. | Haplosclerida (D) | 24.0 (nd) | galactose/lactose | nd | nd | nd | nd/nd |
| HoL-1 | Halichondria okadai | Halichondrida (D) | 84.0 (4) | N-acetyl groups of N-acetyl-d-glucosamine or N-acetyl-d-galactosamine | 4.5 | no | yes 1 | nd/<50 °C |
| HoL-2 | Halichondria okadai | Halichondrida (D) | 42.0 (1) | galactoside β1-4N-acetyl-d-glucosamine units | 4.5 | no | no | nd/<40 °C |
| HpL | Halichondria panicea | Halichondrida (D) | 78.0 (4) | fetuin/galacturonic acid/glucuronic acid/polygalacturonic acid/fucose | nd | no | yes 1 | 7.2–9.5/<30 °C |
3. Sponge Lectin Purification, Sequencing and Expression
3.1. Purification Strategies
| Species | Material | Extraction Buffer | Type I Purification | Column | Elution Buffer I | Type II Purification | Column | Elution Buffer II | |
|---|---|---|---|---|---|---|---|---|---|
| Aphrocallites vastus | 100 g (w) frozen | calcium- and magnesium-free seawater Ca2+ precipitation | gel filtration | Biogel P300 column | Calcium Magnesium free sea water | centrifugation on sucrose gradient | |||
| Axinella corrugata | water/PBS pH 7.2 | affinity chromatography | rabbit stroma-polyacrilamide gel | 0.035 M NH4OH/0.154 M NaCl | gel filtration | Ultrogel—AcA 44 | PBS | ||
| Cinachyrella apion | 1:2 (w/v) in 0.05 M Tris-HCl pH 7.5 acetone precipitation | immunoaffinity chromatography | IgG anti CvL-Sepharose | 0.050 M Tris-HCl pH 11 | gel filtration | Superose 6 10/300 | 0.05 M Tris-HCl pH 7.5 | ||
| Cliona varians | 1:2 (w/v) in 0.05 M Tris-HCl pH 7.5 acetone precipitation | affinity chromatography | Sepharose CL 4B | 0.05 M Tris-HCl/0.1 M EDTA pH 8 | ion exchange | CM cellulose column | acetate buffered saline | ||
| Craniella australiensis | 23 g (w) | 1:10 in 0.9% NaCl dialysis against water | ion exchange | DEAE-Sephacel | gradient of NaCl in 0.010 Tris-HCL pH 7.4 | gel filtration | Sephadex G-150 | 0.1 M PBS pH 7.4 | |
| Geodia cydonium | calcium- and magnesium-free sea water | affinity chromatography | lactose-divinylsulfone-agarose | PBS/0.05% Tween 20 | precipitation with carbohydrates | ||||
| Haliclona caerulea | 1:10 (w/v) in deionized water | affinity chromatography | glutaraldehyde-fixed human erythrocyte stroma-Sephadex G25 | TBS/0.3% NH4OH pH 8.5 | gel filtration | ||||
| Haliclona cratera | 150 g (w) | TBS pH 7.5 polyvinylpolypyrrolidone/+protease inhibitors | affinity chromatography | CM Sepharose 4B CL | 0.020 M phosphate buffer/1 M NaCl pH 7.5 | gel filtration | Bio Gel P-100 | TBS/152 mM NaCl | |
| Halichondria okadai | 200 g (w) frozen | 1:10 (w/v) TBS/0.15 M NaCl/0.01 M protease inhibitor mix pH 7.4 | affinity chromatography | lactosyl agarose | TBS/0.1 M lactose pH 7.4 | gel filtration | Sephadex 75 | TBS pH 7.4 | |
| Halichondria panicea | 30 g (w) | 1:3 (w/v) in calcium- and magnesium-free sea water/0.25 M EDTA/0.01 M 2-mercaptoethanol | affinity chromatography | Sepharose 4B | |||||
| Pellina semitubulosa | 240 g (w) | 1:1 (w/v) in 0.03 M Tris-HCl/2 mM CaCl2 pH 8.5 | affinity chromatography | acid-treated Sepharose 6B | 0.03 M Tris-HCl/0.002 M CaCl2/0.1 M lactose pH 7.5 | gel filtration | Sephadex G-200 | 0.03 M Tris-HCl pH 7.5 | |
3.2. Genome Screening and Lectin Production Strategies
4. Biochemical Properties of Sponge Lectins
4.1. Galectins
4.2. C-Type Lectins
4.3. Tachylectin-Like Lectins
4.4. F-Type Lectin
4.5. Unclassified Sponge Lectins
4.5.1. Intrachain Disulfide Bridge-Containing Lectins
4.5.2. Mucin-Binding Lectins
4.5.3. N-Acetyl-d-glucosamine-/N-Acetyl-d-galactosamine-Binding Lectins
4.5.4. Miscellaneous
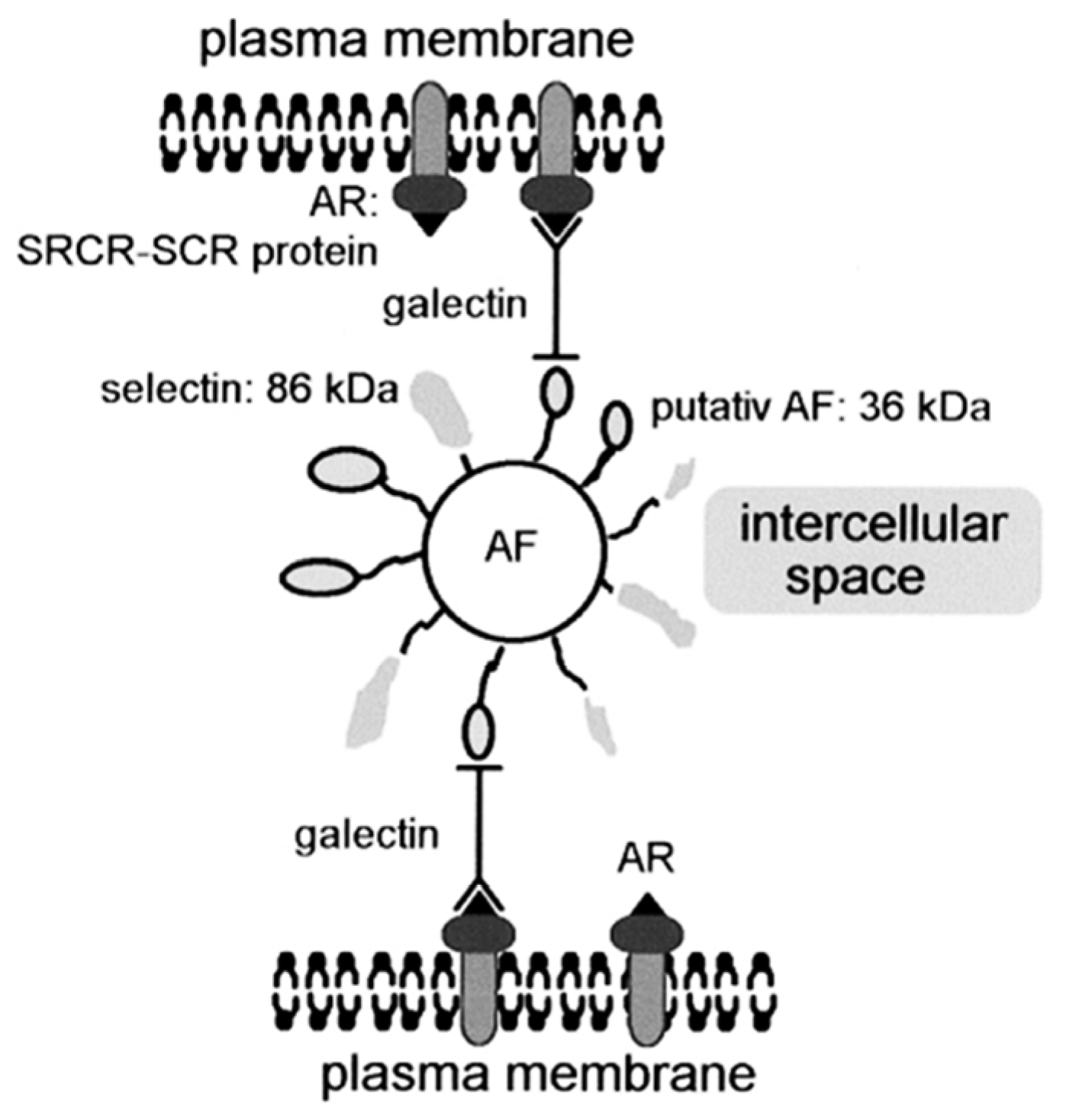
5. Physiological Roles of Sponge Lectins
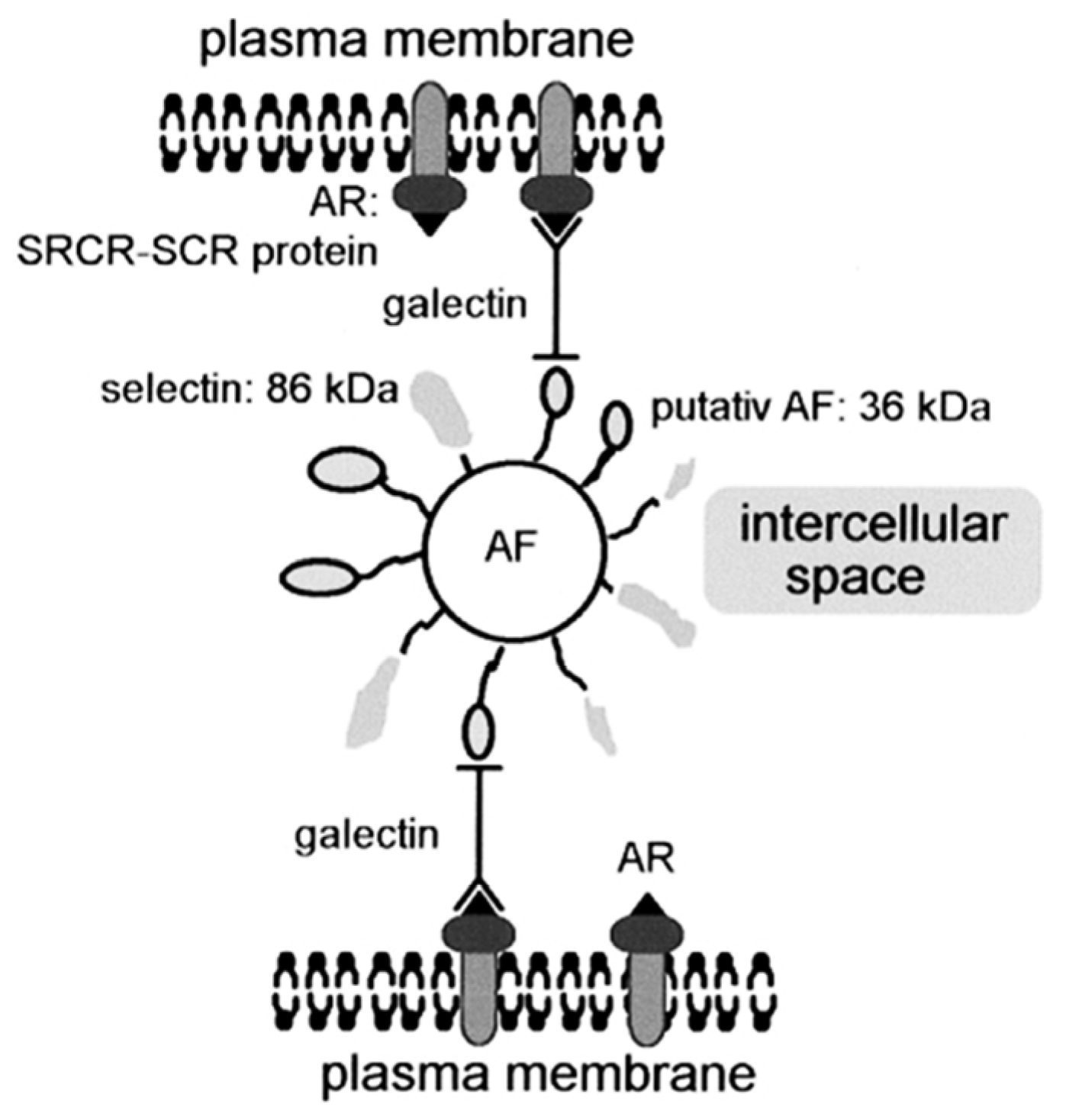
5.1. Morphogenesis and Cell Interaction
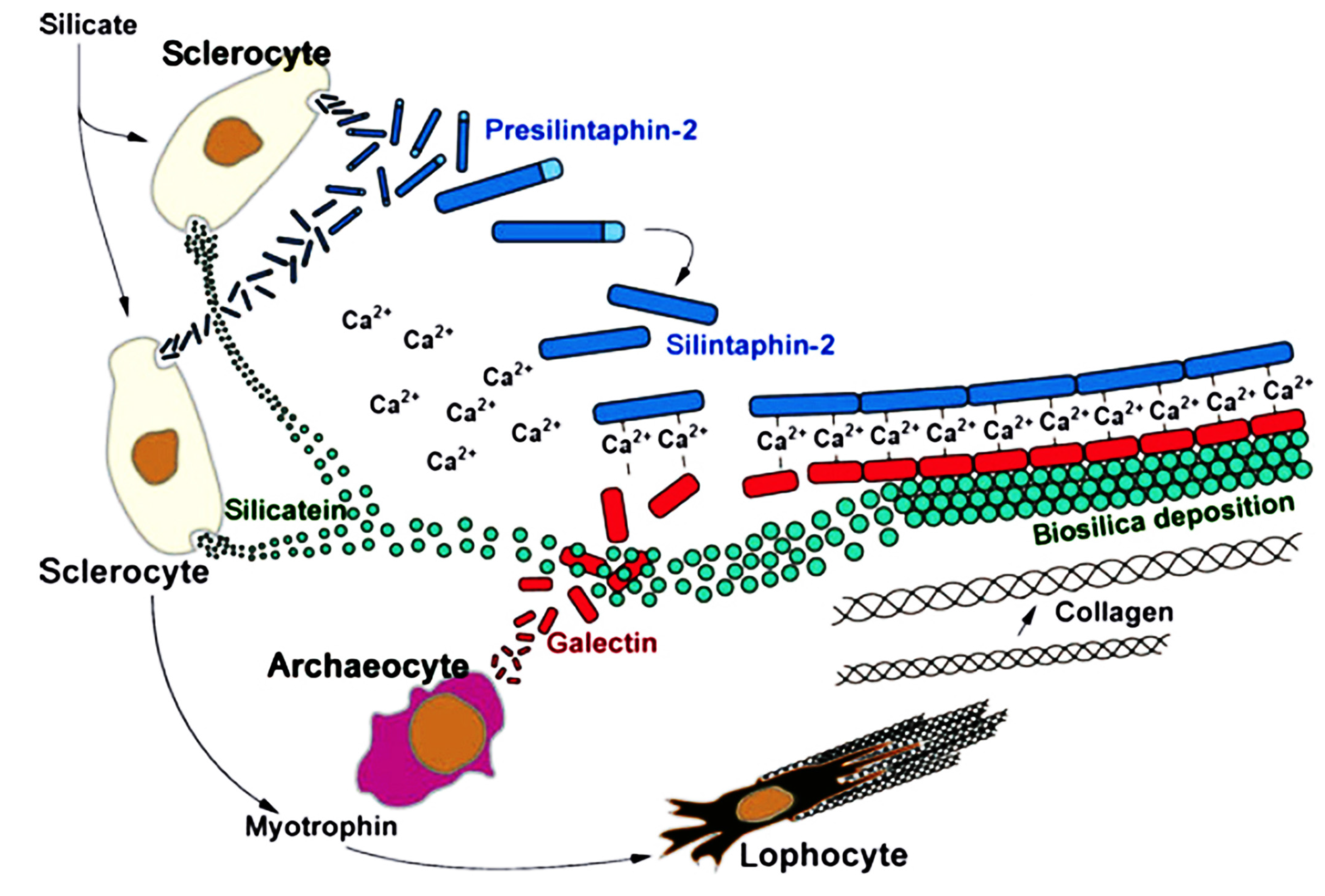
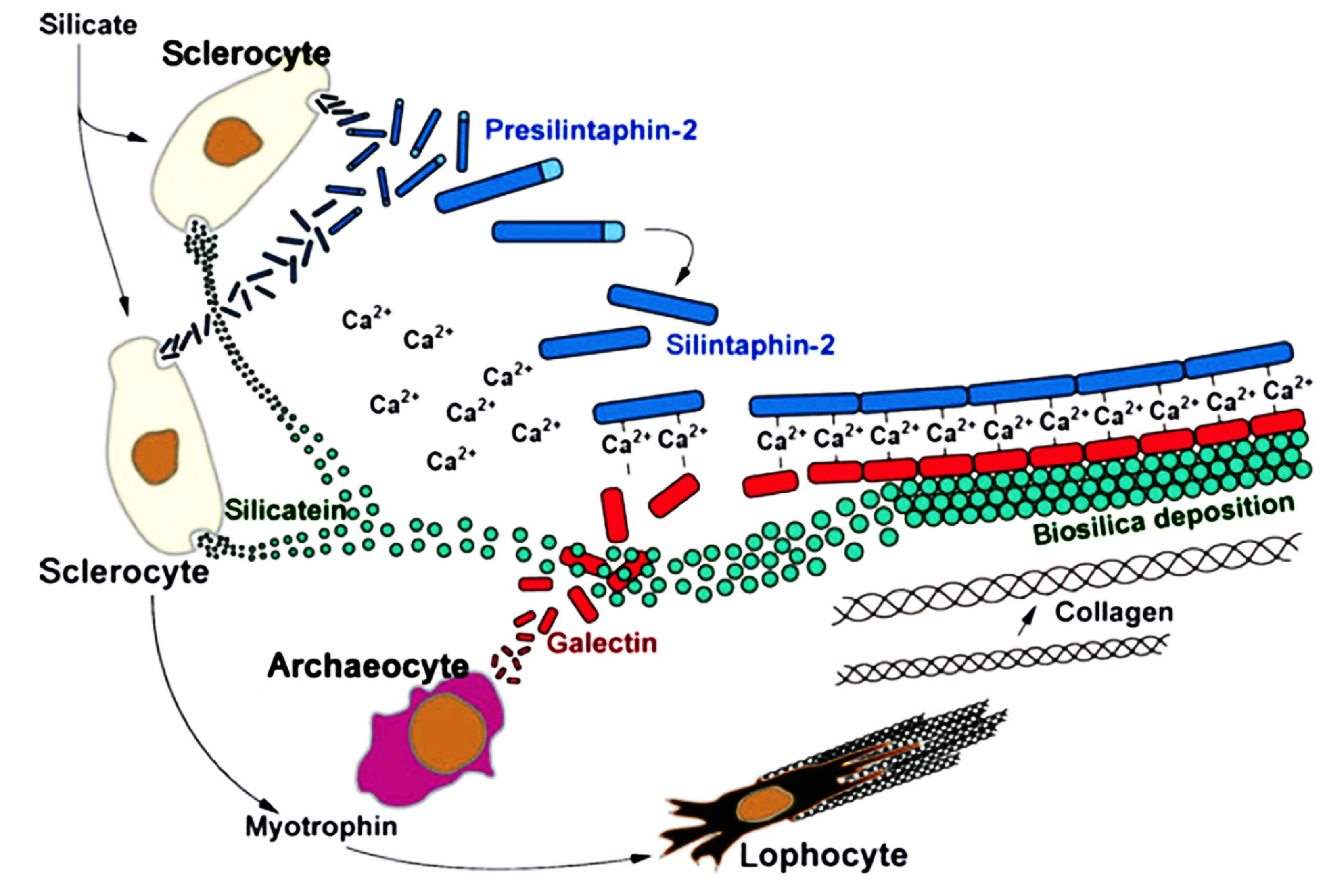
5.2. Biomineralization and Spiculogenesis
5.3. Host Defense
5.4. Association with Microorganisms
6. Biotechnological Potential
6.1. Activities on Mammalian Immune Cells
6.2. Antimicrobial Activities
| Name | Species | Order (Class) | Biological Activities | Physiological Roles | References |
|---|---|---|---|---|---|
| Sponge galectins | |||||
| CchG 1 | Cinachyrella sp. | Spirophorida (D) | rabbit erythrocyte agglutination modulatory activity of human glutamate receptors | nd | [65] |
| CchG 2 | Cinachyrella sp. | Spirophorida (D) | rabbit erythrocyte agglutination | nd | [65] |
| GCG | Geodia cydonium | Astrophorida (D) | modulatory activity of human glutamate receptors increase of the growth rate of L5178y mouse lymphoma cells mitotic activity on human lymphocytes | cell interaction | [68,69,70,71,72] |
| HoL-30 | Halichondria okadai | Halichondrida (D) | rabbit and human erythrocyte agglutination | nd | [62] |
| Sd galectin 1 | Suberites domuncula | Hadromerida (D) | nd | canal system formation in primmorphs | [51] |
| Sd galectin 2 | Suberites domuncula | Hadromerida (D) | nd | biomineralization/spiculogenesis | [67] |
| Sponge C-type lectins | |||||
| AaL | Aplysina archeri | Verongida (D) | hamster, rabbit, bovine and human erythrocyte agglutination | nd | [77] |
| AlL | Aplysina lacunosa | Verongida (D) | hamster, rabbit, bovine and human erythrocyte agglutination | nd | [77] |
| AvL | Aphrocallistes vastus | Hexactinosida (H) | nd | cell interaction | [56] |
| CvL | Cliona varians | Hadromerida (D) | human erythrocte agglutination antibacterial activity against B. subtilis and S. aureus no activity against E. coli and P. aeruginosa Leishmania chagasi agglutination chemotactic on mouse neutrophils in vivo cytotoxic activity against K562 and Jurkat cells no cytotoxicity on human erythrocytes and blood cells | nd | [59,142,157] |
| Lb MBL | Lubomirskia baicalensis | Haplosclerida (D) | nd | biomineralization/spiculogenesis | [52] |
| PsL | Pellina semitubulosa | Halichondrida (D) | sheep, rabbit and human erythrocyte agglutination strong mitogenic effect on spleen lymphocytes of mice interleukin-1 release from mouse peritoneal macrophages nterleukin-2 production by murine lymphocyte cultures | nd | [58] |
| Sponge tachylectin-like lectins | |||||
| Ef lectin | Ephydatia fluviatilis | Haplosclerida (D) | putative antibacterial activity | host defense | [42] |
| Sd lectin | Suberites domuncula | Hadromerida (D) | antibacterial activity against E. coli and S. aureus | host defense | [41] |
| Sponge F-type lectin | |||||
| CcL | Crambe crambe | Poecilosclerida (D) | sheep and human erythrocyte agglutination mitotic activity on human lymphocytes | nd | [78] |
| Unclassified sponge lectins | |||||
| AcL I | Axinella corrugata | Halichondrida (D) | goat, dog and rabbit erythrocyte agglutination chemotaxis activity on rat neutrophils mitotic effect toward human mononuclear cells cytotoxic effect against Artemia salina | host defense | [53,63] |
| AcL II | Axinella corrugata | Halichondrida (D) | rabbit erythrocyte agglutination | nd | [63] |
| ApaL I | Aaptos papillata | Hadromerida (D) | nd | nd | [48] |
| ApaL II | Aaptos papillata | Hadromerida (D) | nd | nd | [48] |
| ApaL III | Aaptos papillata | Hadromerida (D) | nd | nd | [48] |
| ApL I | Axinella polypoides | Halichondrida (D) | mitogenic activation on human lymphocytes | spongin production | [64,87] |
| ApL II | Axinella polypoides | Halichondrida (D) | nd | spongin production | [64,87] |
| ApL III | Axinella polypoides | Halichondrida (D) | nd | nd | [45,79] |
| ApL IV | Axinella polypoides | Halichondrida (D) | nd | nd | [45,79] |
| ApL V | Axinella polypoides | Halichondrida (D) | nd | nd | [45,79] |
| CaL | Cinachyrella apion | Spirophorida (D) | human erythrocte agglutination Leishmania chagasi agglutination antiproliferative activity against HeLa, PC3 and 3T3 cells no cytotoxicity on human erythrocytes and blood cells | host defense | [60,158] |
| CalL | Cinachyrella alloclada | Spirophorida (D) | human erythrocyte aggulitination | nd | [84] |
| CauL | Craniella australiensis | Spirophorida (D) | mitogenic activity on human lymphocytes mouse, sheep, rabbit and human erythrocte agglutination mitogenic activity on BALB/c splenocytes | host defense | [61] |
| CnL | Chondrilla nucula | Chondrosida (D) | mitotic activity on mouse and human lymphocytes increase the activity of the (2′-5′) oligoadenylate modulation of HIV-release period by infected cells | host defense | [85] |
| CtL | Cinachyrella tenuiviolacea | Spiroporida (D) | human erythrocte agglutination | nd | [46] |
| DaL | Desmapsamma anchorata | nd | human erythrocte agglutination mitogenic activity on human lymphocytes | nd | [86] |
| Halilectin 1 (H-1) | Haliclona caerulea | Haplosclerida (D) | rabbit erythrocyte agglutination cytotoxic effect on Artemia nauplii | host defense | [49] |
| Halilectin 2 (H-2) | Haliclona caerulea | Haplosclerida (D) | rabbit erythrocyte agglutination cytotoxic effect on Artemia nauplii | host defense | [49] |
| Halilectin 3 (H-3) | Haliclona caerulea | Haplosclerida (D) | human and rabbit erythrocyte agglutination | nd | [83] |
| HcL | Haliclona cratera | Haplosclerida (D) | sheep and human erythrocyte agglutination cytotoxic activity against HeLa and FemX cells weak mitogenic effect on human T lymphocytes | nd | [54] |
| HL | Haliclona sp. | Haplosclerida (D) | human erythrocte agglutination | nd | [46] |
| HoL-1 | Halichondria okadai | Halichondrida (D) | human erythrocte agglutination | nd | [80] |
| HoL-2 | Halichondria okadai | Halichondrida (D) | human erythrocte agglutination | nd | [80] |
6.3. Cytotoxic and Anticancer Activities
6.4. Neuromodulatory Activity
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Boyd, W.C.; Sharpleigh, E. Specific precipitation activity of plant agglutinins (lectins). Science 1954, 119, 419. [Google Scholar] [CrossRef] [PubMed]
- Gabius, H.J. Animal lectin. J. Biochem. 1997, 243, 543–576. [Google Scholar] [CrossRef]
- Rüdiger, H.; Gabius, H.J. Plant lectins: Occurrence, biochemistry, functions and applications. Glycoconj. J. 2001, 18, 589–613. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, K.S.; Cunha, A.I.; Nascimento, K.S.; Cavada, B.S.; Azevedo, A.M.; Aires-Barros, M.R. An overview of lectins purification strategies. J. Mol. Recognit. 2012, 25, 527–541. [Google Scholar] [CrossRef] [PubMed]
- Sharon, N. Bacterial lectins, cell-cell recognition and infectious disease. FEBS 1987, 217, 145–157. [Google Scholar] [CrossRef]
- Lakhtin, V.; Lakhtin, M.; Alyoshkin, V. Lectins of living organisms. The overview. Anaerobe 2011, 17, 452–455. [Google Scholar] [CrossRef] [PubMed]
- Sumner, J.B.; Howel, S.F. Identification of hemagglutinin of jack bean with Concanavalin A. J. Bacteriol. 1936, 32, 227–237. [Google Scholar] [PubMed]
- Hamblin, J.; Kent, S.P. Possible role of phytohaemagglutinin in Phaseolus vulgaris L. Nat. New Biol. 1973, 245, 28–30. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.L.; Zhang, S.; Tian, M.; Zhang, S.Y.; Xie, T.; Chen, D.Y.; Chen, Y.J.; He, J.; Liu, J.; Ouyang, L.; et al. Plant lectins, from ancient sugar-binding proteins to emerging anti-cancer drugs in apoptosis and autophagy. Cell Prolif. 2015, 48, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Ang, A.S.; Cheung, R.C.; Dan, X.; Chan, Y.S.; Pan, W.; Ng, T.B. Purification and characterization of a glucosamine-binding antifungal lectin from Phaseolus vulgaris cv. Chinese pinto beans with antiproliferative activity towards nasopharyngeal carcinoma cells. Appl. Biochem. Biotechnol. 2014, 172, 672–686. [Google Scholar] [PubMed]
- Ashraf, M.T.; Khan, R.H. Mitogenic lectins. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2003, 9, 265–269. [Google Scholar]
- Valadez-Vega, C.; Guzmán-Partida, A.M.; Soto-Cordova, F.J.; Alvarez-Manilla, G.; Zúñiga-Pérez, C.; Morales-González, J.A.; Madrigal-Santillán, E.; Villagómez-Ibarra, J.R.; Gutiérrez-Salinas, J.; Becerril-Flores, M.A. Purification, biochemical characterization, and bioactive properties of a lectin purified from the seeds of white tepary bean (Phaseolus acutifolius variety latifolius). Molecules 2011, 16, 2561–2582. [Google Scholar] [CrossRef] [PubMed]
- Chernikov, O.V.; Molchanova, V.I.; Chikalovets, I.V.; Kondrashina, A.S.; Li, W.; Lukyanov, P.A. Lectins of marine hydrobionts. Biochem. Mosc. 2013, 78, 760–770. [Google Scholar] [CrossRef] [PubMed]
- Molchanova, V.I.; Chernikov, O.V.; Chikalovets, I.V.; Lukyanov, P.A. Purification and partial characterization of the lectin from the marine red alga Tichocarpus crinitus (Gmelin) Rupr. (Rhodophyta). Bot. Mar. 2010, 53, 69–78. [Google Scholar] [CrossRef]
- Chikalovets, I.V.; Kondrashina, A.S.; Chernikov, O.V.; Molchanova, V.I.; Lukyanov, P.A. Isolation and general characteristics of lectin from the mussel Mytilus trossulus. Chem. Nat. Comp. 2013, 48, 1058–1061. [Google Scholar] [CrossRef]
- O’Keefe, B.R.; Giomarelli, B.; Barnard, D.L.; Shenoy, S.R.; Chan, P.K.; McMahon, J.B.; Palmer, K.E.; Barnett, B.W.; Meyerholz, D.K.; Wohlford-Lenane, C.L.; et al. Broad-spectrum in vitro activity and in vivo efficacy of the antiviral protein griffithsin against emerging viruses of the family Coronaviridae. J. Virol. 2010, 84, 2511–2521. [Google Scholar] [CrossRef] [PubMed]
- Meuleman, P.; Albecka, A.; Belouzard, S.; Vercauteren, K.; Wychowski, C.; Leroux-Roels, G.; Verhoye, L.; Palmer, K.E.; Dubuisson, J. Griffithsin has antiviral activity against hepatitis C virus. Antimicrob. Agents Chemother. 2011, 55, 5159–5167. [Google Scholar] [CrossRef] [PubMed]
- Ishag, H.Z.; Li, C.; Huang, L.; Sun, M.X.; Wang, F.; Ni, B.; Malik, T.; Chen, P.Y.; Mao, X. Griffithsin inhibits Japanese encephalitis virus infection in vitro and in vivo. Arch. Virol. 2013, 158, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Morimoto, K.; Hirayama, M.; Hori, K. High mannose-specific lectin (KAA-2) from the red alga Kappaphycus alvarezii potently inhibits influenza virus infection in a strain-independent manner. Biochem. Biophys. Res. Commun. 2011, 405, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Hirayama, M.; Morimoto, K.; Yamamoto, N.; Okuyama, S.; Hori, K. Boodlea coacta is a potent entry inhibitor of HIV-1 and influenza viruses. J. Biol. Chem. 2012, 286, 19446–19458. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, J.H.; Dong-Yun, O.Y.; Molchanova, V.I.; Chikalovets, I.V.; Chernikov, O.V. Anti-human immunodeficiency virus type 1 (HIV-1) activity of lectins from ascidian Didemnum ternatanum. Glycobiology 2006, 16, 1159–1159. [Google Scholar]
- Wang, J.H.; Kong, J.; Li, W.; Molchanova, V.I.; Chikalovets, I.V.; Belogortseva, N.; Lukyanov, P.A.; Zheng, Y.T. A beta-galactose-specific lectin isolated from the marine worm Chaetopterus variopedatus possesses anti-HIV-1 activity. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 142, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Molchanova, V.I.; Chikalovets, I.V.; Chernikov, O.V.; Belogortseva, N.; Li, W.; Wang, J.H.; Yang, D.Y.; Zheng, Y.T.; Lukyanov, P. A new lectin from the sea worm Serpula vermicularis: Isolation, characterization and anti-HIV activity. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 145, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Kovalchuk, S.N.; Chikalovets, I.V.; Chernikov, O.V.; Molchanova, V.I.; Li, W.; Rasskazov, V.A.; Lukyanov, P.A. cDNA cloning and structural characterization of a lectin from the mussel Crenomytilus grayanus with a unique amino acid sequence and antibacterial activity. Fish Shellfish Immunol. 2013, 35, 1320–1324. [Google Scholar] [CrossRef] [PubMed]
- Van Soest, R.W.; Boury-Esnault, N.; Vacelet, J.; Dohrmann, M.; Erpenbeck, D.; de Voogd, N.J.; Santodomingo, N.; Vanhoorne, B.; Kelly, M.; Hooper, J.N. Global diversity of sponges (Porifera). PLoS ONE 2012, 7, e35105. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Dewapriya, P. Bioactive compounds from marine sponges and their symbiotic microbes: A potential source of nutraceuticals. Adv. Food Nutr. Res. 2012, 65, 137–151. [Google Scholar] [PubMed]
- Roué, M.; Quévrain, E.; Domart-Coulon, I.; Bourguet-Kondracki, M.L. Assessing calcareous sponges and their associated bacteria for the discovery of new bioactive natural products. Nat. Prod. Rep. 2012, 29, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Privat de Garilhe, M.; de Rudder, J. Effet de deux nucleosides de l’arabinose sur la multiplication des virus de l’herpes et de la vaccine en culture cellulaire. C. R. Acad. Sci. 1964, 259, 2725–2728. (In French) [Google Scholar]
- Momparler, R.L. A model for chemotherapy of acute leukemia with 1-β-d-arabinofuranosylcytosine. Cancer Res. 1967, 34, 1775–1787. [Google Scholar]
- Bergmann, W.; Feeney, R.J. Contributions to the study of marine products. XXXII. The nucleosides of sponges. I. J. Org. Chem. 1951, 16, 981–987. [Google Scholar] [CrossRef]
- Hirata, Y.; Uemura, D. Halichondrins-antitumor polyether macrolides from a marine sponge. Pure Appl. Chem. 1986, 58, 701–710. [Google Scholar] [CrossRef]
- Traynor, K. Eribulin approved for advanced breast cancer. Am. J. Health Syst. Pharm. 2011, 68. [Google Scholar] [CrossRef] [PubMed]
- Prokop, O.; Uhlenbruck, G.; Kohler, W. A new source of antibody-like substances having anti-blood group specificity. A discussion on the specificity of Helix agglutinins. Vox Sang. 1968, 14, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Baldo, B.A.; Uhlenbruck, G. Tridacnin, a potent anti-galactan precipitin from the hemolymph of Tridacna maxima (Röding). Adv. Exp. Med. Biol. 1975, 64, 3–11. [Google Scholar] [PubMed]
- Müller, W.E.G.; Kurelec, B.; Zahn, R.K.; Müller, I.M.; Vaith, P.; Uhlenbruck, G. Aggregation of sponge cells. Function of a lectin in its homologous biological system. J. Biol. Chem. 1979, 254, 7479–7481. [Google Scholar] [PubMed]
- Gomes Filho, S.M.; Cardoso, J.D.; Anaya, K.; Silva do Nascimento, E.; de Lacerda, J.T.; Mioso, R.; Santi Gadelha, T.; de Almeida Gadelha, C.A. Marine sponge lectins: Actual status on properties and biological activities. Molecules 2015, 20, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Hirabayashi, J.; Kasai, K. The family of metazoan metal-independent beta-galactoside-binding lectins: Structure, function and molecular evolution. Glycobiology 1993, 3, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.E.G.; Blumbach, B.; Wagner-Hülsmann, C.; Lessel, U. Galectins in the phylogenetically oldest metazoa, the sponge (Porifera). Trends Glycosci. Glycotechnol. 1997, 45, 123–130. [Google Scholar] [CrossRef]
- Vasta, G.R.; Ahmed, H.; Bianchet, M.A.; Fernández-Robledo, J.A.; Amzel, L.M. Diversity in recognition of glycans by F-type lectins and galectins: Molecular, structural, and biophysical aspects. Ann. N. Y. Acad. Sci. 2012, 1253, E14–E26. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Kawabata, S.; Hirata, M.; Iwanaga, S. A novel type of limulus lectin-L6. Purification, primary structure, and antibacterial activity. J. Biol. Chem. 1995, 270, 14493–14499. [Google Scholar] [CrossRef] [PubMed]
- Schröder, H.C.; Ushijima, H.; Krasko, A.; Gamulin, V.; Thakur, N.L.; Diehl-Seifert, B.; Müller, I.M.; Müller, W.E.G. Emergence and disappearance of an immune molecule, an antimicrobial lectin, in basal metazoa. A tachylectin-related protein in the sponge Suberites domuncula. J. Biol. Chem. 2003, 278, 32810–32817. [Google Scholar] [CrossRef] [PubMed]
- Funayama, N.; Nakatsukasa, M.; Kuraku, S.; Takechi, K.; Dohi, M.; Iwabe, N.; Miyata, T.; Agata, K. Isolation of Ef silicatein and Ef lectin as molecular markers for sclerocytes and cells involved in innate immunity in the freshwater sponge Ephydatia fluviatilis. Zool. Sci. 2005, 22, 1113–1122. [Google Scholar] [CrossRef] [PubMed]
- Dodd, R.Y.; MacLennan, A.P.; Alyoshkin, V. Haemagglutinins from marine sponges. Vox Sang. 1968, 15, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.G.; Bretting, H.; Kabat, E.A. A galactose-inhibitable mitogen for human lymphocytes from the sponge Axinella polypoides. J. Immunol. 1976, 117, 1226–1232. [Google Scholar] [PubMed]
- Bretting, H.; Donadey, C.; Vacelet, J.; Jacobs, G. Investigations on the occurrence of lectins in marine sponges with special regard to some species of the familly axinellidae. Comp. Biochem. Physiol. B 1981, 70, 69–76. [Google Scholar]
- Mebs, D.; Weiler, I.; Heinke, H.F. Bioactive proteins from marine sponges: Screening of sponge extracts for hemagglutinating, hemolytic, ichthyotoxic and lethal properties and isolation and characterization of hemagglutinins. Toxicon 1985, 23, 955–962. [Google Scholar] [CrossRef]
- Kljajic, Z. Faculty of Chemistry. Ph.D. Thesis, University of Belgrade, Belgrade, Serbia, 1986. [Google Scholar]
- Bretting, H.; Kabat, E.A.; Liao, J.; Pereira, M.E. Purification and characterization of the agglutinins from the sponge Aaptos papillata and a study of their combining sites. Biochemistry 1976, 15, 5029–5038. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, R.F.; de Melo, A.A.; Nascimento, F.E.; Simplicio, C.A.; Nascimento, K.S.; Rocha, B.A.; Saker-Sampaio, S.; Moura Rda, M.; Mota, S.S.; Cavada, B.S.; et al. Halilectin 1 (H-1) and Halilectin 2 (H-2): Two new lectins isolated from the marine sponge Haliclona caerulea. Int. J. Biochem. Cell Biol. 2013, 45, 51–58. [Google Scholar]
- Müller, W.E.G.; Zahn, R.K.; Kurelec, B.; Lucu, C.; Müller, I.M.; Uhlenbruck, G. Lectin, a possible basis for symbiosis between bacteria and sponges. J. Bacteriol. 1981, 145, 548–558. [Google Scholar] [PubMed]
- Wiens, M.; Mangoni, A.; D’Esposito, M.; Fattorusso, E.; Korchagina, N.; Schröder, H.C.; Grebenjuk, V.A.; Krasko, A.; Batel, R.; Müller, I.M.; et al. The molecular basis for the evolution of the metazoan bodyplan: Extracellular matrix-mediated morphogenesis in marine demosponges. J. Mol. Evol. 2003, 57, S60–S75. [Google Scholar] [CrossRef] [PubMed]
- Wiens, M.; Belikov, S.I.; Kaluzhnaya, O.V.; Krasko, A.; Schröder, H.C.; Perovic-Ottstadt, S.; Müller, W.E.G. Molecular control of serial module formation along the apical-basal axis in the sponge Lubomirskia baicalensis: Silicateins, mannose-binding lectin and mago nashi. Dev. Genes Evol. 2006, 216, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Dresch, R.R.; Zanetti, G.D.; Lerner, C.B.; Mothes, B.; Trindade, V.M.; Vozári-Hampe, M.M.; Henriques, A.T. ACL-I, a lectin from the marine sponge Axinella corrugata: Isolation, characterization and chemotactic activity. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2008, 148, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Pajic, I.; Kljajic, Z.; Dogovic, N.; Sladic, D.; Juranic, Z.; Gasic, M.J. A novel lectin from the sponge Haliclona cratera: Isolation, characterization and biological activity. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2002, 132, 213–221. [Google Scholar] [CrossRef]
- Hanisch, F.G.; Baldus, S.E.; Kümmel, T.A. Forssman disaccharide is the specific ligand of a galectin from the sponge Geodia cydonium but does not mediate its binding to nuclear protein np56. Glycobiology 1996, 6, 321–336. [Google Scholar] [CrossRef] [PubMed]
- Gundacker, D.; Leys, S.P.; Schröder, H.C.; Müller, I.M.; Müller, W.E.G. Isolation and cloning of a C-type lectin from the hexactinellid sponge Aphrocallistes vastus: A putative aggregation factor. Glycobiology 2001, 11, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Wagner-Hülsmann, C.; Bachinski, N.; Diehl-Seifert, B.; Blumbach, B.; Steffen, R.; Pancer, Z.; Müller, W.E.G. A galectin links the aggregation factor to cells in the sponge (Geodia cydonium) system. Glycobiology 1996, 6, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.; Bachmann, M.; Schröder, H.C.; Rinkevich, B.; Kljajic, Z.; Uhlenbruck, G.; Müller, W.E.G. A novel galactose- and arabinose-specific lectin from the sponge Pellina semitubulosa: Isolation, characterization and immunobiological properties. Biochimie 1992, 74, 527–537. [Google Scholar] [CrossRef]
- Moura, R.M.; Queiroz, A.F.; Fook, J.M.; Dias, A.S.; Monteiro, N.K.; Ribeiro, J.K.; Moura, G.E.; Macedo, L.L.; Santos, E.A.; Sales, M.P. CvL, a lectin from the marine sponge Cliona varians: Isolation, characterization and its effects on pathogenic bacteria and Leishmania promastigotes. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2006, 145, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, D.S.; Medeiros, T.L.; Ribeiro, J.K.; Monteiro, N.K.; Migliolo, L.; Vasconcelos, I.M.; Uchoa, A.F.; Oliveira, A.S.; de Sales, M.P.; Santos, E.A. A lactose specific lectin from the sponge Cinachyrella apion: Purification, characterization, N-terminal sequences alignment and agglutinating activity on Leishmania promastigotes. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2010, 155, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Xiong, C.; Li, W.; Liu, H.; Zhang, W.; Dou, J.; Bai, X.; Du, Y.; Ma, X. A normal mucin-binding lectin from the sponge Craniella australiensis. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 143, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Kawsar, S.M.; Fujii, Y.; Matsumoto, R.; Ichikawa, T.; Tateno, H.; Hirabayashi, J.; Yasumitsu, H.; Dogasaki, C.; Hosono, M.; Nitta, K.; et al. Isolation, purification, characterization and glycan-binding profile of a d-galactoside specific lectin from the marine sponge, Halichondria okadai. Physiol. B Biochem. Mol. Biol. 2008, 150, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Dresch, R.R.; Lerner, C.B.; Mothes, B.; Trindade, V.M.; Henriques, A.T.; Vozári-Hampe, M.M. Biological activities of ACL-I and physicochemical properties of ACL-II, lectins isolated from the marine sponge Axinella corrugata. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2012, 161, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Buck, F.; Schulze, C.; Breloer, M.; Strupat, K.; Bretting, H. Amino acid sequence of the d-galactose binding lectin II from the sponge Axinella polypoides (Schmidt) and identification of the carbohydrate binding site in lectin II and related lectin I. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1998, 121, 153–160. [Google Scholar] [CrossRef]
- Ueda, T.; Nakamura, Y.; Smith, C.M.; Copits, B.A.; Inoue, A.; Ojima, T.; Matsunaga, S.; Sakai, R.; Swanson, G.T. Isolation of novel prototype galectins from the marine ball sponge Cinachyrella sp. guided by their modulatory activity on mammalian glutamate-gated ion channels. Glycobiology 2013, 23, 412–425. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, K.; Haasemann, M.; Gamulin, V.; Bretting, H.; Fahrenholz, F.; Müller, W.E.G. S-type lectins occur also in invertebrates: High conservation of the carbohydrate recognition domain in the lectin genes from the marine sponge Geodia cydonium. Glycobiology 1993, 3, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Schröder, H.C.; Boreiko, A.; Korzhev, M.; Tahir, M.N.; Tremel, W.; Eckert, C.; Ushijima, H.; Müller, I.M.; Müller, W.E.G. Co-expression and functional interaction of silicatein with galectin: Matrix-guided formation of siliceous spicules in the marine demosponge Suberites domuncula. J. Biol. Chem. 2006, 281, 12001–12009. [Google Scholar] [CrossRef] [PubMed]
- Bretting, H.; Phillips, S.G.; Klumpart, H.J.; Kabat, E.A. A mitogenic lactose-binding lectin from the sponge Geodia cydonium. J. Immunol. 1981, 127, 1652–1658. [Google Scholar] [PubMed]
- Diehl-Seifert, B.; Zahn, R.K.; Uhlenbruck, G.; Maidhof, A.; Müller, W.E.G. Control of L5178y cell growth by the galactose-specific lectin from Geodia cydonium. Basic Appl. Histochem. 1985, 29, 7–20. [Google Scholar] [PubMed]
- Diehl-Seifert, B.; Uhlenbruck, G.; Geisert, M.; Zahn, R.K.; Müller, W.E.G. Physicochemical and functional characterization of the polymerization process of the Geodia cydonium lectin. Eur. J. Biochem. 1985, 147, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.E.G.; Conrad, J.; Schröder, C.; Zahn, R.K.; Kurelec, B.; Dreesbach, K.; Uhlenbruck, G. Characterization of the trimeric, self-recognizing Geodia cydonium lectin I. Eur. J. Biochem. 1983, 133, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Stalz, H.; Roth, U.; Schleuder, D.; Macht, M.; Haebel, S.; Strupat, K.; Peter-Katalinic, J.; Hanisch, F.G. The Geodia cydonium galectin exhibits prototype and chimera-type characteristics and a unique sequence polymorphism within its carbohydrate recognition domain. Glycobiology 2006, 16, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Ohyama, Y.; Hirabayashi, J.; Oda, Y.; Ohno, S.; Kawasaki, H.; Suzuki, K.; Kaisai, K. Nucleotide sequence of chick 14K beta-galactoside-binding lectin mRNA. Biochem. Biophys. Res. Commun. 1986, 134, 51–56. [Google Scholar] [CrossRef]
- Rothe, B.; Roggentin, P.; Frank, R.; Blocker, H.; Schauer, R. Cloning, sequencing and expression of a sialidase gene from Clostridium sordellii G12. J. Gen. Microbiol. 1989, 135, 3087–3096. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Schloßmacher, U.; Wiens, M.; Batel, R.; Schröder, H.C.; Müller, W.E.G. Silicateins, silicatein interactors and cellular interplay in sponge skeletogenesis: Formation of glass fiber-like spicules. FEBS J. 2012, 279, 1721–1736. [Google Scholar] [CrossRef] [PubMed]
- Freymann, D.M.; Nakamura, Y.; Focia, P.J.; Sakai, R.; Swanson, G.T. Structure of a tetrameric galectin from Cinachyrella sp. (ball sponge). Acta Crystallogr. D Biol. Crystallogr. 2012, 68, 1163–1174. [Google Scholar] [CrossRef] [PubMed]
- Miarons, P.B.; Fresno, M. Lectins from tropical sponges. Purification and characterization of lectins from genus Aplysina. J. Biol. Chem. 2000, 275, 29283–29289. [Google Scholar] [CrossRef] [PubMed]
- Dogovic, N.; Sladic, D.; Kljiajic, Z.; Poznanovic, S.; Gasic, M.J. Isolation and partial characterization of a lectin from the marine sponge Crambe crambe. J. Serb. Chem. Soc. 1996, 61, 83–88. [Google Scholar]
- Buck, F.; Luth, C.; Strupat, K.; Bretting, H. Comparative investigations on the amino-acid sequences of different isolectins from the sponge Axinella polypoides (Schmidt). Biochim. Biophys. Acta 1992, 1159, 1–8. [Google Scholar] [CrossRef]
- Hazes, B. The (QxW)3 domain: A flexible lectin scaffold. Protein Sci. 1996, 5, 1490–1501. [Google Scholar] [CrossRef] [PubMed]
- Kawagishi, H.; Yamawaki, M.; Isobe, S.; Usui, T.; Kimura, A.; Chiba, S. Two lectins from the marine sponge Halichondria okadai. An N-acetyl-sugar-specific lectin (HOL-I) and an N-acetyllactosamine-specific lectin (HOL-II). J. Biol. Chem. 1994, 269, 1375–1379. [Google Scholar] [PubMed]
- Srivastava, M.; Simakov, O.; Chapman, J.; Fahey, B.; Gauthier, M.E.; Mitros, T.; Richards, G.S.; Conaco, C.; Dacre, M.; Hellsten, U.; et al. The Amphimedon queenslandica genome and the evolution of animal complexity. Nature 2010, 466, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, R.F.; de Melo, A.A.; de Almeida, A.S.; Moura Rda, M.; Chaves, R.P.; de Sousa, R.P.; Nascimento, K.S.; Sampaio, S.S.; Lima, J.P.; Cavada, B.S.; et al. H-3, a new lectin from the marine sponge Haliclona caerulea: Purification and mass spectrometric characterization. Int. J. Biochem. Cell Biol. 2013, 45, 2864–2873. [Google Scholar] [CrossRef] [PubMed]
- Atta, A.M.; Barral-Netto, M.; Peixinho, S.; Sousa-Atta, M.L. Isolation and functional characterization of a mitogenic lectin from the marine sponge Cinachyrella alloclada. Braz. J. Med. Biol. Res. 1989, 22, 3379–3385. [Google Scholar]
- Schröder, H.C.; Kljajic, Z.; Weiler, B.E.; Gasic, M.; Uhlenbruck, G.; Kurelec, B.; Müller, W.E.G. The galactose-specific lectin from the sponge Chondrilla nucula displays anti-human immunodeficiency virus activity in vitro via stimulation of the (2′-5′) oligoadenylate metabolism. Antivir. Chem. Chemother. 1990, 1, 99–105. [Google Scholar] [CrossRef]
- Atta, A.M.; Menezes, E.P.; Peixinho, S.; Sousa-Atta, M.L. Isolation of a lectin from the marine sponge Desmapsama anchorata by affinity chromatography on raffinose-sepharose 6B. Braz. J. Med. Biol. Res. 1990, 23, 191–194. [Google Scholar] [PubMed]
- Bretting, H.; Jacobs, G.; Donadey, C.; Vacelet, J. Immunohistochemical studies on the distribution and the function of the d-galactose-specific lectins in the sponge Axinella polypoides (Schmidt). Cell Tissue Res. 1983, 229, 551–571. [Google Scholar] [CrossRef] [PubMed]
- Kaltner, H.; Stierstorfer, B. Animal lectins as cell adhesion molecules. Acta Anat. Basel 1998, 161, 162–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odintsova, N.A.; Belogortseva, N.I.; Ermak, A.V.; Molchanova, V.I.; Luk’yanov, P.A. Adhesive and growth properties of lectin from the ascidian Didemnum ternatanum on cultivated marine invertebrate cells. Biochim. Biophys. Acta 1999, 1448, 381–389. [Google Scholar] [CrossRef]
- Conrad, J.; Zahn, R.K.; Kurelec, B.; Uhlenbruck, G.; Müller, W.E.G. Aggregation of sponge cells: Immunological characterization of the species-specific Geodia aggregation factor. J. Supramol. Struct. Cell. Biochem. 1981, 17, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.E.G.; Conrad, J.; Zahn, R.K.; Gramzow, M.; Kurelec, B.; Uhlenbruck, G. Identification and isolation of the primary aggregation factor from the cell membrane of the sponge Geodia cydonium. Mol. Cell. Biochem. 1985, 67, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Schütze, J.; Krasko, A.; Diehl-Seifert, B.; Müller, W.E.G. Cloning and expression of the putative aggregation factor from the marine sponge Geodia cydonium. J. Cell. Sci. 2001, 114, 3189–3198. [Google Scholar] [PubMed]
- Blumbach, B.; Pancer, Z.; Diehl-Seifert, B.; Steffen, R.; Münkner, J.; Müller, I.M.; Müller, W.E.G. The putative sponge aggregation receptor. Isolation and characterization of a molecule composed of scavenger receptor cysteine-rich domains and short consensus repeats. J. Cell Sci. 1998, 111, 2635–2644. [Google Scholar] [PubMed]
- Müller, W.E.G.; Conrad, J.; Zahn, R.K.; Steffen, R.; Uhlenbruck, G.; Müller, I.M. Cell adhesion molecules in the hexactinellid Aphrocallistes vastus: Species-unspecific aggregation factor. Differentiation 1984, 26, 30–35. [Google Scholar] [CrossRef]
- McEver, R.P. Selectins: Lectins that initiate cell adhesion under flow. Curr. Opin. Cell Biol. 2002, 14, 581–586. [Google Scholar] [CrossRef]
- Phillips, M.L.; Nudelman, E.; Gaeta, F.C.; Perez, M.; Singhal, A.K.; Hakomori, S.; Paulson, J.C. ELAM-1 mediates cell adhesion by recognition of a carbohydrate ligand, sialyl-Lex. Science 1990, 250, 1130–1132. [Google Scholar] [CrossRef] [PubMed]
- Fujii, N.; Minetti, C.A.; Nakhasi, H.L.; Chen, S.W.; Barbehenn, E.; Nunes, P.H.; Nguyen, N.Y. Isolation, cDNA cloning, and characterization of an 18-kDa hemagglutinin and amebocyte aggregation factor from Limulus polyphemus. J. Biol. Chem. 1992, 267, 22452–22459. [Google Scholar] [PubMed]
- Yang, J.; Qiu, L.; Wei, X.; Wang, L.; Wang, L.; Zhou, Z.; Zhang, H.; Liu, L.; Song, L. An ancient C-type lectin in Chlamys farreri (CfLec-2) that mediate pathogen recognition and cellular adhesion. Dev. Comp. Immunol. 2010, 34, 1274–1282. [Google Scholar] [CrossRef] [PubMed]
- Ao, J.; Ling, E.; Yu, X.Q. Drosophila C-type lectins enhance cellular encapsulation. Mol. Immunol. 2007, 44, 2541–2548. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, L.; Kong, P.; Yang, J.; Zhang, H.; Wang, M.; Zhou, Z.; Qiu, L.; Song, L. A novel C1qDC protein acting as pattern recognition receptor in scallop Argopecten irradians. Fish Shellfish Immunol. 2012, 33, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Wiens, M.; Schröder, H.C.; Wang, X.; Link, T.; Steindorf, D.; Müller, W.E.G. Isolation of the silicatein-α interactor silintaphin-2 by a novel solid-phase pull-down assay. Biochemistry 2011, 50, 1981–1990. [Google Scholar] [CrossRef] [PubMed]
- Mann, K.; Siedler, F. Ostrich (Struthio camelus) eggshell matrix contains two different C-type lectin-like proteins. Isolation, amino acid sequence, and posttranslational modifications. Biochim. Biophys. Acta 2004, 1696, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Lakshminarayanan, R.; Kini, R.M.; Valiyaveettil, S. Investigation of the role of ansocalcin in the biomineralization in goose eggshell matrix. Proc. Natl. Acad. Sci. USA 2002, 99, 5155–5159. [Google Scholar] [CrossRef] [PubMed]
- Weiss, I.M.; Kaufmann, S.; Mann, K.; Fritz, M. Purification and characterization of perlucin and perlustrin, two new proteins from the shell of the mollusc Haliotis laevigata. Biochem. Biophys. Res. Commun. 2000, 267, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Mann, K.; Weiss, I.M.; André, S.; Gabius, H.J.; Fritz, M. The amino-acid sequence of the abalone (Haliotis laevigata) nacre protein perlucin. Detection of a functional C-type lectin domain with galactose/mannose specificity. Eur. J. Biochem. 2000, 267, 5257–5264. [Google Scholar] [CrossRef] [PubMed]
- Naganuma, T.; Hoshino, W.; Shikanai, Y.; Sato, R.; Liu, K.; Sato, S.; Muramoto, K.; Osada, M.; Yoshimi, K.; Ogawa, T. Novel matrix proteins of Pteria penguin pearl oyster shell nacre homologous to the jacalin-related β-prism fold lectins. PLoS ONE 2014, 9, e112326. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, H.; Hayashi, T.; Ogawa, T.; Muramoto, K.; Jimbo, M.; Kamiya, H. Modulating effect of acorn barnacle C-type lectins on the crystallization of calcium carbonate. Fish Sci. 2008, 74, 418–424. [Google Scholar] [CrossRef]
- Wiens, M.; Korzhev, M.; Krasko, A.; Thakur, N.L.; Perović-Ottstadt, S.; Breter, H.J.; Ushijima, H.; Diehl-Seifert, B.; Müller, I.M.; Müller, W.E.G. Innate immune defense of the sponge Suberites domuncula against bacteria involves a MyD88-dependent signaling pathway. Induction of a perforin-like molecule. J. Biol. Chem. 2005, 280, 27949–27959. [Google Scholar] [CrossRef] [PubMed]
- Wiens, M.; Korzhev, M.; Perovic-Ottstadt, S.; Luthringer, B.; Brandt, D.; Klein, S.; Müller, W.E.G. Toll-like receptors are part of the innate immune defense system of sponges (demospongiae: Porifera). Mol. Biol. Evol. 2007, 24, 792–804. [Google Scholar] [CrossRef] [PubMed]
- Thakur, N.; Perovic-Ottstadt, S.; Batel, R.; Korzhev, M.; Diehl-Seifert, B.; Müller, I.M.; Müller, W.E.G. Innate immune defense of the sponge Suberites domuncula against gram-positive bacteria: Induction of lysozyme and AdaPTin. Mar. Biol. 2005, 146, 271–282. [Google Scholar] [CrossRef]
- Schröder, H.C.; Natalio, F.; Wiens, M.; Tahir, M.N.; Shukoor, M.I.; Tremel, W.; Belikov, S.I.; Krasko, A.; Müller, W.E.G. The 2′-5′-oligoadenylate synthetase in the lowest metazoa: Isolation, cloning, expression and functional activity in the sponge Lubomirskia baicalensis. Mol. Immunol. 2008, 45, 945–953. [Google Scholar] [CrossRef] [PubMed]
- Perović-Ottstadt, S.; Adell, T.; Proksch, P.; Wiens, M.; Korzhev, M.; Gamulin, V.; Müller, I.M.; Müller, W.E.G. A (1→3)-beta-d-glucan recognition protein from the sponge Suberites domuncula. Mediated activation of fibrinogen-like protein and epidermal growth factor gene expression. Mol. Immunol. 2008, 45, 1924–1937. [Google Scholar] [CrossRef] [PubMed]
- Peumans, W.J.; van Damme, E.J. Lectins as plant defense proteins. Plant Physiol. 1995, 109, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Charungchitrak, S.; Petsom, A.; Sangvanich, P.; Karnchanatat, A. Antifungal and antibacterial activities of lectin from the seeds of Archidendron jiringa Nielsen. Food Chem. 2011, 126, 1025–1032. [Google Scholar] [CrossRef]
- Lis, H.; Sharon, N. Lectins as molecules and as tools. Nature 1986, 55, 35–67. [Google Scholar] [CrossRef] [PubMed]
- Vandenborre, G.; Smagghe, G.; van Damme, E.J. Plant lectins as defense proteins against phytophagous insects. Phytochemistry 2011, 72, 1538–1550. [Google Scholar] [CrossRef] [PubMed]
- Gaynor, C.D.; McCormack, F.X.; Voelker, D.R.; McGowan, S.E.; Schlesinger, L.S. Pulmonary surfactant protein A mediates enhanced phagocytosis of Mycobacterium tuberculosis by a direct interaction with human macrophages. J. Immunol. 1995, 155, 5343–5351. [Google Scholar] [PubMed]
- McNeely, T.B.; Coonrod, J.D. Comparison of the opsonic activity of human surfactant protein A for Staphylococcus aureus and Streptococcus pneumoniae with rabbit and human macrophages. J. Infect. Dis. 1993, 167, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, M.; Endo, Y.; Taira, S.; Sato, Y.; Fujita, T.; Ichikawa, N.; Nakata, M.; Mizuochi, T. A novel human serum lectin with collagen- and fibrinogen-like domains that functions as an opsonin. J. Biol. Chem. 1996, 271, 2448–2454. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, L.; Zhang, D.; Jiang, Q.; Sun, R.; Wang, H.; Zhang, H.; Song, L. A novel multi-domain C1qDC protein from Zhikong scallop Chlamys farreri provides new insights into the function of invertebrate C1qDC proteins. Dev. Comp. Immunol. 2015, 52, 202–214. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Li, H.; Zhang, D.; Zhang, H.; Wang, L.; Sun, J.; Song, L. A C1q domain containing protein from Crassostrea gigas serves as pattern recognition receptor and opsonin with high binding affinity to LPS. Fish Shellfish Immunol. 2015, 45, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.G.; Kuroda, T.; Muroga, K. Purification and antibacterial characterization of a novel isoform of the Manila clam lectin (MCL-4) from the plasma of the Manila clam, Ruditapes philippinarum. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 150, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.D.; Fu, L.D.; Jia, Y.P.; Du, X.J.; Wang, Q.; Wang, Y.H.; Zhao, X.F.; Yu, X.Q.; Wang, J.X. A hepatopancreas-specific C-type lectin from the Chinese shrimp Fenneropenaeus chinensis exhibits antimicrobial activity. Mol. Immunol. 2008, 45, 348–361. [Google Scholar] [CrossRef] [PubMed]
- Hentschel, U.; Usher, K.M.; Taylor, M.W. Marine sponges as microbial fermenters. FEMS Microbiol. Ecol. 2006, 55, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Wehrl, M.; Steinert, M.; Hentschel, U. Bacterial uptake by the marine sponge Aplysina aerophoba. Microb. Ecol. 2007, 53, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Reveillaud, J.; Maignien, L.; Eren, A.M.; Huber, J.A.; Apprill, A.; Sogin, M.L.; Vanreusel, A. Host-specificity among abundant and rare taxa in the sponge microbiome. ISME J. 2014, 8, 1198–1209. [Google Scholar] [CrossRef] [PubMed]
- Fieseler, L.; Horn, M.; Wagner, M.; Hentschell, U. Discovery of the novel candidate phylum “Poribacteria” in marine sponges. Appl. Environ. Microbiol. 2004, 70, 3724–3732. [Google Scholar] [CrossRef] [PubMed]
- Siegl, A.; Kamke, J.; Hochmuth, T.; Piel, J.; Richter, M.; Liang, C.; Dandekar, T.; Hentschel, U. Single-cell genomics reveals the lifestyle of Poribacteria, a candidate phylum symbiotically associated with marine sponges. ISME J. 2011, 5, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Rusch, D.; Demaere, M.Z.; Yung, P.Y.; Lewis, M.; Halpern, A.; Heidelberg, K.B.; Egan, S.; Steinberg, P.D.; Kjelleberg, S. Functional genomic signatures of sponge bacteria reveal unique and shared features of symbiosis. ISME J. 2010, 4, 1557–1567. [Google Scholar] [CrossRef] [PubMed]
- Gardères, J.; Henry, J.; Bernay, B.; Ritter, A.; Zatylny-Gaudin, C.; Wiens, M.; Müller, W.E.G.; Le Pennec, G. Cellular effects of bacterial N-3-oxo-dodecanoyl-l-homoserine lactone on the sponge Suberites domuncula (Olivi, 1792): Insights into an intimate inter-kingdom dialogue. PLoS ONE 2014, 9, e97662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bry, L.; Falk, P.G.; Midtvedt, T.; Gordon, J.I. A model of host-microbial interactions in an open mammalian ecosystem. Science 1996, 273, 1380–1383. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Falk, P.G.; Gordon, J.I. Analyzing the molecular foundations of commensalism in the mouse intestine. Curr. Opin. Microbiol. 2000, 3, 79–85. [Google Scholar] [CrossRef]
- Martens, E.C.; Goodrich-Blair, H. The Steinernema carpocapsae intestinal vesicle contains a subcellular structure with which Xenorhabdus nematophila associates during colonization initiation. Cell Microbiol. 2005, 7, 1723–1735. [Google Scholar] [CrossRef] [PubMed]
- Nyholm, S.V.; Stabb, E.V.; Ruby, E.G.; McFall-Ngai, M.J. Establishment of an animal-bacterial association: Recruiting symbiotic vibrios from the environment. Proc. Natl. Acad. Sci. USA 2000, 97, 10231–10235. [Google Scholar] [CrossRef] [PubMed]
- Vydryakova, G.A.; Bondar’, V.S. Location of lectin exhibiting specificity for N-acetyl-d-galactosamine in cells of the symbiotic marine bacteria Photobacterium phosphoreum. Dokl. Biochem. Biophys. 2008, 420, 155–157. [Google Scholar] [CrossRef] [PubMed]
- Jimbo, M.; Yanohara, T.; Koike, K.; Sakai, R.; Muramoto, K.; Kamiya, H. The d-galactose-binding lectin of the octocoral Sinularia lochmodes: Characterization and possible relationship to the symbiotic dinoflagellates. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2000, 125, 227–236. [Google Scholar] [CrossRef]
- Bulgheresi, S.; Schabussova, I.; Chen, T.; Mullin, N.P.; Maizels, R.M.; Ott, J.A. A new C-type lectin similar to the human immunoreceptor DC-SIGN mediates symbiont acquisition by a marine nematode. Appl. Environ. Microbiol. 2006, 72, 2950–2956. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Snyder, S.; Feng, P.; Azadi, P.; Zhang, S.; Bulgheresi, S.; Sanderson, K.E.; He, J.; Klena, J.; Chen, T. Role of N-acetylglucosamine within core lipopolysaccharide of several species of gram-negative bacteria in targeting the DC-SIGN (CD209). J. Immunol. 2006, 177, 4002–4011. [Google Scholar] [CrossRef] [PubMed]
- De Hoff, P.L.; Brill, L.M.; Hirsch, A.M. Plant lectins: The ties that bind in root symbiosis and plant defense. Mol. Genet. Genom. 2009, 282, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Van Rhijn, P.; Fujishige, N.A.; Lim, P.O.; Hirsch, A.M. Sugar-binding activity of pea lectin enhances heterologous infection of transgenic alfalfa plants by Rhizobium leguminosarum biovar viciae. Plant Physiol. 2001, 126, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Laus, M.C.; Logman, T.J.; Lamers, G.E.; van Brussel, A.A.; Carlson, R.W.; Kijne, J.W. A novel polar surface polysaccharide from Rhizobium leguminosarum binds host plant lectin. Mol. Microbiol. 2006, 59, 1704–1713. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, A.F.; Moura, R.M.; Ribeiro, J.K.; Lyra, I.L.; Cunha, D.C.; Santos, E.A.; de-Sales, M.P. Pro-inflammatory effect in mice of CvL, a lectin from the marine sponge Cliona varians. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2008, 147, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, A.; Kuwabara, I.; Frigeri, L.G.; Liu, F.T. A human lectin, galectin-3 (epsilon bp/Mac-2), stimulates superoxide production by neutrophils. J. Immunol. 1995, 154, 3479–3487. [Google Scholar] [PubMed]
- Hsu, D.K.; Yang, R.Y.; Pan, Z.; Yu, L.; Salomon, D.R.; Fung-Leung, W.P.; Liu, F.T. Targeted disruption of the galectin-3 gene results in attenuated peritoneal inflammatory responses. Am. J. Pathol. 2000, 156, 1073–1083. [Google Scholar] [CrossRef]
- Sano, H.; Hsu, D.K.; Yu, L.; Apgar, J.R.; Kuwabara, I.; Yamanaka, T.; Hirashima, M.; Liu, F.T. Human galectin-3 is a novel chemoattractant for monocytes and macrophages. J. Immunol. 2000, 165, 2156–2164. [Google Scholar] [CrossRef] [PubMed]
- Okino, N.; Kawabata, S.; Saito, T.; Hirata, M.; Takagi, T.; Iwanaga, S. Purification, characterization, and cDNA cloning of a 27-kDa lectin (L10) from horseshoe crab hemocytes. J. Biol. Chem. 1995, 270, 31008–31015. [Google Scholar] [PubMed]
- Kawabata, S.; Iwanaga, S. Role of lectins in the innate immunity of horseshoe crab. Dev. Comp. Immunol. 1999, 23, 391–400. [Google Scholar] [CrossRef]
- Saito, T.; Hatada, M.; Iwanaga, S.; Kawabata, S. A newly identified horseshoe crab lectin with binding specificity to O-antigen of bacterial lipopolysaccharides. J. Biol. Chem. 1997, 272, 30703–30708. [Google Scholar] [CrossRef] [PubMed]
- Inamori, K.; Saito, T.; Iwaki, D.; Nagira, T.; Iwanaga, S.; Arisaka, F.; Kawabata, S. A newly identified horseshoe crab lectin with specificity for blood group A antigen recognizes specific O-antigens of bacterial lipopolysaccharides. J. Biol. Chem. 1999, 274, 3272–3278. [Google Scholar] [CrossRef] [PubMed]
- Dey, B.; Lerner, D.L.; Lusso, P.; Boyd, M.R.; Elder, J.H.; Berger, E.A. Multiple antiviral activities of cyanovirin-N: Blocking of human immunodeficiency virus type 1 gp120 interaction with CD4 and coreceptor and inhibition of diverse enveloped viruses. J. Virol. 2000, 74, 4562–4569. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, R.L.; Schlein, Y. Lectins and toxins in the plant diet of Phlebotomus papatasi (Diptera: Psychodidae) can kill Leishmania major promastigotes in the sandfly and in culture. Ann. Trop. Med. Parasitol. 1999, 93, 351–356. [Google Scholar] [PubMed]
- Gazzinelli, R.T.; Pereira, M.E.; Romanha, A.; Gazzinelli, G.; Brener, Z. Direct lysis of Trypanosoma cruzi: A novel effector mechanism of protection mediated by human anti-gal antibodies. Parasite Immunol. 1991, 13, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, R.L. Lectin-Leishmania interaction. In Lectin-Microorganisms Interaction; Doyle, R.J., Slifkin, M., Eds.; Marcel Dekker Inc.: New York, NY, USA, 1994; pp. 191–223. [Google Scholar]
- Schottelius, J.; Alsien, M.S.O. Lectin-Microorganisms Interaction; Doyle, R.J., Slifkin, M., Eds.; Marcel Dekker Inc.: New York, NY, USA, 1994; pp. 225–248. [Google Scholar]
- Castanheira, L.E.; Nunes, D.C.; Cardoso, T.M.; Santos Pde, S.; Goulart, L.R.; Rodrigues, R.S.; Richardson, M.; Borges, M.H.; Yoneyama, K.A.; Rodrigues, V.M. Biochemical and functional characterization of a C-type lectin (BpLec) from Bothrops pauloensis snake venom. Int. J. Biol. Macromol. 2013, 54, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Afonso-Cardoso, S.R.; Rodrigues, F.H.; Gomes, M.A.; Silva, A.G.; Rocha, A.; Guimaraes, A.H.; Candeloro, I.; Favoreto, S., Jr.; Ferreira, M.S.; de Souza, M.A. Protective effect of lectin from Synadenium carinatum on Leishmania amazonensis infection in BALB/c mice. Korean J. Parasitol. 2007, 45, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, A.F.; Silva, R.A.; Moura, R.M.; Dreyfuss, J.L.; Paredes-Gamero, E.J.; Souza, A.C.; Tersariol, I.L.; Santos, E.A.; Nader, H.B.; Justo, G.Z.; et al. Growth inhibitory activity of a novel lectin from Cliona varians against K562 human erythroleukemia cells. Cancer Chemother. Pharmacol. 2009, 63, 1023–1033. [Google Scholar] [CrossRef] [PubMed]
- Rabelo, L.; Monteiro, N.; Serquiz, R.; Santos, P.; Oliveira, R.; Oliveira, A.; Rocha, H.; Morais, A.H.; Uchoa, A.; Santos, E. A lactose-binding lectin from the marine sponge Cinachyrella apion (Cal) induces cell death in human cervical adenocarcinoma cells. Mar. Drugs 2012, 10, 727–743. [Google Scholar] [CrossRef] [PubMed]
- Bantel, H.; Engels, I.H.; Voelter, W.; Schulze-Osthoff, K.; Wesselborg, S. Mistletoe lectin activates caspase-8/FLICE independently of death receptor signaling and enhances anticancer drug-induced apoptosis. Cancer Res. 1999, 59, 2083–2090. [Google Scholar] [PubMed]
- Opric, M.M.; Poznanovic, S.; Kljajic, Z.; Sladic, D.; Pupic, G.; Perunovic, B.; Gasic, M.J. Labelling of breast carcinoma, thyroid carcinoma and melanoma with manno- and galacto-specific lectins from marine invertebrates. Eur. J. Histochem. 1996, 40, 211–218. [Google Scholar] [PubMed]
- Gorelik, E.; Galili, U.; Raz, A. On the role of cell surface carbohydrates and their binding proteins (lectins) in tumor metastasis. Cancer Metastasis Rev. 2001, 20, 245–277. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.E.G. Review: How was metazoan threshold crossed? The hypothetical Urmetazoa. A. Mol. Integr. Physiol. 2001, 129, 433–460. [Google Scholar] [CrossRef]
- Barauna, S.C.; Kaster, M.P.; Heckert, B.T.; do Nascimento, K.S.; Rossi, F.M.; Teixeira, E.H.; Cavada, B.S.; Rodrigues, A.L.; Leal, R.B. Antidepressant-like effect of lectin from Canavalia brasiliensis (ConBr) administered centrally in mice. Pharmacol. Biochem. Behav. 2006, 85, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Soares, G.D.; Lima, C.B.; Cavalcanti, L.C.; Villacampa, N.; Castellano, B.; Guedes, R.C. Brain effects of the lectin from Canavalia ensiformis in adult rats previously suckled in favorable and unfavorable conditions: A spreading depression and microglia immunolabeling study. Nutr. Neurosci. 2015. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gardères, J.; Bourguet-Kondracki, M.-L.; Hamer, B.; Batel, R.; Schröder, H.C.; Müller, W.E.G. Porifera Lectins: Diversity, Physiological Roles and Biotechnological Potential. Mar. Drugs 2015, 13, 5059-5101. https://doi.org/10.3390/md13085059
Gardères J, Bourguet-Kondracki M-L, Hamer B, Batel R, Schröder HC, Müller WEG. Porifera Lectins: Diversity, Physiological Roles and Biotechnological Potential. Marine Drugs. 2015; 13(8):5059-5101. https://doi.org/10.3390/md13085059
Chicago/Turabian StyleGardères, Johan, Marie-Lise Bourguet-Kondracki, Bojan Hamer, Renato Batel, Heinz C. Schröder, and Werner E. G. Müller. 2015. "Porifera Lectins: Diversity, Physiological Roles and Biotechnological Potential" Marine Drugs 13, no. 8: 5059-5101. https://doi.org/10.3390/md13085059
APA StyleGardères, J., Bourguet-Kondracki, M.-L., Hamer, B., Batel, R., Schröder, H. C., & Müller, W. E. G. (2015). Porifera Lectins: Diversity, Physiological Roles and Biotechnological Potential. Marine Drugs, 13(8), 5059-5101. https://doi.org/10.3390/md13085059